
Advances in Egyptian Mediterranean Coast Climate Change Monitoring

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
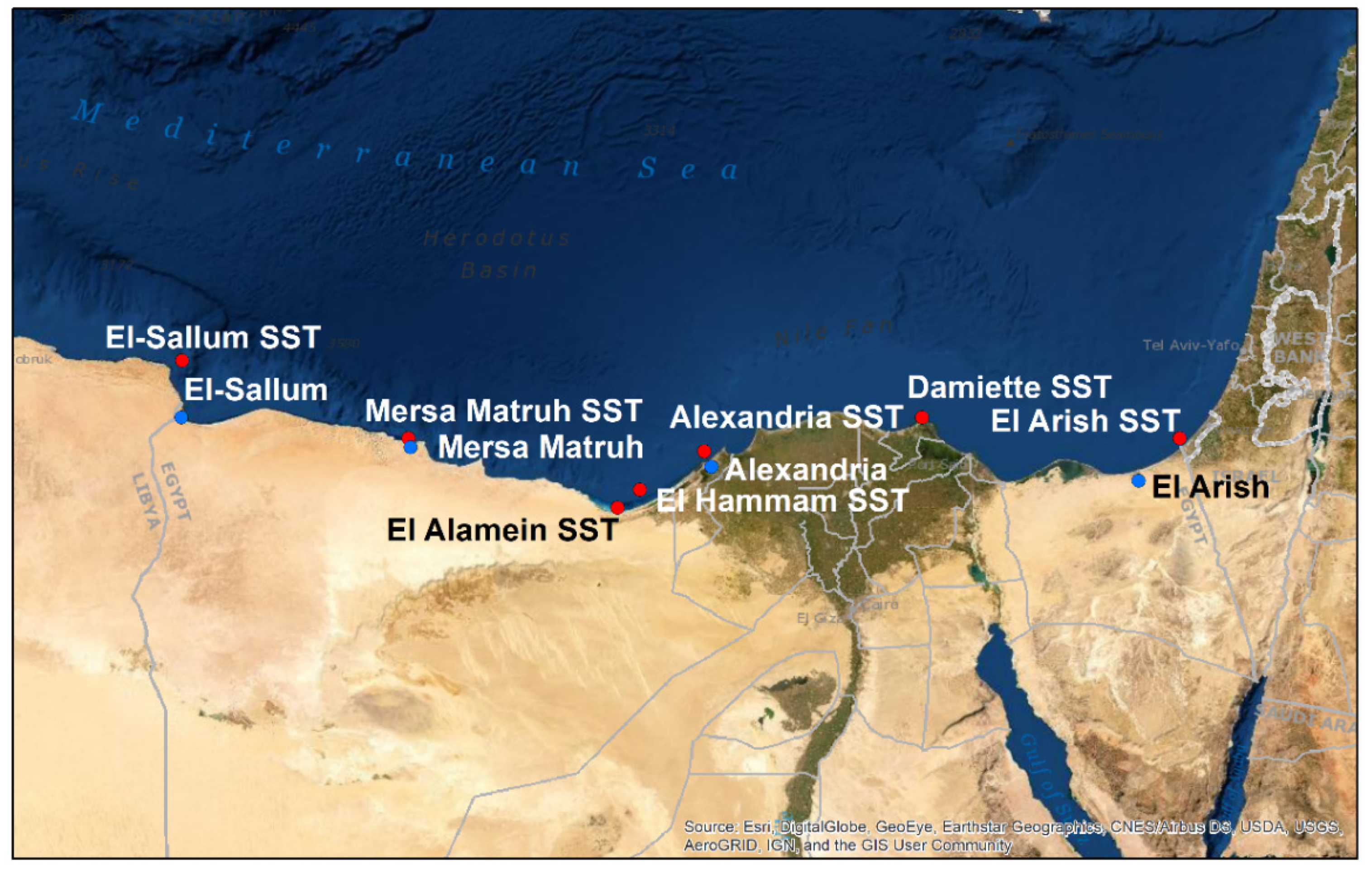
2.1. Egyptian Mediterranean Sites of Study
2.2. Biological Characterization of Invading Species in the Egyptian Mediterranean Coast (1999–2020)
2.3. Temperature Trends along the Egyptian Mediterranean Coast
3. Results
3.1. Analysis of the Biological Invasions along the Egyptian Mediterranean Coast (1999–2020)
