
Omnivorous Carp (Carassius gibelio) Increase Eutrophication in Part by Preventing Development of Large-Bodied Zooplankton and Submerged Macrophytes

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Mesocosm Setup
2.2. Sampling and Analysis
2.3. Statistical Analysis
3. Results
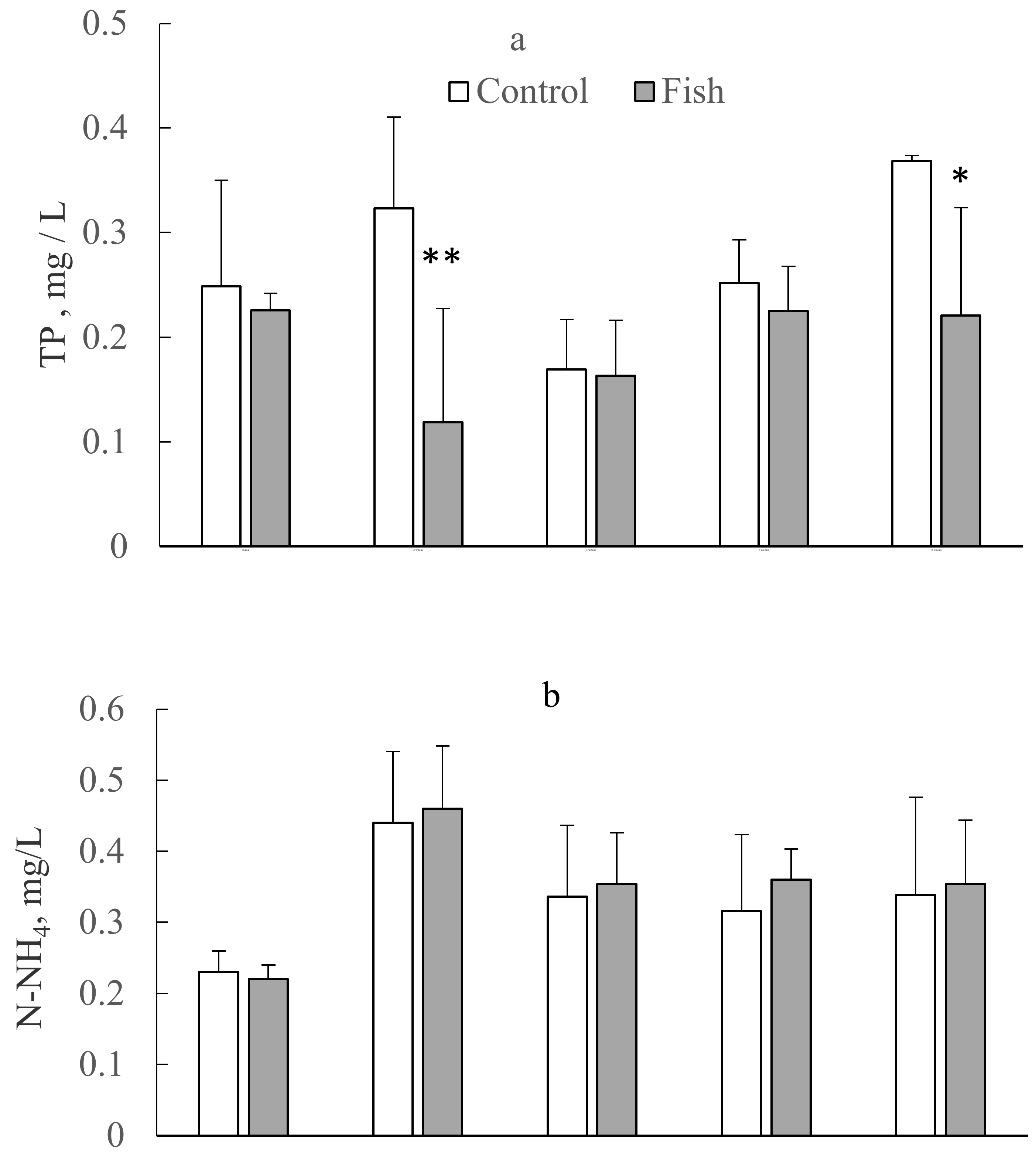
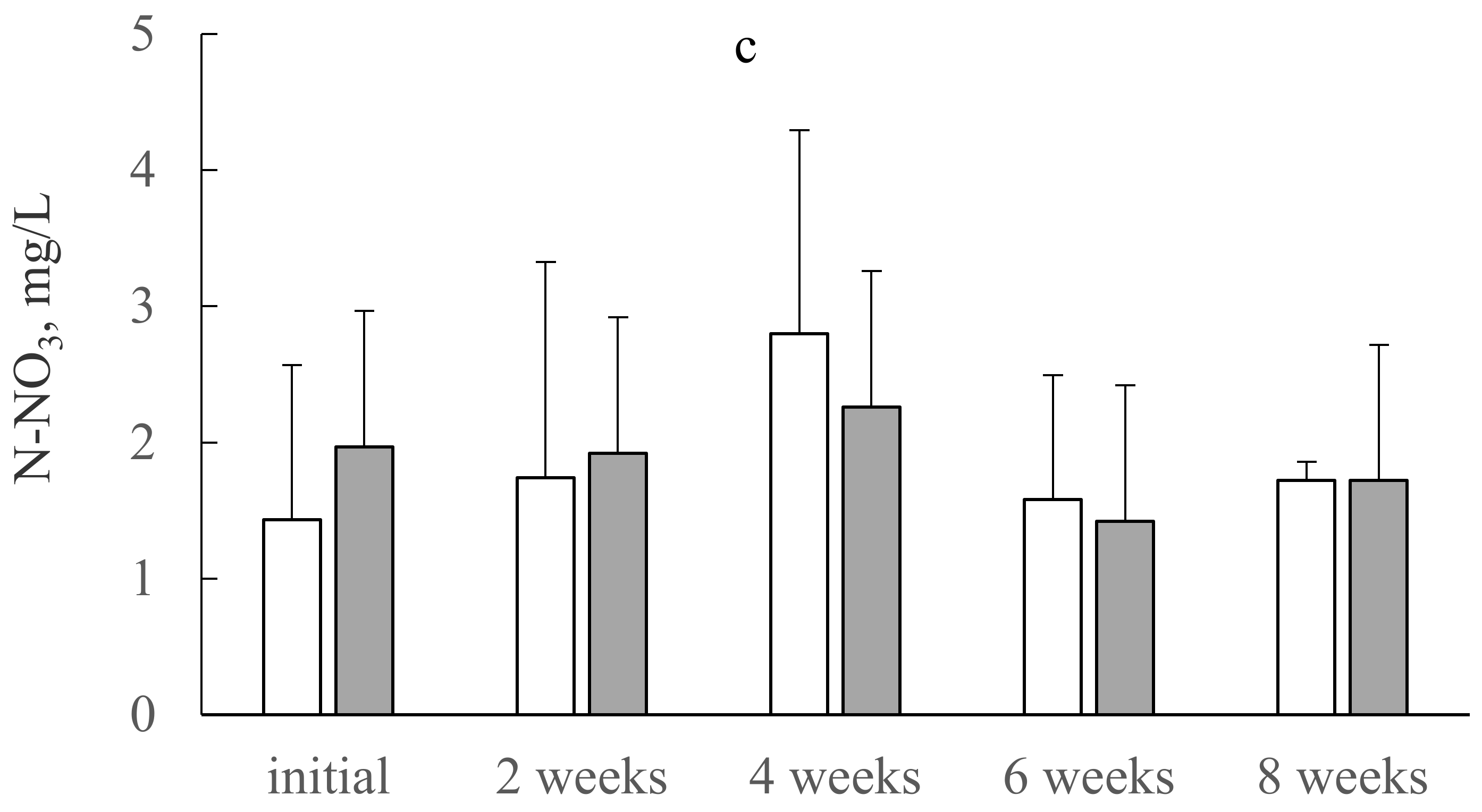
3.1. N and P
3.2. TSS, ISS and Light Intensity
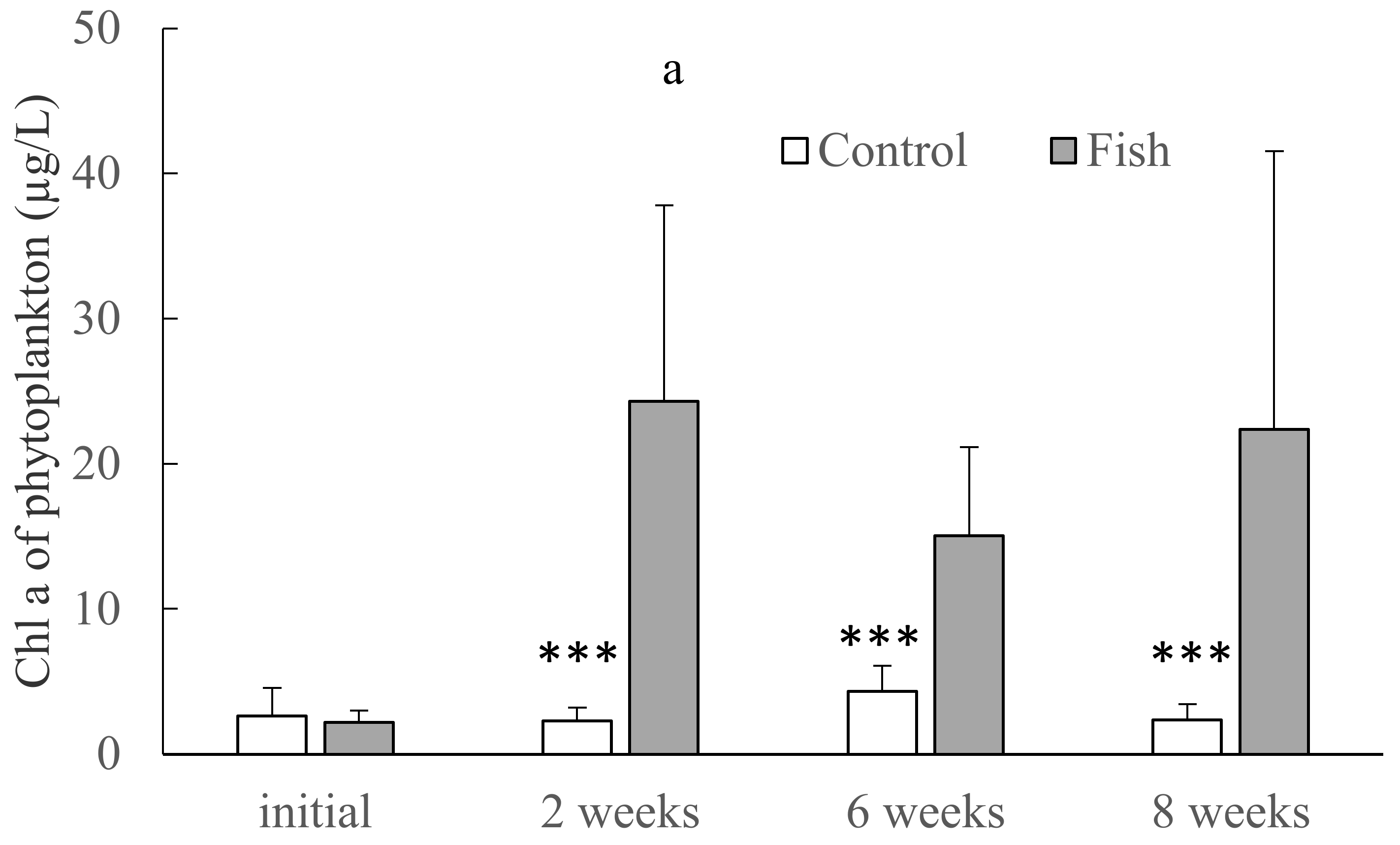
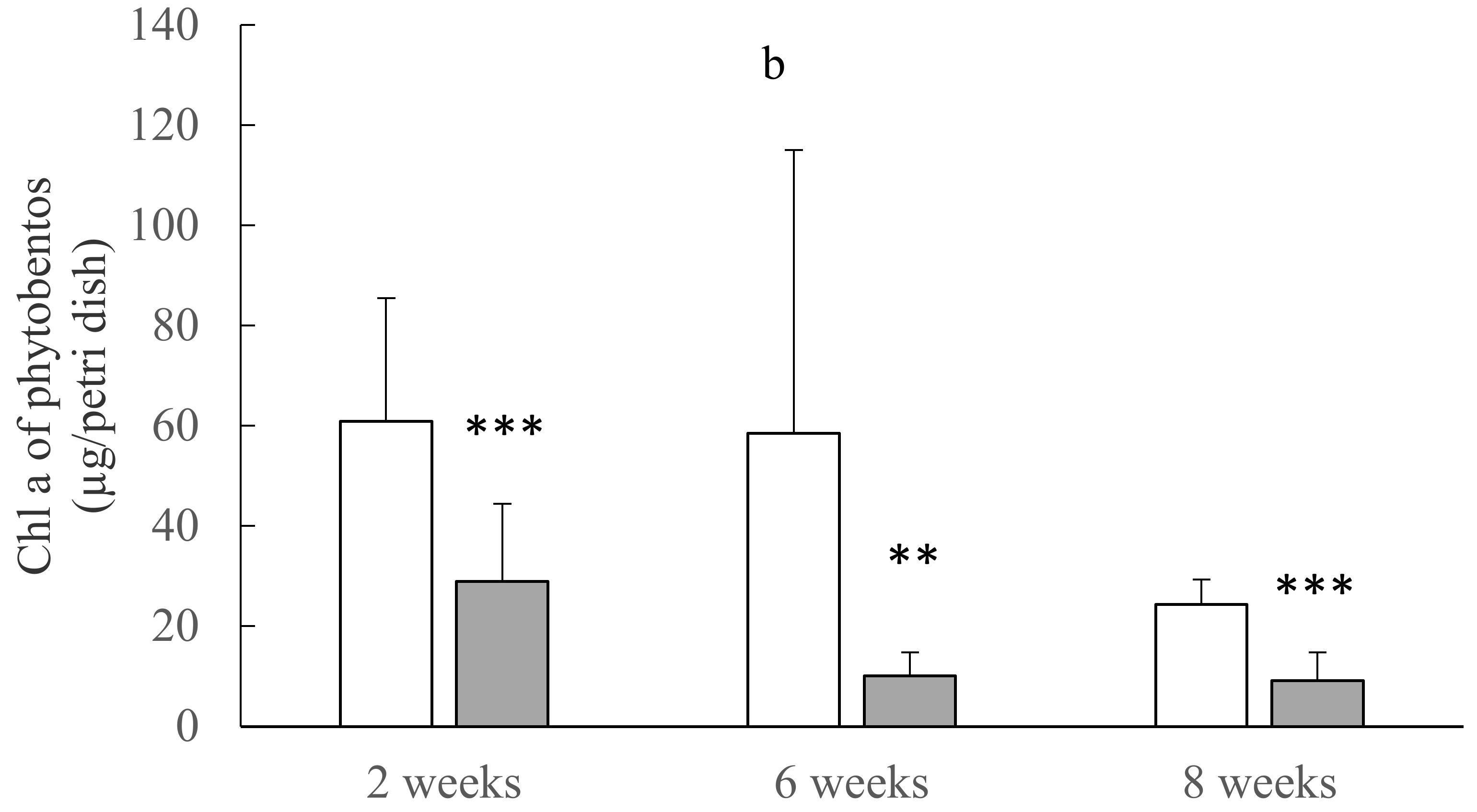
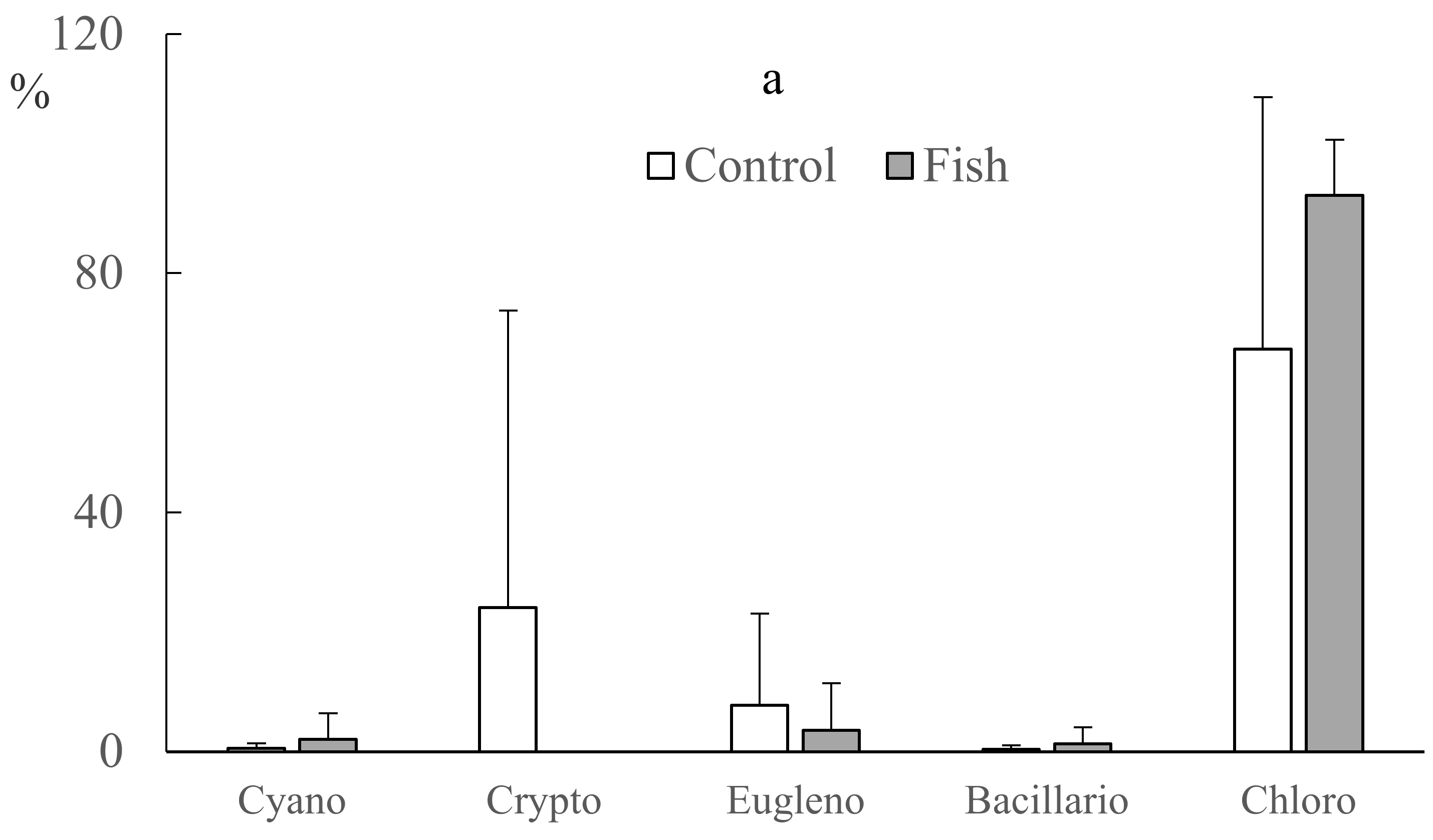
3.3. Biomass (Chl a Concentrations) and Compositions of Planktonic and Benthic Algae
3.4. Submerged Charophytes
3.5. Density and Composition of Zooplankton
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schindler, D.W.; Hecky, R.E.; Findlay, D.L.; Stainton, M.P.; Parker, B.R.; Paterson, M.J.; Beaty, K.G.; Lyng, M.; Kasian, S.E.M. Eutrophication of lakes cannot be controlled by reducing nitrogen input: Results of a 37-year whole-ecosystem experiment. Proc. Natl. Acad. Sci. USA 2008, 105, 11254–11258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, V.H.; Schindler, D.W. Eutrophication science: Where do we go from here? Trends Ecol. Evol. 2009, 24, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Schindler, D.W. Lakes as sentinels and integrators for the effects of climate change on watersheds, airsheds, and landscapes. Limnol. Oceanogr. 2009, 54, 2349–2358. [Google Scholar] [CrossRef]
