Testing Different Membrane Filters for 16S rRNA Gene-Based Metabarcoding in Karstic Springs

 ,
,  ,
,  , ,
, ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Sample Preparation
2.2. Physicochemical Measurements and Laboratory Analysis
2.3. 16S rRNA Protocol
2.4. Statistical Analysis
3. Results
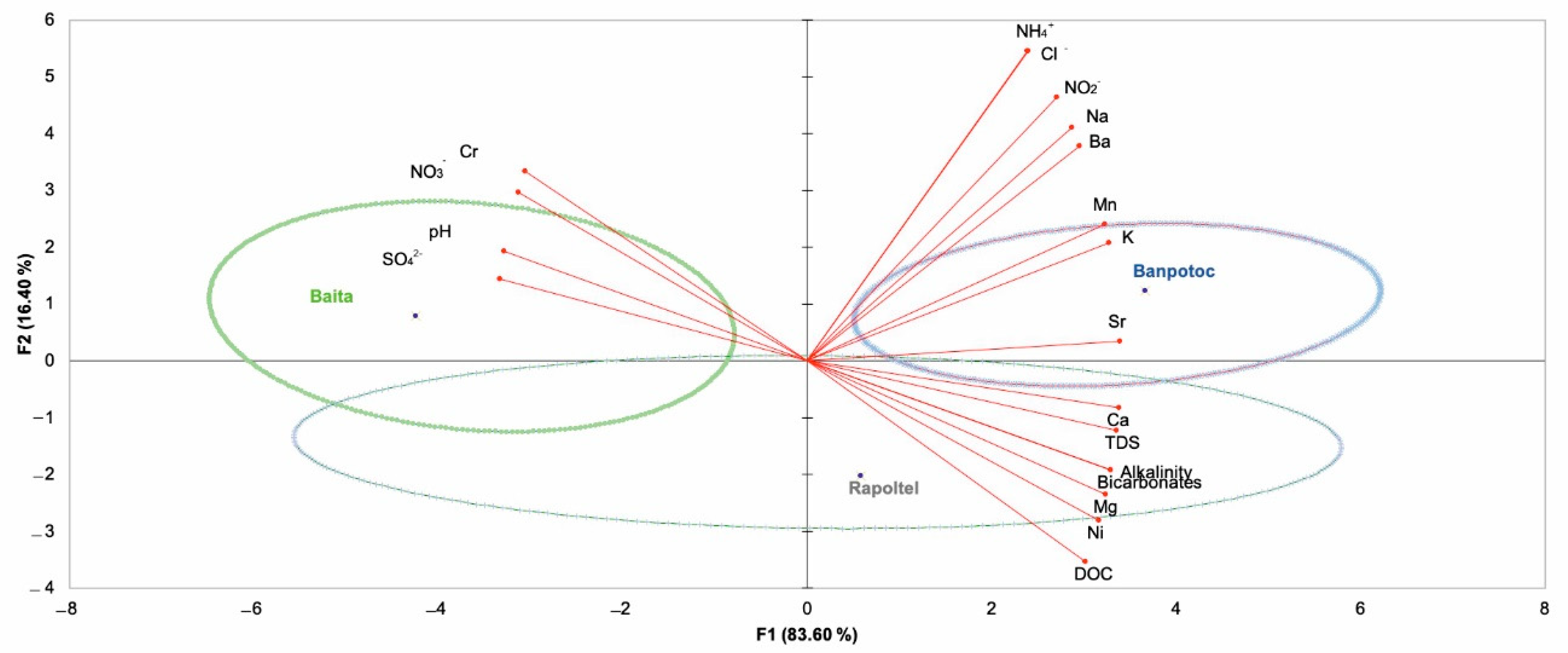
3.1. Chemical Analysis
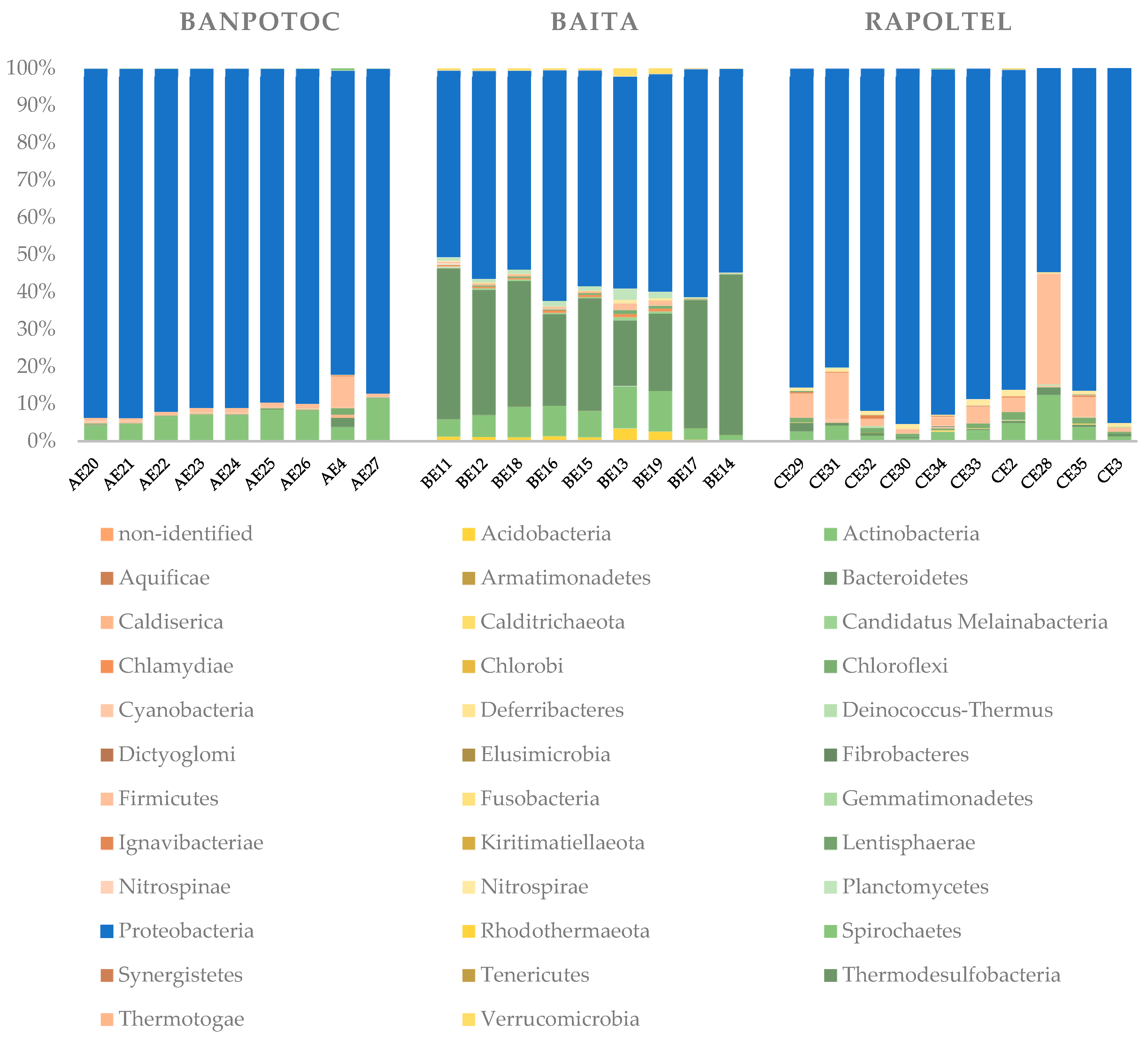
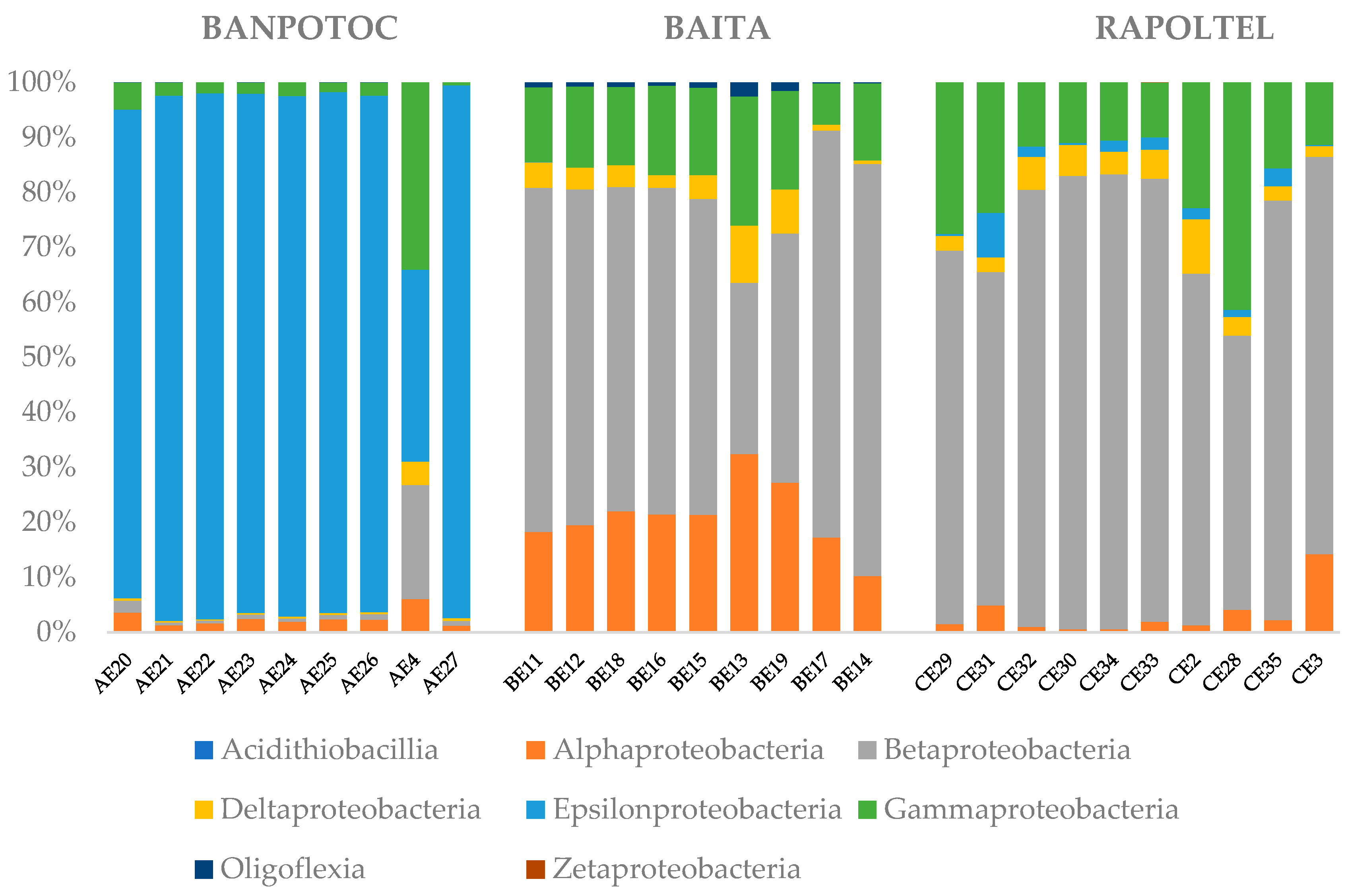
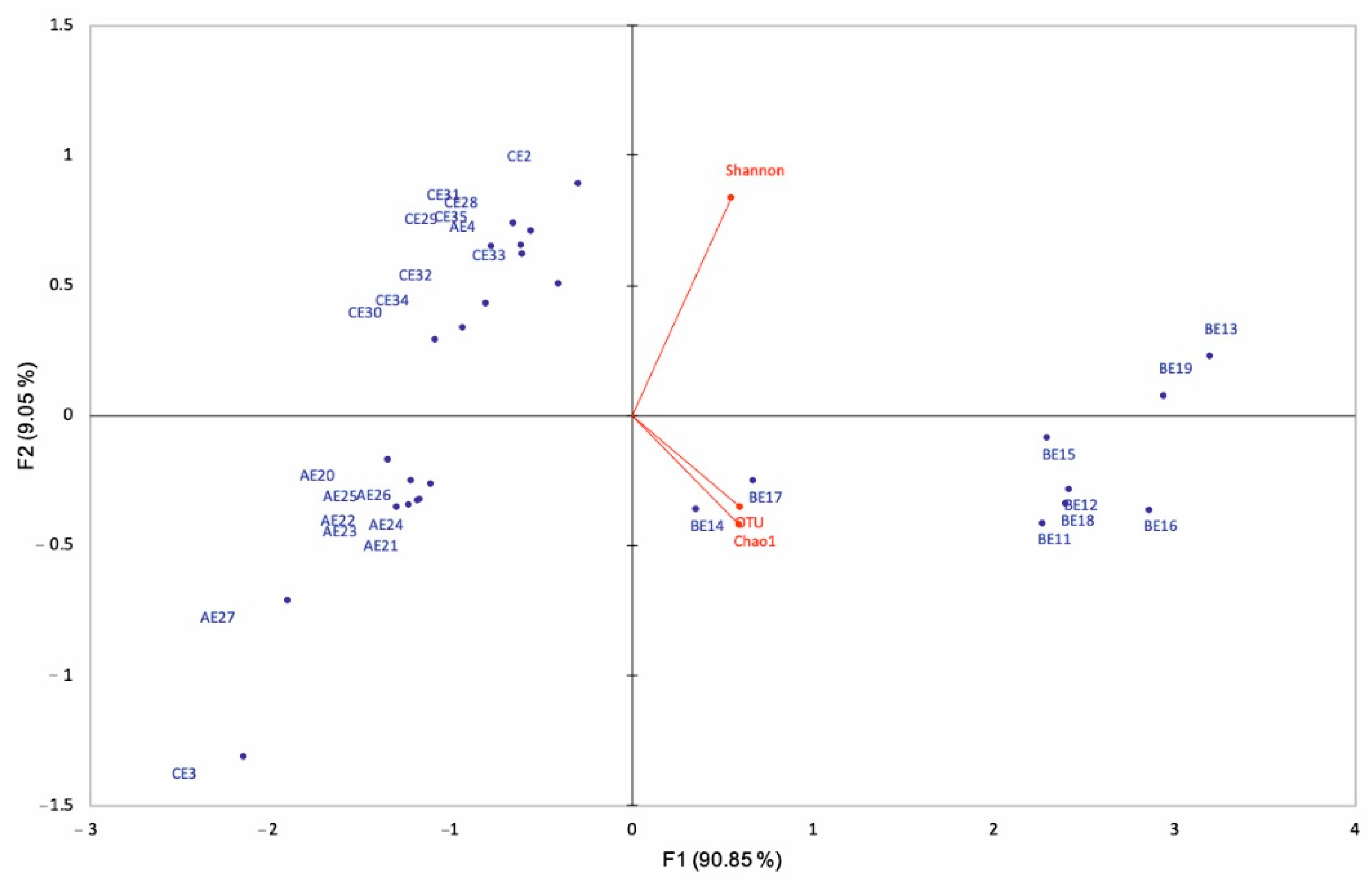
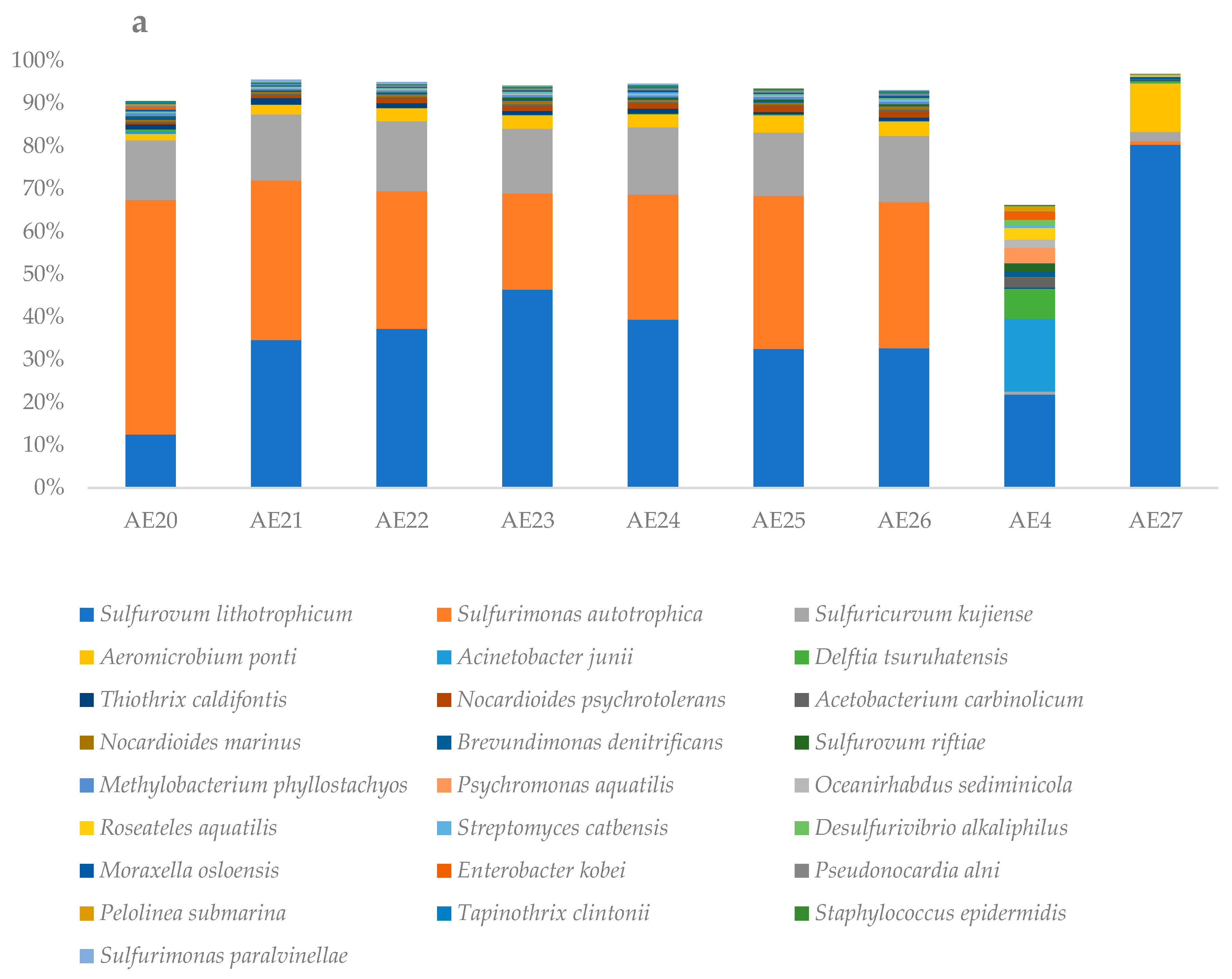
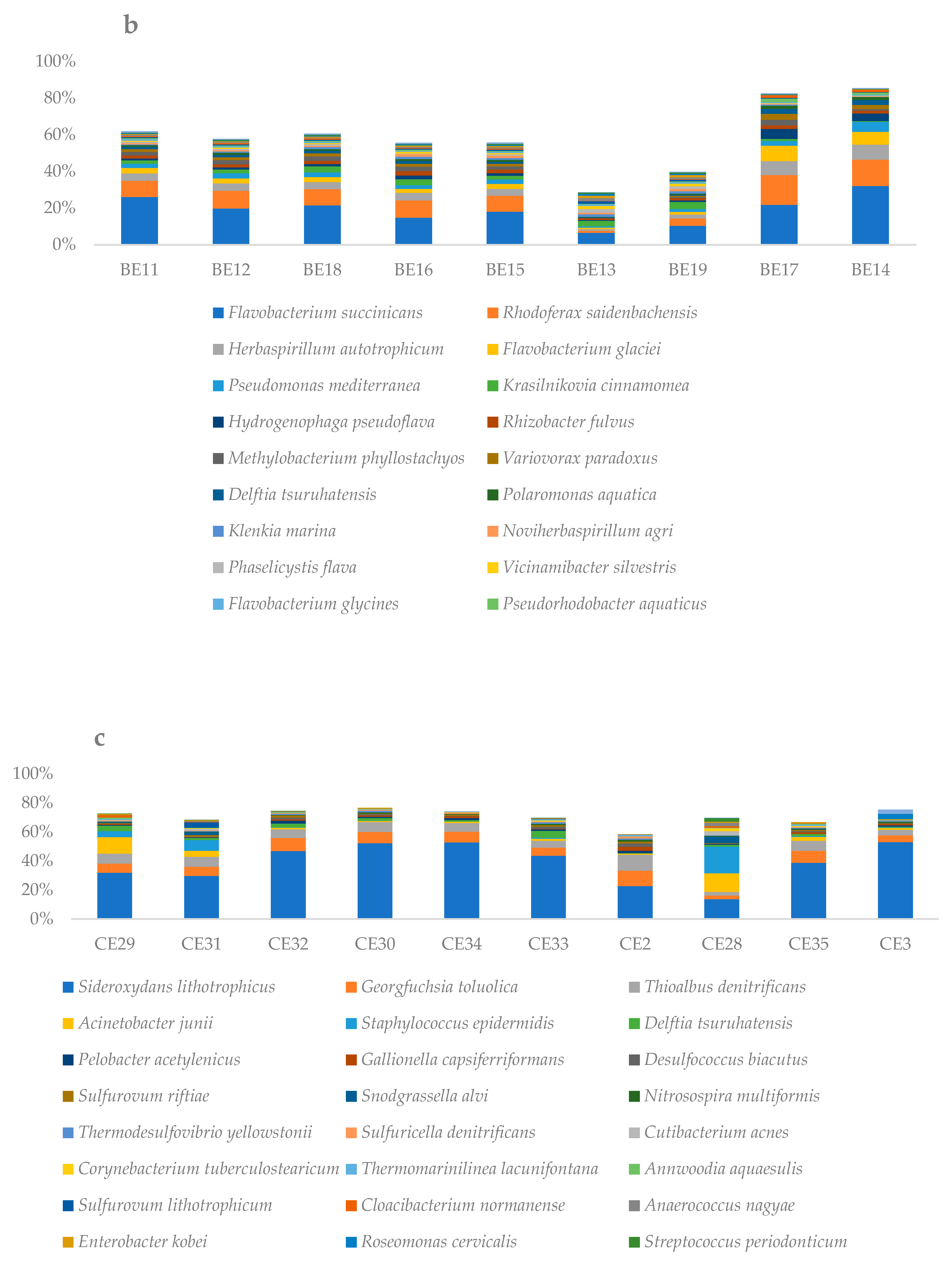
3.2. Microbial Diversity Inferred by 16s RNA Gene-Based Metabarcoding
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rodell, M.; Famiglietti, J.S.; Wiese, D.N.; Reager, J.T.; Beaudoing, H.K.; Landerer, F.W.; Lo, M.-H. Emerging trends in global freshwater availability. Nature 2018, 557, 651–659. [Google Scholar] [CrossRef]
- Zektser, I.S.; Everett, L.G. (Eds.) Groundwater Resources of the World and Their Use; UNESCO: Paris, France, 2004; Available online: http://unesdoc.unesco.org/images/0013/001344/134433e.pdf (accessed on 3 December 2020).
- Siebert, S.; Burke, J.; Faures, J.M.; Frenken, K.; Hoogeveen, J.; Döll, P.; Portmann, F.T. Groundwater use for irrigation—A global inventory. Hydrol. Earth Syst. Sci. 2010, 14, 1863–1880. [Google Scholar] [CrossRef] [Green Version]
- Taylor, R.G.; Scanlon, B.; Döll, P.; Rodell, M.; Van Beek, R.; Wada, Y.; Longuevergne, L.; LeBlanc, M.; Famiglietti, J.; Edmunds, M.; et al. Ground water and climate change. Nat. Clim. Chang. 2013, 3, 322–329. [Google Scholar] [CrossRef] [Green Version]
- Eurostat. Available online: https://ec.europa.eu/eurostat/web/products-datasets/-/env_wat_pop%20last (accessed on 7 June 2020).
