
Fish Assemblage Shift after Japanese Smelt (Hypomesus nipponensis McAllister, 1963) Invasion in Lake Erhai, a Subtropical Plateau Lake in China


1
State Key Laboratory of Freshwater Ecology and Biotechnology, Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan 430072, China
2
University of Chinese Academy of Sciences, Beijing 100049, China
*
Author to whom correspondence should be addressed.
Water 2021, 13(13), 1800; https://doi.org/10.3390/w13131800
Submission received: 28 May 2021
/
Revised: 26 June 2021
/
Accepted: 27 June 2021
/
Published: 29 June 2021
(This article belongs to the Special Issue Biological Invasions in Changing Aquatic Ecosystems: Future Perspectives)
Abstract
:The introduction of non-native fish species is known to have adverse effects on aquatic ecosystems, but their effect on plateau lakes is not well studied. In this study, we examined the effect of the Japanese smelt (Hypomesus nipponensis) invasion on the fish assemblage in Lake Erhai, a subtropical plateau lake in southwestern China. Through cluster analysis and non-metric multidimensional scaling (NMDS), we found a significant fish assemblage shift: the population of sharpbelly (Hemiculter leucisculus) fell by 67% in catch per unit effort (CPUE) from 2.262 to 0.741; topmouth gudgeon (Pseudorasbora parva) fell by 52% from 0.61 to 0.29; and icefish (Neosalanx taihuensis) plummeted by 88% from 0.736 to 0.088. Meanwhile, the numbers for crucian carp (Carassius auratus) improved by almost 185% from 1.82 to 3.36. A Pearson correlation analysis showed that these four species significantly correlated with the invasion of the Japanese smelt: sharpbelly (−0.71), topmouth gudgeon (−0.71), icefish (−0.62), and crucian carp (0.81). This study documented the expansion of invasive fish and their effects on native species over time, thus providing a case study of invasive fish as well as a theoretical basis for further research into interspecies interactions.
1. Introduction
Non-native fish are often introduced to enhance aquaculture and fisheries in freshwater ecosystems [1,2], but many come at a significant ecological, evolutionary, and economic cost [3,4,5]. Numerous studies have found that the introduction of non-native species has significant impacts on ecosystems, including species extinction [6] and biodiversity loss [7,8], a decrease in the number of native aquatic animals [9,10] and environmental pollution [11]. In addition, non-native species cause significant changes to an ecosystem’s nutritional structure [12,13]. Therefore, understanding the risks non-native fish pose is necessary to determine how to introduce them without damaging the ecosystem [14].
The discovery of an invasive fish usually triggers a survey to determine the scope of its proliferation. Subsequent censuses establish a baseline against which to compare its effect on native organisms; however, detection of an invasive species often occurs after the invasive population has reached a critical threshold and begun to alter the ecosystem. Pre-invasion assessments are often lacking or incomplete, making it difficult to address the impact of an invasion [15].
The lake area of the Yunnan–Guizhou Plateau in southwest China has experienced multiple fish invasions: Rhinogobius giurinus, Rhinogobius cliffordpopei and topmouth gudgeon (Pseudorasbora parva), Barcheek goby (Ctenogobius giurinus), icefish (Neosalanx taihuensis) and sharpbelly (Hemiculter leucisculus) [16]. Lake Erhai, the second-largest lake on the plateau, is already considered an invasive hotspot due to constant introductions. Before the 1950s, the lake’s main economic fish were the Dali schizothorax (Schizothorax taliensis), Dali barb (Barbodes daliensis) and Erhai carp (Cyprinus barbatus). In the 1960s, to increase production, the four major species of Chinese carp were successively introduced and the main fishery gradually shifted from native to non-native fish [17]. During the 1970s, the production of Barcheek goby (Rhinogobius giurinus), boshi goby (Rhinogobius cliffordpopei), swinhon’s sleeper (Micropercops swinhonis) and other introduced fish increased dramatically. Afterwards, production of Dali schizothorax dropped sharply, and most native fish such as the Yunnan schizothorax (Schizothorax yunnanensis) and Erhai carp disappeared. In the mid-1980s, icefish was introduced, and it quickly became a dominant species [18,19,20,21,22], so much so that in 1985 the Dali Erhai Conservation Administration was prompted to start compiling long-term fishery catch statistics to evaluate changes in annual yield. In the 1990s, the production of icefish soared, and the average annual yield rose to 3880.4 t.
From 2000 to 2010, several fish researchers conducted a detailed survey of the fish composition of Lake Erhai [21,23] and found that the Japanese smelt (Hypomesus nipponensis) arrived in 2010. We then conducted annual samplings for nearly a decade (2011–2019) to observe variations in its population. Thus, Lake Erhai is one of the few places where a long-term study to record the expansion and effect of a new invasive species could be conducted because its ecosystem is relatively isolated and provides a more productive and protected refuge than an open lake does.
The Japanese smelt is a small zooplanktivorous fish [24] that exerts remarkable grazing pressure on zooplankton [25]. It has a short life cycle, but it spreads rapidly and has strong adaptability [26]. For example, early studies showed that its initial range extended from Japan to the Korean Peninsula, but it was introduced into other bodies of water and developed on a large scale as an economic fish and an excellent food for carnivorous fish [27,28]. Before the 1980s, Japanese smelt were distributed only in northeastern China, and the number of reservoirs that could sustain production was small. Following its successful introduction in the 1980s and 1990s [27], it spread to numerous bodies of water (mainly reservoirs), including the highland lakes of Yunnan. However, the introduction of the Japanese smelt had also had significant impacts on some aquatic ecosystems. In California’s Sacramento-San Joaquin estuary, a morphologically similar non-native congener wakasagi (H. nipponensis) had emerged, and wakasagi may also negatively affect delta smelt (Hypomesus transpacificus) through competition for food and space, and predation on larval delta smelt [29]. The salmonids kokanee (Oncorhynchus nerka) and pond smelt (Hypomesus transpacificus nipponensis) had been released into a number of Japanese lakes, became an important fishery resource and had an important impact on ecosystems [30]. In Bosten Lake, the largest lake in Xinjiang, the Eurasian perch (Perca fluviatilis L.) population declined dramatically after the introduction of the Japanese smelt [31,32], and studies on Lake Ulungu showed that it supplanted the native Leuciscus (Leuciscus baicalensis) as the dominant species [33]. The fight to survive in this fierce interspecific competition has pushed native fish to the edge, resulting in a decrease in populations and deterioration to the watershed environment [34]. Lake Erhai is currently in the early stages of eutrophication, with total nitrogen (TN) values ranging from 0.60–0.80 mg L−1 and total phosphorus (TP) values ranging from 0.015–0.037 mg L−1. This lake has a large population of planktivorous fish (Unpublished data). Based on previous experience, biomanipulation may be away to control the Japanese smelt. Studies of the Lake Wuhu ecosystem showed that stocking it with piscivorous fish (Culter alburnus) reduced the number of planktivorous fish [35]. Additionally, the cold-water piscivorous O. mykiss was selected to mitigate the planktivorous H. transpacificus nipponensis in Lake Shirakaba, Japan [36].
The isolation of highland lakes allows it to provide an isolated environment for Lake Erhai to track the time course of invasions. Many of the non-native species in this lake can be traced back to the introduction of non-native fish species. Since its discovery in Lake Erhai in 2010, the Japanese smelt population quickly began increasing in abundance from 2016 to 2019 [37]. However, it is unknown how native and non-native fish are responding to the growing Japanese smelt population. Therefore, the objective of this study was to examine changes in fish communities and explore the effect of growing populations of the invasive Japanese smelt on the abundance of native and non-native fish populations.
2. Materials and Methods
2.1. Study Area
Lake Erhai is 1973.7 m a.s.l. located in a subtropical monsoon climate zone in the Dali Bai Autonomous Prefecture. With a surface area of 249.8 km2, it is the second-largest plateau freshwater lake in Yunnan province. Its year-round temperature is between 10 and 20 °C [38], average annual rainfall is 1060 mm, and the average and maximum water depths are 10.7 m and 22 m, respectively [16]. Vegetation is the dominant watershed land type, and the highest proportion of riparian vegetation and agricultural land 49 and 26%, respectively, followed by 13% of construction land [39]. Rapid economic development in recent years has led to urbanization and industrialization, which has harmed the watershed [16]. This lake also has experienced a series of fish species invasions and loss of native fish diversity in recent decades [23]. Commercial fish such as four major Chinese carp had been introduced successively in Lake Erhai since the 1960s, and the main fishery had gradually shifted from native fishes to artificially introduced non-native fish [21]. In addition, due to the introduction of commercial fish, the fish production in Lake Erhai increased from less than 2500 tons in 2001 to more than 5000 tons in 2009 [40].
