
Taxonomy and Physiology of Oxyrrhis marina and Oxyrrhis maritima in Korean Waters


1
Climate Change Research Laboratory, Advanced Institute of Convergence Technology, Suwon 16229, Korea
2
Environment and Resource Convergence Center, Advanced Institute of Convergence Technology, Suwon 16229, Korea
*
Authors to whom correspondence should be addressed.
Water 2021, 13(15), 2057; https://doi.org/10.3390/w13152057
Submission received: 10 June 2021
/
Revised: 12 July 2021
/
Accepted: 19 July 2021
/
Published: 28 July 2021
(This article belongs to the Special Issue Plankton Ecology in Shallow Coastal Waters)
Abstract
:The genus Oxyrrhis is a heterotrophic dinoflagellate found in diverse marine environments. Oxyrrhis spp. have received attention owing to their ecological and industrial importance, high lipid contents, and docosahexaenoic acid formation. To the best of our knowledge, contrary to O. marina, ecophysiological characterization studies on O. maritima have not yet been reported. Therefore, we investigated the taxonomy and ecophysiology of four strains of O. marina from coastal waters and two strains of O. maritima from the littoral tidepool waters of Korea. Based on phylogenetic trees constructed using internal transcribed spacer ribosomal DNA (ITS rDNA) and SSU rDNA of dinoflagellates, the clade of all four O. marina strains was divergent from that of the two O. maritima strains. We measured the growth rates of both species at various water temperatures (10–36 °C), salinities (5–90), and light intensities (0–100 µE·m−2·s−1). The lowest (O. marina and O. maritima: 10 °C) and highest temperatures (O. marina: <35 °C, O. maritima: >35 °C) revealed that O. maritima has more tolerance to high salinity. This study provides a basis for understanding the ecophysiology of O. marina and O. maritima and their population dynamics in marine ecosystems.
1. Introduction
Heterotrophic dinoflagellates (HTDs) are of great importance in the marine food web because they coexist as predators and prey for other species [1]. In this study, we focused on HTDs Oxyrrhis marina (O. marina) and Oxyrrhis maritima (O. maritima). There are advantages to using Oxyrrhis as a model for other HTD natural samples, to identify and manipulate in experiments, and hence, to study the model of evolution and exploitation of this species. Oxyrrhis is a widespread, free-living, and ecologically important HTD [2,3,4,5] that has been utilized as an important model organism for a broad range of studies [6,7,8].
The genus Oxyrrhis is commonly found in marine and brackish nearshore waters, tide pools, and salterns. However, it explicitly excluded dinokaryotes in common dinoflagellate classification schemes [9]. Unlike most dinoflagellates, which either have a longitudinal and a transverse flagellum emerging from the sulcus and the cingulum, respectively, or both flagella growing from the apical area (Prorocentrales) [4], Oxyrrhis have both flagella emerging from the ventral side. Therefore, the phylogenetic position of O. marina remains controversial. While some of the morphological and cytological studies support its basal position in the dinoflagellate classification of the higher ranks [10], others infer a highly derived position within the order of Gonyaulacales, and the majority of recent studies support O. marina as an ancestral dinoflagellate [11].
Morphological studies have raised disputes on whether the genus Oxyrrhis contains multiple species (O. marina, O. maritima van Meel 1969, O. phaeocysticola Scherffel, 1900, and O. tentaculifera Conrad, 1939) or only one species (O. marina) [4,12,13,14,15,16,17]. In the literature, the lengths of O. marina Dujardin (1841) and O. marina Kent (1880) are 28–51 µm, whereas that of O. maritima Van Meel (1969) is 16–24 µm (Table 1). Recently, molecular phylogenetic studies have favored the notion that two sibling species exist in this genus. Lowe et al. suggested that this genus consists of two species, O. marina and O. maritima, and each species consists of two clades [4].
However, the existing data are scattered, despite the increasing number of studies on O. marina [11]. A myriad of ecological, physiological, and genetic studies have been conducted without accurate identification of the species. Thus, we established four strains of O. marina and two strains of O. maritima by isolating and culturing from six different locations in the Korean environmental water samples. The morphological features, phylogeny, and nuclear ribosomal DNA were analyzed based on the six strains of Oxyrrhis spp.
The growth rate of heterotrophic protists is affected by various abiotic factors, such as salinity [18,19], temperature [20], and light intensity [21]. Thus, the physiological study of each species is a critical step in understanding the structure and function of the ocean ecosystem. There have been many studies on the effects of temperature, salinity, and light conditions on the growth rates of O. marina [22,23,24]. However, there have been no studies on the ecophysiological characterizations of O. maritima. Therefore, it is worthwhile to explore this topic. Moreover, the difference between the ecophysiological characterizations of these two Oxyrrhis spp. is poorly understood. Thus, we measured and compared the growth rates of O. marina and O. maritima.
In the present study, we measured the growth rates of O. marina and O. maritima at various water temperatures (10–36 °C), salinities (5–90), and light intensities (0–100 µE·m−2·s−1). In particular, we tested whether O. marina could grow in darkness when suitable prey was provided. Furthermore, we determined the lowest and highest temperatures and salinities at which O. marina and O. maritima can survive or grow. The results of the present study provide a basis for understanding the ecophysiology of O. marina and O. maritima and their population dynamics in marine ecosystems.
2. Materials and Methods
2.1. Collection, Isolation, and Culturing of Oxyrrhis spp.
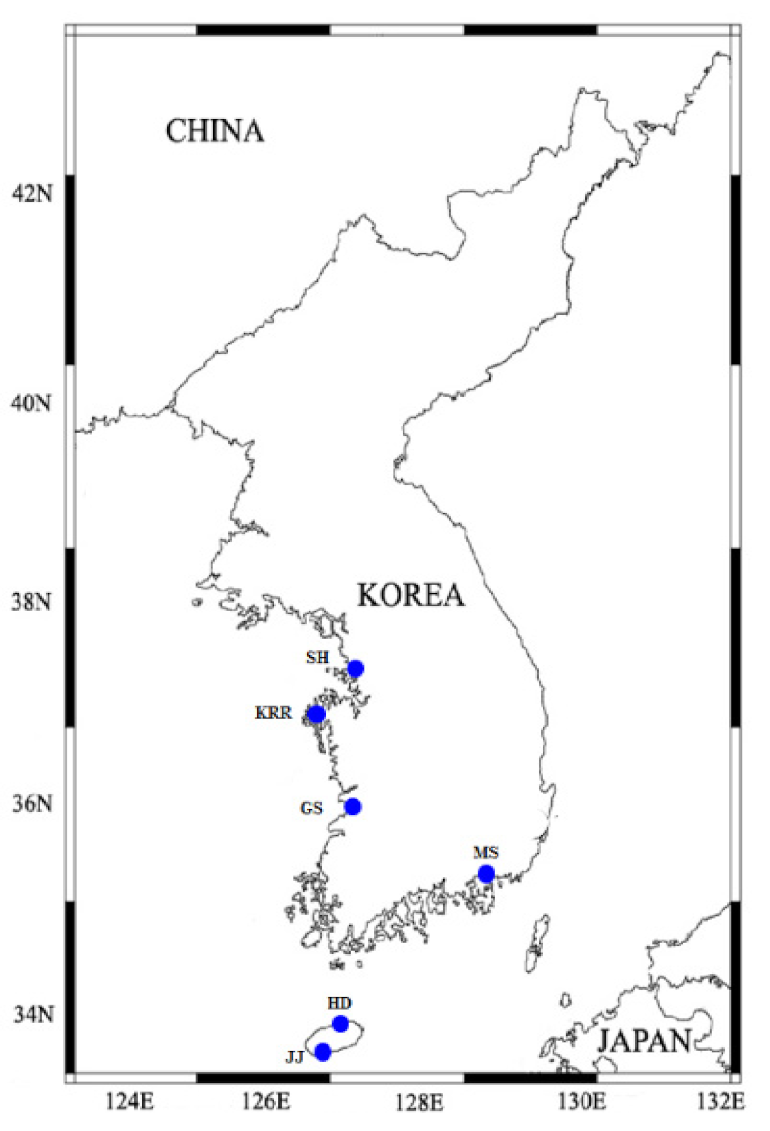
Water samples were collected using a syringe tubing and water sampling bucket from the surface of six different stations at littoral tide pools in Jeju Island and coastal waters in Korea (Shiwha, Gunsan, Masan, and Karorim; Table 2; Figure 1) between May 2001 and May 2013. Water temperature and salinity measured during sampling for O. marina were 13.5–19.7 °C and 27.7–33 °C, respectively, but no physical information was available in the case of O. maritima.
