
Linking Hydrogeology and Ecology in Karst Landscapes: The Response of Epigean and Obligate Groundwater Copepods (Crustacea: Copepoda)

 , , and
, , and
Abstract
:1. Introduction
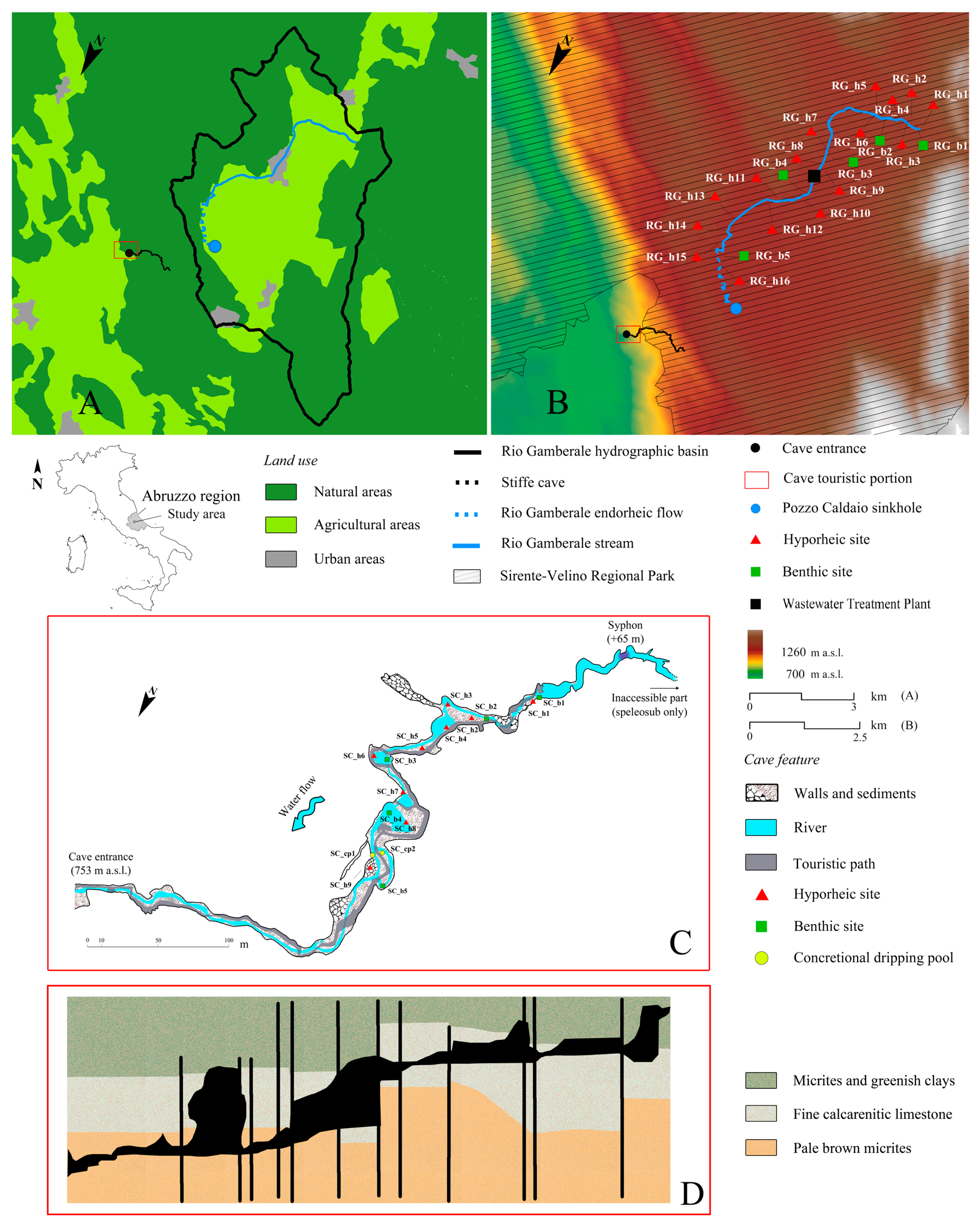
2. Study Area
3. Materials and Methods
3.1. Sampling Survey
3.2. Data Analysis
3.2.1. Environmental Variables
3.2.2. Biological Variables
4. Results
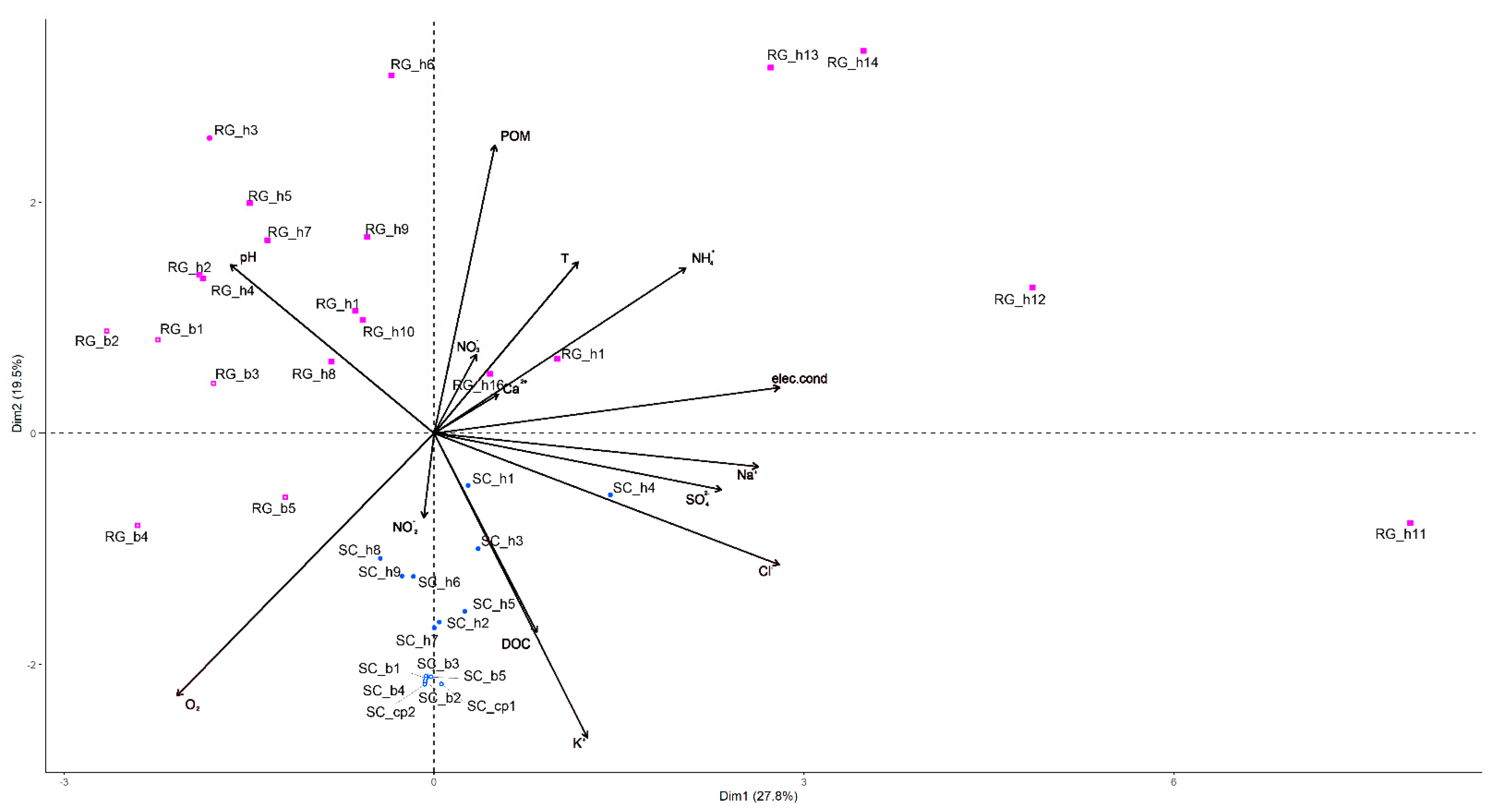
4.1. Environmental Variables
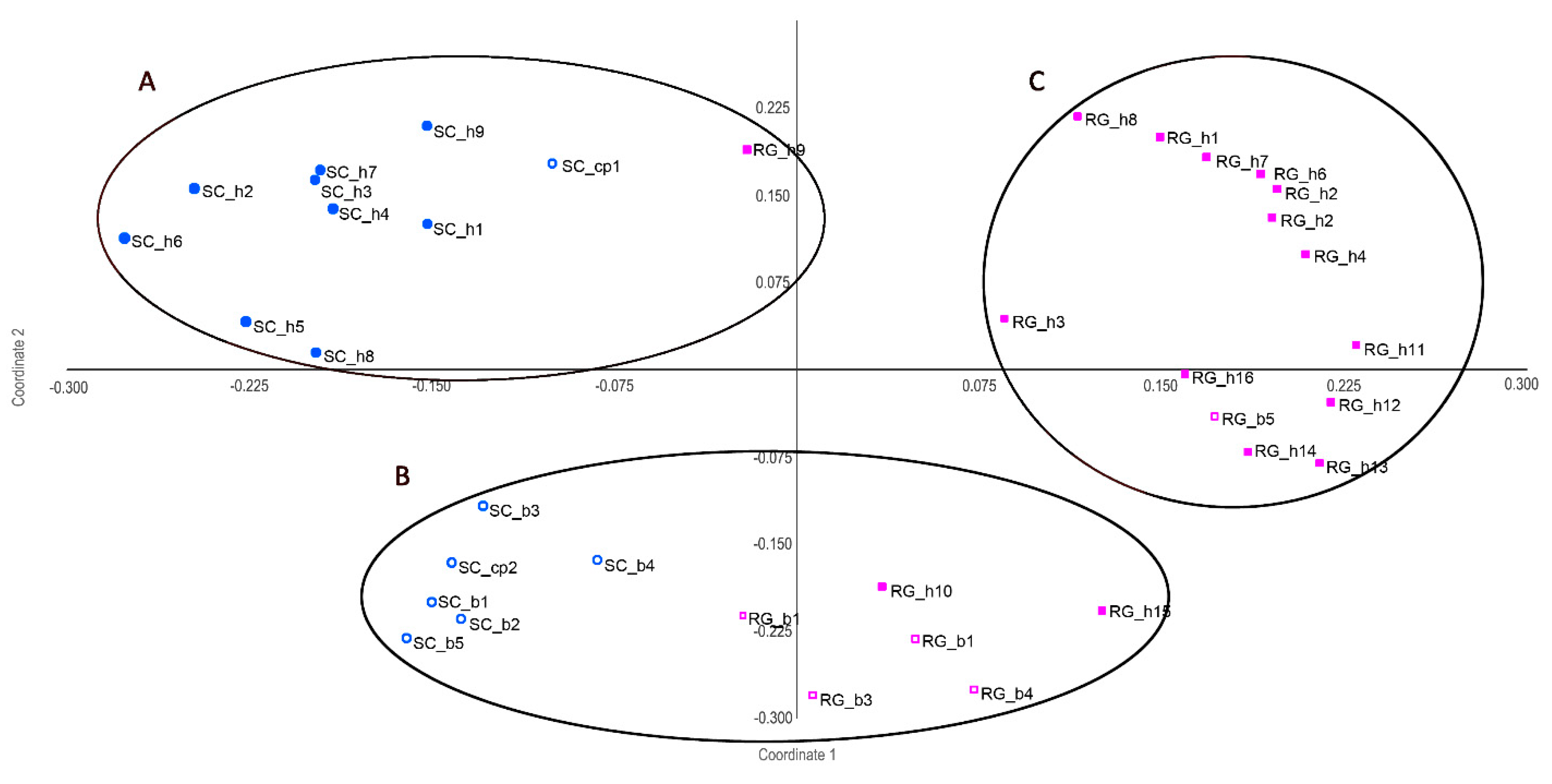
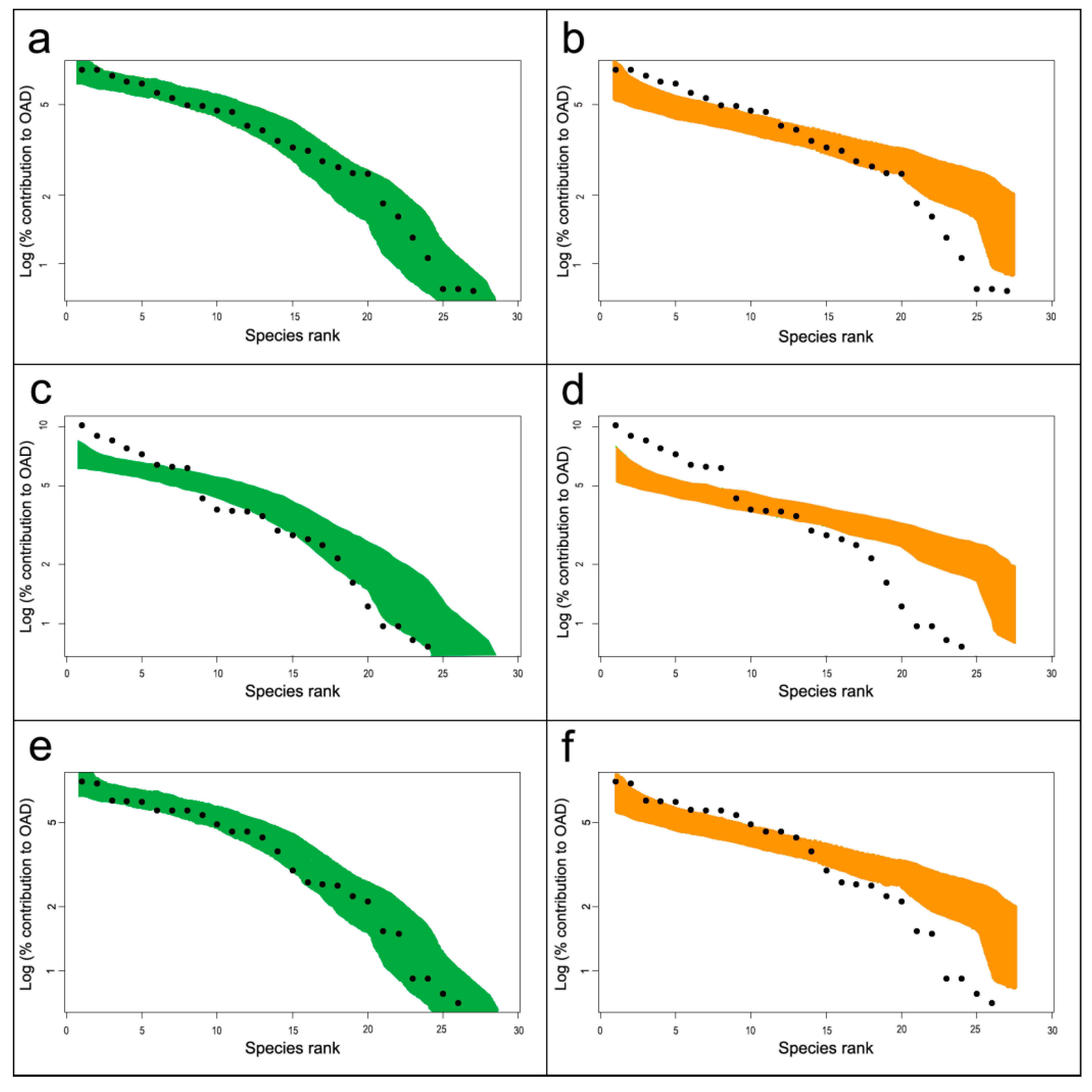
4.2. Biological Variables
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gibert, J.; Vervier, P.; Malard, F.; Laurent, R.; Reygrobellet, J.-L. Dynamics of Communities and Ecology of Karst Ecosystems: Example of Three Karsts in Eastern and Southern France; Academic Press: Cambridge, MA, USA, 1994; Volume 450, p. 425. [Google Scholar] [CrossRef]
- Humphreys, W.F. Hydrogeology and groundwater ecology: Does each inform the other? Hydrogeol. J. 2008, 17, 5–21. [Google Scholar] [CrossRef]
- Maurice, L.; Bloomfield, J. Stygobitic Invertebrates in Groundwater—A Review from a Hydrogeological Perspective. Freshw. Rev. 2012, 5, 51–71. [Google Scholar] [CrossRef] [Green Version]
- Larned, S.T.; Gooseff, M.N.; Packman, A.I.; Rugel, K.; Wondzell, S.M. Groundwater–surface-water interactions: Current research directions. Freshw. Sci. 2015, 34, 92–98. [Google Scholar] [CrossRef] [Green Version]
- Galassi, D.M.; Lombardo, P.; Fiasca, B.; Di Cioccio, A.; Di Lorenzo, T.; Petitta, M.; Di Carlo, P. Earthquakes trigger the loss of groundwater biodiversity. Sci. Rep. 2014, 4, 6273. [Google Scholar] [CrossRef] [PubMed]
- Di Lorenzo, T.; Cipriani, D.; Fiasca, B.; Rusi, S.; Galassi, D.M.P. Groundwater drift monitoring as a tool to assess the spatial distribution of groundwater species into karst aquifers. Hydrobiol. 2018, 813, 137–156. [Google Scholar] [CrossRef]
- Parise, M.; Gabrovsek, F.; Kaufmann, G.; Ravbar, N. Recent advances in karst research: From theory to fieldwork and applications. Geol. Soc. Lond. Spéc. Publ. 2018, 466, 1–24. [Google Scholar] [CrossRef]
- Gabrovšek, F.; Dreybrodt, W. A model of the early evolution of karst aquifers in limestone in the dimensions of length and depth. J. Hydrol. 2001, 240, 206–224. [Google Scholar] [CrossRef]
- Ford, D.; Williams, P. Karst Hydrogeology and Geomorphology; Wiley: Hoboken, NJ, USA, 2007. [Google Scholar] [CrossRef]
- Worthington, S.R.H.; Jeannin, P.-Y.; Alexander, E.C.; Davies, G.J.; Schindel, G.M. Contrasting definitions for the term ‘karst aquifer’. Hydrogeol. J. 2017, 25, 1237–1240. [Google Scholar] [CrossRef]
- Poulain, A.; Watlet, A.; Kaufmann, O.; Van Camp, M.; Jourde, H.; Mazzilli, N.; Rochez, G.; Deleu, R.; Quinif, Y.; Hallet, V. Assessment of groundwater recharge processes through karst vadose zone by cave percolation monitoring. Hydrol. Process. 2018, 32, 2069–2083. [Google Scholar] [CrossRef]