2.2. Sampling Sites
Based on habitat variation and anthropogenic activity, we selected seven sampling sites along Lake Erhai (Xiaoguanyi, Haidong Town, Caicun Village, Xizhou Town, Wase Town, Hewei Village and Shuanglang Town) (Figure 1). These sites were subdivided according to water depth and gradient, substrate and aquatic plant, which basically covered different habitat types in Lake Erhai [17]. The indicators of ecological factors such as water depth range, variation of water depth gradient, substrate type, and aquatic plant abundance at each site are as follows (Table 1).
2.3. Sampling Methods
From January 2011 to December 2019, fish surveys were conducted every season at the sites (Figure 1) and the weights of each species were recorded. The fish were caught 8 × 1.5 × 0.7 m vessels and 4236 vessels were surveyed. The nets used by professional fishermen included 50 standardized gillnets and benthic fyke-nets and 25 surface nets and sink nets. The gillnets were 50 m in length and 1.5–15 m in height (mesh size = 5–160 mm). The length, width, height and mesh of the benthic fyke-nets were 20, 0.62, 0.35 m and 5 mm, respectively and were mainly used to capture demersal fish. The nets were set daily at dusk and hauled in early the following morning. Since data on the fishing effort time for some vessels were missing, and the type of vessel used for fishing was consistent, we used the daily weight of fish per vessel to calculate the catch per unit effort (CPUE), the weight of fish per vessel per day. At the same time, we investigated and collected fish in the township markets near the sampling sites and strengthened the compilation of fish species, which were divided into 6 trophic groups according to diet: omnivore, carnivore, herbivore, phytoplanktivore, zooplanktivore or detritivore [23]. They were classified and identified according to Fishes of Yunnan (vol. 1 and vol. 2) [41,42]. Any unidentified fish species was photographed, placed in an anhydrous ethanol solution and brought to a laboratory for further identification. Then it was given a scientific name according to Fishbase (http://www.fishbase.org/search.php (accessed on 20 May 2019)).
The data on the fish composition of Lake Erhai before 2011 is well recorded in the literature [21], but from 2011 onwards they come from our surveys. According to their main dietary sources, all fish were divided into 6 trophic groups: omnivore, carnivore, herbivore, phytoplanktivore, zooplanktivore and detritivore [23].
2.4. Data Analysis
Based on the presence or absence data of species composition, the Jaccard distance was calculated to obtain a similarity distance matrix. Then, a cluster analysis of a similarity distance matrix was carried out using the unweighted pair group method with arithmetic mean (UPGMA), and species composition was sorted by Non-metric Multidimensional Scaling (NMDS) to identify the structural characteristics of annual fish assemblage. The cluster analysis was conducted using the software package “Vegan” R [43].
Although Japanese smelt had been detected several years earlier, we first collected them in 2013 and began to produce capture yields in 2016. Therefore, for this study the “pre-Japanese smelt period” was 2011–2015 and the “post-Japanese smelt” was 2016–2019. A t-test for the establishment of Japanese smelt was conducted to compare the CPUE for the two periods. Prior to the analysis, the data had been tested for parametric statistical analysis. The fish CPUE data in this study achieved normality (Shapiro-Wilk test) and homogeneity of variance (Levene’s test), and the t-test was used to compare the means between the groups [44]. A Pearson correlation analysis was used to identify the relationship between Japanese smelt and the dominant fish species. CPUE data were reported as means ±1 standard deviation to show trends in catch rate across years. All data analysis was performed with R version 4.0.3 (R Development Core Team, https://www.r-project.org/ (accessed on 10 October 2019)).
