
Interacting Flooding and Competition Negatively Affect Growth of Riparian Species Dominating a Reservoir Shoreline

Abstract
:1. Introduction
2. Material and Methods
2.1. Plant Species
2.2. Plant Material
2.3. Evaluating the Vertical Seed Dispersal Potential
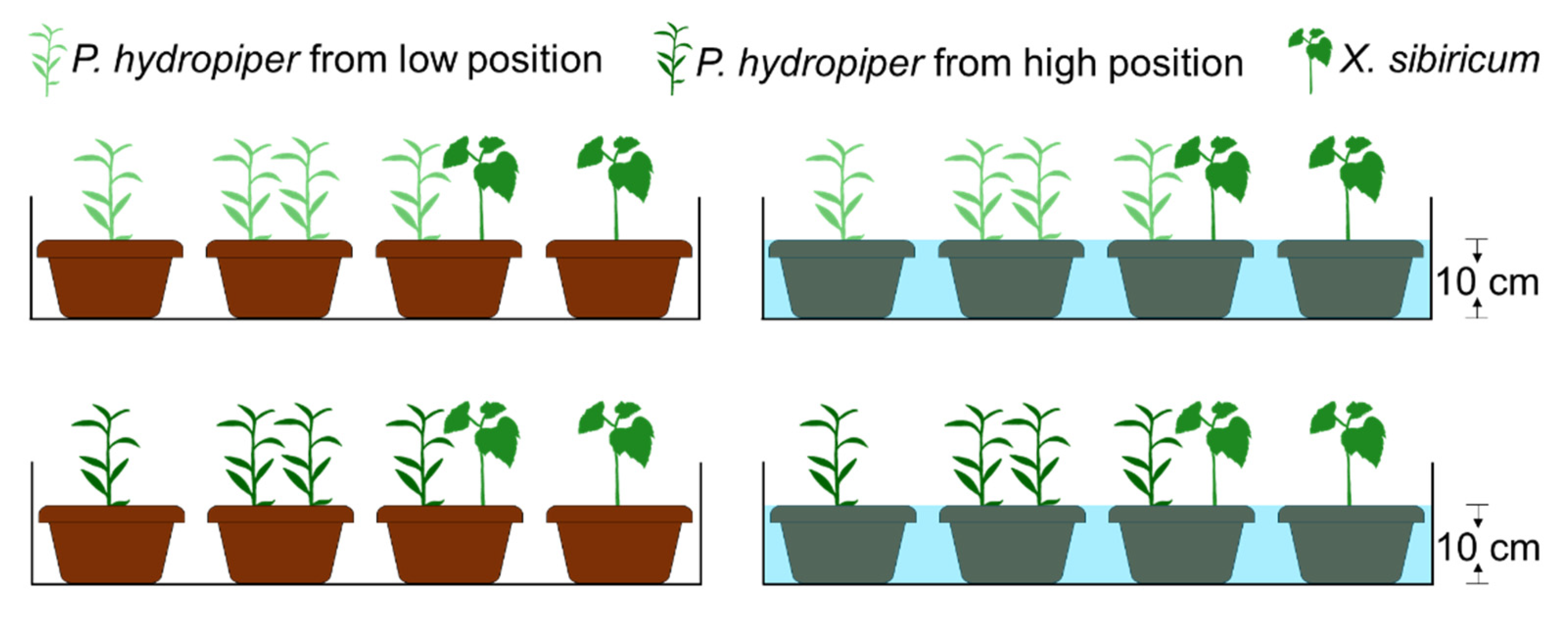
2.4. Experimental Design
2.5. Total Chlorophyll Content and Chlorophyll a Fluorescence Measurements
2.6. Growth Measurements
2.7. Statistical Analyses
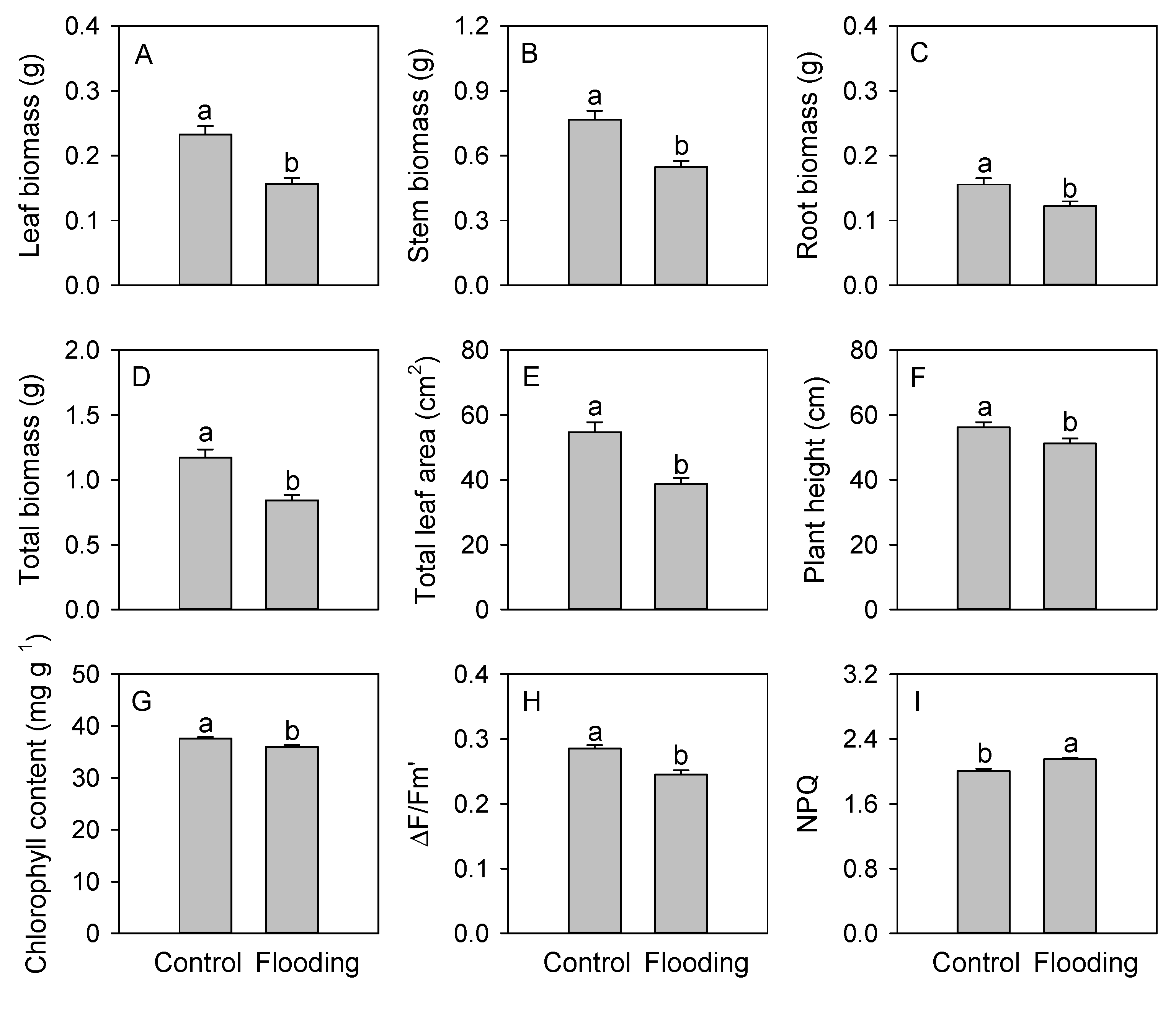
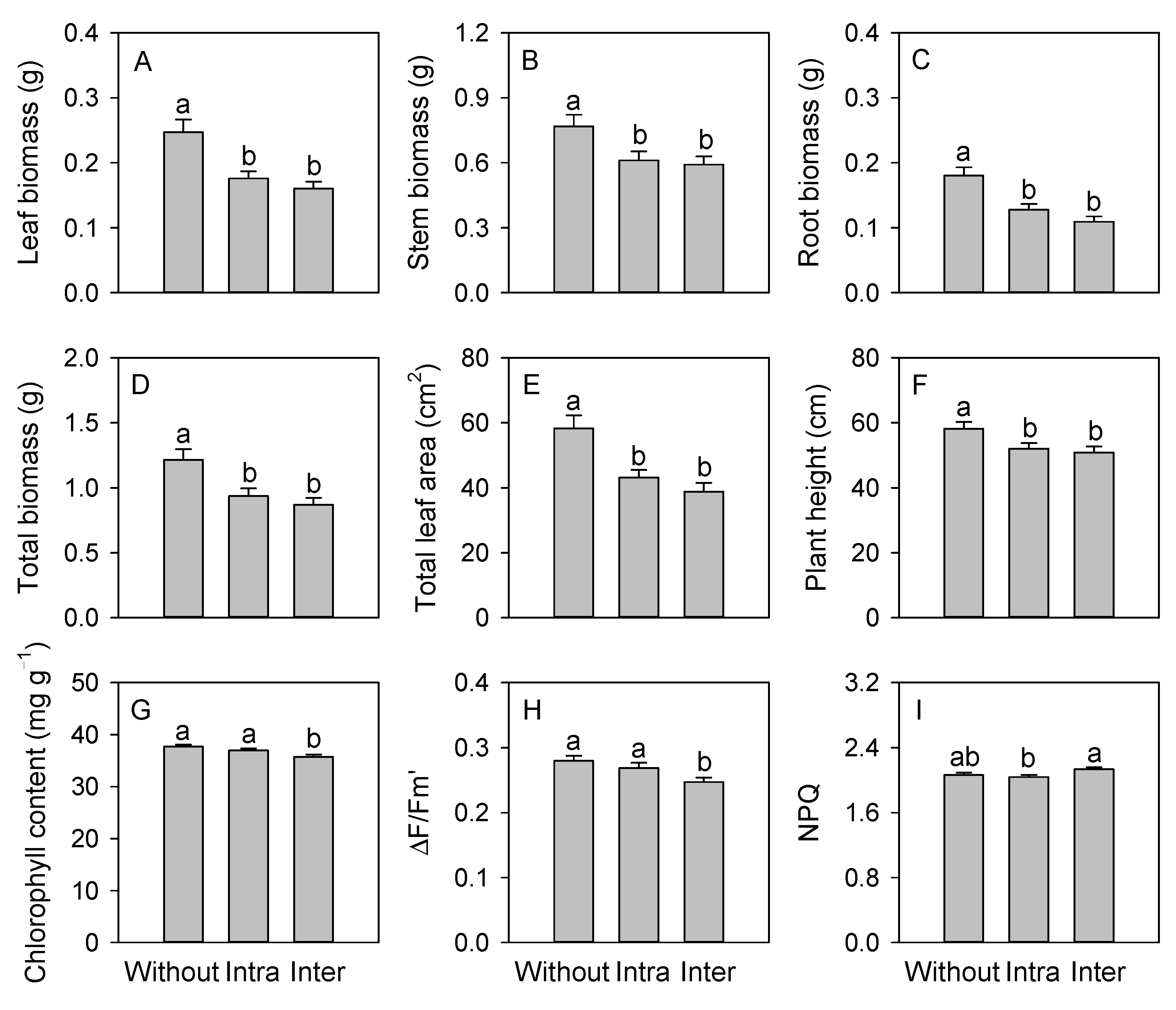
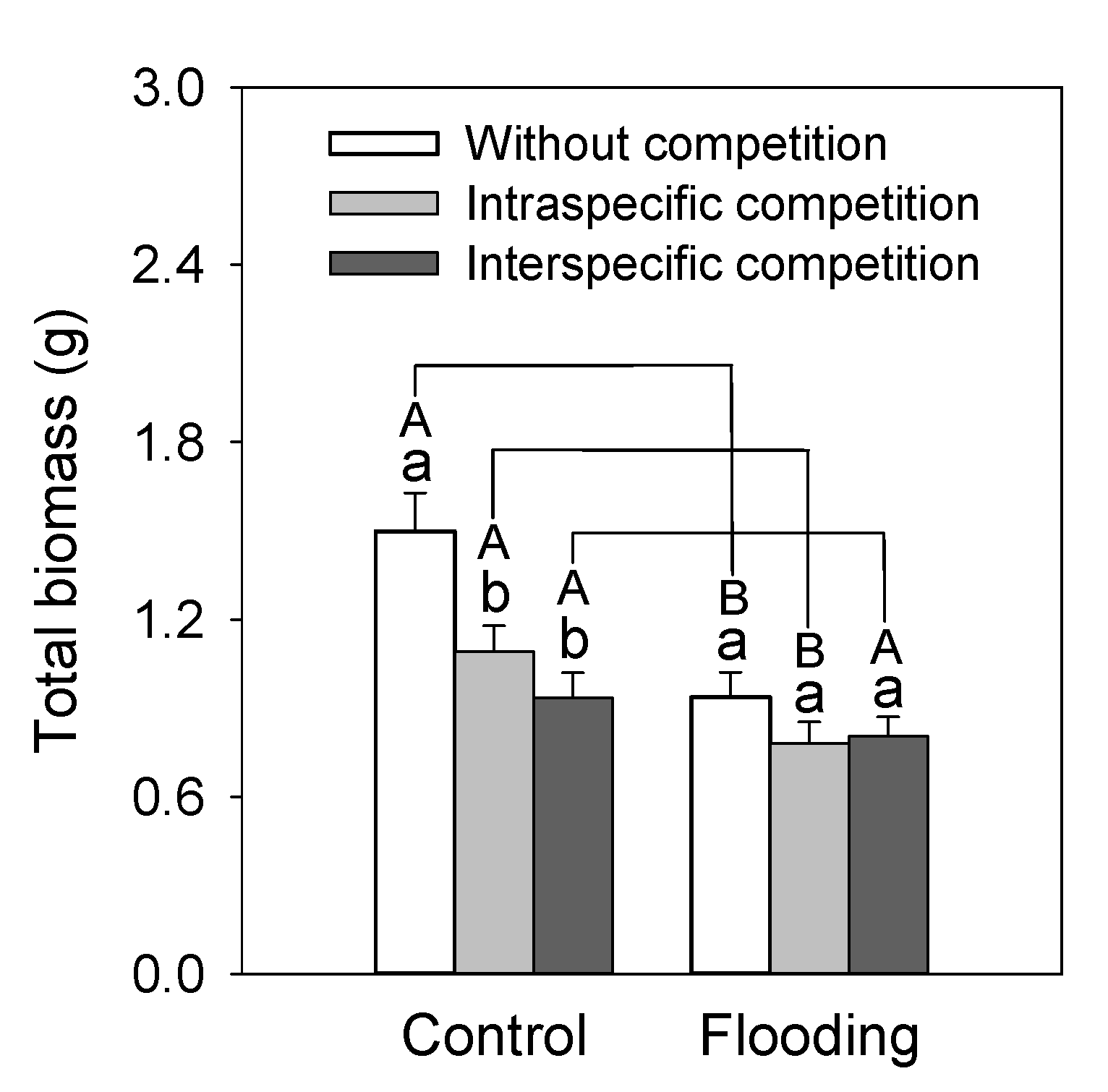
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Loreti, E.; van Veen, H.; Perata, P. Plant responses to flooding stress. Curr. Opin. Plant Biol. 2016, 33, 64–71. [Google Scholar] [CrossRef] [Green Version]
- Voesenek, L.A.C.J.; Bailey-Serres, J. Flood adaptive traits and processes: An overview. New Phytol. 2015, 206, 57–73. [Google Scholar] [CrossRef]
- Winkel, A.; Visser, E.J.W.; Colmer, T.D.; Brodersen, K.P.; Voesenek, L.A.C.J.; Sand-Jensen, K.; Pedersen, O. Leaf gas films, underwater photosynthesis and plant species distributions in a flood gradient. Plant Cell Environ. 2016, 39, 1537–1548. [Google Scholar] [CrossRef]
- Liu, M.Z.; Liu, Y.Y.; Zeng, B.; Niu, H.G.; Liu, S.P.; Pan, X.J.; Xie, J.J.; Shi, S.H.; Lin, F. Lowest elevation of plant growth and soil characteristics in natural drawdown areas in the Jiangjin section of the Yangtze River. Wetlands 2018, 39, 381–391. [Google Scholar] [CrossRef]
- Zhang, Z.Y.; Wan, C.Y.; Zheng, Z.W.; Hu, L.; Feng, K.; Chang, J.B.; Xie, P. Plant community characteristics and their responses to environmental factors in the water level fluctuation zone of the three gorges reservoir in China. Environ. Sci. Pollut. Res. 2013, 20, 7080–7091. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, J.; Taylor, J.P., Jr.; Smith, L.M.; Haukos, D.A. Seedling competition between native cottonwood and exotic saltcedar: Implications for restoration. Biol. Invasions 2009, 11, 1777–1787. [Google Scholar] [CrossRef] [Green Version]