- Mangin, A. Sur La Dynamique Des Transferts En Aquiféres Karstique. In Proceedings of the Sixth International Congress of Speleology, Olomouc, Czech Republic, 2 July 1973 ; pp. 157–162. [Google Scholar]
- Mangin, A. Contribution à l’étude Hydrodynamique Des Aquifères Karstiques, Université de Dijon, France, 1975. (Alain Mangin. Contribution à l’étude hydrodynamique des aquifères karstiques: Première partie: Généralités sur le karst et les lois d’écoulement utilisées (Ann. Spéléol., 1974, 29, 3, p.283-332) Deuxième partie: Concepts méthodologiques adoptés. Systèmes karstiques étudiés. (Ann. Spéléol., 1974, 29, 4, p.495-601) Troisième partie: Constitution et fonctionnement des aquifères karstiques. (Ann. Spéléol.,1975, 30, 1, p.21-124) Sciences de la Terre Université de Dijon, 1975 Français. Available online: https://hal.archives-ouvertes.fr/tel-01575806 (accessed on 2 July 2021).
- Klimchouk, A.; Towards Defining, Delimiting and Classifying Epikarst: Its Origin, Processes and Variants of Geomorphic Evolution. Speleogenesis Evol. Karst Aquifers. Available online: https://speleogenesis.com/pdf/seka_pdf4501.pdf (accessed on 2 July 2021).
- Pipan, T.; Culver, D.C. Epikarst communities: Biodiversity hotspots and potential water tracers. Environ. Earth Sci. 2007, 53, 265–269. [Google Scholar] [CrossRef]
- Pipan, T. David David Culver, American University Forty years of epikarst: What biology have we learned? Int. J. Speleol. 2013, 42, 215–223. [Google Scholar] [CrossRef] [Green Version]
- Kozel, P.; Pipan, T. Specialized aquatic subterranean communities are probably most species-rich in the thickest epikarst. Limnol. 2020, 81, 125756. [Google Scholar] [CrossRef]
- Bonacci, O.; Pipan, T.; Culver, D.C. A framework for karst ecohydrology. Environ. Earth Sci. 2008, 56, 891–900. [Google Scholar] [CrossRef]
- Pipan, T.; Holt, N.; Culver, D.C. How to protect a diverse, poorly known, inaccessible fauna: Identification and protection of source and sink habitats in the epikarst. Aquat. Conserv. Mar. Freshw. Ecosyst. 2010, 20, 748–755. [Google Scholar] [CrossRef]
- Hutchins, B.; Gibson, J.; Diaz, P.; Schwartz, B. Stygobiont Diversity in the San Marcos Artesian Well and Edwards Aquifer Groundwater Ecosystem, Texas, USA. Divers. 2021, 13, 234. [Google Scholar] [CrossRef]
- Palmer, A.N. Hydrogeological control of cave patterns. In Speleogenesis: Evol. Karst Aquifers; Klimchouk, A.B., Ford, D.C., et al., Eds.; National Speleological Society: Huntsville, AL, USA, 2000. [Google Scholar]
- Lerch, R.N.; Wicks, C.M.; Moss, P.L. Hydrologic Characterization of Two Karst Recharge Areas in Boone County, Missouri. J. Cave Karst Stud. 2005, 67, 158–173. [Google Scholar]
