1. Introduction
Unraveling the effects of climate change and its impact on natural systems has been a central goal in ecological studies, being particularly important in the context of global change [
1,
2]. Numerous approaches have been used to study the effects of climate change in shallow lakes [
3], including warming experiments [
4,
5,
6], mathematical models [
7], paleolimnological records [
8], time series [
9], and space-for-time substitution (SFTS) [
10]. In particular, SFTS has provided major empirical evidence of climate change effects in lakes, including shallow lake systems [
3,
10]. In this approach, comparisons were conducted between systems in contrasting climates where the organisms had enough time to evolve and adapt to the local climate. However, apparent climate effects determined by SFTS may in part reflect differences on biogeography, and may be influenced by co-variates of temperature and the latitudinal variation in other climate-related variables, such as seasonality and the length of the growing season. Similar studies focusing on species-poor areas, such as remote islands located in contrasting climates with a recent fish introduction, may be useful to somewhat separate the effect of confounding factors occurring at a large, latitudinal scale in the mainland [
3].
Studies conducted along climate gradients suggest strong effects of increases in ambient temperatures on lake fish community structures and dynamics, as reviewed in [
3]. Reduced abundance of cold stenothermal fish species and enhanced abundance of eurythermal fish species, even in deep, stratified lakes, are expected with decreasing latitude [
11]. Moreover, fish in warmer climates have higher specific metabolic and excretion rates, as well as earlier reproduction and prolonged spawning [
12,
13,
14]. Moreover, a decrease in mean fish body size [
15,
16,
17,
18,
19,
20] is evident with decreasing latitude and have been attributed particularly to a significant switch in life history [
19], and sometimes to intra-species density depended changes [
21]. Moreover an increase in taxonomic and functional richness [
16,
17,
18,
22] as well as changes in functional composition with increased proportion of omnivorous and herbivorous fish species [
23,
24], have been observed with decreasing latitude.
Due to reduced mean size, among other expected changes in fish community structure and function at warmer regions, a higher zooplanktivory is expected [
25,
26,
27]. The higher fish predation pressure on zooplankton reduces the grazing pressure on phytoplankton [
10] and the maintenance of clear water at the ecosystem level is consequently less frequent in warm than in cold climates [
28].
The described differences at community level can affect various food web metrics, including the trophic web length (TWL), commonly measured as range in δ
15N (NR). TWL has been one of the most studied metrics of trophic webs and several hypotheses and mechanisms have been proposed to explain its variations. One is the energetic limitation hypothesis (ELH) [
29,
30,
31]; and references therein, according to which energy losses between trophic levels limit the maximum food web length, is susceptible to temperature changes. A negative correlation between the maximum trophic position and environmental temperature is theoretically expected [
32], a pattern already revealed in empirical studies [
33,
34,
35].
The width of the food web, represented as δ
13C range (hereafter CR), depends largely on the stable isotope signature of primary producers [
36], but as their signal is highly variable among ecosystems, primary consumer’s signatures have been proposed to better integrate the natural variability in space and time of the basal resources (producers) [
36]. The carbon signal (δ
13C) was thought to be transmitted with little change from the primary producers to the consumers; consequently, the consumer signals depend directly on the signals of their diets [
37]. However, a large variability in trophic fractionations has been reported for aquatic consumers [
38]. In contrast to nitrogen, the theoretical framework developed for carbon is patchier, particularly in connection with climate and temperature in particular [
39]. In low productivity lakes, the carbon sources for photosynthesis usually derive from mineralization in the benthic and littoral zones, which are typically depleted in δ
13C compared to atmospheric carbon signature [
40]. In productive lakes, however, endogenous carbon may not suffice to maintain pelagic photosynthesis and pelagic producers therefore have to exploit a higher proportion of atmospheric inorganic carbon, enriched in δ
13C [
40,
41]. An enriched δ
13C signal, due to input of atmospheric carbon in the pelagic zone in the warmer (and consequently, often more productive) lakes is thus expected, which may reduce the range of δ
13C of producers in warmer systems.
Biotic interactions could also lead to differences in CR as fish predation may affect the abundance, composition, size structure, and/or the diet behavior of macroinvertebrates [
42,
43,
44], and consequently reduce their use of resources. Accordingly, lake food webs with stronger fish predation, namely warm low latitude systems, would potentially exhibit a narrower CR; this has previously been observed in streams [
45].