- Stromberg, J.C.; Beauchamp, V.B.; Dixon, M.D.; Lite, S.J.; Paradzick, C. Importance of low-flow and high-flow characteristics to restoration of riparian vegetation along rivers in arid south-western United States. Freshw. Biol. 2007, 52, 651–679. [Google Scholar] [CrossRef]
- Davis, C.A.; Bidwell, J.R.; Hickman, K.R. Effects of hydrological regimes on competitive interactions of Schoenoplectus fluviatilis and two co-occurring wetland plants. Aquat. Bot. 2009, 91, 267–272. [Google Scholar] [CrossRef]
- Moore, J.E.; Franklin, S.B. Understanding the relative roles of disturbance and species interactions in shaping Mississippi River island plant communities. Community Ecol. 2011, 12, 108–116. [Google Scholar] [CrossRef]
- Bertness, M.D.; Callaway, R. Positive interactions in communities. Trends Ecol. Evol. 1994, 9, 191–193. [Google Scholar] [CrossRef]
- He, Q.; Bertness, M.D.; Altieri, A.H. Global shifts towards positive species interactions with increasing environmental stress. Ecol. Lett. 2013, 16, 695–706. [Google Scholar] [CrossRef] [PubMed]
- Maestre, F.T.; Callaway, R.M.; Valladares, F.; Lortie, C.J. Refining the stress-gradient hypothesis for competition and facilitation in plant communities. J. Ecol. 2009, 97, 199–205. [Google Scholar] [CrossRef]
- Hawkins, T.S.; Schiff, N.M.; Leininger, T.D.; Gardiner, E.S.; Devall, M.S.; Hamel, P.B.; Wilson, A.D.; Connor, K.F. Growth and intraspecific competitive abilities of the dioecious Lindera melissifolia (Lauraceae) in varied flooding regimes. J. Torrey Bot. Soc. 2009, 136, 90–101. [Google Scholar] [CrossRef]
- Moore, J.E.; Franklin, S.B. Water stress interacts with early arrival to influence interspecific and intraspecific priority competition: A test using a greenhouse study. J. Veg. Sci. 2012, 23, 647–656. [Google Scholar] [CrossRef]