- Ollivier, C.; Mazzilli, N.; Olioso, A.; Chalikakis, K.; Carrière, S.D.; Danquigny, C.; Emblanch, C. Karst recharge-discharge semi distributed model to assess spatial variability of flows. Sci. Total. Environ. 2020, 703, 134368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonacci, O. Surface Waters and Groundwater in Karst. In Karst Aquifers- Characterization and Engineering; Stevanović, Z., Ed.; Springer: Dordrecht, The Netherlands, 2015; pp. 149–169. [Google Scholar] [CrossRef]
- Butscher, C.; Huggenberger, P. Modeling the Temporal Variability of Karst Groundwater Vulnerability, with Implications for Climate Change. Environ. Sci. Technol. 2009, 43, 1665–1669. [Google Scholar] [CrossRef] [Green Version]
- Bonacci, O.; Terzić, J.; Roje-Bonacci, T.; Frangen, T. An Intermittent Karst River: The Case of the Čikola River (Dinaric Karst, Croatia). Water 2019, 11, 2415. [Google Scholar] [CrossRef] [Green Version]
- Chu, H.; Wei, J.; Wang, R.; Xin, B. Characterizing the interaction of groundwater and surface water in the karst aquifer of Fangshan, Beijing (China). Hydrogeol. J. 2016, 25, 575–588. [Google Scholar] [CrossRef]
- Keshavarzi, M.; Baker, A.; Kelly, B.F.J.; Andersen, M.S. River–groundwater connectivity in a karst system, Wellington, New South Wales, Australia. Hydrogeol. J. 2016, 25, 557–574. [Google Scholar] [CrossRef]
- Ghasemizadeh, R.; Hellweger, F.; Butscher, C.; Padilla, I.; Vesper, D.; Field, M.; Alshawabkeh, A. Review: Groundwater flow and transport modeling of karst aquifers, with particular reference to the North Coast Limestone aquifer system of Puerto Rico. Hydrogeol. J. 2012, 20, 1441–1461. [Google Scholar] [CrossRef] [Green Version]
- Drogue, C. Essai d’identi Fication d’un Type de Structure de Magasins Carbonates, Fissures. Mém H Sér Soc Géol Fr 1980, 11, 101–108. [Google Scholar]
- Meleg, I.N.; Fiers, F.; Robu, M.; Moldovan, O.T. Distribution patterns of subsurface copepods and the impact of environmental parameters. Limnol. 2012, 42, 156–164. [Google Scholar] [CrossRef]
- Petitta, M.; Caschetto, M.; Galassi, D.M.P.; Aravena, R. Dual-flow in karst aquifers toward a steady discharge spring (Presciano, Central Italy): Influences on a subsurface groundwater dependent ecosystem and on changes related to post-earthquake hydrodynamics. Environ. Earth Sci. 2014, 73, 2609–2625. [Google Scholar] [CrossRef]
- Hare, D.K.; Helton, A.M.; Johnson, Z.C.; Lane, J.W.; Briggs, M.A. Continental-scale analysis of shallow and deep groundwater contributions to streams. Nat. Commun. 2021, 12, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Rouch, R.; Carlier, A.; Le Système Karstique Du Baget. XIV. La Communauté Des Harpacticides. Evolution et Comparaison Des Structures Du Peuplement Epigé à l’ Entrée et à La Sortie de l’aquifère. Stygologia 1985, 1, 71–92. [Google Scholar]