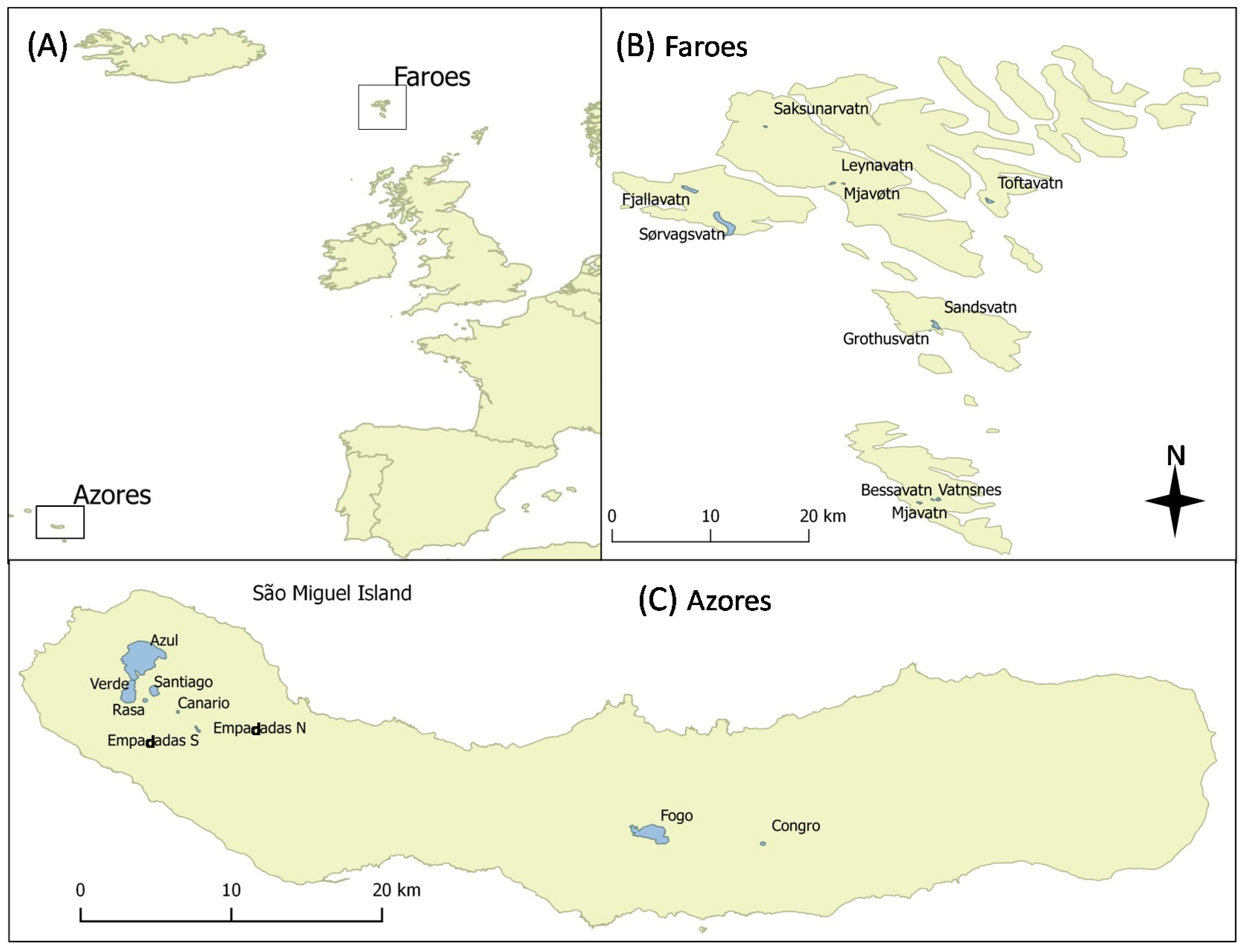
To reduce some of the weakness of SFTS performed in continental lakes, often positioned in distant regions with different seasonality, biogeography, and evolutionary time [
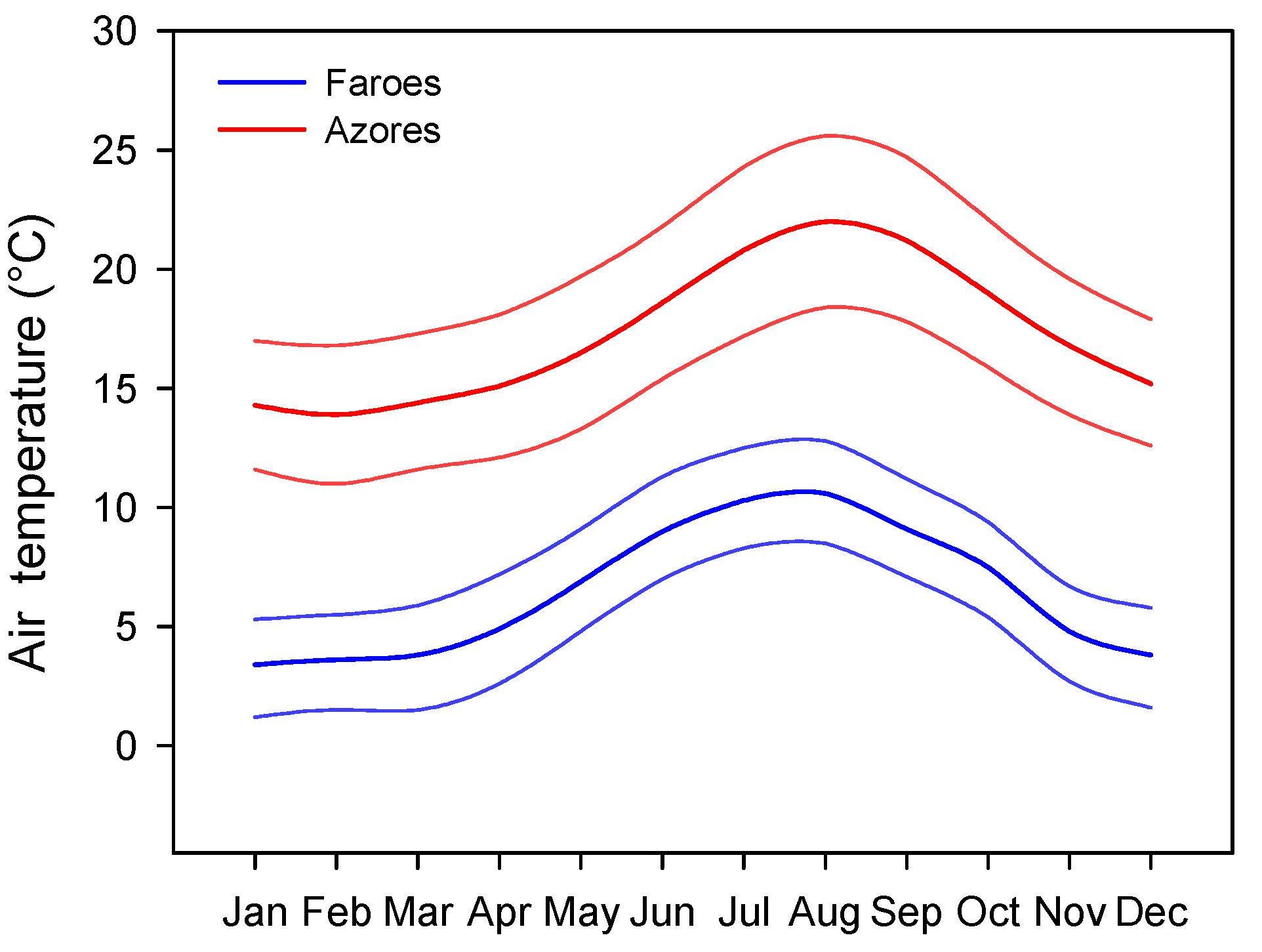
3], we studied two sets of species-poor lakes located in two remote groups of islands in the North Atlantic Ocean with contrasting climates, but comparable seasonality, namely the Azores (warm) and the Faroe (cold) Islands. Since both island groups are located in the North Atlantic Ocean, the studied lakes are subject to the same seasonality (
Figure 1).