The sample was filtered through a 154-μm Nitex mesh screen and placed in six-well tissue culture plates. A clonal culture was established following two serial single-cell isolations, and the dried yeast (Saccharomyces cerevisiae) obtained from Red Star (Lesaffre Yeast Corporation, Milwaukee, WI, USA) were provided as food. As the culture volume increased, cells were transferred to 32-, 250-, and 500-mL polycarbonate (PC) bottles. The bottles were placed on a shelf at 20 °C under continuous illumination of 10 μE·m−2·s−1 provided by a cool white fluorescent light source. As the cultures became dense, approximately every 2–3 d, they were transferred to new 500-mL PC bottles containing dried yeast as prey.
2.2. Microscopic Observations
The morphologies of O. marina and O. maritima were examined using light microscopy. The length and width of live vegetative flagellated cells were measured using an image analysis system for processing images taken with a compound microscope (BX53 microscope, Olympus, Japan).
2.3. DNA Extraction, PCR Amplification, and Sequencing
Three to five cells of O. marina and O. maritima from a clonal culture were transferred to a 0.2-mL PCR tube containing 38.75 μL distilled water. To lyse the cell membranes before PCR analysis, the tube was frozen at −72 °C for 1–3 min and then thawed. The final mixture (50 μL) was vortexed, and PCR was performed in a thermal cycler (Eppendorf AG, Mastercycler® ep, model 5341, Hamburg, Germany). The final concentrations of PCR products were: 5 μL of 10X F-Star Taq buffer, 1 μL of 10 mM of dNTP mix, 0.25 μL of 5 U/μL BioFACT™ F-Star Taq DNA polymerase (BioFACT Co., Ltd., Daejeon, Korea), and 0.02 μM of each primer (Table S1) and a final volume of 50 μL was made. The PCR conditions were as follows: one activation step at 95 °C for 2 min, followed by 40 cycles at 95 °C for 20 s, the selected annealing temperature for 40 s, and 72 °C for 1 min, and a final elongation step at 72 °C for 5 min. The annealing temperature was adjusted depending on the primers used according to the manufacturer’s instructions. Positive and negative controls were used for all the amplification reactions. The purity of the amplicons was checked by performing electrophoresis of 5 μL of PCR products mixed with 1 μL of red fluorescent reagent (Redstar, Daejeon, Korea) using a 0.9% agarose gel at 80 V and observing it under a UV lamp to ensure that a single product was formed. Products containing a single band were then purified using an AccuPrep® PCR purification kit (Bioneer Corp., Daejeon, Korea) according to the manufacturer’s instructions. The purified amplicon was sent to Biomedic Corporation (Gyeonggi, Korea) for sequencing using an ABI PRISM® 3700 DNA Analyzer (Applied Biosystems, Foster City, CA, USA). Each portion of the target DNA was independently sequenced three times in both directions using primer pairs identical to those used for DNA amplification. These sequences were aligned using the ContigExpress alignment program (InforMax, Frederick, MD, USA) to remove low-quality regions and to assemble the individual sequence reads.
2.4. Phylogenetic Analysis
Phylogenetic analyses were performed based on alignments of the partial 3-end small subunit (SSU) and internal transcribed spacer region (ITS1-5.8S-ITS2) sequences. All sequence regions are located in the nuclear genome. Analyses included sequences obtained from GenBank and the present study (Table S2). Multiple sequences were aligned automatically using MEGA-X native implementation of ClustalW [25], and further aligned manually to refine the alignments. Two data sets were analyzed as follows: (1) SSU sequences only, (2) ITS1-5.8S-ITS2 sequences only. SSU sequences were used to investigate the systematic distance between species. Meanwhile, ITS1-5.8S-ITS2 sequences were used to determine intraspecific genetic diversity between Oxyrrhis species. The final alignment of the SSU rDNA contained 19 dinoflagellate species, plus Perkinsus, Apicomplexans, Ciliate, and Stramenopiles sequences that were used as outgroups. The final alignment of the ITS1-5.8S-ITS2 rDNA contained 33 Oxyrrhis sequences, plus Perkinsus, Apicomplexans, Ciliate, and Stramenopiles sequences that were used as outgroups. Two main model-based methods were used to infer phylogenetic relationships: maximum likelihood (ML) and Bayesian inference based on the likelihood function. ML analysis was performed with the RAxML 7.0.3 program [26] using the default GTR + G model. Tree likelihoods were estimated using a heuristic search with 200 random additional sequence replicates and tree bisection and reconnection branch swapping. ML bootstrapping with 1000 replications was also conducted. Bayesian analysis was performed with MrBayes v.3.1.2 [27,28] using the GTR + G + I model according to the model test results. The program was set to operate with a gamma distribution (+G) with five rate categories and by assuming that a certain fraction of sites is evolutionarily invariable(+I), and four Monte Carlo Markov chains (MCMC) starting from a random tree. In total, 2,000,000 generations were calculated with trees sampled every 100 generations and with a prior burn-in of 5000 generations.
2.5. Physiology
Experiments 1 and 2 were designed to investigate the effects of temperatures on the growth rates of O. marina (GS) and O. maritima (JJ) at 5, 10, 15, 20, 25, 30, 35, and 36 °C.
Each of the 5 L PC bottles containing O. marina or O. maritima was maintained at the target temperature and 20 μE·m−2·s−1 of continuous cool white fluorescent continuous light. O. marina and O. maritima cells were acclimated to the target temperatures, except <5 °C and >35 °C, for more than 10 d. For the target temperatures of <5 °C and >35 °C, cells acclimated at 10 °C and 30 °C for 10 d, respectively, were used without further acclimation. Dried yeast was provided as prey every 1–2 d. During this pre-incubation period, one-mL aliquots from each bottle were taken every day to determine the abundance of O. marina or O. maritima using a compound microscope.
In experiments 1 and 2, the initial abundances of O. marina and O. maritima (ca. 200 cell mL−1 at temperatures of 15, 20, 25, and 30 °C; 700 cells mL−1 at 10, 32, 35, and 36 °C) and the concentrations of autoclaved dried yeasts (ca. 20 mg per liter) were established using an auto-pipette to deliver predetermined volumes of known cell concentrations to the bottles.
Experiments 3 and 4 were designed to investigate the effects of salinity on the growth rates of O. marina and O. maritima. For the experiments, artificial seawater (S 9883 Sea salts, Sigma-Aldrich, St. Louis, MO, USA) or sterilized distilled water was added to sterilized seawater to obtain final salinities of 5, 10, 20, 33, 50, 70, and 90. Each bottle was placed on a shelf in a culture room maintained at 20 °C. The bottles were fed with dried yeasts as prey every 1–2 d. O. marina and O. maritima cells were acclimated to the target salinities, except <5 and >70, for more than 10 d. For target salinities <5 and >70, the cells acclimated at a salinity of 10 and 70 for 10 d, respectively, were used without further acclimation.
In experiments 3 and 4, the initial abundances of O. marina and O. maritima (ca. 300 cell mL−1 at salinities of 10, 20, 33, and 50; 500 cell mL−1 at salinities of 10, 70, and 90) and the concentrations of autoclaved dried yeast (ca. 20 mg dried yeast per liter) were established using an auto-pipette to deliver predetermined volumes of known cell concentrations to the bottles.
Experiments 5 and 6 were conducted to investigate the effects of light intensity on the growth rates of O. marina and O. maritima when fed with dried yeast (ca. 4 mg dried yeast per 200 mL) as prey. Dense cultures of O. marina and O. maritima were grown on dried yeast at 10 µE·m−2·s−1 of continuous cool white fluorescent light without any mixing. Bottles were wrapped with aluminum foil for darkness and placed on the shelf for each target light intensity of 0, 1, 20, or 100 µE·m−2·s−1. The experimental bottles were maintained at 20 °C, and every 1–2 d, the contents of each bottle were evenly distributed into two 1-L PC bottles containing the target amount of dried yeast and acclimated to their target light intensities for 10 d.