- Malard, F.; Reygrobellet, J.-L.; Mathieu, J.; Lafont, M. The use of invertebrate communities to describe groundwater flow and contaminant transport in a fractured rock aquifer. Archiv. Hydrobiol. 1994, 131, 93–110. [Google Scholar] [CrossRef]
- Messouli, M. What Can Karstic Organisms Tell Us about Groundwater Functioning and Water Quality? In Proceedings of the BALWOIS 2006 Conference on Water Observation and Information System for Decision Support, Ohrid, Morocco, 23–26 May 2006; Available online: https://horizon.documentation.ird.fr/exl-doc/pleins_textes/divers16-09/010048589.pdf (accessed on 2 July 2021).
- Fiasca, B.; Stoch, F.; Olivier, M.-J.; Maazouzi, C.; Petitta, M.; Di Cioccio, A.; Galassi, D.M. The dark side of springs: What drives small-scale spatial patterns of subsurface meiofaunal assemblages? J. Limnol. 2014, 73. [Google Scholar] [CrossRef]
- Mori, N.; Kanduč, T.; Slabe, M.O.; Brancelj, A. Groundwater Drift as a Tracer for Identifying Sources of Spring Discharge. Ground Water 2015, 53, 123–132. [Google Scholar] [CrossRef]
- Stoch, F.; Fiasca, B.; Di Lorenzo, T.; Porfirio, S.; Petitta, M.; Galassi, D.M. Exploring copepod distribution patterns at three nested spatial scales in a spring system: Habitat partitioning and potential for hydrological bioindication. J. Limnol. 2015. [Google Scholar] [CrossRef] [Green Version]
- Brancelj, A.; Mori, N.; Treu, F.; Stoch, F. The groundwater fauna of the Classical Karst: Hydrogeological indicators and descriptors. Aquat. Ecol. 2019, 54, 205–224. [Google Scholar] [CrossRef]
- Cantonati, M.; Poikane, S.; Pringle, C.M.; Stevens, L.E.; Turak, E.; Heino, J.; Richardson, J.S.; Bolpagni, R.; Borrini, A.; Cid, N.; et al. Characteristics, Main Impacts, and Stewardship of Natural and Artificial Freshwater Environments: Consequences for Biodiversity Conservation. Water 2020, 12, 260. [Google Scholar] [CrossRef] [Green Version]
- Moldovan, O.T.; Meleg, I.N.; Perşoiu, A. Habitat fragmentation and its effects on groundwater populations. Ecohydrol. 2011, 5, 445–452. [Google Scholar] [CrossRef]
- Liu, W.; Zhou, C.; Burnet, J.E.; Brancelj, A. The effect of hydrological and hydrochemical parameters on the microdistribution of aquatic fauna in drip water in the Velika Pasica Cave, Central Slovenia. Ecohydrol. 2017, 10, e1835. [Google Scholar] [CrossRef]
- Pipan, T.; Culver, D.C.; Papi, F.; Kozel, P. Partitioning diversity in subterranean invertebrates: The epikarst fauna of Slovenia. PLoS ONE 2018, 13, e0195991. [Google Scholar] [CrossRef]
- Galassi, D.M.P.; Fiasca, B.; Di Lorenzo, T.; Montanari, A.; Porfirio, S.; Fattorini, S. Groundwater biodiversity in a chemoautotrophic cave ecosystem: How geochemistry regulates microcrustacean community structure. Aquat. Ecol. 2016, 51, 75–90. [Google Scholar] [CrossRef]