Records of freshwater fish communities in both sets of islands trace back to the 17th and 18th centuries [
46,
47]. Fish were first introduced by humans in the Azores Islands in 1792 [
47], while the fish species at the Faroe Island are indigenous, but have been transplanted internally [
46,
48]. Most of the fish species in the Azores Islands were stocked for the first time one to two hundred years ago. The first introduced species in 1792 was crucian carp
Carassius auratus Linnaeus 1758. Most of the exotic species were introduced during the 19th century; ruivaca
Achondrostoma oligolepis (Robalo, Doadrio, Almada and Kottelat 2005) was introduced in 1879; brown trout
Salmo trutta Linnaeus 1758 in 1880; common carp
Cyprinus carpio Linnaeus 1758 in 1890; roach
Rutilus rutilus (Linnaeus 1758) in 1895; largemouth black bass
Micropterus salmoides (Lacepède 1802) in 1898 and perch
Perca fluviatilis Linnaeus 1758 in 1898. Three species have been more recently stocked, rainbow trout
Oncorhynchus mykiss (Walbaum 1792) in 1941, pike
Esox Lucius Linnaeus 1758 in 1979 and finally pikeperch
Sander lucioperca (Linnaeus 1758) in 1980.
We used a stable isotope approach to test the hypothesis that ambient temperature differences between the two island groups play a key role in shaping food webs. Based on evidence from mainland lakes, we expected to find: (1) a fish size structure dominated by small individuals in the warmer lakes (Azores Island); (2) a stronger top-down effect of fish in the warmer lakes than in the colder ones (Faroe Island); (3) shorter food webs in the warmer lakes; and (4) a wider (CR) food web in the colder lakes.
