
Double Trouble: Synergy between Habitat Loss and the Spread of the Alien Species Caulerpa cylindracea (Sonder) in Three Mediterranean Habitats

 , , ,
, , ,
Abstract
:1. Introduction
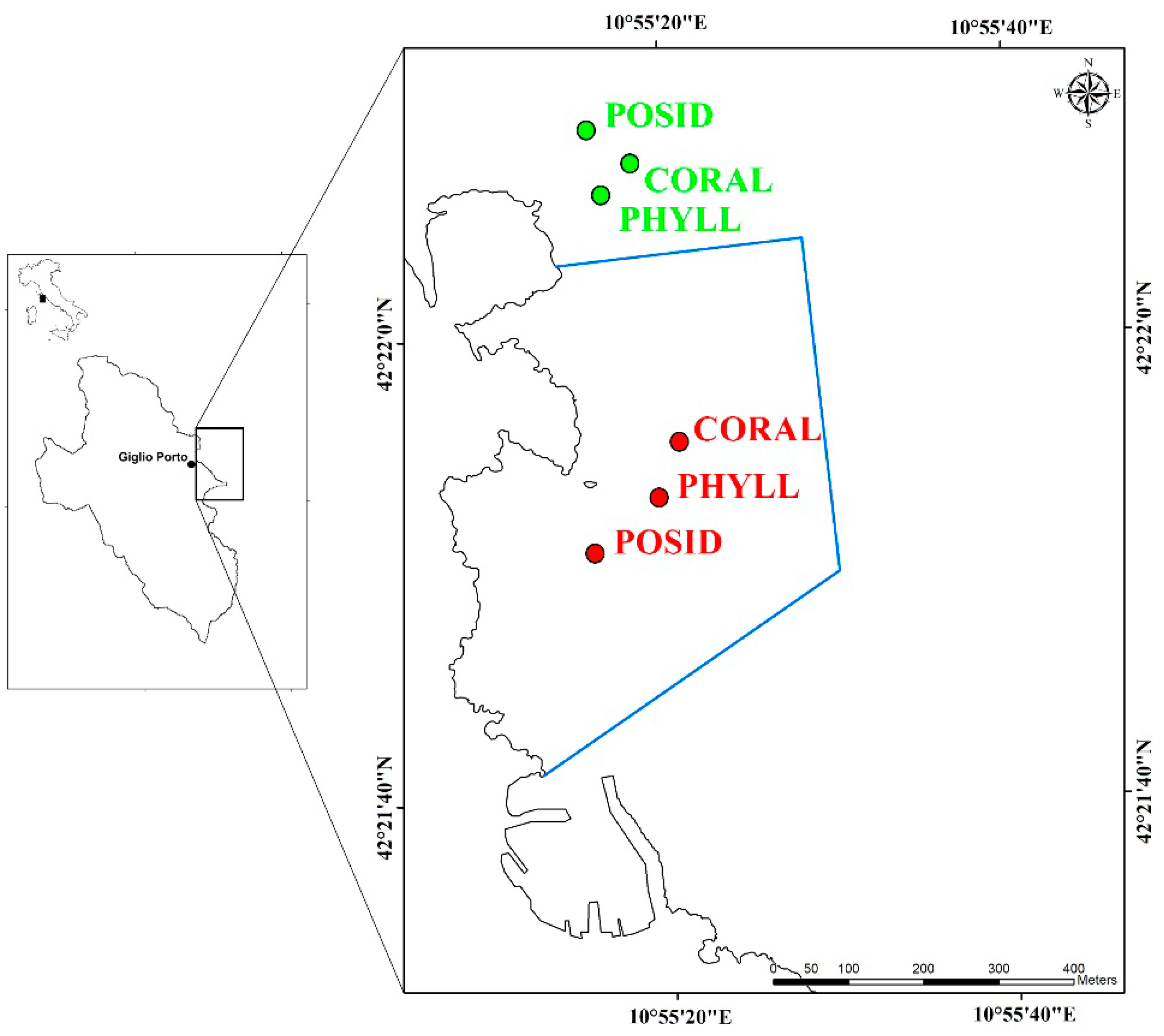
2. Materials and Methods
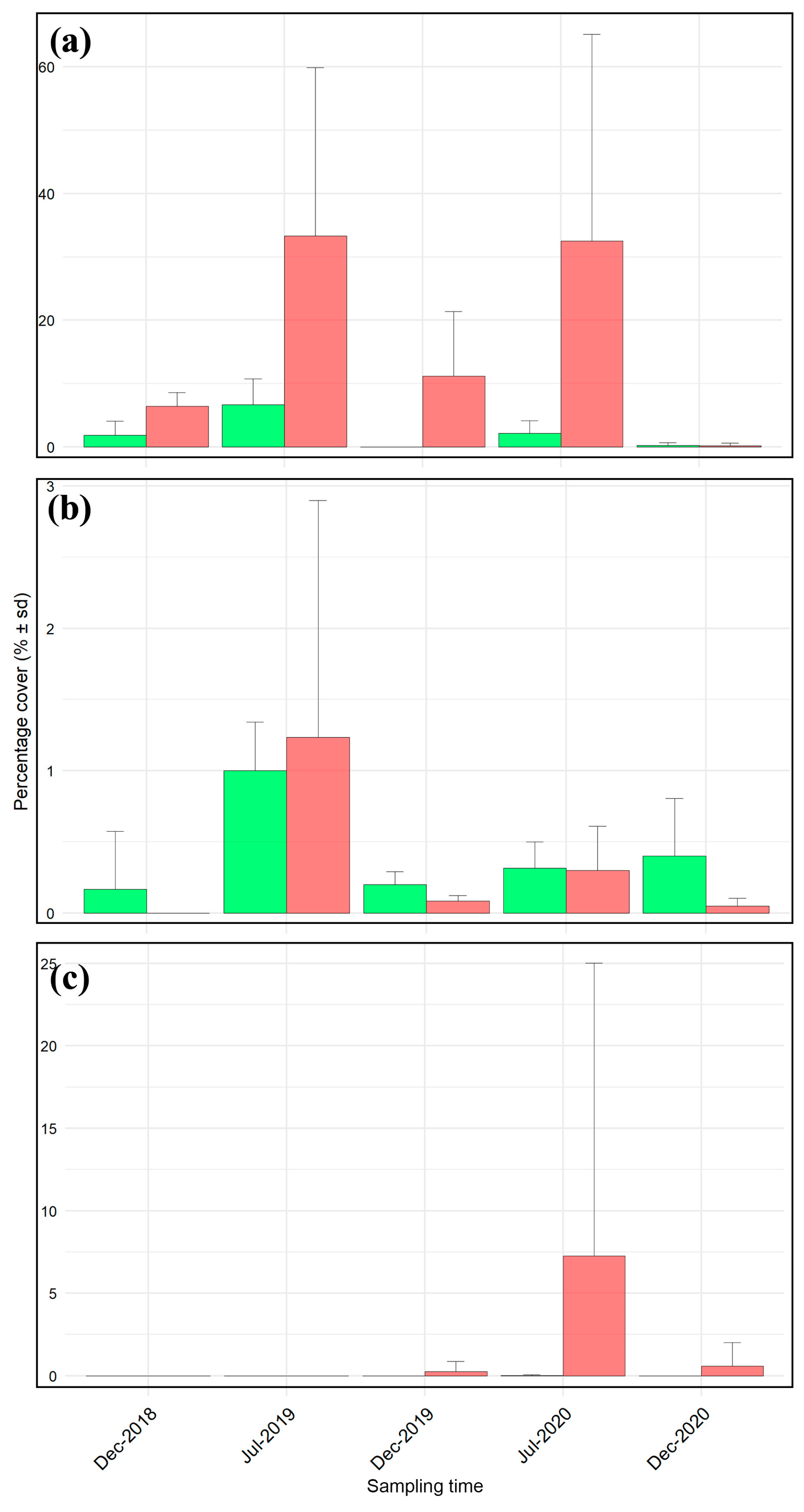
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Coll, M.; Piroddi, C.; Steenbeek, J.; Kaschner, K.; Ben Rais Lasram, F.; Aguzzi, J.; Ballesteros, E.; Bianchi, C.N.; Corbera, J.; Dailianis, J.; et al. The Biodiversity of the Mediterranean Sea: Estimates, patterns, and threats. PLoS ONE 2010, 5, e11842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsiamis, K.; Azzurro, E.; Bariche, M.; Çinar, M.E.; Crocetta, F.; De Clerck, O.; Galil, B.; Gómez, F.; Hoffman, R.; Jensen, K.R.; et al. Prioritizing marine invasive alien species in the European Union through horizon scanning. Aquat. Conserv. Mar. Freshw. Ecosyst. 2020, 30, 794–845. [Google Scholar] [CrossRef]
- Zenetos, A.; Galanidi, M. Mediterranean non indigenous species at the start of the 2020s: Recent changes. Mar. Biodivers. Rec. 2020, 13, 10. [Google Scholar] [CrossRef]
- Orfanidis, S.; Alvito, A.; Azzurro, E.; Badreddine, A.; Ben Souissi, J.; Chamorro, M.; Crocetta, F.; Dalyan, C.; Fortic, A.; Galanti, L.; et al. New alien mediterranean biodiversity records (March 2021). Mediterr. Mar. Sci. 2021, 22, 180–198. [Google Scholar]
- Ragkousis, M.; Abdelali, N.; Azzurro, E.; Badreddine, A.; Bariche, M.; Bitar, G.; Crocetta, F.; Denitto, F.; Digenis, M.; El Zrelli, R. New alien mediterranean biodiversity records (October 2020). Mediterr. Mar. Sci. 2020, 21, 631–652. [Google Scholar]
- Otero, M.; Cebrian, E.; Francour, P.; Galil, B.; Savini, D. Monitoring Marine Invasive Species in Mediterranean Marine Protected Areas (MPAs): A Strategy and Practical Guide for Managers; IUCN Centre for Mediterreanean Cooperation: Malaga, Spain, 2013. [Google Scholar]
- Piazzi, L.; Balata, D.; Bulleri, F.; Gennaro, P.; Ceccherelli, G. The invasion of Caulerpa cylindracea in the Mediterranean: The known, the unknown and the knowable. Mar. Biol. 2016, 163, 1–14. [Google Scholar] [CrossRef]