- Hansson, L.A. Effects of competitive interactions on the biomass development of planktonic and periphytic algae in lakes. Limnol. Oceanogr. 1998, 33, 121–128. [Google Scholar] [CrossRef]
- Vadeboncoeur, Y.; Jeppesen, E.; Zanden, M.J.V.; Schierup, H.H.; Christoffersen, K.; Lodge, D.M. From Greenland to green lakes: Cultural eutrophication and the loss of benthic pathways in lakes. Limnol. Oceanogr. 2003, 48, 1408–1418. [Google Scholar] [CrossRef] [Green Version]
- Cantonati, M.; Lowe, R.L. Lake benthic algae: Toward an understanding of their ecology. Freshw. Sci. 2014, 33, 475–486. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, Z.; Gulati, R.D.; Jeppesen, E. The effect of benthic algae on phosphorus exchange between sediment and overlying water in shallow lakes: A microcosm study using 32P as a tracer. Hydrobiologia 2013, 710, 109–116. [Google Scholar] [CrossRef]
- Zambrano, L.; Scheffer, M.; Martínez-Ramos, M. Catastrophic response of lakes to benthivorous fish introduction. Oikos 2001, 94, 344–350. [Google Scholar] [CrossRef] [Green Version]
- Andersson, G.; Berggren, H.; Cronberg, G.; Gelin, C. Effects of planktivorous and benthivorous fish on organisms and water chemistry in eutrophic lakes. Hydrobiologia 1978, 59, 9–15. [Google Scholar] [CrossRef]