- Fattorini, S.; Fiasca, B.; Di Lorenzo, T.; Di Cicco, M.; Galassi, D.M.P. A new protocol for assessing the conservation priority of groundwater-dependent ecosystems. Aquat. Conserv. Mar. Freshw. Ecosyst. 2020, 30, 1483–1504. [Google Scholar] [CrossRef]
- Goldscheider, N. A holistic approach to groundwater protection and ecosystem services in karst terrains. Carbonates Evaporites 2019, 34, 1241–1249. [Google Scholar] [CrossRef]
- Galassi, D.M.P.; Huys, R.; Reid, J.W. Diversity, ecology and evolution of groundwater copepods. Freshw. Biol. 2009, 54, 691–708. [Google Scholar] [CrossRef] [Green Version]
- Iannella, M.; Fiasca, B.; Di Lorenzo, T.; Biondi, M.; Di Cicco, M.; Galassi, D.M.P. Jumping into the grids: Mapping biodiversity hotspots in groundwater habitat types across Europe. Ecography 2020, 43, 1825–1841. [Google Scholar] [CrossRef]
- Iannella, M.; Fiasca, B.; Di Lorenzo, T.; Biondi, M.; Di Cicco, M.; Galassi, D.M.P. Spatial distribution of stygobitic crustacean harpacticoids at the boundaries of groundwater habitat types in Europe. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Panzanaro, M. La Rete di Monitoraggio delle Grotte di Stiffe. In Le Grotte d’Italia rivista annuale dell’Istituto Italiano di Speleologia e della Società Speleologica Italiana. 2002, pp. 122–132. Available online: http://www.gruppospeleologicoaquilano.it/wp-content/uploads/2017/05/Descrizione-della-rete-di-monitoraggio-delle-Grotte-di-Stif2.pdf (accessed on 2 July 2021).
- Chiarelli, B.; Imprescia, U.; Lorè, M.; Panzanaro, M.; Pettini, A.; Romagnoli, C.; Atti Della Stazione Scientifica Di Stiffe; Panzanaro, M.; Romagnoli, C. (Eds.) Museo Speleologico “V Rivera” Gruppo Speleologico Aquilano. 1995. Available online: http://www.gruppospeleologicoaquilano.it/wp-content/uploads/2017/05/ATTI-DELLA-STAZIONE-SCIENTIFICA-DI-STIFFE-web.pdf (accessed on 2 July 2021).
- Scorzini, A.R.; Leopardi, M. River basin planning: From qualitative to quantitative flood risk assessment: The case of Abruzzo Region (central Italy). Nat. Hazards 2017, 88, 71–93. [Google Scholar] [CrossRef]
- Bou, C.; Rouch, R. Un Nouveau Champ de Recherches Sur La Faune Aquatique Souterraine. CR Acad Sci 1967, 265, 369–370. [Google Scholar]
- Fischer, H.; Wanner, S.C.; Pusch, M. Bacterial abundance and production in river sediments as related to the biochemical composition of particulate organic matter (POM). Biogeochem. 2002, 61, 37–55. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. Primer v6: User Manual/Tutorial; PRIMER-E: Plymouth, 2006. [Google Scholar]
- Magurran, A.E.; McGill, B.J. Frontiers in Measurement and Assessment. In Biological Diversity, 2nd ed.; Oxford University Press: Oxford, UK; New York, NY, USA, 2011; ISBN 978-0-19-958067-5. [Google Scholar]