To determine the actual initial predator densities (cells mL−1) at the beginning of the experiment and after incubation, 5 mL aliquots were removed from each bottle and fixed with 5% Lugol’s solution, and all O. marina or O. maritima cells and (or >300) prey cells from the triplicate SRCs were enumerated. Before taking subsamples, the conditions of O. marina, O. maritima, and prey were assessed under a dissecting microscope.
The specific growth rate of O. marina or O. maritima, μ (d−1), was calculated as follows:
where H0 is the initial concentration (cells mL−1) of O. marina or O. maritima, and Ht is the final concentration after time t (d−1).
μ = ln (Ht/H0)/t,
3. Results
3.1. Morphology
3.1.1. Morphology of O. marina
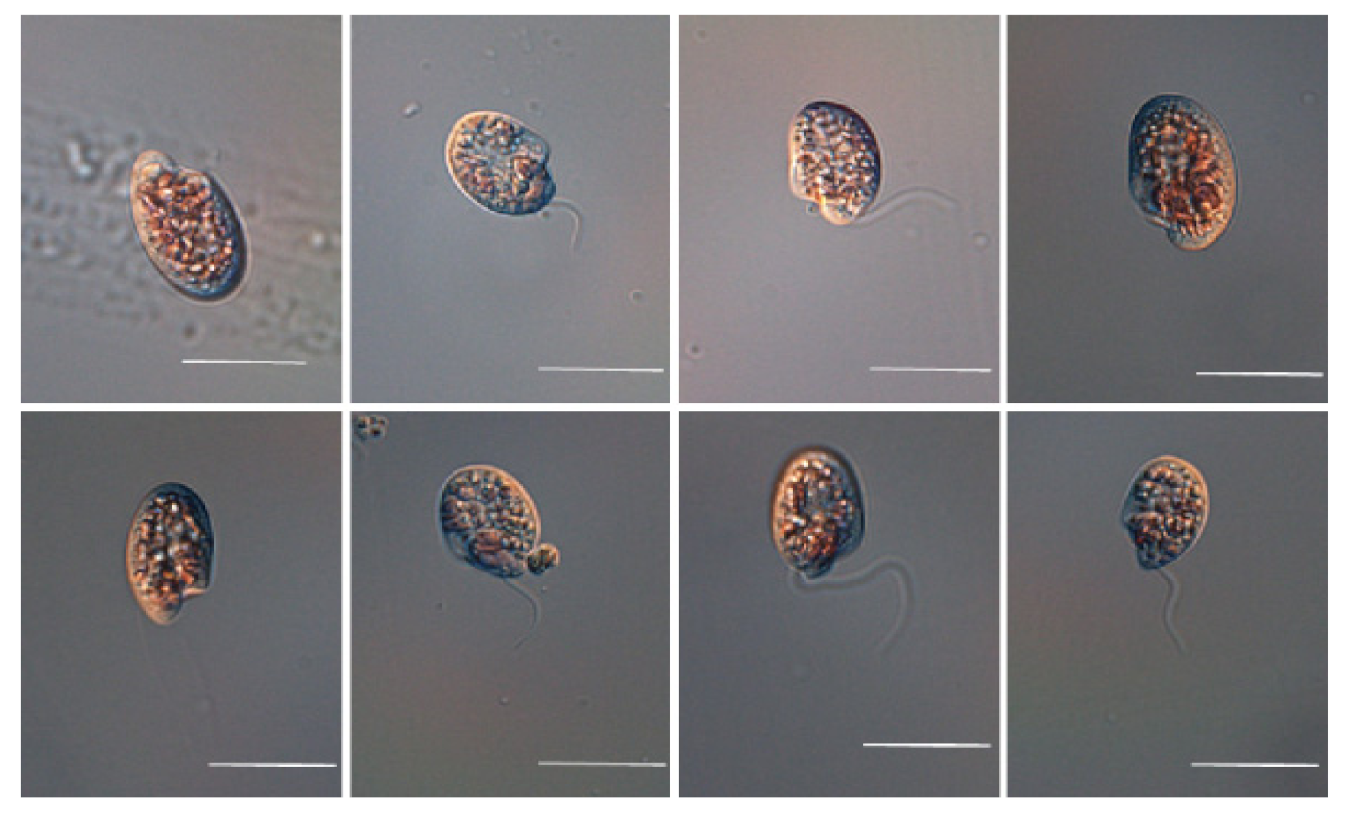
Under a light microscope (LM), the living cell was subovoid, asymmetrical posteriorly; girdle postmedial, not extending to the dorsal surface; sulcus spreading posteroventrally; flagella midventral; tentacular lobe situated between two flagella, dividing the broad undeveloped ventral sulcus (Figure 2). It appeared colorless, but had pink pigmentation, which was apparent in concentrated cultures. Therefore, this species is called a “pink pig” because of the pink colored water at a high concentration of the organisms [29].
The length and width of living cells of starved O. marina (n = 30) ranged 16.0–31.2 µm (mean ± SD = 23.6 ± 4.2 µm) and 8.0–19.2 µm (mean ± SD = 13.8 ± 2.4 µm), respectively, whereas live cells fed with Prorocentrum minimum (n = 15) measured 15.2–33.7 µm (28.4 ± 4.64 µm) and 15.6–24.0 µm (18.9 ± 3.7 µm) in length and width, respectively. Thus, the ratio of the cell length to width of living cells starved for 5 d (mean ± SD = 1.74 ± 0.23; range = 1.2–2.2; n = 30) was larger than that of cells fed with P. minimum (mean ± SD = 1.53 ± 0.17; range = 1.3–1.8; n = 15).
The mean equivalent spherical diameter (ESD, mean ± SD) was measured using an electronic particle counter (Coulter Multisizer II; Coulter Corp., Miami, FL, USA). The ESD of O. marina (OMGS 0105) starved for 5d (n > 2000) was 17.11–22.72 µm (19.85 ± 2.54 µm).
3.1.2. Morphology of O. maritima
As shown in Figure 3, under LM, living cells were observed to be subovoid, posteriorly asymmetrical; girdle postmedial, not extending to the dorsal surface; sulcus spreading posteroventrally; flagella midventral; tentacular lobe present between two flagella, dividing the broad undeveloped ventral sulcus. It appeared colorless but had pink pigmentation, which was apparent in concentrated cultures.
The length and width of living cells of O. maritima starved for 5 d (n = 30) ranged 14.4–28.0 µm (mean + SD = 20.6 ± 3.3 µm) and 9.6–15.2 µm (11.8 ± 1.9 µm), respectively, whereas live cells fed with P. minimum (n = 15) measured 16.8–35.2 µm (27.6 ± 5.1 µm) and 11.2–24.7 µm (19.9 ± 3.6 µm) in length and width, respectively. Thus, the ratio of length to width of living cells starved for 5 d (mean ± SD = 1.74 ± 0.23; range = 1.2–2.2; n = 30) was larger than that of the cells of O. maritima fed with P. minimum (mean ± SD = 1.53 ± 0.17; range = 1.3–1.8; n = 15). The ESD of O. maritima (OMJJ 1305) measured 11.4–15.1 µm (13.3 ± 1.5 µm), respectively. The ESD of O. maritima was approximately 45% smaller than that of O. marina.
3.2. DNA Sequence Analysis (Genetic Variability)
Nuclear sequences of SSU, ITS, and LSU rDNA were obtained from four strains of O. marina and two strains of O. maritima. The SSU, ITS 1 and 2, 5.8S, and LSU rDNA sequences of six strains of Oxyrrhis spp. from Gunsan (GS), Karorim (KRR), Masan (MS), Shiwha (SH), Jeju Anduck (JJ), and Jeju Hamduck (HD) comprised of 2882; 3010; 3033; 3006; 3051; and 3038 nucleotides, respectively.
Sequence alignments were performed using the MEGA-X native implementation of ClustalW. Sequence divergence was exceptionally high among some strains (Table 3). We compared the genetic divergence (%), except gaps, of the total sequences of the six strains of Oxyrrhis spp.
When aligned the sequence using the NCBI blast, the total rDNA sequence of O. maritima (JJ) differed greatly from those of O. marina isolated from KRR, SH, MS, and GS by 445, 494, 433, and 535 bp (except Gap), and a similarity of 82.2%, 81.2%, 82.1%, and 77.5%, respectively (corresponding to 17.8–22.5% dissimilarity).