4. Discussion
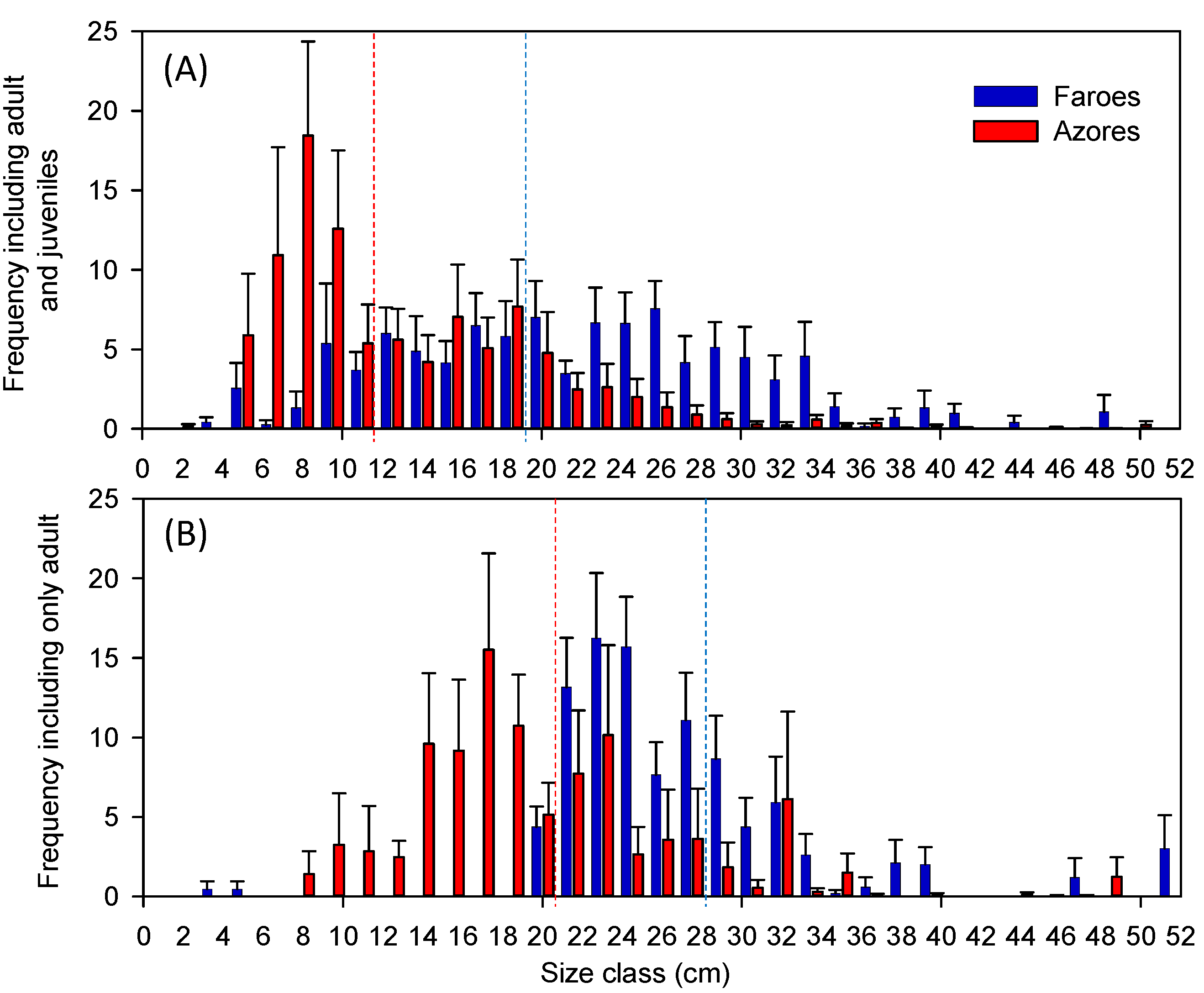
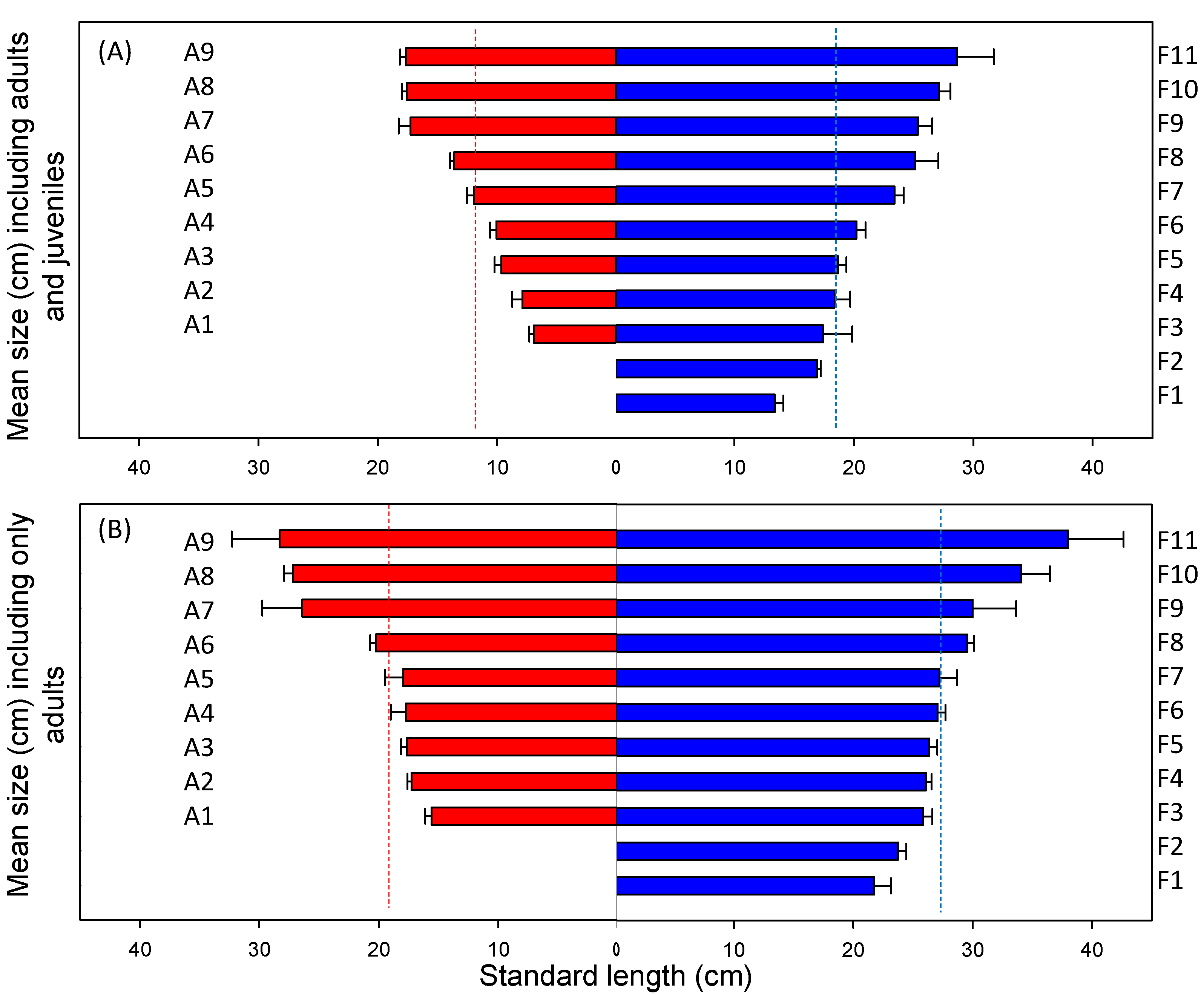
As predicted, we found smaller mean body size and a higher frequency of small-bodied fish in the warmer Azorean region than in the colder Faroese lakes. A random collection of introduced species or random events, for example, a particularly strong recruitment year or differences in life history and habitat use of the juveniles [
19] could, potentially, produce such patterns. Additionally, the composition of fish communities differed between regions (i.e., Faroese lakes being dominated by salmonids), life history, and habitat use may vary as well. A larger mean fish size could, potentially, be due to lack of sufficient habitats where juveniles may occur (e.g., streams where the salmonids spawn [
19], though likely not the case for the Faroe Islands lakes as they are well connected to streams [
46]. However, fish in the warmer Azorean lakes were smaller, even when the size of each fish species was standardized by the maximum body size of resident forms within their continental distribution and when the analysis was performed, excluding juveniles (determined from maturity sizes reported in the literature). The differences in the size structure of the fish communities between the two climatic regions thus followed the pattern obtained along climate gradients or pair-comparisons in continental lakes [
16,
17,
67], as well as within lakes subjected to climate warming [
11,
68].