- Puy, J.; Carmona, C.P.; Dvoáková, H.; Latzel, V.; de Bello, F. Diversity of parental environments increases phenotypic variation in Arabidopsis populations more than genetic diversity but similarly affects productivity. Ann. Bot. 2021, 127, 425–436. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.Y.; Latzel, V.; Fischer, M.; Bossdorf, O. Understanding the evolutionary potential of epigenetic variation: A comparison of heritable phenotypic variation in epiRILs, RILs, and natural ecotypes of Arabidopsis thaliana. Heredity 2018, 121, 257–265. [Google Scholar] [CrossRef]
- Albert, C.H. Intraspecific trait variability matters. J. Veg. Sci. 2015, 26, 7–8. [Google Scholar] [CrossRef]
- Messier, J.; McGill, B.J.; Lechowicz, M.J. How do traits vary across ecological scales? A case for trait-based ecology. Ecol. Lett. 2010, 13, 838–848. [Google Scholar] [CrossRef]
- Siefert, A.; Violle, C.; Chalmandrier, L.; Albert, C.H.; Taudiere, A.; Fajardo, A.; Aarssen, L.W.; Baraloto, C.; Carlucci, M.B.; Cianciaruso, M.V.; et al. A global meta-analysis of the relative extent of intraspecific trait variation in plant communities. Ecol. Lett. 2015, 18, 1406–1419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, X.L.; Zeng, B.; Huang, W.J.; Xu, S.J.; Lei, S.T. Effects of the Three Gorges Dam on preupland and preriparian drawdown zones vegetation in the upper watershed of the Yangtze River, P.R. China. Ecol. Eng. 2012, 44, 123–127. [Google Scholar] [CrossRef]
- Wu, J.G.; Huang, J.H.; Han, X.G.; Gao, X.M.; He, F.L.; Jiang, M.X.; Jiang, Z.G.; Primack, R.B.; Shen, Z.H. The Three Gorges Dam: An ecological perspective. Front. Ecol. Environ. 2004, 2, 241–248. [Google Scholar] [CrossRef]
- Fan, D.Y.; Xiong, G.M.; Zhang, A.Y.; Liu, X.; Xie, Z.Q.; Li, Z.J. Effect of water-lever regulation on species selection for ecological restoration practice in the water-level fluctuation zone of Three Gorges Reservoir. Chin. J. Plant Ecol. 2015, 39, 416–432. [Google Scholar]