- Verlaque, M.; Durand, C.; Huisman, J.M.; Boudouresque, C.F.; Le Parco, Y. On the identity and origin of the Mediterranean invasive Caulerpa racemosa (Caulerpales, Chlorophyta). Eur. J. Phycol. 2003, 38, 325–339. [Google Scholar] [CrossRef] [Green Version]
- Buia, M.C.; Petrocelli, A.; Saracino, O.D. Caulerpa racemosa spread in the Mediterranean Sea: First record in the Gulf of Taranto. Biol. Mar. Mediterr. 1998, 5, 527–529. [Google Scholar]
- Piazzi, L.; Balata, D.; Ceccherelli, G.; Cinelli, F. Comparative study of the growth of the two co-occurring introduced green algae Caulerpa taxifolia and Caulerpa racemosa along the Tuscan coast (Italy, western Mediterranean). Cryptogam. Algol. 2001, 22, 459–466. [Google Scholar] [CrossRef]
- Piazzi, L.; Meinesz, A.; Verlaque, M.; Akcali, B.; Antolic, B.; Argyrou, M.; Balata, D.; Ballesteros, E.; Calvo, S.; Cinelli, F. Invasion of Caulerpa racemosa var. cylindracea (Caulerpales, Chlorophyta) in the Mediterranean Sea: An assessment of the spread. Cryptogamie-Algologie 2005, 26, 189–202. [Google Scholar]
- Streftaris, N.; Zenetos, A. Alien marine species in the Mediterranean-the 100 ‘Worst Invasives’ and their impact. Mediterr. Mar. Sci. 2006, 7, 87–118. [Google Scholar] [CrossRef] [Green Version]
- Ceccherelli, G.; Pinna, S.; Cusseddu, V.; Bulleri, F. The role of disturbance in promoting the spread of the invasive seaweed Caulerpa racemosa in seagrass meadows. Biol. Invasions 2014, 16, 2737–2745. [Google Scholar] [CrossRef]
- Bulleri, F.; Alestra, T.; Ceccherelli, G.; Tamburello, L.; Pinna, S.; Sechi, N.; Benedetti-Cecchi, L. Determinants of Caulerpa racemosa distribution in the north-western Mediterranean. Mar. Ecol. Prog. Ser. 2011, 431, 55–67. [Google Scholar] [CrossRef] [Green Version]
- Gennaro, P.; Piazzi, L.; Persia, E.; Porrello, S. Nutrient exploitation and competition strategies of the invasive seaweed Caulerpa cylindracea. Eur. J. Phycol. 2015, 50, 384–394. [Google Scholar] [CrossRef]