- Eggers, D.M.; Bartoo, N.W.; Rickard, N.A.; Nelson, R.E.; Wissmar, R.C.; Burgner, R.L.; Devol, A.H. The Lake Washington ecosystem: The perspective from the fish community production and forage base. J. Fish. Res. Board Can. 1978, 35, 1553–1571. [Google Scholar] [CrossRef] [Green Version]
- Jeppesen, E.; Jensen, J.P.; Jensen, C.; Faafeng, B.; Hessen, D.O.; Søndergaard, M.; Lauridsen, T.; Brettum, P.; Christoffersen, K. The impact of nutrient state and lake depth on top-down control in the pelagic zone of lakes: A study of 466 lakes from the temperate zone to the arctic. Ecosystems 2003, 6, 313–325. [Google Scholar] [CrossRef]
- Attayde, J.L.; van Nes, E.H.; Araujo, A.I.; Corso, G.; Scheffer, M. Omnivory by planktivores stabilizes plankton dynamics, but may either promote or reduce algal biomass. Ecosystems 2010, 13, 410–420. [Google Scholar] [CrossRef] [Green Version]
- Mao, Z.; Gu, X.; Cao, Y.; Zhang, M.; Zeng, Q.; Chen, H.; Shen, R.; Jeppesen, E. The role of top-down and bottom-up control for phytoplankton in a subtropical shallow eutrophic lake: Evidence based on long-term monitoring and modelling. Ecosystems 2020, 23, 1449–1463. [Google Scholar] [CrossRef]
- Zhang, X.F.; Liu, Z.W.; Jeppesen, E.; Taylor, W.D.; Rudstam, L.G. Effects of benthic-feeding common carp and filterfeeding silver carp on benthic-pelagic coupling: Implications for shallow lake management. Ecol. Eng. 2016, 88, 256–264. [Google Scholar] [CrossRef]