- Yuan, S.H.; Zeng, B.; Su, X.L.; Xu, J.P. Effect of water-level fluctuation discrepancy on the composition of different annuals in Three Gorges Reservoir drawdown zone. Acta Econ. Sin. 2014, 34, 6481–6488. [Google Scholar]
- Ye, C.; Zhang, K.R.; Deng, Q.; Zhang, Q.F. Plant communities in relation to flooding and soil characteristics in the water level fluctuation zone of the Three Gorges Reservoir, China. Environ. Sci. Pollut. Res. 2013, 20, 1794–1802. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.Q.; Li, Y.; Qie, G.W.; Xu, W.N. The morphological responses and endurance of Polygonum hydropiper to flooding stress. J. Wuhan Bot. Res. 2008, 2, 142–146. [Google Scholar]
- Lan, Z.C.; Chen, Y.S.; Li, L.; Li, F.; Jin, B.S.; Chen, J.K. Testing mechanisms underlying elevational patterns of lakeshore plant community assembly in Poyang Lake, China. J. Plant Ecol. 2018, 12, 438–447. [Google Scholar] [CrossRef]
- Intergovermental Panel on Climate Change (IPCC). Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part A: Global and Sectoral Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Field, C.B., Barros, V.R., DOkken, D.J., Mach, K.J., Mastrandrea, M.D., Bilir, T.E., Chatterjhee, M., Ebi, K.L., Estrada, Y.O., Genova, R.C., et al., Eds.; Cambridge University Press: Cambridge, UK, 2014; p. 1132. [Google Scholar]
- Malkinson, D.; Tielbörger, K. What does the stress-gradient hypothesis predict? Resolving the discrepancies. Oikos 2010, 119, 1546–1552. [Google Scholar] [CrossRef]
- Pellissier, L.; Bråthen, K.A.; Pottier, J.; Randin, C.F.; Vittoz, P.; Dubuis, A.; Yoccoz, N.G.; Alm, T.; Zimmermann, N.E.; Guisan, A. Species distribution models reveal apparent competitive and facilitative effects of a dominant species on the distribution of tundra plants. Ecography 2010, 33, 1004–1014. [Google Scholar] [CrossRef]