- Bulleri, F.; Piazzi, L. Variations in importance and intensity of competition underpin context dependency in the effects of an invasive seaweed on resident assemblages. Mar. Biol. 2015, 162, 485–489. [Google Scholar] [CrossRef]
- Piazzi, L.; Ceccherelli, G. Persistence of biological invasion effects: Recovery of macroalgal assemblages after removal of Caulerpa racemosa var. cylindracea. Estuar. Coast. Shelf Sci. 2006, 68, 455–461. [Google Scholar] [CrossRef]
- Mancini, G.; Casoli, E.; Ventura, D.; Jona-Lasinio, G.; Criscoli, A.; Belluscio, A.; Ardizzone, G.D. Impact of the Costa Concordia shipwreck on a Posidonia oceanica meadow: A multi-scale assessment from a population to a landscape level. Mar. Pollut. Bull. 2019, 148, 168–181. [Google Scholar] [CrossRef]
- Casoli, E.; Mancini, G.; Ventura, D.; Pace, D.S.; Belluscio, A.; Ardizzone, G.D. Reteporella spp. success in the re-colonization of bare coralligenous reefs impacted by Costa Concordia shipwreck: The pioneer species you did not expect. Mar. Pollut. Bull. 2020, 161, 111808. [Google Scholar] [CrossRef]
- European Parliament; Council of the European Union. MSFD 2008/56/EC of the European Parliament and of the Council, of 17 June 2008, Establishing a Framework for Community Action in the Field of Marine Environmental Policy (Marine Strategy Framework Directive); Official Journal of the European Union: Luxembourg, 2008. [Google Scholar]
- Bonifazi, A.; Ventura, D.; Gravina, M.F.; Lasinio, G.J.; Belluscio, A.; Ardizzone, G.D. Unusual algal turfs associated with the rhodophyta Phyllophora crispa: Benthic assemblages along a depth gradient in the Central Mediterranean Sea. Estuar. Coast. Shelf Sci. 2017, 185, 77–93. [Google Scholar] [CrossRef]
- Marín-Guirao, L.; Bernardeau-Esteller, J.; Ruiz, J.M.; Sandoval-Gil, J.M. Resistance of Posidonia oceanica seagrass meadows to the spread of the introduced green alga Caulerpa cylindracea: Assessment of the role of light. Biol. Invasions 2015, 17, 1989–2009. [Google Scholar] [CrossRef]