- Skeate, E.R.; Perrow, M.R.; Tomlinson, M.L.; Madgwick, G.; Harwood, A.J.P.; Ottewell, D.; Berridge, R.; Winfield, I.J. Fish stocking for recreational angling is culpable for the poor condition of many English lakes designated for conservation purposes. Inland Waters 2021, 1–14. [Google Scholar] [CrossRef]
- Carpenter, S.R.; Kitchell, J.F.; Hodgson, J.R.; Cochran, P.A.; Elser, J.J.; Elser, M.M.; Lodge, D.M.; Kretchmer, D.; He, X.; von Ende, C.N. Regulation of lake primary productivity by food web structure. Ecology 1987, 68, 1863–1876. [Google Scholar] [CrossRef] [Green Version]
- Vanni, M.J. Nutrient cycling by animals in freshwater ecosystems. Annu. Rev. Ecol. Syst. 2002, 33, 341–370. [Google Scholar] [CrossRef] [Green Version]
- Vanni, M.J. Nutrient transport and recycling by consumers in lake food webs: Implications for algal communities. Food Webs 1996, 81–95. [Google Scholar] [CrossRef]
- Scheffer, M.; Portielje, R.; Zambrano, L. Fish facilitate wave resuspension of sediment. Limnol. Oceanogr. 2003, 48, 1920–1926. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Mei, X.; Gulati, R.D. Effects of omnivorous tilapia on water turbidity and primary production dynamics in shallow lakes: Implications for ecosystem management. Rev. Fish. Biol. Fish. 2017, 27, 245–254. [Google Scholar] [CrossRef]
- Breukelaar, A.W.; Lammens, E.H.R.R.; Breteler, J.G.P.K.; Tátrai, I. Effects of benthivorous bream (Abramis brama) and carp (Cyprinus carpio) on sediment resuspension and concentrations of nutrients and chlorophyll a. Freshw. Biol. 1994, 32, 113–121. [Google Scholar] [CrossRef]