- McDonough, L.K.; Santos, I.R.; Andersen, M.S.; O’Carroll, D.M.; Rutlidge, H.; Meredith, K.; Oudone, P.; Bridgeman, J.; Gooddy, D.C.; Sorensen, J.P.R.; et al. Changes in global groundwater organic carbon driven by climate change and urbanization. Nat. Commun. 2020, 11, 1279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoaghia, M.A.; Cadar, O.; Hognogi, G.G.; Levei, E.; Moisa, C.; Roman, C. Quality and human health risk assessment of metals and nitrogen compounds in drinking water from an urban area near a former non-ferrous ore smelter. Anal. Lett. 2019, 52, 1268–1281. [Google Scholar] [CrossRef]
- Pawlowski, J.; Kelly-Quinn, M.; Altermatt, F.; Apothéloz-Perret-Gentil, L.; Beja, P.; Boggero, A.; Borja, T.; Bouchez, A.; Cordier, T.; Domaizon, I.; et al. The future of biotic indices in the ecogenomic era: Integrating (e) DNA metabarcoding in biological assessment of aquatic ecosystems. Sci. Total Environ. 2018, 637, 1295–1310. [Google Scholar] [CrossRef] [PubMed]
- Deiner, K.; Fronhofer, E.A.; Mächler, E.; Walser, J.C.; Altermatt, F. Environmental DNA reveals that rivers are conveyer belts of biodiversity information. Nat. Commun. 2016, 7, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Kuntke, F.; de Jonge, N.; Hesselsøe, M.; Nielsen, J.L. Stream water quality assessment by metabarcoding of invertebrates. Ecol. Indic. 2020, 111, 105982. [Google Scholar] [CrossRef]
- Tolotti, M.; Cerasino, L.; Donati, C.; Pindo, M.; Rogora, M.; Seppi, R.; Albanese, D. Alpine headwaters emerging from glaciers and rock glaciers host different bacterial communities: Ecological implications for the future. Sci. Total Environ. 2020, 717, 137101. [Google Scholar] [CrossRef]
- Paruch, L.; Paruch, A.M.; Eiken, H.G.; Sørheim, R. Faecal pollution affects abundance and diversity of aquatic microbial community in anthropo-zoogenically influenced lotic ecosystems. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef]