3. Results
3.1. Fish Species Composition
The fish specimens captured were categorized into 29 species of 12 families (Table 2). The most abundant species was Cyprinidae (55.2%), followed by Gobiidae and Siluridae, respectively. Single species were Cobitidae, Synbranchidae, Eleotridae, Salangidae, Channidae, Poeciliidae, Osmeridae, Belontiidae, and Bagridae. In our study, two other invasive fish species besides the Japanese smelt were found: Southern catfish (Silurus meridionalis Chen) and Catfish (Clarias gariepinus), which appeared after 2014 (Table 2).
3.2. Changes in Fish Communities
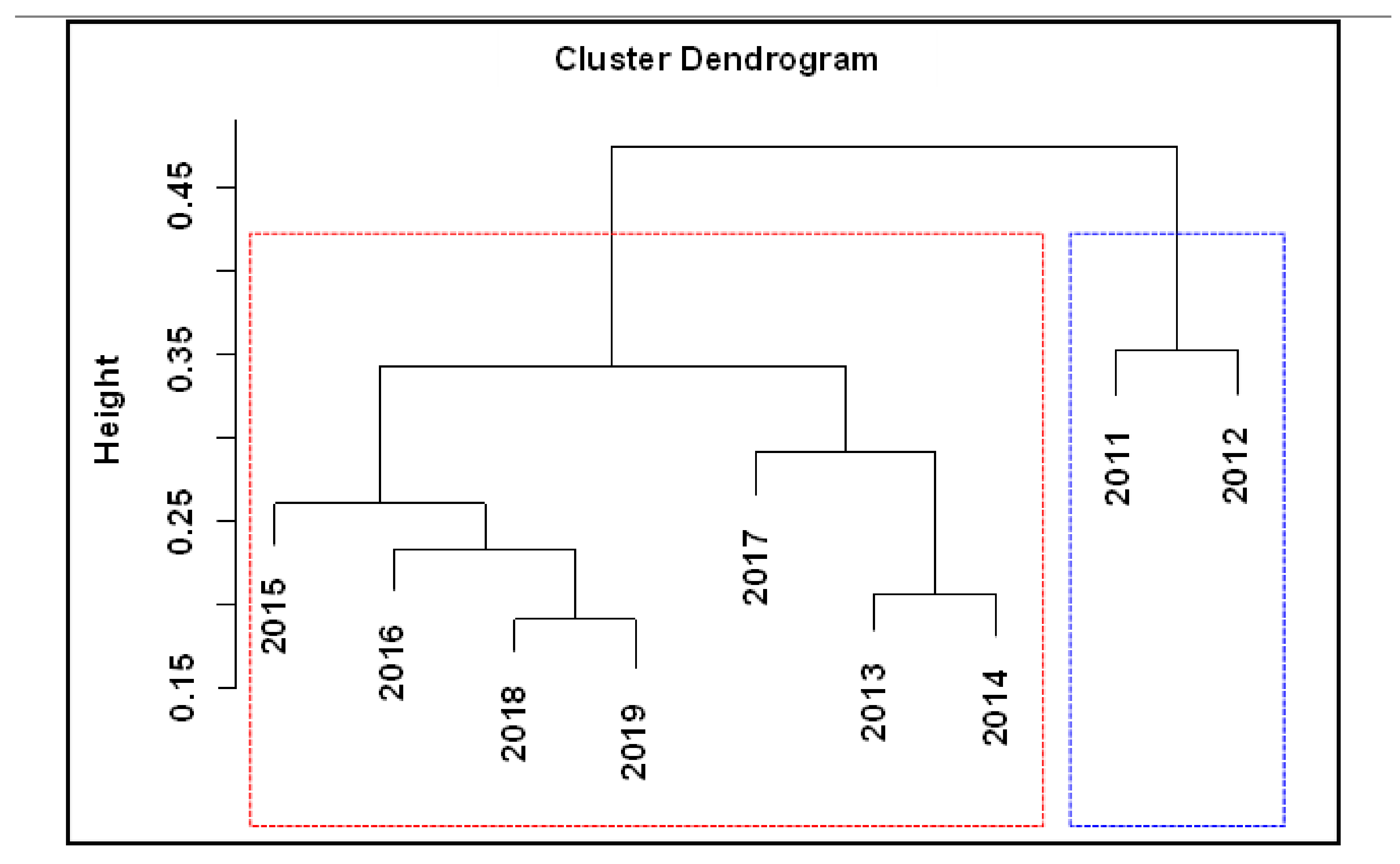
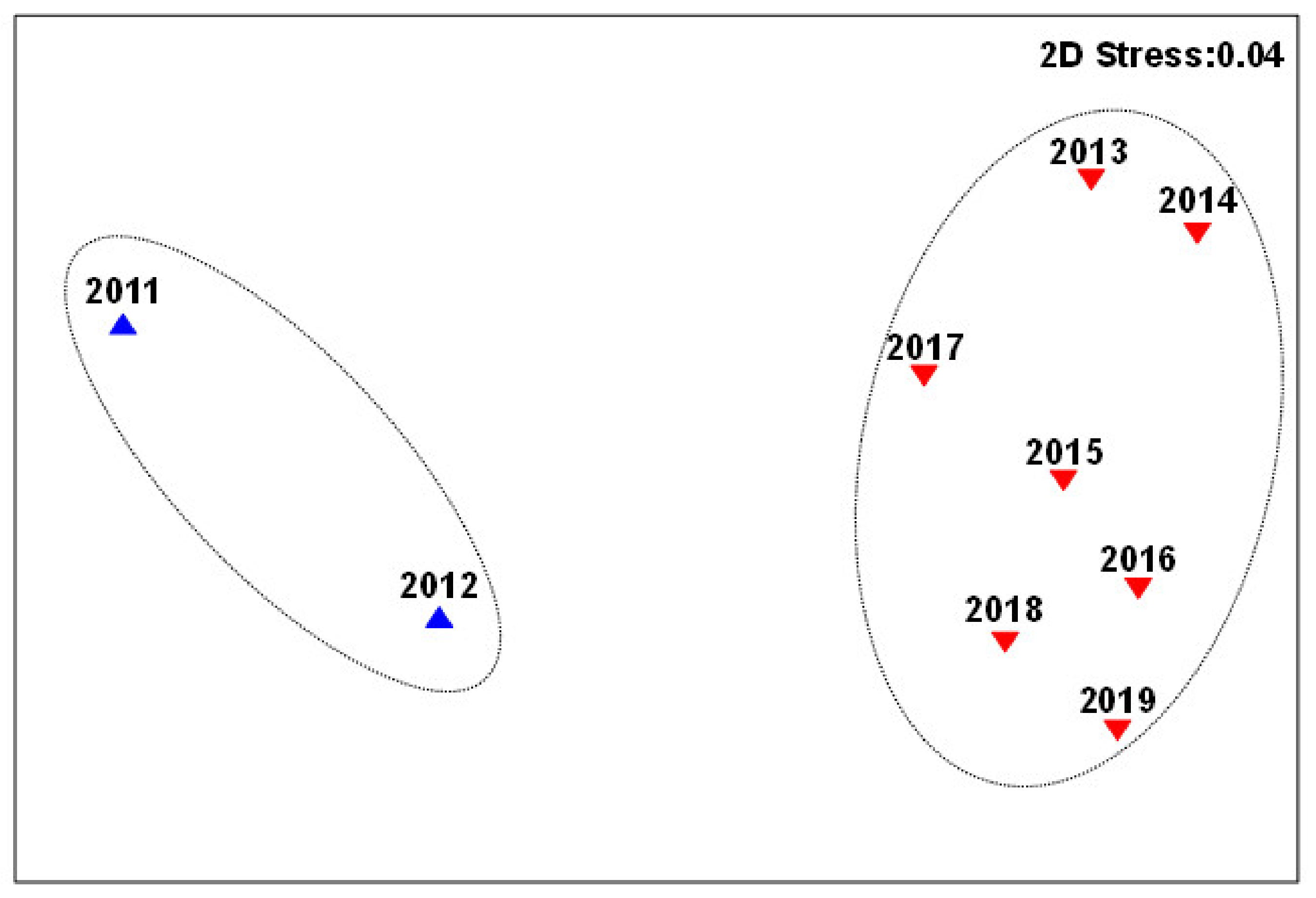
Our cluster analysis divided the fish assemblage into two significantly different relatively homogeneous groups (Figure 2), the first from 2011 to 2012 and the second from 2013 to 2019. Cluster analysis is a set of statistical analysis techniques that classify study subjects into relatively homogeneous groups (clusters). Like the Japanese smelt, the other two invasive species were collected for the first time after 2013; however, bluntsnout bream (Megalobrama amblycephala) and mosquitofish (Gambusia affinis) were not captured. Thus, cluster analysis divided the fish assemblage into two groups with distinct differences among years. NMDS sorting based on species composition divided the fish communities into two groups that had temporal continuity (Figure 3) because the entry of species into an ecosystem creates a continuous disturbance. The NMDS scale results were represented by a two-dimensional image with a stress coefficient stress = 0.04, which is a good ranking, so the component classification was considered reliable.
3.3. Variations in Population Abundance
The abundance of invasive Japanese smelt was extremely low when it was first detected in Lake Erhai (Table 2; Figure 4) but increased rapidly after 2016. By 2019, the annual yield exceeded 2500 t (Table 2), and the annual mean CPUE after 2016 increased abruptly, reaching 4.7 ± 0.89 in 2019. This may be due to the reason that Lake Erhai provides sufficient food sources and a good growing environment for the Japanese smelt, causing its population abundance to increase dramatically. Although southern catfish were also found, their populations were extremely low, with registering a maximum CPUE of only 0.037 (2017) and 0.011 (2019), respectively. Thus, their CPUE was not analyzed due to the low abundance (Figure 4).
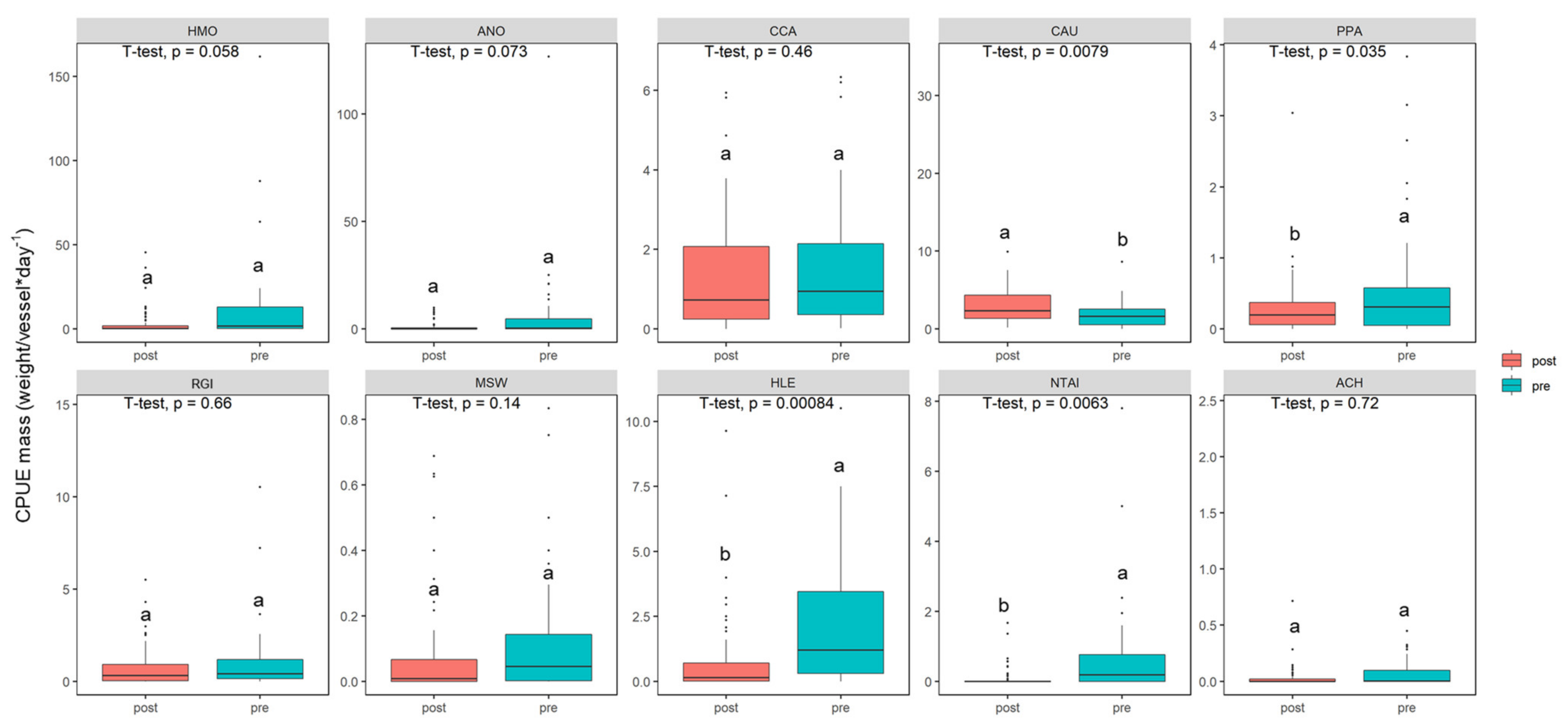
Variation in the CPUE for the 10 dominant fish species (relatively high abundance and frequency of occurrence) for pre- and post-Japanese smelt invasion, respectively, are shown in Figure 5. Pre-Japanese smelt abundance for sharpbelly (2.262 ± 0.39 CPUE) was significantly different in post-Japanese smelt years (0.741 ± 0.18 CPUE) (t-test, p = 0.00084 < 0.001, N = 110). Pre-Japanese smelt abundance for icefish (0.736 ± 0.22 CPUE) was statistically significant (t-test, p = 0.0063 < 0.01, N = 110) from post-Japanese smelt abundance (0.088 ± 0.03 CPUE). The population of crucian carp increased significantly after Japanese smelt detection averaging 3.36 ± 0.51 CPUE versus 1.82 ± 0.25 CPUE pre-Japanese smelt (t-test, p = 0.0079 < 0.01, N = 110). Pre-Japanese smelt CPUE (0.61 ± 0.14) for topmouth gudgeon was significantly higher than post-Japanese smelt median CPUE (0.29 ± 0.05) (t-test, p = 0.035 < 0.05, N = 110). However, there was no significant variation in CPUE for silver carp, bighead carp, common carp, Barcheek goby, Swinhons sleeper, or khanka spiny bitterling (Figure 5).