- Stein, R.A.; DeVries, D.R.; Dettmers, J.M. Food-web regulation by a planktivore: Exploring the generality of the trophic cascade hypothesis. Can. J. Fish. Aquat. Sci. 1995, 52, 2518–2526. [Google Scholar] [CrossRef] [Green Version]
- Dettmers, J.M.; Stein, R.A. Quantifying linkages among gizzard shad, zooplankton, and phytoplankton in reservoirs. Transac. Am. Fish. Soc. 1995, 125, 27–41. [Google Scholar] [CrossRef] [Green Version]
- Jeppesen, E.; Jensen, J.P.; Søndergaard, M.; Lauridsen, T. Trophic dynamics in turbid and clearwater lakes with special emphasis on the role of zooplankton for water clarity. Hydrobiologia 1999, 408, 217–231. [Google Scholar] [CrossRef]
- Richardson, M.J.; Whoriskey, F.G.; Roy, H. Turbidity generation and biological impacts of an exotic Carassius auratus, introduced into shallow seasonally anoxic pounds. J. Fish Biol. 1995, 47, 576–585. [Google Scholar]
- Qiu, X.; Mei, X.; Razlutskij, V.; Rudstam, L.G.; Liu, Z.; Tong, C.; Zhang, X. Effects of common carp (Cyprinus carpio) on water quality in aquatic ecosystems dominated by submerged plants: A mesocosm study. Knowl. Manag. Aquat. Ecosyst. 2019, 420, 28. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Mei, X.; Rudstam, L.G.; Taylor, W.D.; Urabe, J.; Jeppesen, E.; Liu, Z.; Zhang, X. Effects of crucian carp (Carassius auratus) on water quality in aquatic ecosystems: An experimental mesocosm study. Water 2020, 12, 14–44. [Google Scholar] [CrossRef]