3.1.1. Update on Non-Indigenous Species (NIS)
3.1.2. Biodata on Lessepsian Species
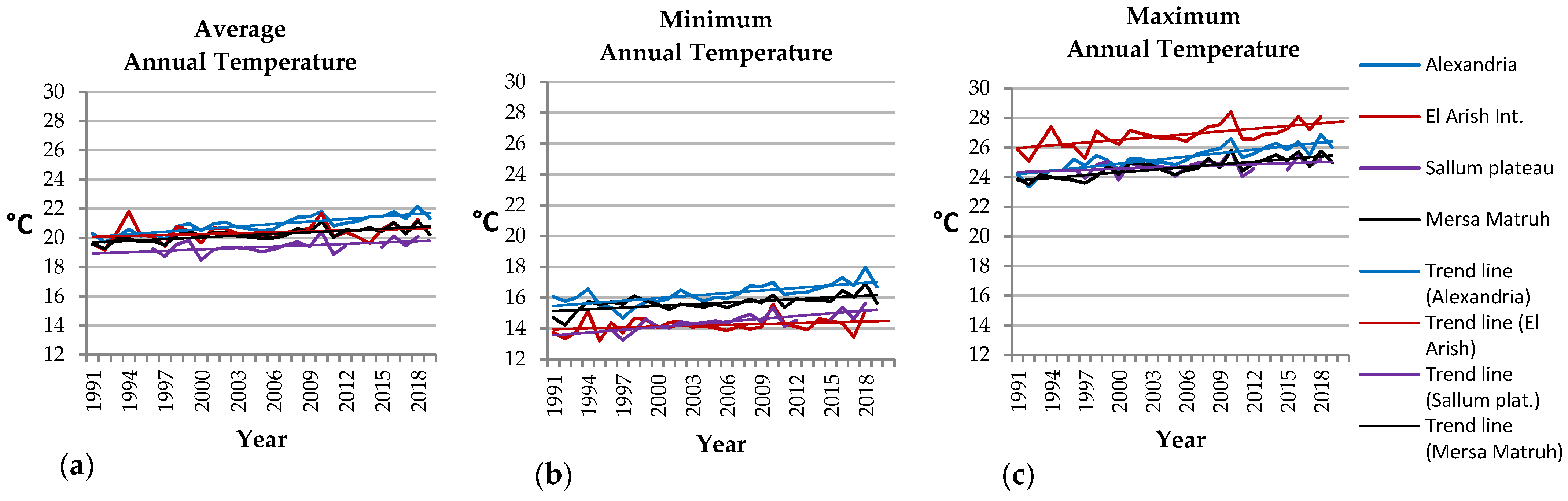
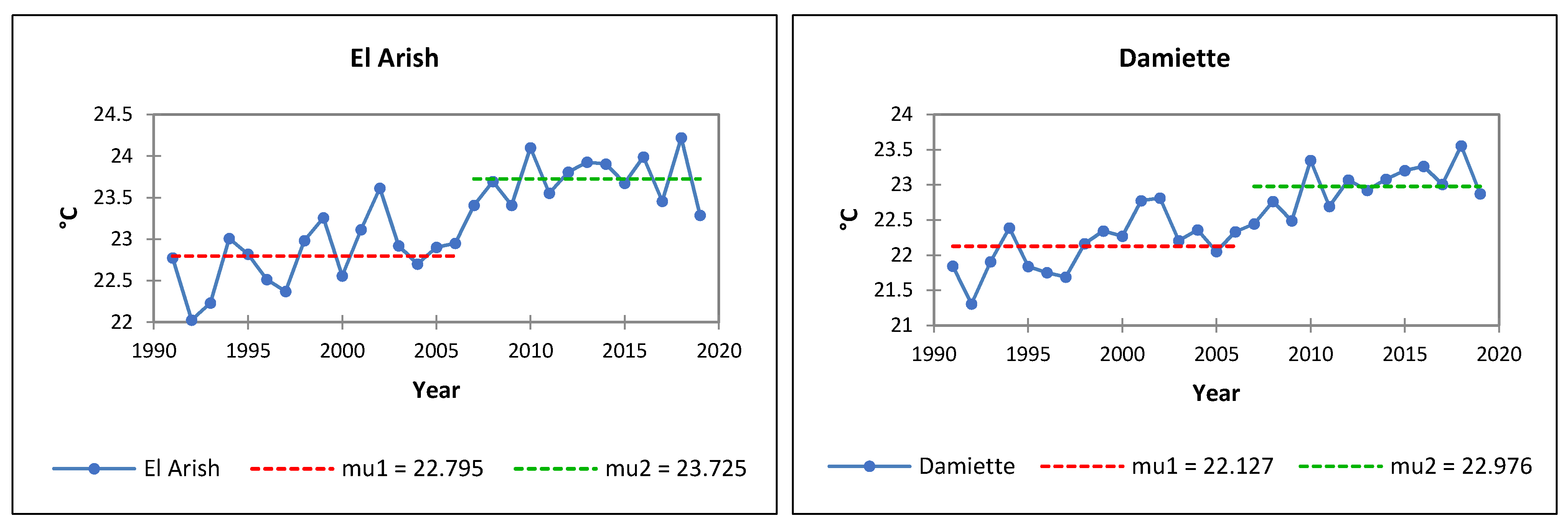
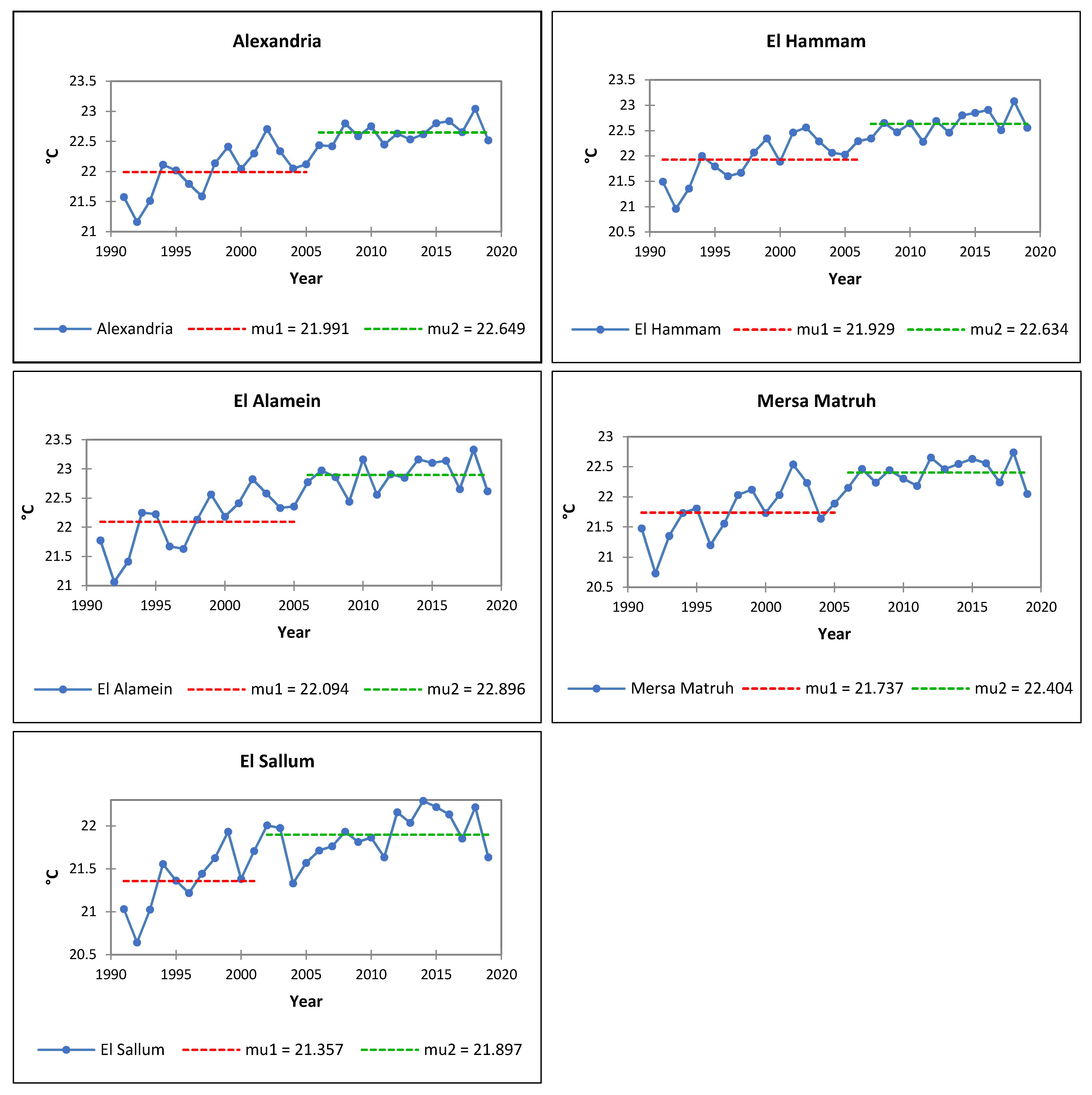
3.2. Analysis of Air Temperatures
- El-Arish: Ta +0.6 °C, Tm +0.5 °C, TM +1.8 °C;
- Alexandria: Ta +1.7 °C, Tm +1.6 °C, TM +2.2 °C;
- Mersa Matruh: Ta +1.1 °C, Tm +1.2 °C, TM +1.6 °C;
- El-Sallum: Ta +0.9 °C, Tm +1.6 °C, TM +0.9 °C.