- Szekeres, E.; Chiriac, C.M.; Baricz, A.; Szőke-Nagy, T.; Lung, I.; Soran, M.L.; Rudi, K.; Dragos, N.; Coman, C. Investigating antibiotics, antibiotic resistance genes, and microbial contaminants in groundwater in relation to the proximity of urban areas. Environ. Pollut. 2018, 236, 734–744. [Google Scholar] [CrossRef] [PubMed]
- Grönwall, J.; Danert, K. Regarding Groundwater and Drinking Water Access through A Human Rights Lens: Self-Supply as A Norm. Water 2020, 12, 419. [Google Scholar] [CrossRef] [Green Version]
- Sanganyado, E.; Gwenzi, W. Antibiotic resistance in drinking water systems: Occurrence, removal, and human health risks. Sci. Total Environ. 2019, 669, 785–797. [Google Scholar] [CrossRef] [PubMed]
- Moldovan, O.T.; Øvrevik Skoglund, R.; Banciu, H.L.; Dinu Cucoș, A.; Levei, E.A.; Perșoiu, A.; Lauritzen, S.-E. Monitoring and risk assessment for groundwater sources in rural communities of Romania (GROUNDWATERISK). Res. Ideas Outcomes 2019, 5, e48898. [Google Scholar] [CrossRef] [Green Version]
- Magoc, T.; Salzberg, S. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Fu, L.; Niu, B.; Wu, S.; Wooley, J. Ultrafast Clustering Algorithms for Metagenomic Sequence Analysis. Brief. Bioinform. 2012, 13, 656–668. [Google Scholar] [CrossRef]
- Caporaso, J.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Law 311. Law 311 for Modifying and Completing Law 458/2002, Regarding the Quality of Drinking Water, Official Monitor, No. 582 from 30 June 2004, Romania. 2004. Available online: http://legislatie.just.ro/Public/DetaliiDocumentAfis/53106 (accessed on 2 December 2020).
- Tarits, C.; Aquilina, L.; Ayraud, V.; Pauwels, H.; Davy, P.; Touchard, F.; Bour, O. Oxido-reduction sequence related to flux variations of groundwater from a fractured basement aquifer (Ploemeur area, France). Appl. Geochem. 2006, 21, 29–47. [Google Scholar] [CrossRef]