- Gross, N.; Liancourt, P.; Choler, P.; Suding, K.N.; Lavorel, S. Strain and vegetation effects on local limiting resources explain the outcomes of biotic interactions. Perspect. Plant Ecol. 2010, 12, 9–19. [Google Scholar] [CrossRef]
- Soliveres, S.; Eldridge, D.J.; Maestre, F.T.; Bowker, M.A.; Tighe, M.; Escudero, A. Microhabitat amelioration and reduced competition among understorey plants as drivers of facilitation across environmental gradients: Towards a unifying framework. Perspect. Plant Ecol. 2011, 13, 247–258. [Google Scholar] [CrossRef] [Green Version]
- Li, A.R. Flora of China; Science Press: Beijing, China, 1998; Volume 25. [Google Scholar]
- Chen, X.S.; Li, Y.F.; Cai, Y.H.; Xie, Y.H.; Deng, Z.M.; Li, F.; Hou, Z.Y. Differential strategies to tolerate flooding in Polygonum hydropiper plants originating from low- and high-elevation habitats. Front. Plant Sci. 2019, 9, 1970. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D. Nlme: Linear and Nonlinear Mixed Effects Models. R package version 3.1-152. Available online: https://CRAN.R-project.org/package=nlme (accessed on 20 February 2021).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Adler, P.B.; Smull, D.; Beard, K.H.; Choi, R.T.; Furniss, T.; Kulmatiski, A.; Meiners, J.M.; Tredennick, A.T.; Veblen, K.E. Competition and coexistence in plant communities: Intraspecific competition is stronger than interspecific competition. Ecol. Lett. 2018, 21, 1319–1329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kula, A.A.R.; Hey, M.H.; Couture, J.J.; Townsend, P.A.; Dalgleish, H.J. Intraspecific competition reduces plant size and quality and damage severity increases defense responses in the herbaceous perennial, Asclepias syriaca. Plant Ecol. 2020, 221, 421–430. [Google Scholar] [CrossRef]