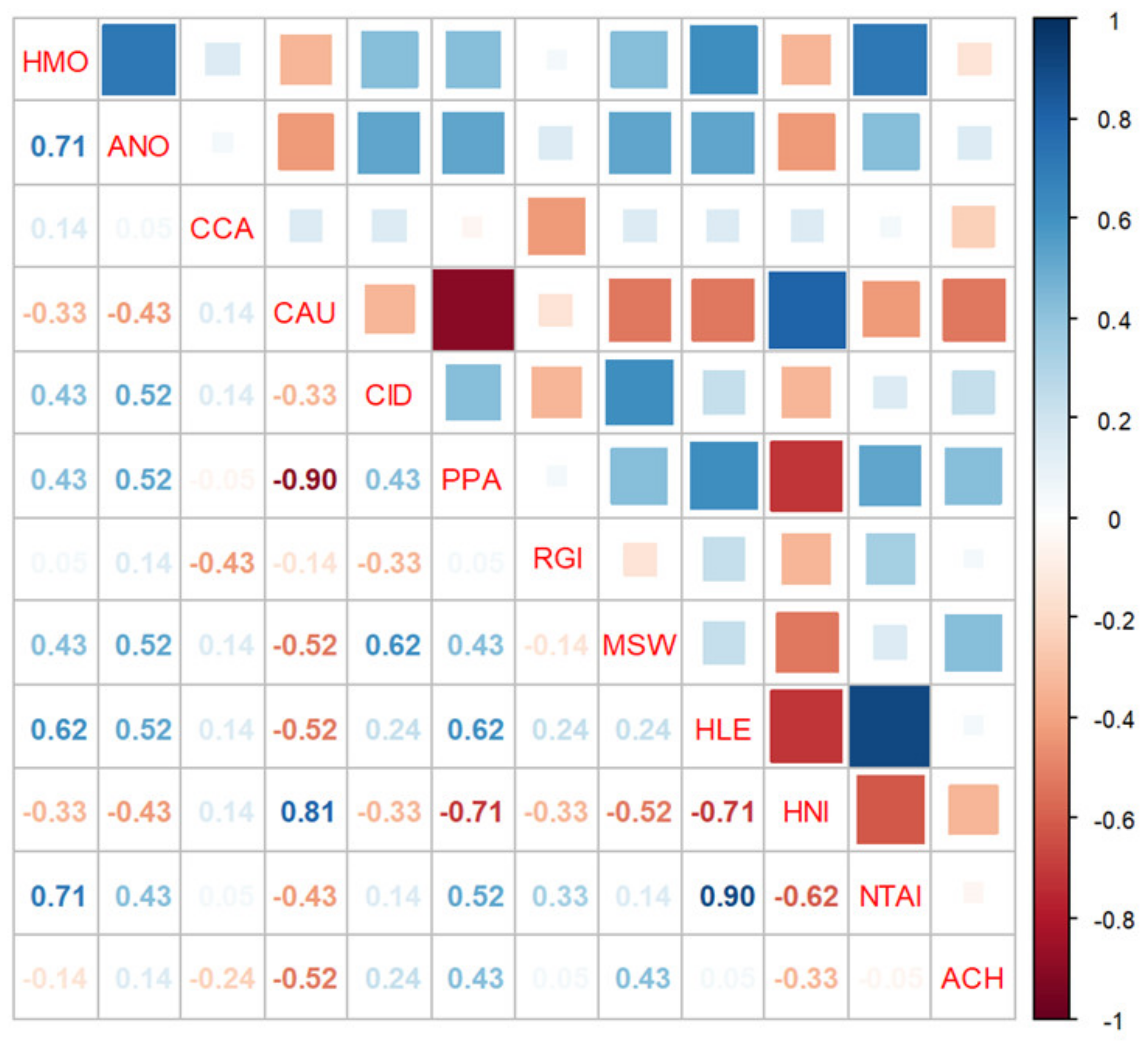
The consequences of the invasive Japanese smelt on the CPUE of dominant fish species are presented in Figure 6. The results showed that the four species that significantly correlated with the invasion of the Japanese smelt were icefish (−0.62), sharpbelly (−0.71), crucian carp (0.81), and topmouth gudgeon (−0.71) (Figure 6). This suggested that the increasing population of Japanese smelt significantly affected the abundance of this fish population.
4. Discussion
The long sampling of this study provided an excellent opportunity to follow a fish community. Collection efforts began with the discovery of the icefish, but its presence allowed us to maintain records for the introduction and rapid spread of the Japanese smelt. This study used the same techniques to sample the same areas over a nine-year period, but a shortcoming of this study needs to be addressed. The detrimental effects of an invasive species may show up in its abundance and size [1,15], but without length measurements, it was difficult to determine whether the Japanese smelt had a negative effect on fish size. Although biomass may be a better indicator, this was not available, so we performed a correlation analysis between the abundance of non-native and native fish. Despite the limitations, this study provided a reliable assessment of the impact of the Japanese smelt invasion on the fish assemblage in Lake Erhai.
During this study, a total of 29 species of fish were found, including 22 non-native species. Three species of non-native fish were the first to be recorded after 2012. Therefore, we used the Jaccard distance matrix and NMDS method to investigate whether the fish assemblage changed after the fish invasion. The results showed that the fish assemblage in Lake Erhai at different times was divided into two distinct groups, as the first group (2011 and 2012) and the second group (2013–2019). As the Japanese smelt, Southern catfish and Catfish were collected for the first time after 2013. Meanwhile, bluntsnout bream (Megalobrama amblycephala) and mosquitofish (Gambusia affinis) were not captured. Thus, cluster analysis divided the fish assemblage into two groups with distinct differences among years. In China, as a means of fishery development, the fish introduction had been implemented widely and had greatly altered fish assemblage since the middle of the twentieth century [45]. Lake Erhai was a plateau lake invaded seriously by non-native fish. Fish species were introduced into the lake accidentally and caused the fish communities to vary considerably [20].
Currently, it is generally believed that fish introduction could impact native ecosystems through predatory interactions that trigger a trophic cascade and alter nutrient recycling [46]. Lake Erhai was a lake with serious fish miniaturization, planktivorous fishes and bottom omnivorous fishes were greater [23]. Our results found that the abundance of Japanese smelt, which is highly selective for large Daphnia zooplankton [47], caused a drastic reduction in the CPUE of small, pelagic icefish, which mainly feed on zooplankton [48,49,50]. The Japanese smelt also feeds mainly on zooplankton and is highly selective for large Daphnia species [47]. Interspecific competition for food caused some fish resources to vary greatly. Previous studies showed that the accidental introduction of non-native fish produced a sharp, rapid expansion into the ecological niche occupied by native fish, leading to a sharp decline in native fish resources and their eventual extinction [19,20]. Furthermore, the time of Japanese smelt reproduction (January–April) [47] precedes that of the icefish [18] and other species like the sharpbelly [51] and the topmouth gudgeon, an omnivorous fish [52,53,54] that feeds on zooplankton and benthic animals. The result led to food shortages during the early stages of the icefish [55,56], thus causing its decline.
The impact of zooplanktivorous invaders is thought to be found primarily through competition for depleted zooplankton [57,58] and shifts in the zooplankton community composition: reducing the dominance of large zooplankton and increasing the medium-sized zooplankton or small rotifer abundance [59,60]. Previous studies have indicated that the small planktivorous sharpbelly fed mainly on zooplankton [61], but recent studies have found that the abundance of macro-zooplankton has decreased, which may explain the decline in the sharpbelly CPUE. Additionally, the reproduction of the Japanese smelt population in Lake Erhai occurred mainly from January to April [47], earlier than that of the majority of other fish species including sharpbelly [51]. For, like most other fishes that have a zooplanktivores stage in their early life history [55], the introduction of the Japanese smelt may lead to food shortages among these species in their early life stages, which might reduce early growth and development and thus increase their early-life mortality [56].