- Montefalcone, M.; Morri, C.; Parravicini, V.; Bianchi, C.N. A tale of two invaders: Divergent spreading kinetics of the alien green algae Caulerpa taxifolia and Caulerpa cylindracea. Biol. Invasions 2015, 17, 2717–2728. [Google Scholar] [CrossRef]
- Montefalcone, M.; Morri, C.; Peirano, A.; Albertelli, G.; Bianchi, C.N. Substitution and phase shift within the Posidonia oceanica seagrass meadows of NW Mediterranean Sea. Estuar. Coast. Shelf Sci. 2007, 75, 63–71. [Google Scholar] [CrossRef]
- Boudouresque, C.F.; Verlaque, M. An overview of species introduction and invasion processes in marine and coastal lagoon habitats. CBM-Cahiers Biol. Mar. 2012, 53, 309. [Google Scholar]
- Piazzi, L.; Cecchi, E.; Gennaro, P.; Penna, M.; Trabucco, B.; Ceccherelli, G. Spread of non-indigenous macroalgae and disturbance: Impact assessment of the Costa Concordia shipwreck (Giglio Island, Italy) using the ALEX index. Ocean Coast. Manag. 2019, 183, 104999. [Google Scholar] [CrossRef]
- Bulleri, F.; Balata, D.; Bertocci, I.; Tamburello, L.; Benedetti-Cecchi, L. The seaweed Caulerpa racemosa on Mediterranean rocky reefs: From passenger to driver of ecological change. Ecology 2010, 91, 2205–2212. [Google Scholar] [CrossRef]
- Piazzi, L.; Cinelli, F. Distribution and dominance of two introduced turf-forming macroalgae on the coast of Tuscany, Italy, northwestern Mediterranean Sea in relation to different habitats and sedimentation. Bot. Mar. 2001, 44, 509–520. [Google Scholar] [CrossRef]
- Gennaro, P.; Piazzi, L. The indirect role of nutrients in enhancing the invasion of Caulerpa racemosa var cylindracea. Biol. Invasions 2014, 16, 1709–1717. [Google Scholar] [CrossRef]
- Piazzi, L.; Balata, D.; Cinelli, F. Invasions of alien macroalgae in Mediterranean coralligenous assemblages. Cryptogam. Algol. 2007, 28, 289–301. [Google Scholar]
- Capiomont, A.; Breugnot, E.; den Haan, M.; Meinesz, A. Phenology of a deep-water population of Caulerpa racemosa var. cylindracea in the northwestern Mediterranean Sea. Bot. Mar. 2005, 48, 80–83. [Google Scholar] [CrossRef]