- Watson, L.; Williams, W.T.; Lance, G.N. Angiosperm taxonomy: A comparative study of some novel numerical techniques. J. Linn. Soc. London, Bot. 1966, 59, 491–501. [Google Scholar] [CrossRef]
- Sörensen, T.A. A Method of Establishing Groups of Equal Amplitude in Plant Sociology Based on Similarity of Species Content and Its Application to Analyses of the Vegetation on Danish Commons. Biol Skar. 1948, 5, 1–34. [Google Scholar]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Austral Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2008, 26, 32–46. [Google Scholar] [CrossRef]
- Anderson, M.J. Distance-Based Tests for Homogeneity of Multivariate Dispersions. Biom. 2005, 62, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Gibert, C.; Escarguel, G. PER-SIMPER-A new tool for inferring community assembly processes from taxon occurrences. Glob. Ecol. Biogeogr. 2018, 28, 374–385. [Google Scholar] [CrossRef]
- Legendre, P.; Galzin, R.; Harmelin-Vivien, M.L. Relating Behavior to Habitat: Solutions to the Fourth-corner Problem. Ecol. 1997, 78, 547–562. [Google Scholar] [CrossRef]
- Gotelli, N.J. Null Model Analysis of Species Co-Occurrence Patterns. Ecol. 2000, 81, 2606–2621. [Google Scholar] [CrossRef]
- Culver, D.C.; Pipan, T. The Biology of Caves and Other Subterranean Habitats. In Biology of Habitats Series, 2nd ed.; Oxford University Press: Oxford, UK, 2019; ISBN 978-0-19-882076-5. [Google Scholar]
- Rouch, R. Considérations Sur l’écosystème Karstique. C.R. Acad. Sci. 1977, 284, 1101–1103. [Google Scholar]
- Simões, L.; Ferreira, T.; Bichuette, M.E.B. Aquatic Biota of Different Karst Habitats in Epigean and Subterranean Systems of Central Brazil – Visibility Versus Relevance of Taxa. Subterr. Biol. 2013, 11, 55–74. [Google Scholar] [CrossRef]
- Mori, N.; Brancelj, A.; Machel, H. Differences in aquatic microcrustacean assemblages between temporary and perennial springs of an alpine karstic aquifer. Int. J. Speleol. 2013, 42, 257–266. [Google Scholar] [CrossRef] [Green Version]
- Devitt, T.J.; Wright, A.M.; Cannatella, D.C.; Hillis, D.M. Species Delimitation in Endangered Groundwater Salamanders: Implications for Aquifer Management and Biodiversity Conservation. Proc. Natl. Acad. Sci. 2019, 116, 2624–2633. [Google Scholar] [CrossRef] [Green Version]
- Brancelj, A. Fauna of an unsaturated karstic zone in Central Slovenia: Two new species of Harpacticoida (Crustacea: Copepoda), Elaphoidella millennii n. sp. and E. tarmani n. sp., their ecology and morphological adaptations. Hydrobiol. 2008, 621, 85–104. [Google Scholar] [CrossRef]
- Pipan, T. Epikarst—A Promising Habitat: Copepod Fauna, Its Diversity and Ecology; a Case Study from Slovenia (Europe); Carsologica; Institut za raziskovanje krasa ZRC SAZU: Postojna, Slovenia, 2005; ISBN 978-961-6500-90-6. [Google Scholar]