Our results also showed that the CPUE of topmouth gudgeon decreased significantly after the invasion of Japanese smelt, as well as the changes in CPUE of Japanese smelt and topmouth gudgeon CPUE were significantly negatively correlated. Topmouth gudgeon is an omnivorous fish [52,53,54], which feeds mainly on zooplankton and benthic animals in Lake Erhai [61]. The reproduction of the Japanese smelt population in Lake Erhai occurs mainly from January to April [47], which is earlier than that of the topmouth gudgeon [62]. Like most other fishes that have a zooplanktivores stage in their early life history [56], the full utilization of food during the reproductive stage of the Japanese smelt may reduce the growth of the topmouth gudgeon.
Numerous studies have been conducted on the effects of invasive piscivorous fish on fish ecosystems [63,64], but studies into the effects of introduced planktivorous fish are very limited [65,66,67]. The introduction of Japanese smelt into Lake Erhai has led to a reduction in resources and even the extinction of some native fish [29,30,33]. The results of this study concluded that the invasion of the Japanese smelt has led to a decrease in the abundance of sharpbelly, icefish, and topmouth gudgeon. We also found a significant increase in CPUE for crucian carp, which could be due to the reduction of planktivorous fish, which compete for the same food: benthic macroinvertebrates such as chironomidae and oligochaeta [68]. However, further research is needed to examine the relationship between the population growth and trophic aspects of invasive zooplanktivorous fish and crucian carp. However, there were no significant changes in CPUE for silver carp, bighead carp, carp, Bacchae goby, Swinhons sleeper and Kanka spiny bittern in our results. Annual stocking of silver carp, bighead carp and common carp may be responsible for the lack of significant change, while other fish were affected to a lesser extent due to the difference between their habitat and those of the Japanese smelt.
Three invasive fish species have emerged in Lake Erhai over the past 10 years, with a major invasive Japanese smelt outbreak that increased rapidly. Interspecific interactions within a given density range have resulted in increased populations of invasive species [69]. Following the introduction of any successful new populations, there is a need to determine if the new populations are displacing natives or exploiting a new niche. From 2011 to 2019, there was a dramatic increase in the abundance of the Japanese smelt, while the total fish caught decreased relatively. This suggests that the invasion of the Japanese smelt has greatly increased its total catch and depressed the native populations already. Lake Erhai has a large and complex ecosystem. It is difficult to identify a single cause for the increase or decrease of any species. However, this long-term survey provides an overview of fish community dynamics and argues for a more in-depth study and management of invasive fish in plateau lakes.
5. Conclusions
In the current study, the results show that the invasion of Japanese smelt significantly changed fish assemblages in Lake Erhai, especially for icefish (Neosalanx taihuensis), sharpbelly (Hemiculter leucisculus), topmouth gudgeon (Pseudorasbora parva), and crucian carp (Carassius auratus). Our results provide a basis for fishery management and a theoretical basis for the next study of interspecific interactions. Meanwhile, Lake Erhai, as one of the numerous lakes on the Yunnan-Guizhou Plateau, and can be used as a case study so that, when a new species arrives in a plateau lake, the progress of its invasion can be documented.
Author Contributions
C.Y.: Conceptualization, methodology, investigation, data curation, software, writing—original draft preparation and writing—review and editing. Y.C.: writing—review and editing. L.G.: funding provision, Conceptualization, writing—review and editing. L.N.: writing—review and editing, supervision. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ecological effects, population regulation and management strategies of invasion of Japanese smelt (Hypomesus nipponensis) in Lake Erhai, entrusted by Dali City, Yunnan Province, China (No. 9, 2018, approved by the Government of Dali City).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We thank all those who contributed to this work.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Britton, J.R.; Gozlan, R.E.; Copp, G.H. Managing non-native fish in the environment. Fish Fish. 2011, 12, 256–274. [Google Scholar] [CrossRef]
- Peeler, E.J.; Oidtmann, B.C.; Midtlyng, P.J.; Miossec, L.; Gozlan, R.E. Non-native aquatic animals introductions have driven disease emergence in Europe. Biol. Invasions 2010, 13, 1291–1303. [Google Scholar] [CrossRef] [Green Version]
- Jeschke, J.M.; Strayer, D.L. Invasion success of vertebrates in Europe and North America. Proc. Natl. Acad. Sci. USA 2005, 102, 7198–7202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricciardi, A.; Kipp, R. Predicting the number of ecologically harmful exotic species in an aquatic system. Divers. Distrib. 2007, 14, 374–380. [Google Scholar] [CrossRef]
- Cucherousset, J.; Olden, J.D. Ecological Impacts of Nonnative Freshwater Fishes. Fisheries 2011, 36, 215–230. [Google Scholar] [CrossRef]
- Gurevitch, J.; Padilla, D.K. Are invasive species a major cause of extinctions? Trends Ecol. Evol. 2004, 19, 470–474. [Google Scholar] [CrossRef] [PubMed]
- Pyšek, P.; Richardson, D.M. Invasive Species, Environmental Change and Management, and Health. Annu. Rev. Environ. Resour. 2010, 35, 25–55. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, F.; Leunda, P.M. Non-native fish impacts on Mediterranean freshwater ecosystems: Current knowledge and research needs. Fish. Manag. Ecol. 2012, 19, 142–156. [Google Scholar] [CrossRef]
- Marr, S.M.; Ellender, B.R.; Woodford, D.J.; Alexander, M.E.; Wasserman, R.J.; Ivey, P.; Zengeya, T.; Weyl, O.L. Evaluating invasion risk for freshwater fishes in South Africa. Bothalia 2017, 47. [Google Scholar] [CrossRef]
- Ellender, B.; Weyl, O. A review of current knowledge, risk and ecological impacts associated with non-native freshwater fish introductions in South Africa. Aquat. Invasions 2014, 9, 117–132. [Google Scholar] [CrossRef]
- Matsuzaki, S.S.; Usio, N.; Takamura, N.; Washitani, I. Contrasting impacts of invasive engineers on freshwater ecosystems: An experiment and meta-analysis. Oecologia 2009, 158, 673–686. [Google Scholar] [CrossRef] [PubMed]
- Ellis, B.K.; Stanford, J.A.; Goodman, D.; Stafford, C.P.; Gustafson, D.L.; Beauchamp, D.A.; Chess, D.W.; Craft, J.A.; Deleray, M.A.; Hansen, B.S. Long-term effects of a trophic cascade in a large lake ecosystem. Proc. Natl. Acad. Sci. USA 2011, 108, 1070–1075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rypel, A.L. Do invasive freshwater fish species grow better when they are invasive? Oikos 2014, 123, 279–289. [Google Scholar] [CrossRef]
- Gozlan, R.E.; Britton, J.R.; Cowx, I.; Copp, G.H. Current knowledge on non-native freshwater fish introductions. J. Fish Biol. 2010, 76, 751–786. [Google Scholar] [CrossRef]
- Leino, J.R.; Mensinger, A.F. The benthic fish assemblage of the soft-bottom community of the Duluth-Superior Harbor before and after round goby invasion (1989–2011). J. Great Lakes Res. 2016, 42, 829–836. [Google Scholar] [CrossRef]
- Wang, S.H.; Wang, J.; Li, M.B.; Du, F.; Yang, Y.M.; Lassoie, J.P.; Hassan, M.Z. Six decades of changes in vascular hydrophyte and fish species in three plateau lakes in Yunnan, China. Biodivers. Conserv. 2013, 22, 3197–3221. [Google Scholar] [CrossRef]
- Fei, J.H. Studies on the Present Condition of Fish Stocks and Its Spatial Structure, Growth Characteristics in Erhai Lake. Master’s Thesis, Hangzhou Normal University, Hangzhou, China, 2012. [Google Scholar]
- Zhu, F.Y.; Ye, S.W.; Li, Z.J.; Zhang, T.L.; Yuan, J.; Guo, Z.Q.; Tang, J.F.; Liu, J.S. Growth and reproduction of the non-native icefish Neosalanx taihuensis Chen, 1956 (Salangidae) in a plateau lake, southwestern China. J. Appl. Ichthyol. 2014, 30, 862–869. [Google Scholar] [CrossRef]
- Du, B.H.; Li, Y.A. Danger Risk to Fish Diversity in Erhai Lake and Proposals to Dispel It. Res. Environ. Sci. 2001, 14, 42–44. [Google Scholar] [CrossRef]
- Wu, Q.L.; Wang, Y.F. On the succession of Aquatic Communities in Erhai Lake. J. Lake Sci. 1999, 11, pp. 267–273. Available online: https://www.semanticscholar.org/paper/On-the-Succession-of-Aquatic-Communities-in-Erhai-Qinglon/af6872c768d2839fc7b5355bc8144bb4ef37a153.