- Roques, C.; Aquilina, L.; Bour, O.; Maréchal, J.-C.; Dewandel, B.; Pauwels, H.; Labasque, T.; Vergnaud-Ayraud, V.; Hochreutener, R. Groundwater sources and geochemical processes in a crystalline fault aquifer. J. Hydrol. 2014, 519, 3110–3128. [Google Scholar] [CrossRef]
- Ben Maamar, S.; Aquilina, L.; Quaiser, A.; Pauwels, H.; Michon-Coudouel, S.; Vergnaud-Ayraud, V.; Labasque, T.; Roques, C.; Abbott, B.W.; Dufresne, A. Groundwater isolation governs chemistry and microbial community structure along hydrologic flowpaths. Front. Microbiol. 2015, 6, 1457. [Google Scholar] [CrossRef]
- Akob, D.M.; Mills, H.J.; Kostka, J.E. Metabolically active microbial communities in uranium-contaminated subsurface sediments. FEMS Microbiol. Ecol. 2007, 59, 95–107. [Google Scholar] [CrossRef] [Green Version]
- Boyd, E.S.; Cummings, D.E.; Geesey, G.G. Mineralogy influences structure and diversity of bacterial communities associated with geological substrata in a pristine aquifer. Microb. Ecol. 2007, 54, 170–182. [Google Scholar] [CrossRef] [PubMed]
- Flynn, T.M.; Sanford, R.A.; Ryu, H.; Bethke, C.M.; Levine, A.D.; Ashbolt, N.J.; Santo Domingo, J.W. Functional microbial diversity explains groundwater chemistry in a pristine aquifer. BMC Microbiol. 2013, 13, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flynn, T.M.; Sanford, R.A.; Santo Domingo, J.W.; Ashbolt, N.J.; Levine, A.D.; Bethke, C.M. The active bacterial community in a pristine confined aquifer. Water Resour. Res. 2012, 48, W09510. [Google Scholar] [CrossRef]
- Nyyssönen, M.; Bomberg, M.; Kapanen, A.; Nousiainen, A.; Pitkänen, P.; Itävaara, M. Methanogenic and sulphate-reducing microbial communities in deep groundwater of crystalline rock fractures in olkiluoto, finland. Geomicrobiol. J. 2012, 29, 863–878. [Google Scholar] [CrossRef]