- Baldacconi, R.; Corriero, G. Effects of the spread of the alga Caulerpa racemosa var. cylindracea on the sponge assemblage from coralligenous concretions of the Apulian coast (Ionian Sea, Italy). Mar. Ecol. 2009, 30, 337–345. [Google Scholar] [CrossRef]
- Gennaro, P.; Piazzi, L. Synergism between two anthropic impacts: Caulerpa racemosa var. cylindracea invasion and seawater nutrient enrichment. Mar. Ecol. Prog. Ser. 2011, 427, 59–70. [Google Scholar] [CrossRef] [Green Version]
- Piazzi, L.; Balata, D.; Cecchi, E.; Cinelli, F.; Sartoni, G. Species composition and patterns of diversity of macroalgal coralligenous assemblages in the north-western Mediterranean Sea. J. Nat. Hist. 2009, 44, 1–22. [Google Scholar] [CrossRef]
- Piazzi, L.; Gennaro, P.; Balata, D. Threats to macroalgal coralligenous assemblages in the Mediterranean Sea. Mar. Pollut. Bull. 2012, 64, 2623–2629. [Google Scholar] [CrossRef]
- Balata, D.; Piazzi, L.; Bulleri, F. Sediment deposition dampens positive effects of substratum complexity on the diversity of macroalgal assemblages. J. Exp. Mar. Biol. Ecol. 2015, 467, 45–51. [Google Scholar] [CrossRef]
- Piazzi, L.; Atzori, F.; Cadoni, N.; Cinti, M.F.; Frau, F.; Ceccherelli, G. Monitoring non-indigenous macroalgae in a Mediterranean MPA: Lessons from a short-temporal variability of pristine habitats invasion. Ocean Coast. Manag. 2021, 207, 105608. [Google Scholar] [CrossRef]
- Ruitton, S.; Verlaque, M.; Boudouresque, C.F. Seasonal changes of the introduced Caulerpa racemosa var. cylindracea (Caulerpales, Chlorophyta) at the northwest limit of its Mediterranean range. Aquat. Bot. 2005, 82, 55–70. [Google Scholar] [CrossRef]
- Madricardo, F.; Foglini, F.; Kruss, A.; Ferrarin, C.; Pizzeghello, N.M.; Murri, C.; Rossi, M.; Bajo, M.; Bellafiore, D.; Campiani, E.; et al. High resolution multibeam and hydrodynamic datasets of tidal channels and inlets of the Venice Lagoon. Sci. Data 2017, 4, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Janowski, L.; Madricardo, F.; Fogarin, S.; Kruss, A.; Molinaroli, E.; Kubowicz-Grajewska, A.; Tegowski, J. Spatial and temporal changes of tidal inlet using object-based image analysis of multibeam echosounder measurements: A case from the Lagoon of Venice, Italy. Remote Sens. 2020, 12, 2117. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| P. oceanica meadow | |||||
| Source of variation | Df | SS | MS | F-value | p-value |
| Condition | 1 | 25.47 | 25.46 | 38.97 | <0.001 |
| Time | 4 | 37.49 | 9.37 | 14.34 | <0.001 |
| Condition × Time | 4 | 8.50 | 2.12 | 3.25 | 0.02 |
| Residuals | 50 | 32.67 | 0.65 | ||
| P. crispa turf | |||||
| Source of variation | Df | SS | MS | F-value | p-value |
| Condition | 1 | 0.18 | 0.18 | 2.87 | 0.09 |
| Time | 4 | 2.65 | 0.66 | 10.53 | <0.001 |
| Condition × Time | 4 | 0.09 | 0.02 | 0.38 | 0.82 |
| Residuals | 50 | 3.14 | 0.06 | ||
| Coralligenous reef | |||||
| Source of variation | Df | SS | MS | F-value | p-value |
| Condition | 1 | 0.62 | 0.62 | 2.14 | 0.15 |
| Time | 4 | 0.85 | 0.21 | 0.73 | 0.57 |
| Condition × Time | 4 | 0.77 | 0.19 | 0.66 | 0.62 |
| Residuals | 50 | 14.59 | 0.29 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Casoli, E.; Mancini, G.; Ventura, D.; Belluscio, A.; Ardizzone, G. Double Trouble: Synergy between Habitat Loss and the Spread of the Alien Species Caulerpa cylindracea (Sonder) in Three Mediterranean Habitats. Water 2021, 13, 1342. https://doi.org/10.3390/w13101342
Casoli E, Mancini G, Ventura D, Belluscio A, Ardizzone G. Double Trouble: Synergy between Habitat Loss and the Spread of the Alien Species Caulerpa cylindracea (Sonder) in Three Mediterranean Habitats. Water. 2021; 13(10):1342. https://doi.org/10.3390/w13101342
Chicago/Turabian StyleCasoli, Edoardo, Gianluca Mancini, Daniele Ventura, Andrea Belluscio, and Giandomenico Ardizzone. 2021. "Double Trouble: Synergy between Habitat Loss and the Spread of the Alien Species Caulerpa cylindracea (Sonder) in Three Mediterranean Habitats" Water 13, no. 10: 1342. https://doi.org/10.3390/w13101342