- Sánchez-Fernández, D.; Galassi, D.M.P.; Wynne, J.J.; Cardoso, P.; Mammola, S. Don’t Forget Subterranean Ecosystems in Climate Change Agendas. Nat. Clim. Chang. 2021, 11, 458–459. [Google Scholar] [CrossRef]
- Iannella, M.; Fiasca, B.; Lorenzo, T.D.; Biondi, M.; Di Cicco, M.; Mammola, S.; Galassi, D.M.P. Getting the “most out of the Hotspot” for Practical Conservation of Groundwater Biodiversity. Glob. Ecol. Conserv. under review.
- Mammola, S.; Lunghi, E.; Bilandžija, H.; Cardoso, P.; Grimm, V.; Schmidt, S.I.; Hesselberg, T.; Martínez, A. Collecting eco-evolutionary data in the dark: Impediments to subterranean research and how to overcome them. Ecol. Evol. 2021, 11, 5911–5926. [Google Scholar] [CrossRef] [PubMed]
- Francois, C.M.; Mermillod-Blondin, F.; Malard, F.; Fourel, F.; Lécuyer, C.; Douady, C.; Simon, L. Trophic ecology of groundwater species reveals specialization in a low-productivity environment. Funct. Ecol. 2015, 30, 262–273. [Google Scholar] [CrossRef]
- Fišer, Z.; Altermatt, F.; Zakšek, V.; Knapič, T.; Fišer, C. Morphologically Cryptic Amphipod Species Are “Ecological Clones” at Regional but Not at Local Scale: A Case Study of Four Niphargus Species. PLoS ONE 2015, 10, e0134384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fattorini, S.; Lombardo, P.; Fiasca, B.; Di Cioccio, A.; Di Lorenzo, T.; Galassi, D.M. Earthquake-Related Changes in Species Spatial Niche Overlaps in Spring Communities. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Ercoli, F.; Lefebvre, F.; Delangle, M.; Godé, N.; Caillon, M.; Raimond, R.; Souty-Grosset, C. Differing trophic niches of three French stygobionts and their implications for conservation of endemic stygofauna. Aquat. Conserv. Mar. Freshw. Ecosyst. 2019, 29, 2193–2203. [Google Scholar] [CrossRef]
- Hutchins, B.T.; Engel, A.S.; Nowlin, W.H.; Schwartz, B.F. Chemolithoautotrophy supports macroinvertebrate food webs and affects diversity and stability in groundwater communities. Ecol. 2016, 97, 1530–1542. [Google Scholar] [CrossRef]
- Saccò, M.; Blyth, A.J.; Humphreys, W.F.; Kuhl, A.; Mazumder, D.; Smith, C.; Grice, K. Elucidating stygofaunal trophic web interactions via isotopic ecology. PLoS ONE 2019, 14, e0223982. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RG Benthic | RG Hyporheic | SC Benthic | SC Hyporheic | |
|---|---|---|---|---|
| Number of samples | 9 | 31 | 14 | 18 |
| Electrical conductivity (µS cm−1, 25 °C) | 480.66 ± 36.60 | 513.00 ± 105.59 | 476.71 ± 14.84 | 503.22 ± 79.21 |
| Temperature (°C) | 6.58 ± 4.16 | 9.61 ± 4.71 | 7.85 ± 1.83 | 8.33 ± 1.90 |
| O2 (mg L−1) | 11.02 ± 1.21 | 6.21 ± 2.74 | 10.07 ± 0.66 | 8.86 ± 1.05 |
| pH | 8.28 ± 0.16 | 7.93 ± 0.23 | 7.61 ± 0.13 | 7.66 ± 0.13 |
| NO3- (mg L−1) | 2.85 ± 2.79 | 3.81 ± 3.08 | 2.32 ± 1.07 | 3.97 ± 1.81 |