- Zhou, J.; Zheng, L.D.; Pan, X.; Li, W.; Kang, X.M.; Li, J.; Ning, Y.; Zhang, M.X.; Cui, L.J. Hydrological conditions affect the interspecific interaction between two emergent wetland species. Front. Plant Sci. 2018, 8, 2253. [Google Scholar] [CrossRef] [Green Version]
- Muench, A.; Elsey-Quirk, T. Competitive reversal between plant species is driven by species-specific tolerance to flooding stress and nutrient acquisition during early marsh succession. J. Appl. Ecol. 2019, 56, 2236–2247. [Google Scholar] [CrossRef]
- Yue, M.F.; Shen, H.; Li, W.H.; Chen, J.F.; Ye, W.H.; Tian, X.S.; Yin, A.G.; Cheng, S.M. Waterlogging tolerance of Bidens pilosa translates to increased competitiveness compared to native Bidens biternata. Plant Soil 2019, 437, 301–311. [Google Scholar] [CrossRef]
- Lysenko, E.A.; Klaus, A.A.; Pshybytko, N.L.; Kusnetsov, V.V. Cadmium accumulation in chloroplasts and its impact on chloroplastic processes in barley and maize. Photosynth. Res. 2015, 125, 291–303. [Google Scholar] [CrossRef]
- García-Cervigón, A.I.; Gazol, A.; Sanz, V.; Camarero, J.J.; Olano, J.M. Intraspecific competition replaces interspecific facilitation as abiotic stress decreases: The shifting nature of plant-plant interactions. Perspect. Plant Ecol. 2013, 15, 226–236. [Google Scholar] [CrossRef]
- Qiu, S.Y.; Liu, S.S.; Wei, S.J.; Cui, X.H.; Nie, M.; Huang, J.X.; He, Q.; Ju, R.T.; Li, B. Changes in multiple environmental factors additively enhance the dominance of an exotic plant with a novel trade-off pattern. J. Ecol. 2020, 108, 1989–1999. [Google Scholar] [CrossRef]
- Ye, C.; Butler, O.M.; Chen, C.; Liu, W.; Du, M.; Zhang, Q. Shifts in characteristics of the plant-soil system associated with flooding and revegetation in the riparian zone of Three Gorges Reservoir, China. Geoderma 2020, 361, 114015. [Google Scholar] [CrossRef]