- Jiang, Z.G.; Brosse, S.; Jiang, X.M.; Zhang, E. Measuring ecosystem degradation through half a century of fish species introductions and extirpations in a large isolated lake. Ecol. Indic. 2015, 58, 104–112. [Google Scholar] [CrossRef]
- Lin, Q.Q.; Zhao, P.P.; Wu, Q.T.; Wei, Z.H.; Li, H.M.; Han, B.P. Predation pressure induced by seasonal fishing moratorium changes the dynamics of subtropical Cladocera populations. Hydrobiologia 2013, 710, 73–81. [Google Scholar] [CrossRef]
- Tang, J.F.; Ye, S.W.; Li, W.; Liu, J.S.; Zhang, T.L.; Guo, Z.Q.; Zhu, F.Y.; Li, Z.J. Status and historical changes in the fish community in Erhai Lake. Chin. J. Oceanol. Limnol. 2013, 31, 712–723. [Google Scholar] [CrossRef]
- Saruwatari, T.; Lopez, J.A.; Pietsch, T.W. A revision of the osmerid genus Hypomesus Gill (Teleostei: Salmoniformes), with the description of a new species from the southern Kuril Islands. Species Divers. 1997, 2, 59–82. [Google Scholar] [CrossRef] [Green Version]
- Chang, K.H.; Hanazato, T.; Ueshima, G.; Tahara, H. Feeding habit of pond smelt (Hypomesus transpacificus nipponensis) and its impact on the zooplankton community in lake suwa, Japan. J. Freshw. Ecol. 2005, 20, 129–138. [Google Scholar] [CrossRef]
- Sakamoto, D.; Nemoto, T.; Sunoh, N.; Iwasaki, J.; Niwa, S.; Arayama, K.; Suzuki, N.; Pai, S.; Takagi, K.; Sakuramoto, K. Population size estimation of the pond smelt Hypomesus nipponensis in Lake Kasumigaura and Lake Kitaura, Japan. Fish. Sci. 2014, 80, 907–914. [Google Scholar] [CrossRef]
- Xie, Y.H.; Li, B.; Li, W.L.; Liu, C.X.; Xu, S.Q.; Zhao, X.L. The Fishes of Genus Hypomesus and Utilization of Its Resource; Liaoning Science and Technology Press: Liaoning, China, 1992; pp. 159–164. [Google Scholar]
- Katano, O.; Hakoyama, H.; Matsuzaki, S.S. Japanese inland fisheries and aquaculture: Status and trends. Freshw. Fish. Ecol. 2015, 231–240. [Google Scholar] [CrossRef]
- Swanson, C.; Reid, T.; Young, P.S.; Cech, J.J. Comparative environmental tolerances of threatened delta smelt (Hypomesus transpacificus) and introduced wakasagi (H. nipponensis) in an altered California estuary. Oecologia 2000, 123, 384–390. [Google Scholar] [CrossRef]
- Makino, W.; Mikami, H.; Katano, N.; Nakagawa, M.; Takamura, N. Biological productivity of Lake Towada, a north temperate, oligotrophic, kokanee fishery lake. Limnology 2003, 4, 79–90. [Google Scholar] [CrossRef]
- Guo, Y.; Zhang, R.M.; Cai, L.G.; Tian, Y.; Tilawalti, T.; Lai, Y. Fish Resources and Fishery in the Lake Bositenghu; The Science and Technology Press of Xinjiang: Xinjiang, China, 2005; pp. 1–160. [Google Scholar]
- Zhou, X.Y.; Hu, Z.J.; Liu, Q.G.; Yang, L.L.; Wang, Y.B. Feeding ecology of the non-indigenous fish Hypomesus nipponensis in Lake Ulungur, China: Insight into the relationship between its introduction and the collapse of the native Eurasian perch population. Mar. Freshw. Res. 2013, 64, 549–557. [Google Scholar] [CrossRef]
- Adakbek, K.; Su, D.X.; Yang, Y.; Ye, S.M.; Huderat. The current situation and the countermeasures of protection and development of fish resources in Ulungur Lake. J. Shanghai Fish. Univ. 2006, 15, 308–314. [Google Scholar] [CrossRef]
- Guo, Y.; Zhang, R.M.; Li, H. Degeneration Causes of the Aboriginal Fishes and Some Measures for Protecting Them in the Ergis River, Xinjiang, China. Arid Zone Res. 2003, 20, 152–155, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Lin, M.L. Studies on Stocking Techniques of Culter mongolicus and Ecosystem Effects of Piscivorous Fishes Releasing in Lakes along the Middle and Lower Reach of the Yangtze River. Ph.D. Thesis, Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan, China, 2012. (In Chinese with English Abstract). [Google Scholar]
- Ha, J.Y.; Hanazato, T.; Chang, K.H.; Jeong, K.S.; Kim, D.K. Assessment of the lake biomanipulation mediated by piscivorous rainbow trout and herbivorous daphnids using a self-organizing map: A case study in Lake Shirakaba, Japan. Ecol. Inform. 2015, 29, 182–191. [Google Scholar] [CrossRef]
- Gong, L.; Guo, L.G.; Yin, C.J.; Yang, J.J.; Yang, Y.L. Study on growth characteristics and population regulation effect of Hypomesus nipponesis and Neosalanx taihuensis in Lake Erhai. J. Hydroecology 2020, in press. [Google Scholar]
- Li, Y.; Yen, H.; Lei, Q.L.; Qiu, W.W.; Luo, J.F.; Lindsey, S.; Qin, L.H.; Zhai, L.M.; Wang, H.Y.; Wu, S.X.; et al. Impact of human activities on phosphorus flows on an early eutrophic plateau: A case study in Southwest China. Sci. Total Environ. 2020, 714, 136851. [Google Scholar] [CrossRef] [PubMed]
- Xiang, S.; Wang, L.; Pang, Y. Spatial-temporal variation of inflow river water quality under land use effect. J. Agro-Environ. Sci. 2020, 39, 160–170. [Google Scholar]
- Fei, J.H.; Tang, T.; Shao, X.Y. Fishery resources and developmental patterns of fishery in Erhai Lake. Wetl. Sci. 2011, 9, 277–283. [Google Scholar] [CrossRef]
- Chu, X.L.; Chen, Y.R. Fishes of Yunnan, Vol. I.; Science Press: Beijing, China, 1989. (In Chinese) [Google Scholar]
- Chu, X.L.; Chen, Y.R. Fishes of Yunnan, Vol. II.; Science Press: Beijing, China, 1990. (In Chinese) [Google Scholar]
- Knights, D.; Kuczynski, J.; Charlson, E.S.; Zaneveld, J.; Mozer, M.C.; Collman, R.G.; Bushman, F.D.; Knight, R.; Kelley, S.T. Bayesian community-wide culture-independent microbial source tracking. Nat. Methods 2011, 8, 761–763. [Google Scholar] [CrossRef] [Green Version]
- Quinn, G.; Keough, M. Experimental Design and Data Analysis for Biologists; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Liu, G.M.; Liu, Z.W.; Li, Y.L.; Chen, F.Z.; Gu, B.H.; Smoak, J.M. Effects of fish introduction and eutrophication on the cladoceran community in Lake Fuxian, a deep oligotrophic lake in southwest China. J. Paleolimnol. 2008, 42, 427–435. [Google Scholar] [CrossRef] [Green Version]
- Sarnelle, O.; Knapp, R.A. Nutrient recycling by fish versus zooplankton grazing as drivers of the trophic cascade in alpine lakes. Limnol. Oceanogr. 2005, 50, 2032–2042. [Google Scholar] [CrossRef]
- Gong, L. Study on Effects of Invasion of Hypomesus nipponensis on Crustacean Zooplankton in Lake Erhai. Master’s Thesis, Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan, China, 2020. [Google Scholar]
- Guo, N.C.; Zhang, M.; Yu, Y.; Qian, S.Q.; Li, D.M.; Kong, F.X. Crustacean zooplankton communities in 13 lakes of Yunnan-Guizhou plateau: Relationship between crustacean zooplankton biomass or size structure and trophic indicators after invasion by exotic fish. Ann. Limnol. Int. J. Limnol. 2009, 45, 279–288. [Google Scholar] [CrossRef]
- Wang, X.L.; Xiang, J.G.; Liu, J.S.; Liu, M.; Wu, L.; Murphy, B.R.; Xie, S.G. Reduced growth and reproductive investment of Hemiculter leucisculus (Cyprinidae) in a reservoir with introduced icefish Neosalanx taihuensis (Salangidae). Environ. Biol. Fishes 2013, 96, 895–903. [Google Scholar] [CrossRef]
- Hu, C.L.; Guo, L.G.; Wang, S.R. Diet and feeding ecology of invasive icefish Neosalanx taihuensis in Erhai Lake, a Chinese plateau meso-eutrophicated lake. Chin. J. Oceanol. Limnol. 2015, 33, 372–380. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Huang, D.; Zhao, Y.; Wang, H.; Hu, S.; Shen, J. Age, growth and mortality of invasive sharpbelly, Hemiculter leucisculus (Basilewski, 1855) in Erhai Lake, China. J. Appl. Ichthyol. 2013, 29, 1279–1285. [Google Scholar] [CrossRef]