The difference among the sequences of the four strains of O. marina was 0.1–0.3%, whereas that between the two strains of O. maritima was only 0.1%. However, the difference between the sequences of O. marina and O. maritima was 17.8–22.5%. In the phylogenetic trees based on the ITS1-5.8s-ITS2 rDNA of dinoflagellates, the clade of all four strains of O. marina belonged to lineage I (clades 1 and 2) and was clearly divergent from that of the two strains of O. maritima that belonged to lineage II (clade 4).
3.3. Phylogeny
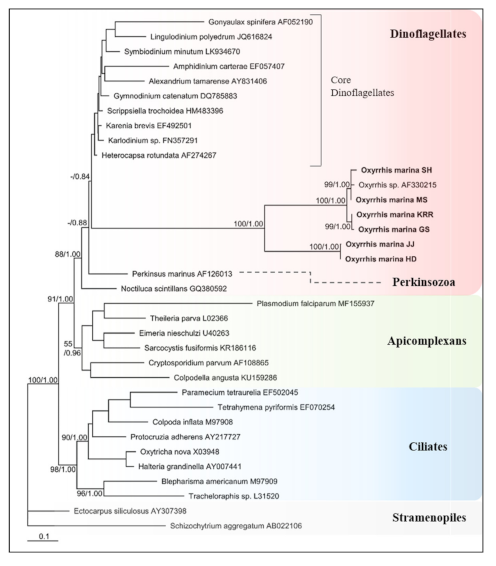
Figure 4 shows the phylogenetic trees constructed based on SSU rDNA sequences from nucleotide DNA to identify the phylogenetic position of Oxyrrhis spp. This confirmed that Oxyrrhis spp. were extremely divergent from core dinoflagellates, such as Alexandrium spp., and Amphidinium spp. (Figure 4). Oxyrrhis branched after Perkinsus marinus, which is between the dinoflagellates and Apicomplexans, with high bootstrap support in Bayesian analysis. Moreover, its branches proved that the Oxyrrhis was positioned at the base of dinoflagellate lineages. The branch length of SSU rDNA sequences of Oxyrrhis was approximately eight times as long as that of P. marinus. In all phylogenetic analyses, Oxyrrhis had two highly different species, and the four strains of O. marina and two strains of O. maritima branched together with high support. We used only Stramenopiles as outgroups, but Apicomplexans and Ciliates were apparently divergent with over 0.99 (Bayesian) probability.
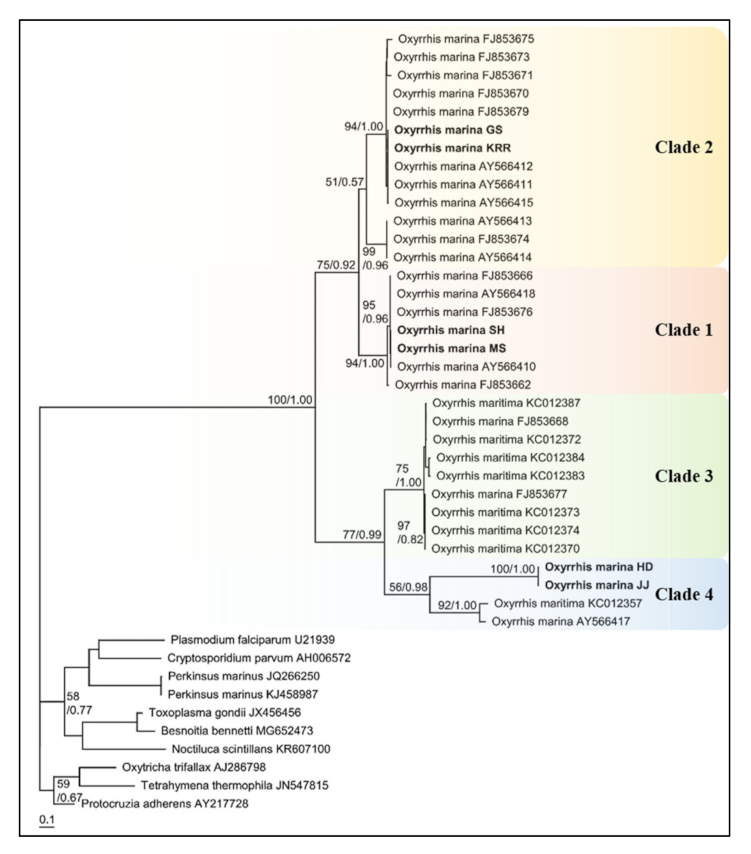
Furthermore, the phylogenetic tree based on ITS rDNA sequences proved the two highly divergent lineages within O. marina morphospecies (Figure 5). The two Oxyrrhis lineages divided four clades with high bootstrap support as follows: lineages I and II—1.00 (Bayesian)/100 (ML), clades 1 and 2—0.92 (Bayesian)/75 (ML), clades 3 and 4—0.99 (Bayesian)/77 (ML). According to the pairwise distance analysis, the net distance between clades 1 and 2 was 0.219, and this was 0.309 between clades 3 and 4. Moreover, the longest distance between clades was 0.456 between clades 1 and 3. Clades 1, 2, and 3 had a very short distance within each clade, but clade 4 had a slightly longer distance (0.32) than the others. Based on Lowe et al., clades 1 and 2 were classified as O. marina and clades 3 and 4 was O. maritima. The JJ strain formed a clade within the O. maritima HD, and both strains were closely related to O. maritima KC012357 and O. marina CCAP 1133/4, which were assigned to clade 4 in Lowe et al. In addition, other O. maritima species belonged to clade 3 together with two O. marina strains (FJ853668, FJ853677), and O. marina strains, except for these two strains, belonged to clades 1 and 2, respectively. In addition, four strains isolated from Korean coastal waters showed a closer relationship with each other. Thus, these two species are genetically different from each other, and the strains of Oxyrrhis in the Korean coastal waters are split into three clades.
3.4. Physiology
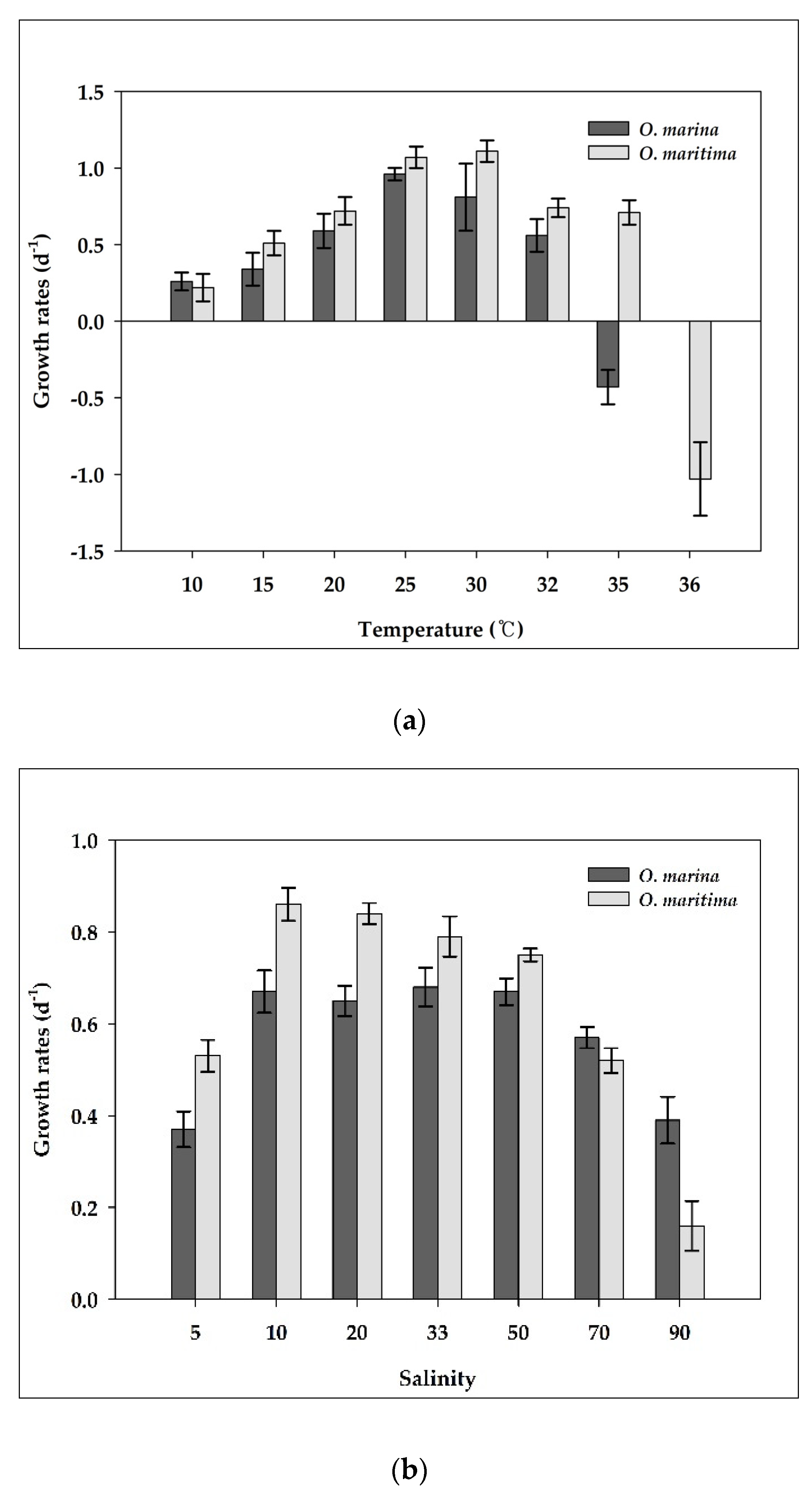
O. marina did not grow at temperatures >35 °C (Figure 6a). With increasing temperature from 10 to 25 °C, the growth rate of O. marina increased continuously from 0.26 d−1 at 10 °C to 0.96 d−1 at 25 °C, but it rapidly decreased between 30 and 35 °C (Figure 6a). However, O. maritima strains grew at temperatures >35 °C. Furthermore, the optimal temperature for supporting the maximum growth rate of O. marina was 25 °C, whereas that of O. maritima was 30 °C. With increasing temperature from 10 to 30 °C, the growth rate of O. maritima increased continuously from 0.22 d−1 at 10 °C to 1.11 d−1 at 30 °C, but it rapidly decreased between 32 and 36 °C. The growth rates of O. marina and O. maritima were significantly affected by temperature (ANOVA test, p < 0.05, Figure 6a). At 35 °C, the growth rate of O. maritima was significantly greater than that of O. marina (p < 0.05, one-tailed t-test). However, at temperatures of 10, 15, 20, 25, 30, and 32 °C, the growth rates of O. marina were not significantly different from those of O. maritima (p > 0.05, two-tailed t-test).