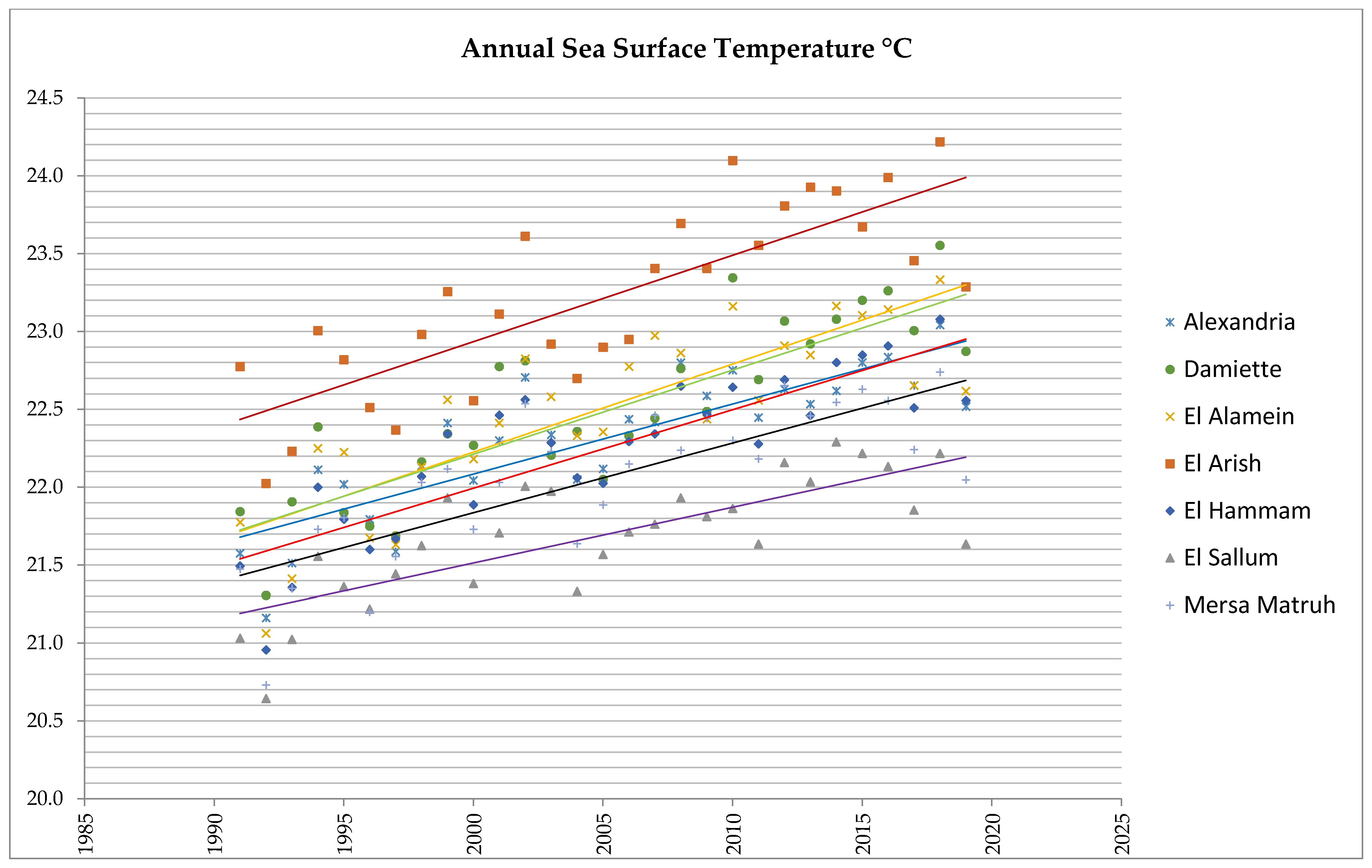
3.3. Analysis of Sea Surface Temperatures
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Roberts, N.; Jones, M.D.; Benkaddour, A.; Eastwood, W.J.; Filippi, M.L.; Frogley, M.R.; Zanchetta, G. Stable isotope records of Late Quaternary climate and hydrology from Mediterranean lakes: The ISOMED synthesis. Quat. Sci. Rev. 2008, 27, 2426–2441. [Google Scholar] [CrossRef]
- Gentilucci, M.; Barbieri, M.; D’Aprile, F.; Zardi, D. Analysis of extreme precipitation indices in the Marche region (central Italy), combined with the assessment of energy implications and hydrogeological risk. Energy Rep. 2020, 6, 804–810. [Google Scholar] [CrossRef]
- Ren, G.; Ding, Y.; Zhao, Z.; Zheng, J.; Wu, T.; Tang, G.; Xu, Y. Recent progress in studies of climate change in China. Adv. Atmos. Sci. 2012, 29, 958–977. [Google Scholar] [CrossRef]
- Cramer, W.; Guiot, J.; Fader, M.; Garrabou, J.; Gattuso, J.P.; Iglesias, A.; Lange, M.A.; Lionello, P.; Llasat, M.C.; Paz, S.; et al. Climate change and interconnected risks to sustainable development in the Mediterranean. Nat. Clim. Chang. 2018, 8, 972–980. [Google Scholar] [CrossRef] [Green Version]
- Georges, A.; Holleley, C.E. How does temperature determine sex? Science 2018, 360, 601–602. [Google Scholar] [CrossRef]
- Giangrande, A.; Pierri, C.; Del Pasqua, M.; Gravili, C.; Gambi, M.C.; Gravina, M.F. The Mediterranean in check: Biological invasions in a changing sea. Mar. Ecol. 2020, 41, e12583. [Google Scholar] [CrossRef]
- Gentilucci, M.; Materazzi, M.; Pambianchi, G.; Burt, P.; Guerriero, G. Temperature variations in Central Italy (Marche region) and effects on wine grape production. Theor. Appl. Climatol. 2020, 140, 303–312. [Google Scholar] [CrossRef]
- Visser, M.E.; Both, C. Shifts in phenology due to global climate change: The need for a yardstick. Proc. R. Soc. B Biol. Sci. 2005, 272, 2561–2569. [Google Scholar] [CrossRef]
- Seppälä, R. Adaptation of Forests and People to Climate Change: A Global Assessment Report; IUFRO: Vienna, Austria, 2009. [Google Scholar]
- Gentilucci, M.; Bisci, C.; Burt, P.; Fazzini, M.; Vaccaro, C. Interpolation of rainfall through polynomial regression in the Marche region (Central Italy). In The Annual International Conference on Geographic Information Science; Springer: Cham, Switzerland, 2018; pp. 55–73. [Google Scholar]
- Gentilucci, M.; Materazzi, M.; Pambianchi, G.; Burt, P.; Guerriero, G. Assessment of variations in the temperature-rainfall trend in the province of Macerata (Central Italy), comparing the last three climatological standard normals (1961–1990; 1971–2000; 1981–2010) for biosustainability studies. Environ. Process. 2019, 6, 391–412. [Google Scholar] [CrossRef]
- Mohammat, A.; Wang, X.; Xu, X.; Peng, L.; Yang, Y.; Zhang, X.; Myneni, R.B.; Piao, S. Drought and spring cooling induced recent decrease in vegetation growth in Inner Asia. Agric. For. Meteorol. 2013, 178, 21–30. [Google Scholar] [CrossRef]
- Maracchi, G.; Sirotenko, O.; Bindi, M. Impacts of present and future climate variability on agriculture and forestry in the temperate regions: Europe. Clim. Chang. 2005, 70, 117–135. [Google Scholar] [CrossRef]
- Harvell, C.D.; Mitchell, C.E.; Ward, J.R.; Altizer, S.; Dobson, A.P.; Ostfeld, R.S.; Samuel, M.D. Climate warming and disease risks for terrestrial and marine biota. Science 2002, 296, 2158–2162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Lierop, P.; Lindquist, E.; Sathyapala, S.; Franceschini, G. Global forest area disturbance from fire, insect pests, diseases and severe weather events. Forest Ecol. Manag. 2015, 352, 78–88. [Google Scholar] [CrossRef] [Green Version]
- Regev, Y.; Davidovich, N.; Berzak, R.; Lau, S.C.; Scheinin, A.P.; Tchernov, D.; Morick, D. Molecular Identification and Characterization of Vibrio Species and Mycobacterium Species in Wild and Cultured Marine Fish from the Eastern Mediterranean Sea. Microorganisms 2002, 8, 863. [Google Scholar] [CrossRef]
- Harrould-Kolieb, E.R. Framing ocean acidification in the context of the United Nations Framework Convention on climate change (UNFCCC) and Paris Agreement. Clim. Policy 2019, 19, 1225–1238. [Google Scholar] [CrossRef]
- Guerriero, G. Seasonal steroids variations and maturity stages in the female chub, Leuciscus cephalus L. (Pisces, Cyprinidae). Ital. J. Zool. 2007, 74, 317–324. [Google Scholar] [CrossRef]
- Cheng, C.; MacIntyre, L.; Abdelmohsen, U.R.; Horn, H.; Polymenakou, P.N.; Edrada-Ebel, R.; Hentschel, U. Biodiversity, anti-trypanosomal activity screening, and metabolomic profiling of actinomycetes isolated from Mediterranean sponges. PLoS ONE 2015, 10, e0138528. [Google Scholar] [CrossRef]
- Guerriero, G.; Bassem, S.M.; Khalil, W.K.; Temraz, T.A.; Ciarcia, G.; Abdel-Gawad, F.K. Temperature changes and marine fish species (Epinephelus coioides and Sparus aurata): Role of oxidative stress biomarkers in toxicological food studies. Emir. J. Food Agric. 2018, 30, 205–211. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Cadotte, M.W.; Chen, Y.; Fraser, L.H.; Zhang, Y.; Huang, F.; Luo, S.; Shi, N.; Loreau, M. Global evidence of positive biodiversity effects on spatial ecosystem stability in natural grasslands. Nat. Commun. 2019, 10, 3207. [Google Scholar] [CrossRef]