- Czerniejewski, P.; Rybczyk, A.; Linowska, A.; Sobecka, E. New location, food composition, and parasitic fauna of the invasive fish Pseudorasbora parva (Temminck & Schlegel, 1846) (Cyprinidae) in Poland. Turk. J. Zool. 2019, 43, 94–105. [Google Scholar] [CrossRef]
- Yalcin-Ozdilek, S.; Kirankaya, S.G.; Ekmekci, F.G. Feeding Ecology of the Topmouth Gudgeon Pseudorasbora parva (Temminck and Schlegel, 1846) in the Gelingullu Reservoir, Turkey. Turk. J. Fish. Aquat. Sci. 2013, 13, 87–94. [Google Scholar] [CrossRef]
- Xie, S.G.; Cui, Y.; Zhang, T.L.; Li, Z. Seasonal patterns in feeding ecology of three small fishes in the Biandantang Lake, China. J. Fish Biol. 2000, 57, 867–880. [Google Scholar] [CrossRef]
- Werner, E.E.; Gilliam, J.F. The ontogenetic niche and species interactions in size structured populations. Annu. Rev. Ecol. Syst. 1984, 15, 393–425. [Google Scholar] [CrossRef]
- Yin, M.C. Feeding, growth and survival of yolk-sac larvae of silver, bighead, grass and crucian carps. Trans. Chin. Ichthyol. Soc. 1997, 6, 69–79. [Google Scholar]
- Amundsen, P.-A.; Siwertsson, A.; Primicerio, R.; Bøhn, T. Long-term responses of zooplankton to invasion by a planktivorous fish in a subarctic watercourse. Freshw. Biol. 2009, 54, 24–34. [Google Scholar] [CrossRef] [Green Version]
- Angeler, D.G.; Álvarez-Cobelas, M.; Sánchez-Carrillo, S.; Rodrigo, M.A. Assessment of exotic fish impacts on water quality and zooplankton in a degraded semi-arid floodplain wetland. Aquat. Sci. 2002, 64, 76–86. [Google Scholar] [CrossRef]
- Bohn, T.; Amundsen, P.-A. Effects of invading vendace (Coregonus Albula L.) on species composition and body size in two zooplankton communities of the Pasvik River System, northern Norway. J. Plankton Res. 1998, 20, 243–256. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.F.; Huang, X.F.; Liu, J.K.; Jiao, N.Z. Effects of fish stocking on the zooplankton community structure in a shallow lake in China. Fish. Manag. Ecol. 2005, 12, 81–89. [Google Scholar] [CrossRef]
- Tang, J.F. Studies on Fisheries Ecology in a Large Yunnan Plateau Lake, Lake Erhai, China. Ph.D. Thesis, Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan, China, 2013. [Google Scholar]
- Davies, G.D.; Britton, J.R. Consistency in the life history traits of four invasive Pseudorasbora parva populations in Southern England. J. Appl. Ichthyol. 2021, 37, 295–302. [Google Scholar] [CrossRef]
- Goudswaard, K.P.C.; Witte, F.; Katunzi, E.F.B. The invasion of an introduced predator, Nile perch (Lates niloticus L.) in Lake Victoria (East Africa): Chronology and causes. Environ. Biol. Fishes 2008, 81, 127–139. [Google Scholar] [CrossRef]
- Copp, G.H.; Britton, J.R.; Cucherousset, J.; García-Berthou, E.; Kirk, R.; Peeler, E.; Stakènas, S. Voracious invader or benign feline? A review of the environmental biology of European catfish (Silurus glanis) in its native and introduced ranges. Fish Fish. 2009, 10, 252–282. [Google Scholar] [CrossRef]
- Brooks, J.L.; Dodson, S.I. Predation body size and composition of plankton. Science 1965, 150, 28–35. [Google Scholar] [CrossRef] [PubMed]
- McCrary, J.K.; Murphy, B.R.; Stauffer, J.R.; Hendrix, S.S. Tilapia (teleostei: Cichlidae) status in Nicaraguan natural waters. Environ. Biol. Fishes 2007, 78, 107–114. [Google Scholar] [CrossRef]
- Britton, J.R.; Davies, G.D.; Brazier, M. Eradication of the invasive Pseudorasbora parva results in increased growth and production of native fishes. Ecol. Freshw. Fish 2009, 18, 8–14. [Google Scholar] [CrossRef]
- Bergman, E.; Greenberg, L.A. Competition between a planktivore, a benthivore, and a species with ontogenetic diet shifts. Ecology 1994, 75, 1233–1245. [Google Scholar] [CrossRef]
- Kornis, M.S.; Mercado-Silva, N.; Vander, Z.M.J. Twenty years of invasion: A review of round goby Neogobius melanostomus biology, spread and ecological implications. J. Fish Biol. 2014, 80, 235–285. [Google Scholar] [CrossRef]
Figure 1.
Location of Lake Erhai and its sampling sites.

Figure 2.
Cluster analysis of fish assemblages in Lake Erhai. Blue boxes represent the first group, red boxes represent the second group.
Figure 2.
Cluster analysis of fish assemblages in Lake Erhai. Blue boxes represent the first group, red boxes represent the second group.

Figure 3.
Non-Metric Multidimensional Scaling (NMDS) analysis of the fish assemblages in Lake Erhai. Blue triangles represent the first group, red triangles represented the second group (Stress = 0.04).
Figure 3.
Non-Metric Multidimensional Scaling (NMDS) analysis of the fish assemblages in Lake Erhai. Blue triangles represent the first group, red triangles represented the second group (Stress = 0.04).

Figure 4.
The mean catch per unit effort (CPUE) is plotted for each calendar year for three invasive species (AS1 = Japanese smelt; AS2 = Southern catfish; AS3 = Catfish) and other fish. Annual yield data (AS1) were derived from the literature [37].
Figure 4.
The mean catch per unit effort (CPUE) is plotted for each calendar year for three invasive species (AS1 = Japanese smelt; AS2 = Southern catfish; AS3 = Catfish) and other fish. Annual yield data (AS1) were derived from the literature [37].

Figure 5.
The box whisker plots comparing pre- and post-Japanese smelt CPUEs for 10 dominant fish species. Each box shows the median value, the bottom and top of the box indicate the first and third quartiles, respectively, and the error bars indicate the 10th and 90th percentiles. A t-test was conducted to compare CPUEs for two different time periods (p < 0.05 indicated a significant difference). Lowercase letters a and b were used to determine whether there was a difference between pre- and post-Japanese smelt (p < 0.05). HMO (Silver carp): Hypophthalmichthys molitrix, ANO (Bighead carp): Aristichthys nobilis, CCA (Common carp): Cyprinus carpio, CAU (Crucian carp): Carassius auratus, PPA (Topmouth gudgeon): Pseudorasbora parva, RGI (Barcheek goby): Rhinogobius giurinus, MSW (Swinhon’s sleeper): Micropercops swinhonis, HLE (Sharpbelly): Hemiculter leucisculus, NTAI (icefish): Neosalanx taihuensis, ACH (Khanka spiny bitterling): Acheilognathus chankaensis.
Figure 5.
The box whisker plots comparing pre- and post-Japanese smelt CPUEs for 10 dominant fish species. Each box shows the median value, the bottom and top of the box indicate the first and third quartiles, respectively, and the error bars indicate the 10th and 90th percentiles. A t-test was conducted to compare CPUEs for two different time periods (p < 0.05 indicated a significant difference). Lowercase letters a and b were used to determine whether there was a difference between pre- and post-Japanese smelt (p < 0.05). HMO (Silver carp): Hypophthalmichthys molitrix, ANO (Bighead carp): Aristichthys nobilis, CCA (Common carp): Cyprinus carpio, CAU (Crucian carp): Carassius auratus, PPA (Topmouth gudgeon): Pseudorasbora parva, RGI (Barcheek goby): Rhinogobius giurinus, MSW (Swinhon’s sleeper): Micropercops swinhonis, HLE (Sharpbelly): Hemiculter leucisculus, NTAI (icefish): Neosalanx taihuensis, ACH (Khanka spiny bitterling): Acheilognathus chankaensis.

Figure 6.