Many ectothermic animals experience faster growth at higher temperatures and, consequently, have a smaller body size at the same age than animals subject to lower temperatures (the temperature-size rule, [
69,
70]. High temperatures often extend beyond the reproduction season of a particular species and enhance fry survival, increasing the abundance of small, young individuals, thus leading to a reduction in the mean size of the fish assemblage [
12,
15]. This may have contributed likely the smaller-sized fish community structure in the Azores Islands. Other mechanisms, more indirectly linked with higher water temperatures, could contribute to the observed patterns. Small fish usually cope better with low oxygen than their bigger counterparts [
71] and particularly a reduced growth under low oxygen levels has been described for salmonids [
72]. Consequently, high-temperature driven hypoxia may act as a filter, affecting especially large piscivorous fish, and contributing to the dominance of smaller omnivorous individuals.
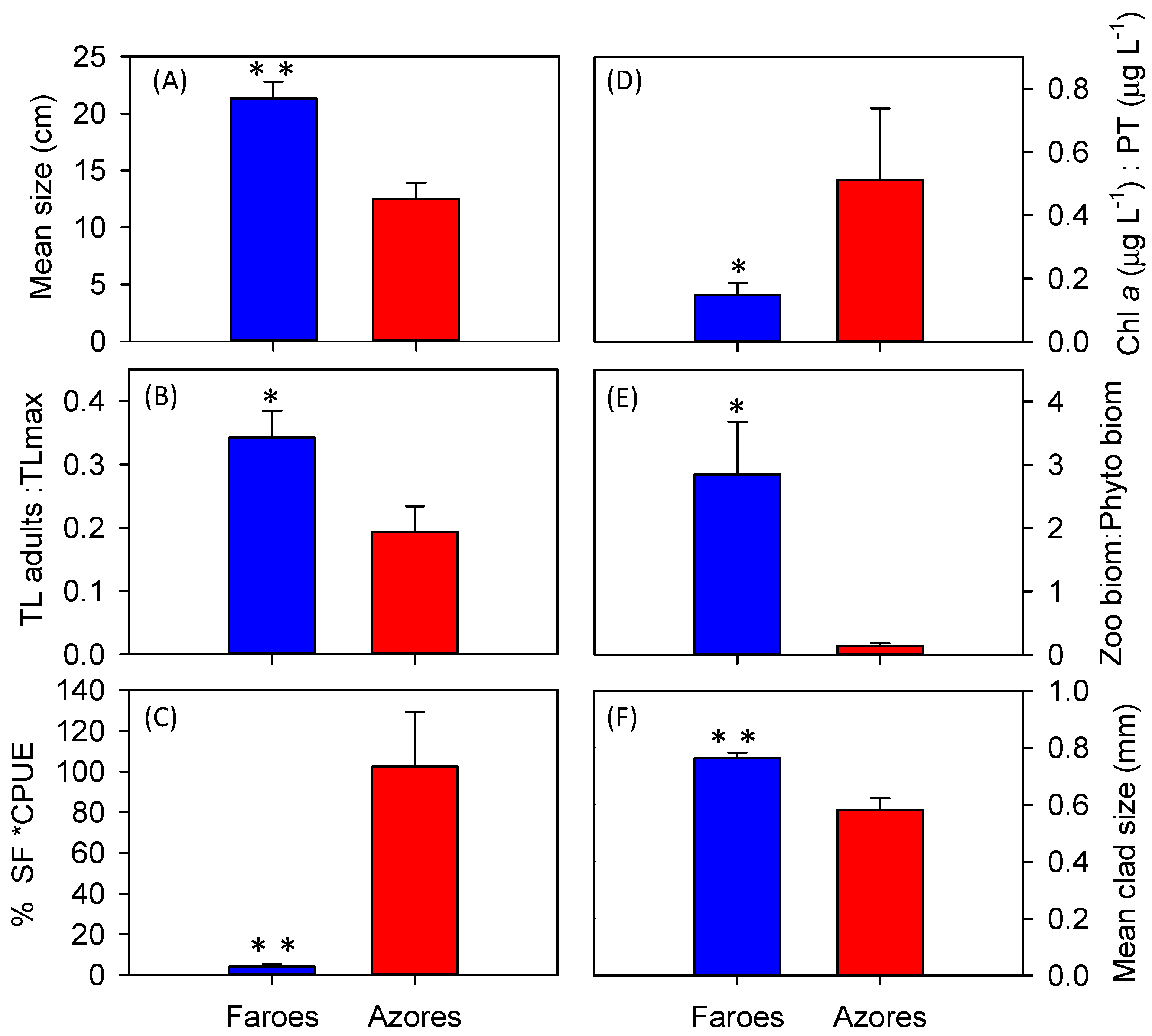
Whatever the mechanism, the smaller size of fish in warmer lakes typically leads to a higher predation pressure on zooplankton as showed by [
10,
25,
26,
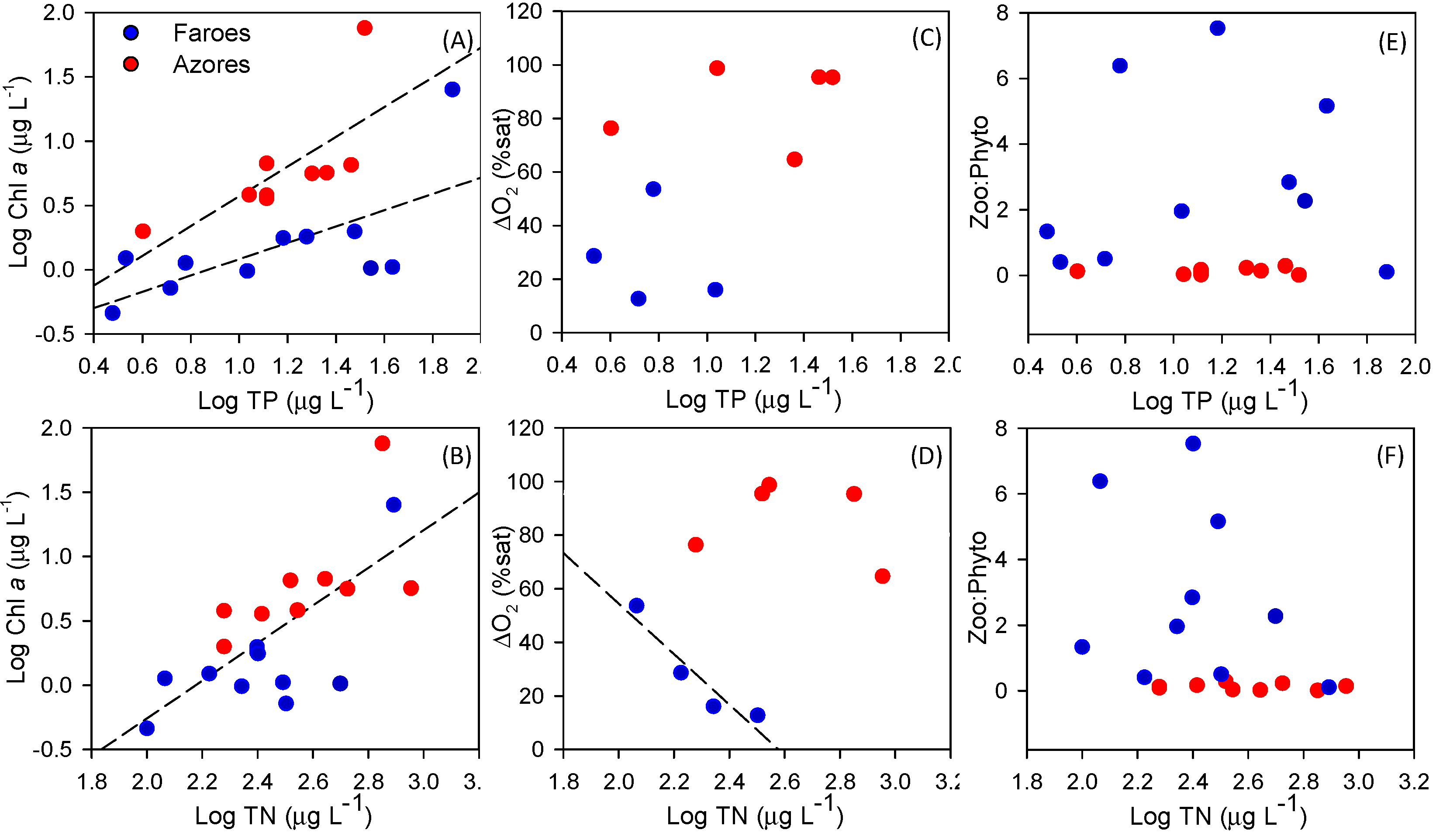
27]. Supporting our second hypothesis, about stronger cascading effects of fish on zooplankton and phytoplankton in warm lakes, we found a smaller mean size of cladocerans and lower zooplankton:phytoplankton biomass ratio in the Azorean lakes, suggesting the occurrence of lower grazing pressure on phytoplankton compared to the Faroese lakes. This may explain the higher Chl-
a per unit of TP and TN in the Azores Islands. Other mechanisms, indirectly related to higher temperatures, could also enhance phytoplankton biomass. The near-bottom depletion of oxygen may have promoted phosphorus release from the sediments [
73] in the warm, stratified lakes (TP concentration were higher here); whereas, temperature increases as well as reduced body size could lead to an increase in nutrient cycling by the fish community [
74,
75].