- Badiou, P.H.J.; Goldsborough, L.G. Ecological impacts of an exotic benthivorous fish, the common carp (Cyprinus carpio L.), on water quality, sedimentation, and submerged macrophyte biomass in wetland mesocosms. Hydrobiologia 2015, 755, 107–121. [Google Scholar] [CrossRef]
- Gabaldóna, C.; Buseva, Z.; Illyovác, M.; Seda, J. Littoral vegetation improves the productivity of drainable fish ponds: Interactive effects of refuge for Daphnia individuals and resting eggs. Aquaculture 2018, 485, 111–118. [Google Scholar] [CrossRef]
- Roberts, J.; Chick, A.; Oswald, L.; Thompson, P. Effect of carp, Cyprinus carpio L., an exotic benthivorous fish, on aquatic plants and water quality in experimental ponds. Mar. Freshw. Res. 1995, 46, 1171–1180. [Google Scholar] [CrossRef]
- Stephen, D.; Balayla, D.M.; Bécares, E.; Collings, S.E.; Fernández-Aaláez, C.; Fernández-Aláez, M.; Ferriol, C.; García, P.; Gomá, J.; Gyllström, M.; et al. Continental-scale patterns of nutrient and fish effects on shallow lakes: Introduction to a pan-European mesocosm experiment. Freshw. Biol. 2004, 49, 1517–1524. [Google Scholar] [CrossRef] [Green Version]
- Nico, L.; Schofield, P.J. Carassius auratus; USGS Non-indigenous Aquatic Species Database: Gainesville, FL, USA, 2006.