- Parisi, C.; Guerriero, G. Antioxidative defense and fertility rate in the assessment of reprotoxicity risk posed by global warming. Antioxidants 2019, 8, 622. [Google Scholar] [CrossRef] [Green Version]
- Bethoux, J.P.; Gentili, B. Functioning of the Mediterranean Sea: Past and present changes related to freshwater input and climate changes. J. Mar. Syst. 1999, 20, 33–47. [Google Scholar] [CrossRef]
- Bianchi, C.N.; Morri, C. Marine biodiversity of the Mediterranean Sea: Situation, problems and prospects for future research. Mar. Pollut. Bull. 2000, 40, 367–376. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Lejeusne, C.; Chevaldonné, P.; Pergent-Martini, C.; Boudouresque, C.F.; Pérez, T. Climate change effects on a miniature ocean: The highly diverse, highly impacted Mediterranean Sea. Trends Ecol. Evol. 2010, 25, 250–260. [Google Scholar] [CrossRef]
- Metaxas, D.A.; Bartzokas, A.; Vitsas, A. Temperature fluctuations in the Mediterranean area during the last 120 years. Int. J. Climatol. 1991, 11, 897–908. [Google Scholar] [CrossRef]
- Abudaya, M. Seasonal and spatial variation in Sea Surface Temperature in the South-East Mediterranean Sea. J. Environ. Earth Sci. 2013, 3, 42–52. [Google Scholar]
- Mohamed, B.; Abdallah, A.M.; El-Din, K.A.; Nagy, H.; Shaltout, M. Inter-annual variability and trends of sea level and sea surface temperature in the mediterranean sea over the last 25 years. Pure Appl. 2019, 176, 3787–3810. [Google Scholar]
- Kubin, E.; Poulain, P.M.; Mauri, E.; Menna, M.; Notarstefano, G. Levantine Intermediate and Levantine Deep Water Formation: An Argo Float Study from 2001 to 2017. Water 2019, 11, 1781. [Google Scholar] [CrossRef] [Green Version]
- Pisano, A.; Marullo, S.; Artale, V.; Falcini, F.; Yang, C.; Leonelli, F.E.; Santoleri, R.; Buongiorno Nardelli, B. New evidence of mediterranean climate change and variability from sea surface temperature observations. Remote Sens. 2020, 12, 132. [Google Scholar] [CrossRef] [Green Version]
- Galil, B.S. Alien species in the Mediterranean Sea—which, when, where, why? In Challenges to Marine Ecosystems; Springer: Dordrecht, The Netherlands, 2008; pp. 105–116. [Google Scholar]
- Galil, B.S.; Zenetos, A. A sea change—exotics in the Eastern Mediterranean Sea. In Invasive Aquatic Species of Europe. Distribution, Impacts and Management; Springer: Dordrecht, The Netherlands, 2002; pp. 325–336. [Google Scholar]
- Galil, B.S.; Marchini, A.; Occhipinti-Ambrogi, A. East is east and West is west? Management of marine bioinvasions in the Mediterranean Sea. Estuar. Coast. Shelf Sci. 2018, 201, 7–16. [Google Scholar] [CrossRef]
- Galil, B.S.; Marchini, A.; Occhipinti-Ambrogi, A. Mare Nostrum, mare quod invaditur—the history of bioinvasions in the mediterranean sea. In Histories of Bioinvasions in the Mediterranean; Springer: Cham, Switzerland, 2018; pp. 21–49. [Google Scholar]
- Yasuhara, M.; Danovaro, R. Temperature impacts on deep-sea biodiversity. Biol. Rev. 2016, 91, 275–287. [Google Scholar] [CrossRef]
- Galil, B.S.; Boero, F.; Campbell, M.L.; Carlton, J.T.; Cook, E.; Fraschetti, S.; Gollasch, S.; Hewitt, C.L.; Jelmert, A.; Macpherson, E.; et al. ‘Double trouble’: The expansion of the Suez Canal and marine bioinvasions in the Mediterranean Sea. Biol. Invasions 2015, 17, 973–976. [Google Scholar] [CrossRef] [Green Version]
- Azzurro, E.; Sbragaglia, V.; Cerri, J.; Bariche, M.; Bolognini, L.; Ben Souissi, J.; Busoni, G.; Coco, S.; Chryssanthi, A.; Fanelli, E.; et al. Climate change, biological invasions, and the shifting distribution of Mediterranean fishes: A large-scale survey based on local ecological knowledge. Glob. Chang. Biol. 2019, 25, 2779–2792. [Google Scholar] [CrossRef] [PubMed]
- Katsanevakis, S.; Poursanidis, D.; Hoffman, R.; Rizgalla, J.; Rothman, S.B.S.; Levitt-Barmats, Y.A.; Espinosa Torre, F. Unpublished Mediterranean records of marine alien and cryptogenic species. BioInvasions Rec. 2020, 9, 165–182. [Google Scholar] [CrossRef]
- Gentilucci, M.; Parisi, C.; Coppola, M.R.; Majdoubi, F.Z.; Madonna, A.; Guerriero, G. Influence of Mediterranean Sea Temperature Increase on Gaeta Gulf (Tyrrhenian Sea) Biodiversity. In Proceedings of the Zoological Society; Springer: Kolkata, India, 2021; pp. 1–3. [Google Scholar]
- Guerriero, G.; Di Finizio, A.; Ciarcia, G. Biological pollution: Molecular identification of non-native species in the Central Tyrrhenian Sea. Catrina Int. J. Environ. Sci. 2010, 5, 41–47. [Google Scholar]
- Guerriero, G.; Rabbito, D.; Alwany, M.A.; Madonna, A.; Temraz, T.A.; Sulaiman, O.O.; Bassem, S.M.; Trocchia, S.; Abdel-Gawad, F.K.; Ciarcia, G. Fisheries and biodiversity along Mediterranean Sea: Italian and Egyptian coast overview. EuroMediterr. J. Environ. Integr. 2017, 2, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Redolat, D.; Monjo, R.; Lopez-Bustins, J.A.; Martin-Vide, J. Upper-Level Mediterranean Oscillation index and seasonal variability of rainfall and temperature. Theor. Appl. Climatol. 2019, 135, 1059–1077. [Google Scholar] [CrossRef]