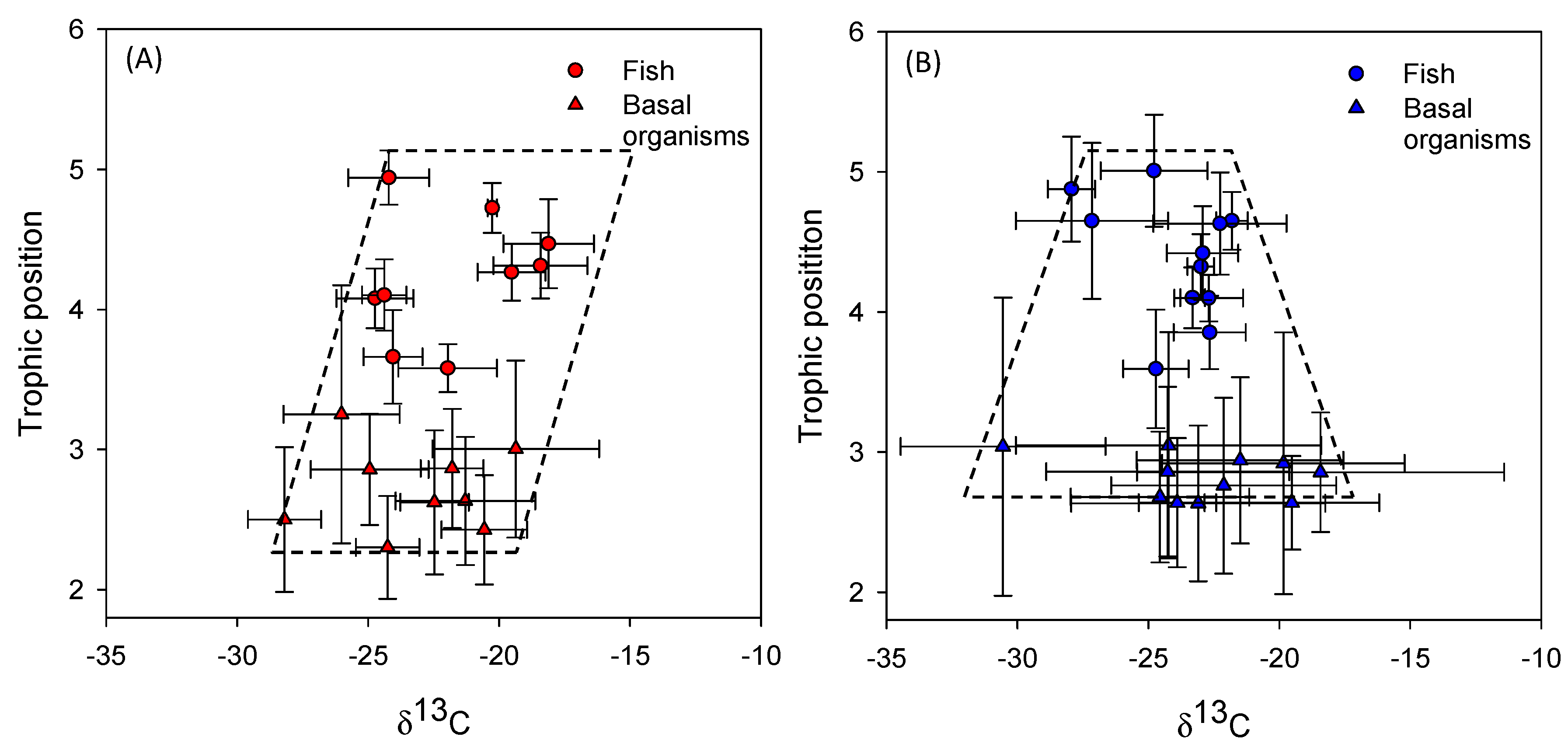
As expected, we found a wider CR for basal organisms in the Faroese lakes. However, contrary to our expectations, the CR of the basal organisms in the Azorean lakes was not related to the proportion of small fish or the fish predation indicators as in the Faroese lakes. Morphometric variables (i.e., area and depth) and transparency (Secchi depth) were positively correlated with the CR of basal organisms in the Faroese lakes. Similarly, lake area and trophic state have been observed to affect the δ
13C signature in the pelagic zone of a wide range of lake areas [
41]. This indicates that a combination of biotic and abiotic factors influenced the CR of basal organisms in the colder Faroese lakes. As the Faroese lakes were relatively (i.e., considering water depth) more transparent than the Azorean lakes, a higher benthic production could supposedly be sustained [
76,
77] in the former. This is supported by the positive relationships between the CR of basal organisms and Secchi disk depth. No such relationships were detected in the Azorean lakes, indicating that in this region other factors may be important in determining the CR of basal organisms. Low dissolved oxygen concentrations at the bottom can negatively affect benthic macroinvertebrates [
78,
79], often leading to the dominance of a few hypoxic-tolerant species, such as oligochaetes and chironomids [
80], with likely lower trophic diversity. Supporting this view, we found a smaller CR and reduced richness of macroinvertebrates in the benthic samples of the Azorean lakes compared to the Faroese lakes.
The δ
13C signal in the pelagic zone is expected to be higher (enriched) in productive [
41] and hydrologically stable systems [
81], both characteristics stimulated by higher temperatures [
82,
83]. The potentially higher productivity of the warmer systems may indirectly narrow CR by increasing the pelagic δ
13C signal. Moreover, the CO
2 flux between a lake and the atmosphere may be affected by the food web structure [
84,
85]. A higher phytoplankton biomass due to diminished zooplankton biomass as a consequence of, for example, high fish predation, may enhance the influx of CO
2 from the atmosphere and thus enrich the δ
13C signal of the pelagic biota [
84].
Despite the differences in CR at the consumer level, no differences were found in fish community CR between the two climatic regions. However, the Azorean fishes showed higher CR when standardized relative to the basal organisms, indicating a greater diversification and a higher efficiency in the use of the resources.