- Hemme, C.L.; VanNostrand, J.D.; Wu, L.; He, Z.; Zhou, J.; Green, S.J.; Rishishwar, L.; Jordan, I.K.; Kostka, J.E.; Prakash, O.; et al. Lateral gene transfer in a heavy metal-contaminated-groundwater microbial community. MBio 2016, 7, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Z.; Zhang, P.; Wu, L.; Rocha, A.M.; Tu, Q.; Shi, Z.; Wu, B.; Qin, Y.; Wang, J.; Yan, Q.; et al. Microbial functional gene diversity predicts groundwater contamination and ecosystem functioning. MBio 2018, 9, e02435-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonthiphand, P.; Ruangroengkulrith, S.; Mhuantong, W.; Charoensawan, V.; Chotpantarat, S.; Boonkaewwan, S. Metagenomic insights into microbial diversity in a groundwater basin impacted by a variety of anthropogenic activities. Environ. Sci. Pollut. Res. 2019, 26, 26765–26781. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Membrane Code | Fabric | Producer | Porosity (µm) | Sterile * | Price ** |
|---|---|---|---|---|---|
| PES | polyethersulfone | Millipore | 0.22 | sterilized | High |
| PVDF | polyvinylidene fluoride | Millipore | 0.22 | sterilized | High |
| NC | cellulose nitrate | Macherey-Nagel | 0.20 | sterilized | Low |
| NYLON | nylon | Fioroni | 0.22 | sterilized | Low |
| MCE_a | mixed cellulose ester | Fioroni | 0.22 | sterile | Low |
| MCE_b | mixed cellulose ester | Whatman | 0.20 | sterile | Medium |
| S-Pak | mixed cellulose ester | Merck | 0.20 | sterile | Low |
| PSR009 | cellulose acetate and nitrate | Nahita | 0.22 | sterilized | Low |
| PSR010 | cellulose acetate and nitrate | Nahita | 0.45 | sterilized | Low |
| Parameter | Unit | Spring | ||
|---|---|---|---|---|
| Banpotoc | Baita | Rapoltel | ||
| TDS | mg/L | 1210 | 298 | 987 |
| DOC | mg/L | 17.7 | 1.4 | 19.3 |
| pH | - | 6.3 | 7.6 | 6.5 |
| Bicarbonates | mg/L | 1122 | 256 | 988 |
| Alkalinity | mmol/L | 18.4 | 4.2 | 16.2 |
| Na | mg/L | 28.5 | 6 | 8.1 |