- Abramenko, M.I.; Kravchenko, O.V.; Velikoivanenko, A.E. Population genetic structure of the goldfish Carassius auratus gibelio diploid-triploid complex from the Don River Basin. J. Ichthyol. 1997, 37, 56–65. [Google Scholar]
- Xie, J.; Wen, J.; Chen, B.; Gui, J.F. Differential gene expression in fully-grown oocytes between gynogenetic and gonochoristic crucian carp. Gene 2001, 272, 109–116. [Google Scholar] [CrossRef]
- Kuznetsov, V.A. Changes in the population structure and biological indices of the goldfish Carassius auratus gibelio in the Volga Stretch of the Kuibyshev Reservoir under conditions of intense anthropogenic load on the ecosystem. J. Ichthyol. 2004, 44, 167–174. [Google Scholar]
- Spotila, J.R.; Terpin, K.M.; Koons, R.R.; Bonati, R.L. Temperature requirements of fishes from eastern Lake Erie and upper Niagara River. Environ. Biol. Fishes 1979, 4, 281–307. [Google Scholar] [CrossRef]
- Pinder, A.C. Keys to the Freshwater Fish of Britain and Ireland, with Notes on Their Distribution and Ecology; Freshwater Biological Association: Ambleside, UK, 2003; 248p. [Google Scholar]
- Rylková, K.; Kalous, L.; Bohlen, J.; Lamatsch, D.K.; Petrtý, M. Phylogeny and biogeographic history of the cyprinid fish genus Carassius (Teleostei: Cyprinidae) with focus on natural and anthropogenic arrivals in Europe. Aquaculture 2013, 380–383, 13–20. [Google Scholar] [CrossRef]
- Grichik, V.V.; Burko, L.D. Animal World of Belarus. Vertebrate: A Tutorial; BSU: Minsk, Belarus, 2013; 399p. [Google Scholar]
- Semenchenko, V.P.; Rizevsky, V.K.; Mastitsky, S.E.; Vezhnovets, V.V.; Pluta, M.V.; Razlutsky, V.I.; Laenko, T. Checklist of aquatic alien species established in large river basins of Belarus. Aquat. Invas. 2009, 4, 311–320. [Google Scholar] [CrossRef]
- Barbour, M.T.; Gerritsen, J.; Snyder, B.D.; Stribling, J.B. Rapid Bioassessment Protocols for Use in Streams and Wadeable Rivers: Periphyton, Benthic Macroinvertebrates and Fish; USEPA: Washington, DC, USA, 1999; 337p.
- Jespersen, A.M.; Christoffersen, K. Measurements of chlorophyll a from phytoplankton using ethanol as extraction solvent. Arch. Hydrobiol. 1987, 109, 445–454. [Google Scholar]
- Kutikova, L.A. Rotatoria of the Fauna of the USSR; Leningrad: Leningrad, Russia, 1970; 744p. [Google Scholar]
- Błędzki, L.A.; Rybak, J.I. Cladocera Morphology. In Freshwater Crustacean Zooplankton of Europe; Springer: Berlin/Heidelberg, Germany, 2016; pp. 95–101. [Google Scholar]
- Kaya, M.; Erdoğan, S. Morphological examination of the resting egg structure of 3 cladoceran species [Ceriodaphnia quadrangula (O. F. Müller, 1785), Daphnia longispina (O. F. Müller, 1776), and D. magna Straus, 1820]. Turk. J. Zool. 2014, 38, 131–135. [Google Scholar] [CrossRef]
- Vandekerkhove, J.; Declerck, S.; Vanhove, M.; Brendonck, L.; Jeppesen, E.; Conde Porcuna, J.M.; de Meester, L. Use of ephippial morphology to assess richness of anomopods: Potentials and pitfalls. J. Limnol. 2004, 63, 131–135. [Google Scholar] [CrossRef] [Green Version]
- Vanni, M.J.; Boros, G.; Mcintyre, P.B. When are fish sources vs. sinks of nutrients in lake ecosystems? Ecology 2013, 94, 2195–2206. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.T.; Liu, F.; Wang, F. Carbon, nitrogen, and phosphorus stoichiometry of three freshwater cultured fishes in growth stage. Turk. J. Fish Aquat. Sci. 2018, 18, 239–245. [Google Scholar] [CrossRef]
- Rhee, G.Y. Competition between an alga and an aquatic bacterium for phosphate. Limnol. Oceanogr. 1972, 17, 505–514. [Google Scholar] [CrossRef]
- Grover, J.P. Resource competition and community structure in aquatic microorganisms: Experimental studies of algae and bacteria along a gradient of organic carbon to inorganic phosphorus supply. J. Plankton Res. 2000, 22, 1591–1610. [Google Scholar] [CrossRef] [Green Version]
- Danger, M.; Oumarou, C.; Benest, D.; Lacroix, G.G. Bacteria can control stoichiometry and nutrient limitation of phytoplankton. Funct. Ecol. 2007, 21, 202–210. [Google Scholar] [CrossRef]