CPUE correlations among dominant fish species and Japanese smelt in Lake Erhai. HMO (Silver carp): Hypophthalmichthys molitrix, ANO (Bighead carp): Aristichthys nobilis, CCA (common carp): Cyprinus carpio, CAU (crucian carp): Carassius auratus, CID (grass carp): Ctenopharyngodon idellus, PPA (topmouth gudgeon): Pseudorasbora parva, RGI (Barcheek goby): Rhinogobius giurinus, MSW (Swinhon’s sleeper): Micropercops swinhonis, HLE (sharpbelly): Hemiculter leucisculus, HNI (Japanese smelt): Hypomesus nipponensis, NTAI (Icefish): Neosalanx taihuensis, ACH (khanka spiny bitterling): Acheilognathus chankaensis.
Figure 6.
CPUE correlations among dominant fish species and Japanese smelt in Lake Erhai. HMO (Silver carp): Hypophthalmichthys molitrix, ANO (Bighead carp): Aristichthys nobilis, CCA (common carp): Cyprinus carpio, CAU (crucian carp): Carassius auratus, CID (grass carp): Ctenopharyngodon idellus, PPA (topmouth gudgeon): Pseudorasbora parva, RGI (Barcheek goby): Rhinogobius giurinus, MSW (Swinhon’s sleeper): Micropercops swinhonis, HLE (sharpbelly): Hemiculter leucisculus, HNI (Japanese smelt): Hypomesus nipponensis, NTAI (Icefish): Neosalanx taihuensis, ACH (khanka spiny bitterling): Acheilognathus chankaensis.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Environmental variables for each of the sampling sites in Lake Erhai.
| Station | Water Depth Range (m) | Water Depth Gradient | Substrate Type | Aquatic Plant Richness |
|---|---|---|---|---|
| Haidong | 1–13 | Medium | Mud | Medium |
| Wase | 2–20 | Large | Silt | Medium |
| Shuanglang | 6–10 | Large | Silt | Low |
| Hewei | 2–6 | Small | Silt | Low |
| Xizhou | 2–15 | Large | Silt | High |
| Caicun | 2–5 | Small | Silt | High |
| Xiaoguanyi | 2–10 | Medium | Silt | High |
Table 2.
Fish composition and CPUE in Lake Erhai from 2011 to 2019.
| Family | Species | Common Names | Code | Trophic Guild | Year | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2000–2010 | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | |||||
| Cyprinidae | Carassius auratus * | Crucian carp | CAU | Omnivore | Y | 0.01 | 0.01 | 1.46 | 1.73 | 1.41 | 2.46 | 4.01 | 3.82 | 4.81 |
| Mylopharyngodon piceus ξ | Black carp | MPI | Carnivore | Y | 0.09 | 0.02 | 0.02 | 0.02 | 0.00 | 0.00 | ||||
| Ctenopharyngodon idellus ξ | Grass carp | CID | Herbivore | Y | 0.07 | 0.06 | 0.39 | 0.23 | 0.17 | 0.20 | 0.10 | 0.38 | 0.17 | |
| Hypophthalmichthys molitrix ξ | Silver carp | HMO | Phytoplanktivore | Y | 3.64 | 15.78 | 6.32 | 3.29 | 11.16 | 5.99 | 0.97 | 3.06 | 1.92 | |
| Aristichthys nobilis ξ | Bighead carp | ANO | Zooplanktivore | Y | 0.00 | 0.01 | 2.87 | 2.52 | 4.34 | 1.98 | 0.41 | 1.40 | 0.99 | |
| Cyprinus carpio ξ | Common carp | CCA | Omnivore | Y | 0.02 | 0.02 | 1.57 | 0.61 | 1.24 | 1.41 | 1.40 | 1.30 | 1.70 | |
| Cyprinus carpio chi1ia * | Chilu carp | CCHI | Omnivore | Y | 0.00 | 0.00 | 0.00 | |||||||
| Cyprinus longipectoralis * | Long-pectoral carp | CLO | Carnivore | Y | 0.00 | |||||||||
| Megalobrama amblycephala ξ | Bluntsnout bream | MAM | Herbivore | Y | 0.00 | 0.00 | 0.09 | 0.02 | 0.00 | |||||
| Pseudorasbora parva ξ | Topmouth gudgeon | PPA | Omnivore | Y | 0.16 | 0.19 | 0.37 | 0.57 | 0.32 | 0.33 | 0.40 | 0.21 | 0.17 | |
| Rhodeus ocellatus ξ | Rosy bitterling | ROC | Detritivore | Y | 0.01 | 0.01 | 0.20 | 0.21 | 0.31 | 0.00 | ||||
| Acheilognathus chankaensis ξ | Khanka spiny bitterling | ACH | Detritivore | Y | 0.01 | 0.15 | 0.21 | 0.03 | 0.00 | 0.20 | 0.07 | 0.06 | ||
| Hemiculter leucisculus ξ | Sharpbelly | HLE | Zooplanktivore | Y | 1.77 | 0.08 | 0.52 | 0.87 | 2.00 | 1.23 | 0.63 | 0.26 | 0.39 | |
| Schizothorax wangchiachii * | Duanxu schizothorax | SWA | Zooplanktivore | Y | 0.00 | 0.00 | 0.00 | 0.00 | ||||||
| Abbottina rivularis ξ | Chinese false gudgeon | ARI | Omnivore | Y | 0.00 | 0.00 | 0.01 | |||||||
| Cyprinus carpio haematopterus * | Mirror carp | CCH | Omnivore | Y | 0.07 | |||||||||
| Cobitidae | Misgurnus anguillicaudatus * | Pond loach | MAN | Omnivore | Y | 0.07 | 0.13 | 0.08 | 0.86 | 0.62 | 0.06 | 0.16 | ||
| Synbranchidae | Monopterus albus * | Ricefield eel | MAL | Carnivore | Y | 0.00 | 0.01 | 0.00 | 0.01 | |||||
| Eleotridae | Micropercops swinhonis ξ | Swinhon’s sleeper | MSW | Omnivore | Y | 0.10 | 0.15 | 0.06 | 0.10 | 0.06 | 0.10 | 0.08 | ||
| Gobiidae | Rhinogobius cliffordpopei ξ | RCL | Carnivore | Y | 1.04 | 0.10 | 0.02 | 0.19 | 0.06 | 0.13 | 0.11 | 0.03 | 0.04 | |
| Rhinogobius giurinus ξ | Barcheek goby | RGI | Carnivore | Y | 9.36 | 0.88 | 0.14 | 1.71 | 0.51 | 1.17 | 0.95 | 0.25 | 0.34 | |
| Salangidae | Neosalanx taihuensis ξ | Icefish | NTAI | Zooplanktivore | Y | 0.27 | 7.80 | 0.03 | 0.08 | 0.63 | 0.67 | 0.21 | 0.00 | 0.52 |
| Channidae | Channa argus * | Snakeheaded fish | CCR | Carnivore | Y | 0.00 | 0.02 | 0.04 | 0.02 | 0.02 | 0.08 | 0.01 | 0.01 | |
| Poeciliidae | Gambusia affinis ξ | Mosquitofish | GAF | Carnivore | Y | 0.00 | 0.00 | |||||||
| Osmeridae | Hypomesus nipponensis ξ | Japanese smelt | HNI | Zooplanktivore | 0.00 | 0.01 | 0.46 | 1.32 | 1.86 | 4.41 | 4.70 | |||
| Belontiidae | Macropodus chinensis ξ | Roundtail paradise fish | MAO | Carnivore | Y | |||||||||
| Siluridae | Clarias gariepinus ξ | Catfish | CGA | Carnivore | 0.01 | |||||||||
| Silurus meridionalis Chen ξ | Southern catfish | SMC | Carnivore | 0.01 | 0.01 | 0.04 | ||||||||
| Bagridae | Pelteobagrus fulvidraco ξ | Yellow catfish | PFU | Carnivore | Y | 0.00 | 0.00 | 0.01 | 0.00 | 0.01 | 0.00 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Yin, C.; Chen, Y.; Guo, L.; Ni, L. Fish Assemblage Shift after Japanese Smelt (Hypomesus nipponensis McAllister, 1963) Invasion in Lake Erhai, a Subtropical Plateau Lake in China. Water 2021, 13, 1800. https://doi.org/10.3390/w13131800
AMA Style
Yin C, Chen Y, Guo L, Ni L. Fish Assemblage Shift after Japanese Smelt (Hypomesus nipponensis McAllister, 1963) Invasion in Lake Erhai, a Subtropical Plateau Lake in China. Water. 2021; 13(13):1800. https://doi.org/10.3390/w13131800
Chicago/Turabian StyleYin, Chengjie, Yushun Chen, Longgen Guo, and Leyi Ni. 2021. "Fish Assemblage Shift after Japanese Smelt (Hypomesus nipponensis McAllister, 1963) Invasion in Lake Erhai, a Subtropical Plateau Lake in China" Water 13, no. 13: 1800. https://doi.org/10.3390/w13131800
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.