O. marina did not grow at salinities <4. With increasing salinity from 5–50, the growth rate of O. marina increased from 0.37 d−1 at 5 to 0.67 d−1 at 50, but it rapidly decreased between 50 and 90 (Figure 6b). However, O. maritima could grow at a salinity of 2. The growth rates of O. marina and O. maritima were significantly affected by salinity (ANOVA test, p < 0.05, Figure 6b). At salinities of 5, 10, and 20, the growth rates of O. maritima were significantly greater than those of O. marina (p < 0.05, one-tailed t-test), whereas at a salinity of 90, the growth rate of O. maritima was significantly lower than that of O. marina (p < 0.05, one-tailed t-test). However, at salinities of 33, 50, and 70, the growth rates of O. marina were not significantly different from those of O. maritima (p > 0.05, two-tailed t-test).
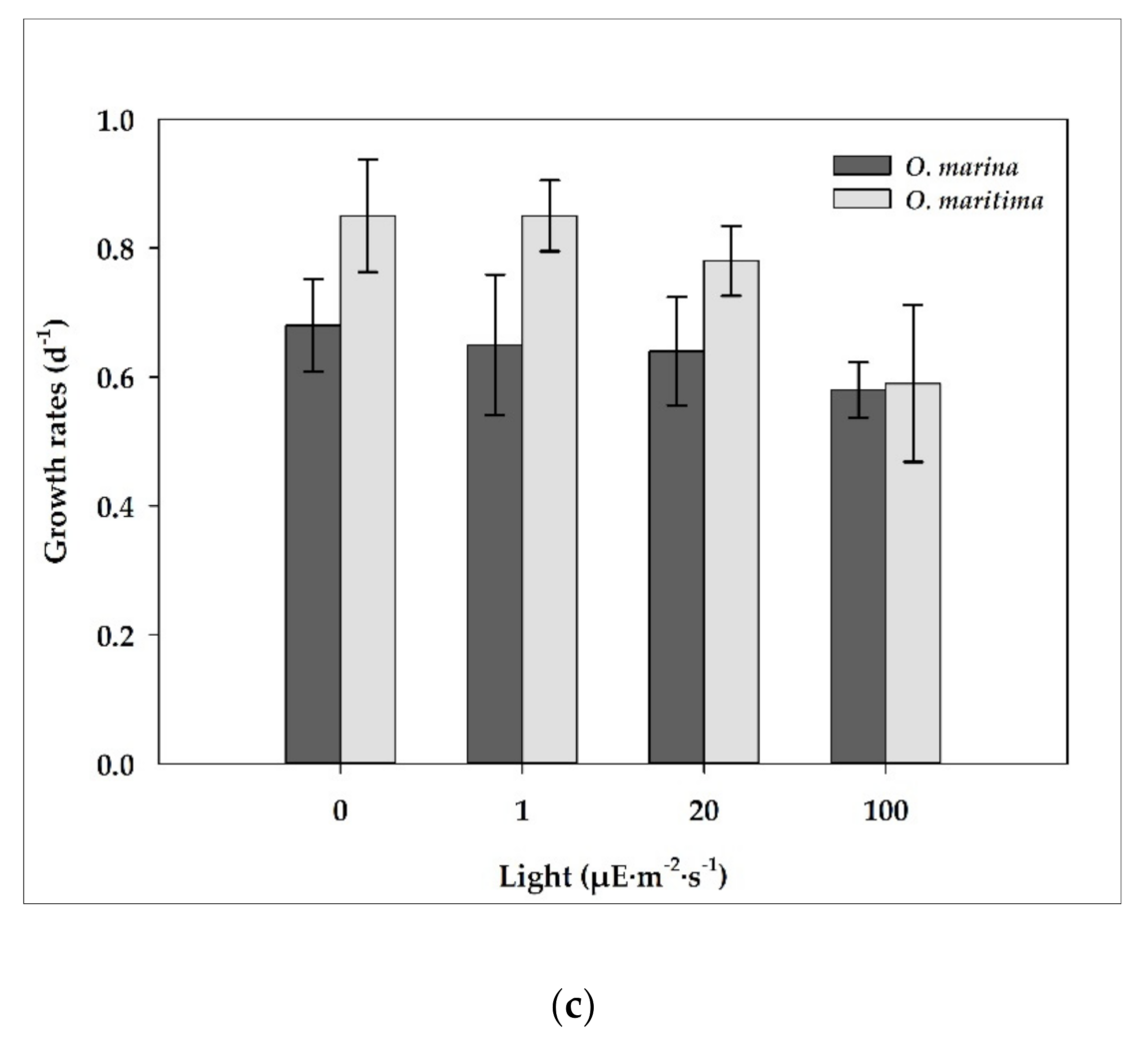
At the light intensities of 0–100 μE·m−2·s−1, the growth rates of dried yeast-fed O. marina did not largely change (i.e., 0.58 d−1 at 0 μE·m−2·s−1 to 0.59 d−1 at 100 μE·m−2·s−1). However, with increasing light intensity from 0 to 100 μE·m−2·s−1, the growth rate of O. maritima decreased from 0.85 d−1 at 0 μE·m−2·s−1 to 0.59 d−1 at 100 μE·m−2·s−1 (Figure 6c). The growth rates of O. marina and O. maritima were not affected by the light intensity (ANOVA test, p > 0.1, Figure 6c). In addition, the growth rate of O. marina was not significantly different from that of O. maritima (p > 0.1, two-tailed t-test).
4. Discussion
4.1. Comparison of Cell Size
In the literature, the lengths of O. marina Dujardin (1841) and O. marina Kent (1880) are 28–51 µm, whereas those of O. maritima Van Meel (1969) are 16–24 µm (Table 1). Some studies have reported that O. maritima could be distinguished from O. marina by its rounder and larger shape [4]. However, in contrast to previous studies, the species collected in this study showed that O. maritima was longer and sharper than O. marina. In the present study, the lengths and ESD of O. maritima were approximately 14% and 45% smaller than that of O. marina, respectively. Most of the studies have analyzed the genetic diversity of Oxyrrhis strains, either by providing only one genetic or morphological information about O. maritima, or based on data from strains collected long ago. The results of this study confirmed that the cell size of O. maritima is generally smaller than that of O. marina by direct analysis of the species collected within 10 years. Additionally, this study attempted to construct integrated data for species classification of Oxyrrhis strains by linking morphological and genetic information.