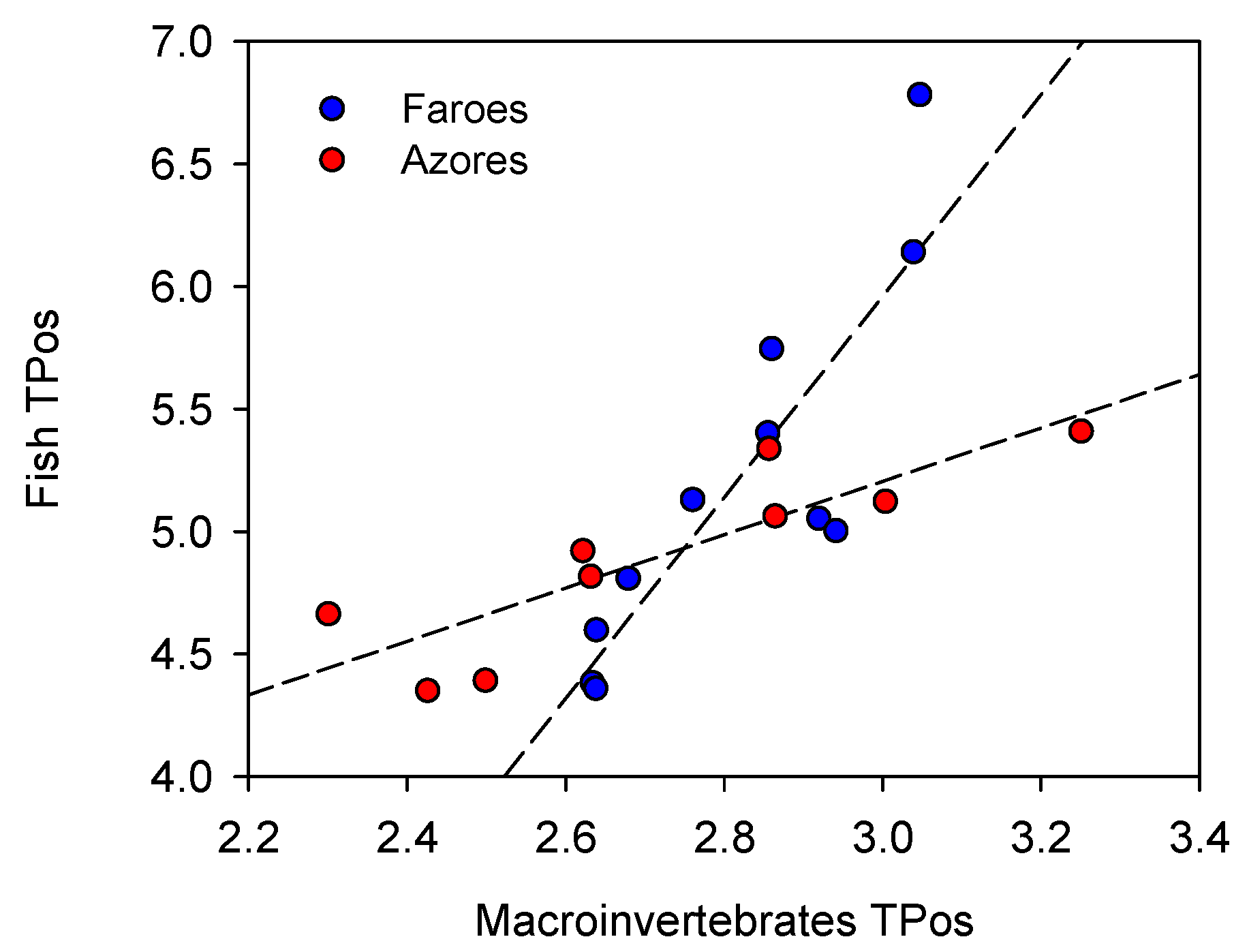
According to the energetic limitation hypothesis (ELH) [
29,
30,
31,
32], we should expect lower trophic web lengths (TPos and NR) in the warmer regions, but this was not the case in our study. Additionally, a higher proportion of herbivorous fish species is expected in warm lakes [
23,
24]. Accordingly, the Azorean fish communities were dominated by potentially omnivorous and herbivorous species (up to 50% of abundance the total capture): roach [
86] (19%), ruivaca [
87] (18%), and common carp [
88] (13%). It is accepted that fractionation of nitrogen vary between carnivorous and herbivorous fish, depending on the diet quality (in terms of C:N ratio) and food processing mechanisms [
89]. Typically, herbivorous fish exhibit higher per-trophic-level isotope enrichment of nitrogen [
90], and consequently, may increase their trophic position more than expected for carnivorous fish. This perhaps explains why we did not find a shorter total food web length in the Azorean lakes. Supporting this view, the fish in the Azorean lakes showed a higher CR relative to the CR of basal organisms, likely reflecting that fish in warmer regions include additional items to the diet such as vegetal material. We cannot, however, fully rule out that the difference in initial fish stock compositions and successions might have affected the food web structure and, thus, the reliability of the comparison of the food web metrics between the two sets of islands.
The space-for-time substitution approach can be criticized as it does not account for biogeographical factors (often leading to high fish species richness at low latitudes) and seasonality differences, thus potentially confounding the effects of climate variability and climate change. The insular lakes used in our study are exposed to the same seasonality in temperature and inhabited by few fish species (some capable of reaching large body sizes) that were introduced from different climatic regions after the 18th century, as in the case of the Azorean lakes. Our data confirmed the dominance of small-sized fishes in the Azorean lakes, which allows us to cautiously suggest that, in insular lakes, fish assemblages under warm conditions reach smaller sizes. Consequently, we conclude that climate differences, and particularly differences in ambient temperature, have played a key role in the divergence in body size of fish between the two groups of islands leading to different cascading effects among climatic regions. Our results further strengthen the argument that biography is not of key importance for the functional changes occurring in lakes along a temperature gradient. Moreover, our findings suggest that changes in trophic web dynamics and functioning, such as the width at the base of the food web, but not necessarily in the trophic web length, can be expected under current and future warmer climates.


 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}