- Wei, G.W.; Chen, Y.H.; Sun, X.S.; Matsubara, S.; Luo, F.L.; Yu, F.H. Elevation-dependent selection for plasticity in leaf and root traits of Polygonum hydropiper in response to flooding. Environ. Exp. Bot. 2021, 182, 104331. [Google Scholar] [CrossRef]
- Wei, G.W.; Sun, X.S.; Chen, Y.H.; Luo, F.L.; Yu, F.H. Growth and reproductive responses of Polygonum hydropiper populations to elevational difference associated with flooding. Glob. Ecol. Conserv. 2020, 23, e01156. [Google Scholar] [CrossRef]
- Harpole, W.S.; Sullivan, L.L.; Lind, E.M.; Firn, J.; Adler, P.B.; Borer, E.T.; Chase, J.; Fay, P.A.; Hautier, Y.; Hillebrand, H.; et al. Addition of multiple limiting resources reduces grassland diversity. Nature 2016, 537, 93–96. [Google Scholar] [CrossRef]
- Li, Y.; Kang, J.; Li, Z.; Korpelainen, H.; Li, C. Ecophysiological responses of two poplar species to intraspecific and interspecific competition under different nitrogen levels. J. Plant Ecol. 2020, 13, 693–703. [Google Scholar] [CrossRef]
- Pedersen, O.; Perata, P.; Voesenek, L.A.C.J. Flooding and low oxygen responses in plants. Funct. Plant Biol. 2017, 44, iii–vi. [Google Scholar] [CrossRef]
- Yamauchi, T.; Colmer, T.D.; Pedersen, O.; Nakazono, M. Regulation of root traits for internal aeration and tolerance to soil waterlogging-flooding stress. Plant Physiol. 2017, 176, 1118–1130. [Google Scholar] [CrossRef] [Green Version]
- Polacik, K.A.; Maricle, B.R. Effects of flooding on photosynthesis and root respiration in saltcedar (Tamarix ramosissima), an invasive riparian shrub. Environ. Exp. Bot. 2013, 89, 19–27. [Google Scholar] [CrossRef] [Green Version]
- van Veen, H.; Sasidharan, R. Shape shifting by amphibious plants in dynamic hydrological niches. New Phytol. 2019, 229, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Sher, A.A.; Marshall, D.L. Seedling competition between native Populus deltoides (Salicaceae) and exotic Tamarix ramosissima (Tamaricaceae) across water regimes and substrate types. Am. J. Bot. 2003, 90, 413–422. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site (S) | Flooding (F) | Competition (C) | Position (P) | F × C | F × P | C × P | F × C × P | |