4.2. Phylogeny
The phylogenetic positions of O. marina are great of importance for understanding cell biology and evolutionary diversity. In particular, the relationships between the Apicomplexa and dinoflagellates have a great impact on disease-causing Apicomplexans and fundamental diversity in eukaryotic cell biology [30]. The dinoflagellate lineage includes abstruse organisms such as syndinians, noctilucoids, Pavilucifera, Perkinsus, and Oxyrrhis. These protists are highly related to Oxyrrhis and their phylogenetic positions are still a matter of debate [31]. Thus, our study conducted the phylogenetic analysis of Oxyrrhis using nuclear SSU rDNA with P. marinus, Noctiluca scintillans, the Apicomplexa (Colphodella), and Ciliates. The relationship with Amoebophrya spp. (Syndinians) was also considered (not shown data), but it was excluded since the sequence length was too long to align with others.
In this study, the phylogenetic tree based on nuclear SSU rDNA sequences was divided into Apicomplexa and dinoflagellates, and then branches of Noctiluca, Perkinsus, Oxyrrhis, and other dinoflagellates. The internal node, where Oxyrrhis and other core dinoflagellates were bound together, proved that Oxyrrhis occupies a basal position in the dinoflagellate lineage. Moreover, branches classified as Oxyrrhis JJ with HD, SH with MS, and GS with KRR prove that there is a big difference for each clade in the SSU sequence as well.
However, the bootstrapping number was low and the branch length of Oxyrrhis was irregularly long. This result is from the limit of the nuclear SSU rDNA. Ribosomal DNA has been utilized in myriad of studies since its advantage is the number of taxa or alignment depth, but it has limitations in that the alignment length is limited, and calculations become more difficult with insertions and deletions in the rDNA sequence. Moreover, some studies asserted that O. marina and N. scintillans can invalidate phylogeny because of their long branches [32,33]. To solve these limitations and determine the more accurate phylogenetic location of Oxyrrhis, more sequence information of taxa is needed, and this is not limited to nuclear rDNA, but also mitochondrial DNA or protein DNA information, which should be added to the concatenated alignment of the phylogenetic tree. However, in the case of mitochondrial DNA or protein DNA, information on various taxa is still lacking, and the degree of differences for each matrix is large, and thus, caution should be taken in the analysis.
Genetic differences between Oxyrrhis species can be seen more clearly in the phylogenetic tree based on the nuclear ITS rDNA sequence. According to Lowe et al., the SH and MS strains belonged to O. marina clade 1, KRR and GS strains belonged to O. marina clade 2, and two Jeju strains belonged to O. maritima clade 4 [34]. Lowe et al. conducted a phylogenetic analysis of 5.8S rDNA internal transcribed spacer (5.8S rDNA-ITS) and found two highly divergent lineages within O. marina morphospecies, thus proposing the existence of two Oxyrrhis spp.: O. marina and O. maritima. [34]. Based on ITS1-5.8S-ITS2 phylogenetic trees, O. marina from SH and MS strains were positioned together with O. marina (AY566418, UK) in clade 1, whereas O. marina from GS and KRR were positioned with O. marina (AY566415, USA) in clade 2. However, O. maritima from JJ and HD strains were present on the same branch as that of O. marina CCAP 1133/4(AY566417, UK), which showed high divergence from O. marina (clades 1 and 2). Moreover, both strains placed highly distinct from O. maritima in clade 3 [35]. This study confirmed that the two Oxyrrhis species are genetically distinct from each other. Furthermore, we suggest that some strains of O. marina (AY566417, UK/ FJ853668, UK/ FJ853677, GRE) mentioned in previous studies should be transferred to O. maritima.
The genetic diversity of O. maritima has been examined in various studies. Lowe et al. reported that O. marina was more widely distributed, from the Atlantic Ocean to the Mediterranean, than O. maritima that was partially distributed and elusive in the EU Ocean coasters [35]. Moreover, from previous reports, the genetic divergence in O. maritima was limited to clade 3 and clade 4 that contained only 2–3 haplotypes. In this study, however, three clades were discovered throughout the Korean coast. This difference in the dominant biological community might be due to the geographic characteristics of South Korea, which is adjacent to three seas with distinct ecological properties. Therefore, it is worth investigating South Korea for the genetic diversity of Oxyrrhis spp. Since O. marina is adaptable to various environments, we cannot explain its genetic diversity without analyzing the environmental conditions. Therefore, it is necessary to collect the strains of O. marina and O. maritima from more diverse environments for analyzing their morphological and genetic characteristics.
4.3. Physiology
The environmental factors affecting the growth of O. marina and O. maritima have been measured in the laboratory. O. marina did not grow at temperatures ≥35 °C, whereas O. maritima showed tolerance to high temperatures, which can explain its habitat. O. maritima was collected in tidal waters, which has a significant variation in temperature depending on direct sunlight or weather conditions until high tide.
The effects of salinity on the growth rate examined in this study were also correlated with their habitat. O. marina did not grow at a salinity below 4, whereas O. maritima species could grow at a salinity of 2. As mentioned above, tidal waters are greatly affected by environmental conditions, and heavy rainfalls can dramatically decrease the salinity close to that of freshwater.
The results indicate that O. maritima could survive in a wider range of environmental conditions than O. marina, and their ecological characteristics are distinct. In recent years, studies have investigated changes in species and habitats due to an increase in seawater temperature. The comparison of the optimal growth temperature and ecological characteristics of O. marina and O. maritima in this study can contribute to the accurate distinction between the two species.
Regarding Oxyrrhis strains used in the nuclear ITS sequence-based phylogenetic tree, there are no data on their collection areas and ecological characteristics, and thus it is difficult to make a comprehensive judgment on the species classification. As mentioned, the nuclear genome has several limitations such as sequence length. Thus, it should be supplemented by matching genetic information with ecological and morphological information. In this study, by systematically combining morphological information, ecological information, and genetic information, O. marina and O. maritima were accurately distinguished, and it was confirmed that they show distinct differences in all three aspects.
5. Conclusions
O. marina is a type species of the genus Oxyrrhis and has been used as a model species for a broad range of ecological, biological, and evolutionary studies. However, the taxonomy and classification of Oxyrrhis spp. are still vague and controversial, and the other species, O. maritima, have not been well investigated to date. For these reasons, we established four different strains of O. marina isolated from the coastal waters off GS, MS, SH, and KRR, Korea, and two strains of O. maritima isolated from littoral tide pool waters in Jeju Island, Korea, and confirmed that O. maritima from JJ and HD were highly divergent from O. marina (clades 1 and 2).
Environmental conditions are an important factor in determining the growth of O. marina and O. maritima. O. marina did not grow at water temperatures ≥ 35 °C, whereas O. maritima did. Thus, the highest temperature for positive growth of O. marina is likely to be approximately 32 °C, whereas that for O. maritima is ≥32 °C. O. marina did not grow at salinities below 4, and as salinity increased from 5 to 50, the growth rate increased continuously but rapidly decreased when salinity was over 70. However, O. maritima could grow at a salinity of 2, and its growth rate increased as salinity increased from 3 to 50, and rapidly decreased as salinity increased over 70. Thus, O. marina and O. maritima may have different ecophysiology, reflecting their habitats; O. maritima inhabits areas of higher salinity and temperatures (tidal pools) than O. marina (water columns).
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/w13152057/s1, Table S1: List of primer pairs used for amplification of SSU rDNA segments, ITS region, and LSU rDNA sequences, Table S2: List of species used in constructing nuclear SSU ribosomal (r) DNA phylogenetic trees. Genbank accession numbers are listed to the right of each species, Table S3: List of species used in constructing nuclear ITS1-5.8S-ITS2 phylogenetic trees. Genbank accession numbers are listed to the right of each species.