- Ruela, R.; Sousa, M.C.; deCastro, M.; Dias, J.M. Global and regional evolution of sea surface temperature under climate change. Glob. Planet. Chang. 2002, 190, 103190. [Google Scholar] [CrossRef]
- Mačić, V.; Albano, P.G.; Almpanidou, V.; Claudet, J.; Corrales, X.; Essl, F.; Evagelopoulos, A.; Giovos, L.; Jimenez, C.; Kark, S.; et al. Biological invasions in conservation planning: A global systematic review. Front. Mar. Sci. 2018, 5, 178. [Google Scholar] [CrossRef] [Green Version]
- Saygu, İ.; Heymans, J.J.; Fox, C.J.; Özbilgin, H.; Eryaşar, A.R.; Gökçe, G. The importance of alien species to the food web and bottom trawl fisheries of the Northeastern Mediterranean, a modelling approach. J. Mar. Syst. 2020, 202, 103253. [Google Scholar] [CrossRef]
- Trasforming Our World: The 2030 Agenda for Sustainable Development. Available online: https://sustainabledevelopment.un.org/post2015/transformingourworld (accessed on 16 March 2021).
- Bonan, G.B. Forests and climate change: Forcings, feedbacks, and the climate benefits of forests. Science 2008, 320, 1444–1449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouizgane, A.; Farid, S.; Majdoubi, F.Z.; Droussi, M.; Guerriero, G.; Hasnaoui, M. Assessment of climate change effects on predation activity and growth of largemouth bass, Micropterus salmoides (Lacepède, 1802) by water temperature variations. Emir. J. Food Agric. 2018, 30. [Google Scholar] [CrossRef]
- Van Rijn, I.; Kiflawi, M.; Belmaker, J. Alien species stabilize local fisheries catch in a highly invaded ecosystem. Can. J. Fish. Aquat. 2020, 77, 752–761. [Google Scholar] [CrossRef] [Green Version]
- Halim, Y.; Rizkalla, S. Aliens in Egyptian Mediterranean waters. A check-list of Erythrean fish with new records. Mediterr. Mar. Sci. 2011, 12, 479–490. [Google Scholar] [CrossRef] [Green Version]
- Farrag, M.; El-Haweet, A.; Akel, E.S.; Moustafa, M. Occurrence of puffer fishes (Tetraodontidae) in the eastern Mediterranean, Egyptian coast—Filling in the gap. BioInvasions Rec. 2016, 5, 47–54. [Google Scholar] [CrossRef]
- Akel, E.H.; Karachle, P. The Marine Ichthyofauna of Egypt. Egypt. J. Aquat. Biol. Fish. 2017, 21, 81–116. [Google Scholar] [CrossRef] [Green Version]
- Lipej, L.; Acevedo, I.; Akel, E.H.K.; Anastasopoulou, A.; Angelidis, A.; Azzurro, E.; Castriota, M.; Çelik, L.; Cilenti, F.; Crocetta, A.; et al. “New Mediterranean Biodiversity Records” (March 2017). Mediterr. Mar. Sci. 2017, 18, 179. [Google Scholar] [CrossRef] [Green Version]
- Osman, A.G.M.; Farrag, M.M.S.; El Sayed, H.K.; Moustafa, M.A. Feeding behavior of lessepsian fish Etrumeus teres (Dekay, 1842) from the Mediterranean Waters, Egypt. Egypt. J. Aquat. Res. 2013, 39, 275–282. [Google Scholar] [CrossRef] [Green Version]
- Akel, E.H.K.; Philips, A.E. Fisheries and biodiversity of the beach seine catch from the Eastern Harbor, Alexandria, Egypt. Egypt. J. Aquat. Res 2014, 40, 79–91. [Google Scholar] [CrossRef] [Green Version]
- Sahar, F.M.; Usama, M.M.; Eman, M.H. First occurrence of the Red Sea goatfish, Parupeneus forsskali (Fourmanoir Guz, 1976) in the coastal waters of Egyptian Mediterranean Sea. Int. J. Fish. Aquac. 2016, 8, 94–97. [Google Scholar] [CrossRef] [Green Version]
- Farrag, M.M.S.; El-Naggar, H.A.; Abou-Mahmoud, M.M.A.; Alabssawy, A.N.; Ahmed, H.O.; Abo-Taleb, H.A.; Kostas, K. Marine biodiversity patterns off Alexandria area, southeastern Mediterranean Sea, Egypt. Environ. Monit. Assess. 2019, 191, 367. [Google Scholar] [CrossRef]
- Ragheb, E.; Rizkalla, S.I. First record of the Fang tooth morayeel Enchelycore anatina (Lowe, 1838) and the diagnostic features of three fish species and a decapod crab from the southeastern Mediterranean Sea, off Alexandria (Egypt). Egypt. J. Aquat. Biol. Fish. 2020, 24, 639–653. [Google Scholar] [CrossRef] [Green Version]
- Sghaier, Y.R.; Zakhama-Sraieb, R.; Hmida, A.B.; Charfi, F. An inventory of non-indigenous species (NIS) inside and outside three tourist marinas from the southern Mediterranean coast. J. Black Sea Mediterr. Environ. 2019, 25, 29–48. [Google Scholar]
- Al Mabruk, S.A.A.; Rizgalla, J.; Giovos, I.; Bariche, M. Social media reveals the first records of the invasive lionfish Pterois miles (Bennett, 1828) and parrotfish Scarus ghobban Forsskål, 1775 from Egypt (Mediterranean Sea). BioInvasions Rec. 2020, 9. in press. Available online: https://www.researchgate.net/publication/341425846 (accessed on 2 March 2021). [CrossRef]
- Gentilucci, M.; Barbieri, M.; Burt, P.; D’Aprile, F. Preliminary data validation and reconstruction of temperature and precipitation in Central Italy. Geosciences 2018, 8, 202. [Google Scholar] [CrossRef] [Green Version]
- Hirsch, R.M.; Slack, J.R.; Smith, R.A. Techniques of trend analysis for monthly water quality data. Water Resour. Res. 1982, 18, 107–121. [Google Scholar] [CrossRef] [Green Version]
- Alexandersson, H.; Moberg, A. Homogenization of Swedish temperature data. Part I: Homogeneity test for linear trends. Int. J. Climatol. 1997, 17, 25–34. [Google Scholar] [CrossRef]