|---|---|---|---|---|---|---|---|---|
| Leaf biomass | 24.09 *** | 26.82 *** | 9.59 *** | 1.58 | 2.18 | 0.04 | 1.69 | 0.64 |
| Stem biomass | 24.21 *** | 16.25 *** | 4.33 * | 0.89 | 2.35 | 0.05 | 1.90 | 0.49 |
| Root biomass | 24.11 *** | 6.45 * | 14.19 *** | 0.30 | 0.58 | 0.21 | 1.52 | 0.42 |
| Total biomass | 21.99 *** | 23.39 *** | 10.24 *** | 0.44 | 3.27 * | 0.10 | 1.20 | 0.85 |
| Total leaf area | 18.68 *** | 17.61 *** | 11.01 *** | 0.52 | 2.12 | 0.18 | 2.59 | 0.78 |
| SLA | 1.45 | 0.09 | 0.53 | 3.29 | 1.53 | 0.18 | 2.11 | 2.34 |
| Plant height | 47.98 *** | 6.72 * | 6.72 ** | 1.80 | 2.39 | 0.36 | 1.22 | 0.70 |
| Chl content | 1.69 | 12.58 *** | 6.48 ** | 11.79 *** | 2.30 | 2.35 | 0.78 | 0.06 |
| Fv/Fm | 16.44 *** | 0.48 | 1.12 | 0.33 | 0.89 | 1.37 | 1.40 | 0.54 |
| ΔF/Fm′ | 9.17 ** | 26.41 *** | 5.98 ** | 1.61 | 0.29 | 1.57 | 0.41 | 0.69 |
| NPQ | <0.01 | 23.06 *** | 3.48 * | 0.71 | 0.87 | 0.86 | 0.60 | 0.34 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.-H.; Sun, X.-S.; Cui, Y.; Zhuo, N.; Wei, G.-W.; Luo, F.-L.; Zhang, M.-X. Interacting Flooding and Competition Negatively Affect Growth of Riparian Species Dominating a Reservoir Shoreline. Water 2021, 13, 1471. https://doi.org/10.3390/w13111471
Chen Y-H, Sun X-S, Cui Y, Zhuo N, Wei G-W, Luo F-L, Zhang M-X. Interacting Flooding and Competition Negatively Affect Growth of Riparian Species Dominating a Reservoir Shoreline. Water. 2021; 13(11):1471. https://doi.org/10.3390/w13111471
Chicago/Turabian StyleChen, Yu-Han, Xin-Sheng Sun, Yuan Cui, Na Zhuo, Guan-Wen Wei, Fang-Li Luo, and Ming-Xiang Zhang. 2021. "Interacting Flooding and Competition Negatively Affect Growth of Riparian Species Dominating a Reservoir Shoreline" Water 13, no. 11: 1471. https://doi.org/10.3390/w13111471