Author Contributions
Data analyses, methodology, writing—original draft preparation, M.K.J.; Culture collection, strain setup, culturing and microscopic analysis, T.Y.Y. and S.J.M.; Conceptualization—review and editing, J.P.; conceptualization, supervision, project administration, writing—review and editing, E.Y.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research was a part of the project titled “Development of eco-friendly functional strategic materials for improving blood circulation derived from marine microalgae (No. 20210515)” and “Improvement of management strategies on marine disturbing and harmful organisms (No. 20190518)” funded by the Ministry of Oceans and Fisheries, Korea.
Data Availability Statement
The data presented in this study are available on request from the corresponding authors.
Acknowledgments
We thank H. J. Jeong for comments on data analysis, J. H. Hyung and E. J. Kim for technical support and Y. D. Yoo for analysis of ESD.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Agmour, I.; Baba, N.; Bentounsi, M.; Achtaich, N. Mathematical study and optimal control of bioeconomic model concerning harmful dinoflagellate phytoplankton. Comp. Appl. Math. 2021, 40, 1–16. [Google Scholar] [CrossRef]
- Slamovits, C.H.; Keeling, P.J. Plastid-derived genes in the nonphotosynthetic alveolate Oxyrrhis marina. Mol. Biol. Evol. 2008, 25, 1297–1306. [Google Scholar] [CrossRef] [PubMed]
- Lowe, C.D.; Martin, L.E.; Roberts, E.C.; Watts, P.C.; Wootton, E.C.; Montagnes, D.J.S. Collection, isolation, and culturing strategies for Oxyrrhis marina. J. Plankton Res. 2011, 33, 569–578. [Google Scholar] [CrossRef] [Green Version]
- Lowe, C.D.; Keeling, P.J.; Laura, E.; Martin, L.E.; Slamovits, C.; Phillip, C.; Watts, P.C.; Montagnes, D.J.S. Who is Oxyrrhis marina? Morphological and phylogenetic studies on an unusual dinoflagellate. J. Plankton Res. 2011, 33, 555–567. [Google Scholar] [CrossRef] [Green Version]
- Watts, P.C.; Martin, L.E.; Kimmance, S.A.; Montagnes, D.J.; Lowe, C.D. The distribution of Oxyrrhis marina: A global disperser or poorly characterized endemic? J. Plankton Res. 2011, 33, 579–589. [Google Scholar] [CrossRef]
- Roberts, E.C.; Wootton, E.C.; Davidson, K.; Jeong, H.J.; Chris, D.; Lowe, C.D.; Montagnes, D.J.S. Feeding in the dinoflagellate Oxyrrhis marina: Linking behavior with mechanisms. J. Plankton Res. 2011, 33, 603–614. [Google Scholar] [CrossRef]
- Hartz, A.J.; Sherr, B.F.; Sherr, E.B. Photoresponse in the heterotrophic marine dinoflagellate Oxyrrhis marina. J. Eukaryot. Microbiol. 2011, 58, 171–177. [Google Scholar] [CrossRef]
- Yang, Z.; Jeong, H.J.; Montagnes, D.J.S. The role of Oxyrrhis marina as a model prey: Current work and future directions. J. Plankton Res. 2011, 33, 665–675. [Google Scholar] [CrossRef] [Green Version]
- Fensome, R.A. A classification of living and fossil dinoflagellates. Micropaleontology 1993, 7, 1–351. [Google Scholar]
- Hoppenrath, M. Dinoflagellate taxonomy—A review and proposal of a revised classification. Mar. Biodivers. 2017, 47, 381–403. [Google Scholar] [CrossRef]
- Guo, Z.; Zhang, H.; Liu, S.; Lin, S. Biology of the marine heterotrophic dinoflagellate Oxyrrhis marina: Current status and future directions. Microorganisms 2013, 1, 33–57. [Google Scholar] [CrossRef] [PubMed]
- Scheffel, A. Phaeocystis globosa nov. Spec. Nebst einigen betrachtungen Über die phylogenie niederer, insbesondere brauner organismen. In Wissenschaftliche Meeresuntersuchungen; Lipsius & Tischer: Abteilung Helgoland, German, 1900; Volume 4, pp. 1–29. (In Germany) [Google Scholar]
- Kofoid, C.A.; Swezy, O. The Free-Living Unarmored Dinoflagellate: Memoirs of the University of California; University of California Press: Berkeley, CA, USA, 1921; Volume 5, p. 538. [Google Scholar]
- Conrad, W. Notes protistologiques ix surtroisdinoflagellates de l’eausaumatre. Bull. Mus. Roy. Hist. Nat. Belg. 1939, 15, 1–10. [Google Scholar]
- Van Meel, L. Etudes hydrobiologiques des eaux saumâtres de belgique: 3. Les étangs galgenweelen à anvers (rive gauche). Bull. K. Belg. Inst. Nat. Wet. 1958, 34, 1–20. [Google Scholar]
- Dodge, J.D.; Hart-Jones, B. Marine Dinoflagellates of the British Isles; HMSO: London, UK, 1982.
- Cavalier-Smith, T.; Chao, E. Protalveolate phylogeny and systematics and the origins of Sporozoa and dinoflagellates (phylum Myzozoa nom. Nov.). Eur. J. Protistol. 2004, 40, 185–212. [Google Scholar] [CrossRef]
- Droop, M.R. A note on some physical conditions for cultivating Oxyrrhis marina. J. Mar. Biol. Assoc. UK 1959, 38, 599–604. [Google Scholar] [CrossRef] [Green Version]
- Lowe, C.D.; Day, A.; Kemp, S.J.; Montagnes, D.J. There are high levels of functional and genetic diversity in Oxyrrhis marina. J. Eukaryot. Microbiol. 2005, 52, 250–257. [Google Scholar] [CrossRef]
- Kimmance, S.; Atkinson, D.; Montagnes, D.J.S. Do temperature—food interactions matter? Responses of production and its components in the model heterotrophic flagellate Oxyrrhis marina. Aquat. Microb. Ecol. 2006, 42, 63–73. [Google Scholar] [CrossRef] [Green Version]
- Jakobsen, H.H.; Hansen, P.J.; Larsen, J. Growth and grazing responses of two chloroplast-retaining dinoflagellates: Effect of irradiance and prey species. MEPS 2000, 201, 121–128. [Google Scholar] [CrossRef] [Green Version]
- Li, A.; Stoecker, D.K.; Coats, D.W. Mixotrophy in Gyrodinium galatheanum (dinophyceae): Grazing responses to light intensity and inorganic nutrients. J. Phycol. 2000, 36, 33–45. [Google Scholar] [CrossRef]
- Montagnes, D.J.S.; Kimmance, S.A.; Atkinson, D. Using Q10: Can growth rates increase linearly with temperature? Aquat. Microb. Ecol. 2003, 32, 307–313. [Google Scholar] [CrossRef]
- Guerrini, F.; Ciminiello, P.; Aversano, C.D.; Tartaglione, L.; Fattorusso, E.; Boni, L.; Pistocchi, R. Influence of temperature, salinity and nutrient limitation on yessotoxin production and release by the dinoflagellate Protoceratium reticulatum in batch-cultures. Harmful Algae. 2007, 6, 707–717. [Google Scholar] [CrossRef]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef] [PubMed]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, B.; Gieskes, W.W.; Krray, G.G. Digestion of chlorophylls and carotenoids by the marine protozoan Oxyrrhis marina studied by HPLC analysis of algal pigments. J. Plankton Res. 1986, 8, 827–836. [Google Scholar] [CrossRef]
- Bachvaroff, T.R.; Handy, S.M.; Place, A.R.; Delwiche, C.F. Alveolate Phylogeny Inferred using Concatenated Ribosomal Proteins. J. Eukaryot. Microbiol. 2011, 58, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Claudio, H.S.; Patrick, J.K. Contributions of Oxyrrhis marina to molecular biology, genomics and organelle evolution of dinoflagellates, J. Plankton Res. 2011, 33, 591–602. [Google Scholar] [CrossRef] [Green Version]
- Hoppenrath, M.; Leander, B.S. Dinoflagellate phylogeny as inferred from heat shock protein 90 and ribosomal gene sequences. PLoS ONE 2010, 5, e13220. [Google Scholar] [CrossRef] [Green Version]
- Saldarriaga, J.F.; McEwan, M.L.; Fast, N.M.; Taylor, F.J.R.; Keeling, P.J. Multiple protein phylogenies show that Oxyrrhis marina and Perkinsus marinus are early branches of the dinoflagellate lineage. Int. J. Syst. Evol. Microbiol 2003, 53, 355–365. [Google Scholar] [CrossRef] [Green Version]
- Lowe, C.D.; Montagnes, D.J.S.; Martin, L.E.; Watts, P.C. Patterns of genetic diversity in the marine heterotrophic flagellate Oxyrrhis marina (alveolata: Dinophyceae). Protist 2010, 161, 212–221. [Google Scholar] [CrossRef] [PubMed]
- Lowe, C.D.; Martin, L.E.; Montagnes, D.J.; Watts, P.C. A legacy of contrasting spatial genetic structure on either side of the Atlantic-Mediterranean transition zone in a marine protest. Proc. Natl. Acad. Sci. USA 2012, 109, 20998–21003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Map of the sampling stations of Oxyrrhis marina (Shiwha [SH], Karorim [KRR], Gunsan [GS], Masan [MS], Korea) and O. maritima (Jeju-Anduck [JJ] and Hamduck [HD], which are located in Jeju Island, Korea).