| Ca | mg/L | 280 | 72.9 | 219 |
| Mg | mg/L | 37 | 4.3 | 33.9 |
| K | mg/L | 5.7 | 0.45 | 2.4 |
| Ba | µg/L | 671 | 13.6 | 109 |
| Sr | µg/L | 351 | 92.7 | 240 |
| Mn | µg/L | 74.6 | 1.5 | 25.7 |
| Ni | µg/L | 13.6 | 5.7 | 13.4 |
| Cr | µg/L | 1.48 | 2.61 | 1.41 |
| NH4+ | mg/L | 0.57 | 0.12 | 0.04 |
| Cl− | mg/L | 4.53 | 2.55 | 2.2 |
| SO42− | mg/L | 2.2 | 19.6 | 6 |
| NO3− | mg/L | <0.2 | 2.4 | <0.2 |
| NO2− | mg/L | 0.32 | <0.05 | <0.05 |
| Spring | Code | Fabric | Membrane Code | OTUs | Diversity Indices | |
|---|---|---|---|---|---|---|
| Chao1 | Shannon | |||||
| Banpotoc | AE20 | polyethersulfone | PES | 186 | 188.6250 | 2.8280 |
| AE21 | polyvinylidene fluoride | PVDF | 226 | 264.2778 | 2.5738 | |
| AE22 | cellulose nitrate | NC | 228 | 306.9643 | 2.6665 | |
| AE23 | nylon | NYLON | 264 | 294.3571 | 2.7375 | |
| AE24 | mixed cellulose ester | MCE_a | 251 | 318.0000 | 2.7688 | |
| AE25 | mixed cellulose ester | MCE_b | 241 | 257.5000 | 2.8318 | |
| AE26 | mixed cellulose ester | S-Pak | 261 | 320.2941 | 2.9322 | |
| AE4 | cellulose acetate and nitrate | PSR009 | 263 | 267.5000 | 4.9879 | |
| AE27 | cellulose acetate and nitrate | PSR010 | 115 | 126.5500 | 1.2992 | |
| Baita | BE11 | polyethersulfone | PES | 1280 | 1699.2250 | 6.4304 |
| BE12 | polyvinylidene fluoride | PVDF | 1335 | 1679.3142 | 6.8048 | |
| BE18 | cellulose nitrate | NC | 1368 | 1652.6980 | 6.6819 | |
| BE16 | nylon | NYLON | 1538 | 1803.8583 | 7.1422 | |
| BE15 | mixed cellulose ester | MCE_a | 1271 | 1569.4933 | 7.0039 | |
| BE13 | mixed cellulose ester | MCE_b | 1566 | 1734.7447 | 8.5057 | |
| BE19 | mixed cellulose ester | S-Pak | 1455 | 1752.9286 | 7.9813 | |
| BE17 | cellulose acetate and nitrate | PSR009 | 794 | 1009.5301 | 4.9305 | |
| BE14 | cellulose acetate and nitrate | PSR010 | 718 | 915.3356 | 4.3866 | |
| Rapoltel | CE29 | polyethersulfone | PES | 200 | 202.0000 | 4.8509 |