- Costa, A.C.M.; Negreiros, J.; Soares, A. Identification of inhomogeneities in precipitation time series using stochastic simulation. In geoENV VI–Geostatistics for Environmental Applications; Springer: Dordrecht, The Netherlands, 2008; pp. 275–282. [Google Scholar]
- Albouy, C.; Guilhaumon, F.; Leprieur, F.; Lasram, F.B.R.; Somot, S.; Aznar, R.; Velez, L.; Le Loc’h, F.; Mouillot, D. Projected climate change and the changing biogeography of coastal Mediterranean fishes. J. Biogeogr. 2013, 40, 534–547. [Google Scholar] [CrossRef]
- Schilling, J.; Hertig, E.; Tramblay, Y.; Scheffran, J. Climate change vulnerability, water resources and social implications in North Africa. Reg. Environ. Chang. 2020, 20, 15. [Google Scholar] [CrossRef] [Green Version]
- Rilov, G. Multi-species collapses at the warm edge of a warming sea. Sci. Rep. 2016, 6, 36897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stern, N.; Levitt, Y.; Galil, B.S.; Diamant, A.; Yokeş, M.B.; Goren, M. Distribution and population structure of the alien Indo-Pacific Randall’s threadfin bream Nemipterus randalli in the eastern Mediterranean Sea. J. Fish Biol. 2014, 85, 394–406. [Google Scholar] [CrossRef] [PubMed]
- Dikbas, F.; Firat, M.; Koc, A.C.; Güngör, M. Homogeneity test for Turkish temperature series. In Proceedings of the BALWOIS 2010, Ohrid, Republic of Macedonia, 25–29 May 2010; pp. 1–5. [Google Scholar]
- Ben Rais Lasram, F.; Tomasini, J.A.; Guilhaumon, F.; Romdhane, M.S.; Do Chi, T.; Mouillot, D. Ecological correlates of dispersal success of Lessepsian fishes. Mar. Ecol. Prog. Ser. 2008, 363, 273–286. [Google Scholar] [CrossRef] [Green Version]
- D’Amen, M.; Azzurro, E. Lessepsian fish invasion in Mediterranean marine protected areas: A risk assessment under climate change scenarios. ICES J. Mar. Sci. 2020, 77, 388–397. [Google Scholar] [CrossRef]
- Azzurro, E.; Soto, S.; Garofalo, G.; Maynou, F. Fistularia commersonii in the Mediterranean Sea: Invasion history and distribution modeling based on presence-only records. Biol. Invasions 2013, 15, 977–990. [Google Scholar] [CrossRef] [Green Version]
- Azzurro, E.; Franzitta, G.; Milazzo, M.; Bariche, M.; Fanelli, E. Abundance patterns at the invasion front: The case of Siganus luridus in Linosa (Strait of Sicily, Central Mediterranean Sea). Mar. Freshw. Res. 2017, 68, 697–702. [Google Scholar] [CrossRef]
- Coro, G.; Vilas, L.G.; Magliozzi, C.; Ellenbroek, A.; Scarponi, P.; Pagano, P. Forecasting the ongoing invasion of Lagocephalus sceleratus in the Mediterranean Sea. Ecol. Model. 2018, 371, 37–49. [Google Scholar] [CrossRef]
- Azzurro, E.; Tuset, V.M.; Lombarte, A.; Maynou, F.; Simberloff, D.; Rodríguez-Pérez, A.; Solé, R.V. External morphology explains the success of biological invasions. Ecol. Let. 2014, 17, 1455–1463. [Google Scholar] [CrossRef]
- Sih, A.; Bolnick, D.I.; Luttbeg, B.; Orrock, J.L.; Peacor, S.D.; Pintor, L.M.; Preisser, E.; Rehage, J.S.; Vonesh, J.R. Predator–prey naïveté, antipredator behavior, and the ecology of predator invasions. Oikos 2010, 119, 610–621. [Google Scholar] [CrossRef]
- Fanelli, E.; Azzurro, E.; Bariche, M.; Cartes, J.E.; Maynou, F. Depicting the novel Eastern Mediterranean food web: A stable isotopes study following Lessepsian fish invasion. Biol. Invasions 2015, 17, 2163–2178. [Google Scholar] [CrossRef]
- Giakoumi, S. Distribution patterns of the invasive herbivore Siganus luridus (Rüppell, 1829) and its relation to native benthic communities in the central Aegean Sea, Northeastern Mediterranean. Mar. Ecol. 2014, 35, 96–105. [Google Scholar] [CrossRef]
- Schlaepfer, M.A.; Sax, D.F.; Olden, J.D. The potential conservation value of non-native species. Conserv. Biol. 2011, 25, 428–437. [Google Scholar] [CrossRef] [PubMed]
- Henderson, P.A. The growth of tropical fishes. Fish Physiol. 2005, 21, 85–100. [Google Scholar]
- Goldberg, S.R. Notes on Reproduction of Hurter’s Spadefoot Toads, Scaphiopus hurterii (Anura: Scaphiopodidae), from Oklahoma. Chic. Herpetol. Soc. 2019, 54, 9–11. [Google Scholar]
- Poos, J.J.; Turenhout, M.N.; AE van Oostenbrugge, H.; Rijnsdorp, A.D. Adaptive response of beam trawl fishers to rising fuel cost. ICES J. Mar. Sci. 2013, 70, 675–684. [Google Scholar] [CrossRef]
- Guerriero, G. Non-native species and biodiversity assessment. J. Biodivers. Endanger. Species 2013, 1, e104. [Google Scholar] [CrossRef] [Green Version]
- Gentilucci, M.; Madonna, A.; Guerriero, G. The Influence of Sea Surface Temperatures on Biodiversity of Gaeta Gulf, Italy. In Recent Advances in Environmental Science from the Euro-Mediterranean and Surrounding Regions, 2nd ed.; Ksibi, M., Ed.; Springer: Cham, Switzerland, 2019. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Area | Coordinates | Year of Non-Indigenous Species (NIS) Fish Collection | Reference |
|---|---|---|---|
| From Al-Arish To El-Sallum | From 31°15′00″ N; 34°12′36″ E To 31°60′00″ N; 25°08′42″ E | Until 2010 | Halim and Rizkalla [51] |
| 2012–2015 | Farrag et al. [52] | ||