Figure 1.
Map of the sampling stations of Oxyrrhis marina (Shiwha [SH], Karorim [KRR], Gunsan [GS], Masan [MS], Korea) and O. maritima (Jeju-Anduck [JJ] and Hamduck [HD], which are located in Jeju Island, Korea).

Figure 2.
Micrographs of Oxyrrhis marina (OMGS 0105) using light microscopy. Scale bars = 20 µm.

Figure 3.
Micrographs of Oxyrrhis maritima (OMJJ 305) using light microscopy. Scale bars = 20 µm.

Figure 4.
Consensus Maximum likelihood tree of nuclear small subunit (SSU) sequences for 6 strains of Oxyrrhis spp. with other Dinoflagellates, Perkinsozoa and Apicomplexans, Ciliates and outgroup Stramenopiles. The branch lengths are proportional to the number of base changes. The numbers near the branches indicate the Bayesian posterior probability (left) and Maximum likelihood bootstrap values (right). Posterior probabilities ≥0.5 are shown.
Figure 4.
Consensus Maximum likelihood tree of nuclear small subunit (SSU) sequences for 6 strains of Oxyrrhis spp. with other Dinoflagellates, Perkinsozoa and Apicomplexans, Ciliates and outgroup Stramenopiles. The branch lengths are proportional to the number of base changes. The numbers near the branches indicate the Bayesian posterior probability (left) and Maximum likelihood bootstrap values (right). Posterior probabilities ≥0.5 are shown.

Figure 5.
Consensus Maximum likelihood tree of nuclear internal transcribed spacer 1 (ITS1)-5.8S-ITS2 sequences for 6 strains of Oxyrrhis spp. with other dinoflagellates and outgroup, Perkinsus sp., Ciliates, and Apicomplexans. The branch lengths are proportional to the number of base changes. The numbers near the branches indicate the Bayesian posterior probability (left) and maximum likelihood bootstrap values (right). Posterior probabilities ≥0.5 are shown.
Figure 5.
Consensus Maximum likelihood tree of nuclear internal transcribed spacer 1 (ITS1)-5.8S-ITS2 sequences for 6 strains of Oxyrrhis spp. with other dinoflagellates and outgroup, Perkinsus sp., Ciliates, and Apicomplexans. The branch lengths are proportional to the number of base changes. The numbers near the branches indicate the Bayesian posterior probability (left) and maximum likelihood bootstrap values (right). Posterior probabilities ≥0.5 are shown.

Figure 6.
The growth rates (d−1) of O. marina (dark gray) and O. maritime (light gray) under various conditions; (a) Temperature; (b) Salinity; (c) Light. The error bar represents the standard deviation from the mean of triplicate data (n = 3).
Figure 6.
The growth rates (d−1) of O. marina (dark gray) and O. maritime (light gray) under various conditions; (a) Temperature; (b) Salinity; (c) Light. The error bar represents the standard deviation from the mean of triplicate data (n = 3).



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The designations, and taxonomic authorities, for species in the genus Oxyrrhis.
| Oxyrrhis | Year | Length (µm) | Flagella | Shape | Location |
|---|---|---|---|---|---|
| O. marina Dujardin | 1841 | 44 | Several | Oblong, oval bodied, rounded posteriorly | Mediterranean |
| O. marina Kent | 1880 | 28–51 | 2 | Body conical, sub-cylindrical, rounded posteriorly | St Helier, Jersey |
| O. marina (starved) | 2001 | 16–31 | 2 | Oblong, oval bodied, rounded posteriorly | Gunsan, Korea |
| O. phaeocysticola | 1900 | 20 | 2 | Rounded posterior, pointed anterior, excavated oral region with trunk-like projection | Helgoland, Germany |
| O. tentaculifera Conrad | 1939 | 38 | 2 | More voluminous than O. marina | Belgium |
| O. maritima Van Meel | 1969 | 16–24 | 2 | Twice as long as wide, has a tentacle | Belgium |
| O. maritima (starved) | 2013 | 15–28 | 2 | Body conical, sub-cylindrical, rounded posteriorly | Jeju, Korea |
Table 2.
Environmental conditions of collected Oxyrrhis spp. in the coastal waters of Korea.
| Location | Date | GPS | Temperature (°C) | Salinity | Habitats | Strain Name |
|---|---|---|---|---|---|---|
| Shiwha (SH) | November 2011 | 37°18′00.1″ N 126°40′19.5″ E | 19.7 | 31.0 | Coastal | OMSH_1111 |
| Karorim (KRR) | May 2010 | 36°45′19.1″ N 126°11′28.0″ E | 19.5 | 33.0 | Saltern | OMKRR_1005 |
| Gunsan (GS) | May 2001 | 35°56′20.1″ N 126°31′50.5″ E | 16.0 | 27.7 | Coastal | OMGS_0105 |
| Masan (MS) | April 2010 | 35°11′41.2″ N 128°34′23.3″ E | 13.5 | 29.8 | Coastal | OMMS_1004 |
| Jeju Island (JJ) | May 2013 | 33°12′47.7″ N 126°17′40.6″ E | - | - | Tide pool | OMJJ_1305 |
| Jeju Island (HD) | May 2013 | 33°32′33.6″ N 126°40′10.7″ E | - | - | Tide pool | OMHD_1305 |
Table 3.
Comparison of the total sequences of O. marina (KRR, SH, MS, GS) and O. maritima (JJ and HD). The numbers are base pairs different from each other. The numbers in parenthesis are similar (%) except gaps.
Table 3.
Comparison of the total sequences of O. marina (KRR, SH, MS, GS) and O. maritima (JJ and HD). The numbers are base pairs different from each other. The numbers in parenthesis are similar (%) except gaps.
| Species | O. marina | O. maritima | ||||
|---|---|---|---|---|---|---|
| Location | KRR | SH | MS | GS | JJ | HD |
| KRR | 0 (100%) | |||||
| SH | 215 (92.9%) | 0 (100%) | ||||
| MS | 202 (93.3%) | 2 (99.9%) | 0 (100%) | |||
| GS | 4 (99.9%) | 211 (92.7%) | 201 (93.1%) | 0 (100%) | ||
| JJ | 445 (82.2%) | 494 (81.2%) | 433 (82.1%) | 535 (77.5%) | 0 (100%) | |
| HD | 427 (83.0%) | 492 (81.3%) | 430 (82.3%) | 421 (82.3%) | 4 (99.9%) | 0 (100%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Jung, M.K.; Yin, T.Y.; Moon, S.J.; Park, J.; Yoon, E.Y. Taxonomy and Physiology of Oxyrrhis marina and Oxyrrhis maritima in Korean Waters. Water 2021, 13, 2057. https://doi.org/10.3390/w13152057
AMA Style
Jung MK, Yin TY, Moon SJ, Park J, Yoon EY. Taxonomy and Physiology of Oxyrrhis marina and Oxyrrhis maritima in Korean Waters. Water. 2021; 13(15):2057. https://doi.org/10.3390/w13152057
Chicago/Turabian StyleJung, Min Kyoung, Tae Yeon Yin, Seung Joo Moon, Jaeyeon Park, and Eun Young Yoon. 2021. "Taxonomy and Physiology of Oxyrrhis marina and Oxyrrhis maritima in Korean Waters" Water 13, no. 15: 2057. https://doi.org/10.3390/w13152057
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.