| CE31 | polyvinylidene fluoride | PVDF | 221 | 224.7500 | 5.1441 | |
| CE32 | cellulose nitrate | NC | 235 | 242.2000 | 4.4465 | |
| CE30 | nylon | NYLON | 174 | 174.6000 | 3.9005 | |
| CE34 | mixed cellulose ester | MCE_a | 214 | 217.3333 | 4.1399 | |
| CE33 | mixed cellulose ester | MCE_b | 358 | 360.5000 | 5.0208 | |
| CE2 | mixed cellulose ester | S-Pak | 310 | 312.5000 | 5.7927 | |
| CE28 | cellulose acetate and nitrate | PSR009 | 260 | 265.1429 | 5.1996 | |
| CE35 | cellulose acetate and nitrate | PSR009 | 255 | 255.4286 | 5.0427 | |
| CE3 | cellulose acetate and nitrate | PSR010 | 168 | 169.5000 | 3.8749 | |
| Spring | Sample Code | Membrane Code | Biosafety Risk Species for Humans & Animals | ||
|---|---|---|---|---|---|
| 100–500 reads/L | 501–1000 reads/L | >1001 reads/L | |||
| Banpotoc | AE20 | PES | |||
| AE21 | PVDF | ||||
| AE22 | NC | ||||
| AE23 | NYLON | ||||
| AE24 | MCE_a | Acinetobacter junii | |||
| AE25 | MCE_b | ||||
| AE26 | S-Pak | ||||
| AE4 | PSR009 | Comamonas testosteroni Enterobacter kobei | |||
| AE27 | PSR010 | ||||
| Baita | BE11 | PES | |||
| BE12 | PVDF | ||||
| BE18 | NC | ||||
| BE16 | NYLON | ||||
| BE15 | MCE_a | ||||
| BE13 | MCE_b | ||||
| BE19 | S-Pak | ||||
| BE17 | PSR009 | ||||
| BE14 | PSR010 | ||||
| Rapolțel | CE29 | PES | Staphylococcus epidermidis | Acinetobacter junii | |
| CE31 | PVDF | Acinetobacter junii Escherichiafergusonii | Staphylococcus epidermidis | ||
| CE32 | NC | Acinetobacter junii | |||
| CE30 | NYLON | Acinetobacter junii | |||
| CE34 | MCE_a | Acinetobacter junii | |||
| CE33 | MCE_b | Acinetobacter junii | |||
| CE2 | S-Pak | ||||
| CE28 | PSR009 | Atopobium vaginae Corynebacterium tuberculostearicum Gardnerella vaginalis Moraxella osloensis | Acinetobacter junii Staphylococcus epidermidis | ||
| CE35 | PSR009 | Acinetobacter junii Veillonella dispar | |||