| Akel et al. [53] | |||
| Port Said | 31°15′12″ N; 32°18′28″ E | 2010–2012 | Katsanevakis et al. [39] |
| Damiette | 31°33′39″ N; 31°51′43″ E | 2016 | Lipej et al. [54] |
| Alexandria | From 31°00′ N; 29°18′ E To 31°36′ N; 30°05′ E | 2009 | Akel and Philips [56] |
| 31°19′29″ N; 30°3′11″ E | 2012–2014 | Katsanevakis et al. [39] | |
| 31°16′00″ N; 30°10′00″ E | 31 January 2016 | Sahar et al. [57] | |
| From 31°00′ N; 29°18′ E To 31°36′ N; 30°05′ E | 2017 | Farrag et al. [58] | |
| 31°13′1″ N; 29°53′1″ E | March–May 2018 | Ragheb and Rizkalla [59] | |
| From El Hamam (Alexandria) to Rashid (Alexandria) | 31°00′ N; 29°18′00″ E 31°35′00″ N; 30°30′00″ E | 2008 | Osman et al. [55] |
| El Alamain (Mersa Matrouh) | 30°50′1″ N; 28°59′5″ E | December 2013 | Sghaier et al. [60] |
| Mersa Matrouh | 31°22′23″ N; 27°12′13″ E | 2010–2019 | Katsanevakis et al. [39] |
| 2018 | Al Mabruk et al. [61] |
| Species | Taxonomy (Order Family) | Egg Type | Native Depth | Native T | COI GenBank ID | Value |
|---|---|---|---|---|---|---|
| (a) | ||||||
| Perciformes | ||||||
| Aulopareia unicolor | Gobiidae | n.d. | n.d. | t | n.d. | n.d. |
| Coryogalops ochetica | Gobiidae | b | n.d. | t | n.d. | n.c. |
| Pteragogus pelycus | Labridae | p | 50–100 m | st | KY176585.1 | n.c. |
| Equulites klunzingeri | Leiognathidae | p | 10–100 m | t | KM538355.1 | n.c. |
| Upeneus pori | Mullidae | p | 50–100 m | st | KM538636.1 | n.c. |
| Upeneus moluccensis | Mullidae | p | 0–50 m | st | JN313347.1 | c. |
| Parupeneus forsskali | Mullidae | n.d. | 1–45 m | st | LC543903.1 | n.c. |
| Nemipterus randalli | Nemipteridae | p-b | 20–450 m | t | KM538438.1 | c. (l.) |
| Nemipterus japonicus | Nemipteridae | p | 0–250 m | t | HQ561008.1 | c. |
| Pempheris rhomboidea | Pempheridae | n.d. | n.d. | t | KJ020198.1 | n.d. |
| Pempheris vanicolensis | Pempheridae | p | 0–25 m | t | MF123980.1 | n.c. |
| Scarus ghobban | Scaridae | p | 0–90 m | t | JQ350329.1 | n.c. |
| Scomberomorus commerson | Scombridae | p | 0–100 m | t | KY372120.1 | c. (h.) |
| Epinephelus malabricus | Serranidae | p | 0–150 m | t | MH707766.1 | c. |
| Siganus luridus | Siganidae | b | 50–100 m | st | MF409629.1 | c. |
| Siganus rivulatus | Siganidae | b | 0–100 m | st | MF124055.1 | c. |
| Sillago sihama | Sillaginidae | p | n.d | t | JN312946.1 | c. |
| Crenidens crenidens | Sparidae | p | n.d | st | JF493282.1 | c. |
| Sphyraena chrysotaenia | Sphyraenidae | p | 50 m | t | JF494556.1 | n.d. |
| Sphyraena flavicauda | Sphyraenidae | p | 0–25 m | t | MF124057.1 | c. |
| Terapon puta | Terapontidae | p | 0–50 m | t | KR861565.1 | n.c. |
| Pelates quadrilineatus | Terapontidae | p | n.d | st | JN313121.1 | c. (l.) |
| (b) | ||||||
| Beloniformes | ||||||
| Tylosurus choram | Belonidae | b | n.d. | t | MF124076.1 | c. |
| Parexocoetus mento | Exocoetidae | b | >20 m | t | HQ956399.1 | c. (l.) |
| Hemiramphus far | Hemiramphidae | b | 0–6 m | st | HQ564418.1 | c. |
| Clupeiformes | ||||||
| Herklotsichthys punctatus | Clupeidae | p | 0–50 m | st | KM538362.1 | c. (l.) |
| Dussumieria elopsoides | Dussumieriidae | p | 0–50 m | st | JF93400.1 | c. (l.) |
| Etrumeus teres | p | <50 m | st | JF952733.1 | n.c. | |
| Scorpaeniformes | ||||||
| Sebastapistes nuchalis | Scorpaenidae | n.d. | n.d. | t | n.d. | n.d. |
| Pterois miles | p | 0–85 m | t | MF124021.1 | c. | |
| Tetraodontiformes | ||||||
| Stephanolepis diaspros | Monacanthidae | p | 0–50 m | t | KM538599.1 | c. |
| Lagocephalus sceleratus | Tetraodontidae | b | 50–100 m | t | KM538364.1 | n.c. |
| Lagocephalus spadiceus | b | 50–100 m | st | JN312835.1 | n.c. | |
| Lagocephalus suezensis | b | 40 m | st | KM538405.1 | n.c. | |
| Kurtiformes | ||||||
| Apogonichthyoides pharaonic | Apogonidae | mb | 0–50 m | t | KM538173.1 | n.c. |
| Jaydia smithi | mb | 22–230 m | t | HQ956586.1 | n.c. | |
| (c) | ||||||
| Atheriniformes | ||||||
| Atherinomorus lacunosus | Atherinidae | b | 0–39 m | st | HQ956401.1 | c. |
| Aulopiformes | ||||||
| Saurida undosquamis | Synodontidae | p | 50–250 m | st | HQ956239.1 | c. |
| Beryciformes | ||||||
| Sargocentron rubrum | Holocentridae | p | 0–84 m | st | KJ202198.1 | c. (l.) |
| Callionymiformes | ||||||
| Callionymus filamentosus | Callionymidae | p | 0–350 m | st | n.d. | n.c. |
| Carangiformes | ||||||
| Alepes djedaba | Carangidae | p | <50 m | st | HQ560999.1 | c. |
| Gobiiformes | ||||||
| Oxyurichtys petersi | Oxudercidae | b | n.d. | t | n.d. | n.c. |
| Mugilifomes | ||||||
| Liza carinata | Mugilidae | p | n.d. | t | KX929970.1 | c. |
| Myliobatiformes | ||||||
| Himantura uarnak | Dasyatidae | b | 0–50 m | st | JX263360.1 | n.c. |
| Pleuronectiformes | ||||||
| Cynoglossus sinusarabici | Cynoglossidae | p | 0–25 m | t | KM538311.1 | n.c. |
| Scombriformes | ||||||
| Rastrelliger kanagurta | Scombridae | p | 20–90 m | t | KY372082.1 | c. (h.) |
| Syngnathiformes | ||||||
| Fistularia commersonii | Fistulariidae | p | 50–100 m | t | MK567525.1 | c. |
| El-Arish | Alexandria | Mersa Matruh | El-Sallum | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Month | Ta | TM | Tm | Ta | TM | Tm | Ta | TM | Tm | Ta | TM | Tm |