- Schwaderer, A.; Yoshiyama, K.; de Tezanos Pinto, P.; Swenson, N.G.; Christopher, A.; Klausmeier, C.A.; Litchman, E. Eco-evolutionary differences in light utilization traits and distributions of freshwater phytoplankton. Limnol. Oceanogr. 2011, 56, 589–598. [Google Scholar] [CrossRef]
- Van Donk, E.; van de Bund, W.J. Impact of submerged macrophytes including charophytes on phyto-and zooplankton communities: Allelopathy versus other mechanisms. Aquat. Bot. 2002, 72, 261–274. [Google Scholar] [CrossRef]
- Rybak, A.S.; Gąbka, M. The influence of abiotic factors on the bloom-forming alga Ulva flexuosa (Ulvaceae, Chlorophyta): Possibilities for the control of the green tides in freshwater ecosystems. J. Appl. Phycol. 2018, 30, 1405–1416. [Google Scholar] [CrossRef] [Green Version]
- Erhard, D.; Gross, E.M. Allelopathic activity of Elodea canadensis and Elodea nuttallii against epiphytes and phytoplankton. Aquat. Bot. 2006, 85, 203–211. [Google Scholar] [CrossRef] [Green Version]
- Pakdel, F.M.; Sim, L.; Beardall, J.; Davis, J. Allelopathic inhibition of microalgae by the freshwater stonewort, Chara australis, and a submerged angiosperm, Potamogeton crispus. Aquat. Bot. 2013, 110, 24–30. [Google Scholar] [CrossRef]
- Ibelings, B.W.; Portielje, R.; Lammens, E.H.R.R.; Noordhuis, R.; van den Berg, M.; Willemien, J.; Meijer, M.L. Resilience of alternative stable states during the recovery of shallow lakes from eutrophication: Lake Veluwe as a case study. Ecosystems 2007, 10, 4–16. [Google Scholar] [CrossRef] [Green Version]
- De Winton, M.D.; Taumoepeau, A.T.; Clayton, J.S. Fish effects on charophyte establishment in a shallow, eutrophic New Zealand lake. N. Z. J. Mar. Freshw. Res. 2002, 36, 815–823. [Google Scholar] [CrossRef] [Green Version]
- Blindow, I. Decline of charophytes during eutrophication: Comparison with angiosperms. Freshw. Biol. 1992, 28, 9–14. [Google Scholar] [CrossRef]
- Liboriussen, L.; Jeppesen, E. Temporal dynamics in epipelic, pelagic and epiphytic algal production in a clear and turbid shallow lake. Freshw. Biol. 2003, 48, 418–431. [Google Scholar] [CrossRef]
- Barko, J.W.; James, W.F. Effects of Submerged Aquatic Macrophytes on Nutrient Dynamics, Sedimentation, Resuspension; Jeppesen, E., Søndergaard, M., Christoffersen, K., Eds.; Springer: New York, NY, USA, 1998; pp. 197–214. [Google Scholar]
- Carlton, R.G.; Wetzel, R.G. Phosphorus flux from lake sediments: Effects of epipelic algal oxygen production. Limnol. Oceanogr. 1988, 33, 562–570. [Google Scholar] [CrossRef]
- Brooks, J.; Dodson, S. Predation, body size and composition of plankton. Science 1965, 150, 28–35. [Google Scholar] [CrossRef]
- Mellors, W.K. Selective predation of ephippal daphnia and the resistance of ephippal eggs to digestion. Ecology 1975, 56, 974–980. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Zooplankton Groups | Control | Fish |
|---|---|---|
| Nauplies | 6.7 ± 9.2 | 0.06 ± 0.07 ** |
| Copepodites | 45.2 ± 37.0 | 1.3 ± 1.5 ** |
| Adalt copepods | 2.0 ± 2.1 | 0.05 ± 0.09 ** |
| Cladocera | 111.3 ± 113.3 | 0.1 ± 0.1 ** |
| Rotifera | 0.4 ± 0.9 | 3.0 ± 2.8 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Razlutskij, V.; Mei, X.; Maisak, N.; Sysova, E.; Lukashanets, D.; Makaranka, A.; Jeppesen, E.; Zhang, X. Omnivorous Carp (Carassius gibelio) Increase Eutrophication in Part by Preventing Development of Large-Bodied Zooplankton and Submerged Macrophytes. Water 2021, 13, 1497. https://doi.org/10.3390/w13111497
Razlutskij V, Mei X, Maisak N, Sysova E, Lukashanets D, Makaranka A, Jeppesen E, Zhang X. Omnivorous Carp (Carassius gibelio) Increase Eutrophication in Part by Preventing Development of Large-Bodied Zooplankton and Submerged Macrophytes. Water. 2021; 13(11):1497. https://doi.org/10.3390/w13111497
Chicago/Turabian StyleRazlutskij, Vladimir, Xueying Mei, Natallia Maisak, Elena Sysova, Dzmitry Lukashanets, Andrei Makaranka, Erik Jeppesen, and Xiufeng Zhang. 2021. "Omnivorous Carp (Carassius gibelio) Increase Eutrophication in Part by Preventing Development of Large-Bodied Zooplankton and Submerged Macrophytes" Water 13, no. 11: 1497. https://doi.org/10.3390/w13111497