| CE3 | PSR010 | Acinetobacter junii | |||
| Spring/ Membrane | Pathogens Identification | Diversity Assessment | Proposed Membrane for Pathogens | Proposed Membrane for Diversity |
|---|---|---|---|---|
| Banpotoc | cellulose actetate and nitrate (PSR009) | nylon (Nylon) | cellulose actetate and nitrate (PSR009) | S-Pak, cellulose actetate and nitrate (PSR009) |
| mixed cellulose ester (MCE_a) | cellulose actetate and nitrate (PSR009) | |||
| mixed cellulose ester (S-Pak) | ||||
| Baita | mixed cellulose ester (MCE-B) | |||
| nylon (Nylon) | ||||
| mixed cellulose ester (S-Pak) | ||||
| Rapoltel | cellulose actetate and nitrate (PSR009) | mixed cellulose ester (MCE_b) | ||
| polyvinylidene fluoride (PVDF) | mixed cellulose ester (S-Pak) | |||
| polyethersulfone (PES) | cellulose actetate and nitrate (PSR009) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moldovan, O.T.; Baricz, A.; Szekeres, E.; Kenesz, M.; Hoaghia, M.A.; Levei, E.A.; Mirea, I.C.; Năstase-Bucur, R.; Brad, T.; Chiciudean, I.; et al. Testing Different Membrane Filters for 16S rRNA Gene-Based Metabarcoding in Karstic Springs. Water 2020, 12, 3400. https://doi.org/10.3390/w12123400
Moldovan OT, Baricz A, Szekeres E, Kenesz M, Hoaghia MA, Levei EA, Mirea IC, Năstase-Bucur R, Brad T, Chiciudean I, et al. Testing Different Membrane Filters for 16S rRNA Gene-Based Metabarcoding in Karstic Springs. Water. 2020; 12(12):3400. https://doi.org/10.3390/w12123400
Chicago/Turabian StyleMoldovan, Oana Teodora, Andreea Baricz, Edina Szekeres, Marius Kenesz, Marial Alexandra Hoaghia, Erika Andrea Levei, Ionuț Cornel Mirea, Ruxandra Năstase-Bucur, Traian Brad, Iulia Chiciudean, and et al. 2020. "Testing Different Membrane Filters for 16S rRNA Gene-Based Metabarcoding in Karstic Springs" Water 12, no. 12: 3400. https://doi.org/10.3390/w12123400