| January | 12.9 | 19.4 | 7.0 | 13.9 | 18.9 | 8.9 | 13.6 | 18.2 | 9.5 | 12.1 | 17.2 | 7.6 |
| February | 13.8 | 20.5 | 7.5 | 14.4 | 19.5 | 9.1 | 14.1 | 18.7 | 9.5 | 12.8 | 17.9 | 8.0 |
| March | 15.9 | 22.6 | 9.5 | 16.4 | 21.2 | 11.1 | 15.8 | 20.5 | 10.8 | 15.2 | 20.6 | 10.0 |
| April | 18.9 | 25.9 | 11.9 | 19.0 | 23.7 | 13.8 | 18.2 | 23.2 | 12.9 | 17.7 | 23.3 | 12.3 |
| May | 21.8 | 28.5 | 15.2 | 22.1 | 26.7 | 17.2 | 21.0 | 25.7 | 15.9 | 20.8 | 26.5 | 15.3 |
| June | 24.8 | 31.1 | 18.4 | 25.2 | 29.2 | 21.4 | 24.2 | 28.3 | 19.5 | 24.0 | 29.5 | 18.8 |
| July | 26.8 | 32.6 | 21.0 | 27.0 | 30.5 | 23.7 | 26.2 | 29.9 | 22.0 | 26.1 | 31.7 | 20.9 |
| August | 27.3 | 32.9 | 21.7 | 27.6 | 31.2 | 24.0 | 26.8 | 30.5 | 22.6 | 26.3 | 31.7 | 21.4 |
| September | 25.9 | 31.6 | 20.0 | 26.3 | 30.6 | 22.1 | 25.7 | 29.6 | 21.2 | 24.9 | 30.3 | 20.0 |
| October | 23.2 | 29.4 | 17.1 | 23.6 | 28.2 | 18.9 | 23.0 | 27.4 | 18.4 | 21.6 | 26.7 | 16.8 |
| November | 18.6 | 25.5 | 12.5 | 19.6 | 24.2 | 14.4 | 19.0 | 23.6 | 14.5 | 17.8 | 22.8 | 13.2 |
| December | 14.4 | 21.4 | 8.5 | 15.5 | 20.8 | 10.4 | 15.2 | 19.7 | 11.0 | 13.8 | 18.9 | 9.4 |
| Annual | 20.4 | 26.8 | 14.2 | 20.9 | 25.3 | 16.2 | 20.2 | 24.6 | 15.6 | 19.4 | 24.8 | 14.5 |
| El-Arish | Alexandria | Mersa Matruh | El-Sallum | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Parameter | Ta | TM | Tm | Ta | TM | Tm | Ta | TM | Tm | Ta | TM | Tm |
| Tau | 0.22 | 0.48 | 0.21 | 0.65 | 0.69 | 0.58 | 0.54 | 0.58 | 0.46 | 0.031 | 0.02 | 0.61 |
| S’ | 90 | 196 | 86 | 281 | 301 | 253 | 237 | 253 | 199 | 66 | 40 | 128 |
| p-value | 0.10 | <0.001 | 0.11 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.049 | 0.24 | <0.001 |
| alfa | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 |
| Sen | 0.03 | 0.05 | 0.02 | 0.06 | 0.08 | 0.05 | 0.04 | 0.06 | 0.03 | 0.03 | 0.02 | 0.06 |
| CI− | −0.27 | −0.22 | −0.20 | −0.12 | −0.14 | −0.19 | −0.12 | −0.17 | −0.12 | −0.24 | −0.30 | −0.15 |
| CI+ | 0.34 | 0.38 | 0.26 | 0.24 | 0.34 | 0.29 | 0.21 | 0.31 | 0.23 | 0.32 | 0.44 | 0.34 |
| El-Arish | Alexandria | Mersa Matruh | El-Sallum | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Parameter | Ta | TM | Tm | Ta | TM | Tm | Ta | TM | Tm | Ta | TM | Tm |
| Tau | 0.18 | 0.29 | 0.15 | 0.39 | 0.38 | 0.33 | 0.29 | 0.31 | 0.25 | 0.14 | 0.08 | 0.3 |
| S’ | 882 | 1412 | 752 | 2007 | 1998 | 1739 | 1519 | 1616 | 1282 | 386 | 218 | 833 |
| p-value | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.0017 | 0.08 | <0.001 |
| alfa | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 |
| El-Arish | Damiette | Alexandria | El-Hammam | El-Alamain | Mersa Matruh | El-Sallum | |
|---|---|---|---|---|---|---|---|
| Month | SST | SST | SST | SST | SST | SST | SST |
| January | 18.4 | 17.2 | 18.5 | 18.5 | 18.1 | 18.3 | 18.1 |
| February | 17.4 | 16.6 | 17.4 | 17.5 | 17.2 | 17.2 | 17.0 |
| March | 17.8 | 17.4 | 17.4 | 17.3 | 17.5 | 17.2 | 16.9 |
| April | 19.3 | 19.1 | 18.5 | 18.4 | 18.8 | 18.0 | 17.7 |
| May | 22.4 | 21.9 | 21.2 | 20.9 | 21.7 | 20.3 | 20.0 |
| June | 25.5 | 25.2 | 24.2 | 24.0 | 24.8 | 23.9 | 23.3 |
| July | 28.2 | 27.4 | 26.4 | 26.2 | 26.9 | 26.2 | 25.6 |
| August | 29.3 | 28.5 | 27.6 | 27.4 | 28.1 | 27.4 | 26.9 |
| September | 28.8 | 28.0 | 27.2 | 27.2 | 27.5 | 27.1 | 26.7 |
| October | 26.9 | 26.2 | 25.8 | 25.8 | 25.7 | 25.7 | 25.2 |
| November | 23.8 | 23.0 | 23.3 | 23.2 | 23.2 | 23.2 | 22.7 |
| December | 20.5 | 19.6 | 20.4 | 20.5 | 20.1 | 20.2 | 20.1 |
| Annual | 23.2 | 22.5 | 22.3 | 22.2 | 22.5 | 22.1 | 21.7 |
| El-Arish | Damiette | Alexandria | El-Hammam | El-Alamain | Mersa Matruh | El-Sallum | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Parameter | MK | MK* | MK | MK* | MK | MK* | MK | MK* | MK | MK* | MK | MK* | MK | MK* |
| Tau | 0.42 | 0.39 | 0.69 | 0.65 | 0.67 | 0.61 | 0.36 | |||||||
| S’ | 2013 | 1873 | 280 | 262 | 270 | 248 | 1759 | |||||||
| p-value | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | |||||||
| alfa | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | |||||||
| Sen | 0.043 | 0.052 | 0.058 | 0.057 | ||||||||||
| CI− | −0.08 | −0.12 | −0.10 | −0.15 | ||||||||||
| CI+ | 0.18 | 0.22 | 0.21 | 0.25 | ||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gentilucci, M.; Moustafa, A.A.; Abdel-Gawad, F.K.; Mansour, S.R.; Coppola, M.R.; Caserta, L.; Inglese, S.; Pambianchi, G.; Guerriero, G. Advances in Egyptian Mediterranean Coast Climate Change Monitoring. Water 2021, 13, 1870. https://doi.org/10.3390/w13131870
Gentilucci M, Moustafa AA, Abdel-Gawad FK, Mansour SR, Coppola MR, Caserta L, Inglese S, Pambianchi G, Guerriero G. Advances in Egyptian Mediterranean Coast Climate Change Monitoring. Water. 2021; 13(13):1870. https://doi.org/10.3390/w13131870
Chicago/Turabian StyleGentilucci, Matteo, Abdelraouf A. Moustafa, Fagr Kh. Abdel-Gawad, Samira R. Mansour, Maria Rosaria Coppola, Lidia Caserta, Sara Inglese, Gilberto Pambianchi, and Giulia Guerriero. 2021. "Advances in Egyptian Mediterranean Coast Climate Change Monitoring" Water 13, no. 13: 1870. https://doi.org/10.3390/w13131870