| NO2- (mg L−1) | 0.10 ± 0.11 | 0.14 ± 0.18 | 0.03 (LOD) | 1.03 ± 1.46 |
| NH4+ (mg L−1) | 0.03 (LOD) | 0.45 ± 0.68 | 0.03 (LOD) | 0.03 (LOD) |
| SO42- (mg L−1) | 6.13 ± 2.28 | 8.08 ± 7.61 | 9.60 ± 1.21 | 8.67 ± 0.97 |
| Cl- (mg L−1) | 3.67 ± 2.83 | 7.31 ± 16.2 | 10.40 ± 2.58 | 7.56 ± 5.54 |
| K+ (mg L−1) | 0.90 ± 0.50 | 1.30 ± 1.00 | 2.9 ± 0.20 | 2.7 ± 0.90 |
| Ca2+ (mg L−1) | 98.30 ± 9.01 | 92.44 ± 32.28 | 93.64 ± 11.79 | 92.83 ± 7.81 |
| Na+ (mg L−1) | 6.10 ± 1.33 | 6.07 ± 1.44 | 7.79 ± 1.85 | 8.31 ± 1.06 |
| DOC (mg L−1) | 1.52 ± 0.45 | 1.67 ± 0.43 | 2.16 ± 0.89 | 1.98 ± 0.83 |
| POM (mg L−1) | 110.44 ± 79.28 | 338.45 ± 275.55 | 35.50 ± 18.36 | 195.78 ± 258.95 |
| Species Name | Ecology | RG | SC | RG_b | RG_h | SC_b | SC_h | |
|---|---|---|---|---|---|---|---|---|
| Macrocyclops albidus | (Jurine, 1820) | nSB | 1 | 0 | 0 | 1 | 0 | 0 |
| Tropocyclops prasinus | (Fischer, 1860) | nSB | 3 | 0 | 3 | 0 | 0 | 0 |
| Paracyclops fimbriatus | (Fischer, 1853) | nSB | 13 | 0 | 4 | 9 | 0 | 0 |
| Paracyclops imminutus | Kiefer, 1929 | nSB | 3 | 6 | 3 | 0 | 2 | 4 |
| Eucyclops serrulatus | (Fischer, 1851) | nSB | 10 | 1 | 3 | 7 | 1 | 0 |
| Eucyclops intermedius | (Damian, 1955) | SB | 3 | 0 | 0 | 3 | 0 | 0 |
| Acanthocyclops robustus | (Sars G.O., 1863) | nSB | 6 | 2 | 3 | 3 | 2 | 0 |
| Megacyclops viridis | (Jurine, 1820) | nSB | 3 | 3 | 1 | 2 | 1 | 2 |
| Diacyclops bisetosus | (Rehberg, 1880) | nSB | 4 | 0 | 2 | 2 | 0 | 0 |
| Diacyclops italianus | (Kiefer, 1931) | SB | 1 | 0 | 1 | 0 | 0 | 0 |
| Diacyclops paolae | Pesce & Galassi, 1987 | SB | 0 | 12 | 0 | 0 | 3 | 9 |
| Diacyclops clandestinus | (Yeatman, 1964) | SB | 2 | 1 | 0 | 2 | 0 | 1 |
| Diacyclops paralanguidoides | Pesce & Galassi, 1987 | SB | 0 | 1 | 0 | 0 | 0 | 1 |
| Diacyclops sp. 1 | SB | 0 | 3 | 0 | 0 | 0 | 3 | |
| Hypocamptus brehmi | (Douwe, 1922) | nSB | 0 | 2 | 0 | 0 | 2 | 0 |
| Canthocamptus staphylinus | (Jurine, 1820) | nSB | 10 | 5 | 4 | 6 | 5 | 0 |
| Attheyella paranaphtalica | Pesce & Galassi, 1988 | SB | 0 | 9 | 0 | 0 | 6 | 3 |
| Attheyella crassa | (Sars G.O., 1863) | nSB | 9 | 7 | 4 | 5 | 5 | 2 |
| Bryocamptus zschokkei | (Schmeil, 1893) | nSB | 2 | 5 | 2 | 0 | 4 | 1 |
| Bryocamptus pygmaeus | (Sars, G.O., 1863) | nSB | 15 | 10 | 5 | 10 | 6 | 4 |
| Bryocamptus echinatus | (Mrázek, 1893) | nSB | 7 | 16 | 3 | 4 | 7 | 9 |
| Pesceus schmeili | (Mrázek, 1893) | nSB | 0 | 14 | 0 | 0 | 7 | 7 |
| Elaphoidella plutonis | Chappuis, 1938 | SB | 0 | 4 | 0 | 0 | 2 | 2 |
| Elaphoidella phreatica | (Chappuis, 1925) | SB | 0 | 9 | 0 | 0 | 6 | 3 |
| Moraria poppei meridionalis | Chappuis, 1929 | nSB | 3 | 0 | 2 | 1 | 0 | 0 |
| Epactophanes richardi | Mrázek, 1893 | nSB | 0 | 7 | 0 | 0 | 3 | 4 |
| Phyllognathopus viguieri | (Maupas, 1892) | nSB | 1 | 0 | 1 | 0 | 0 | 0 |
| Parastenocaris crenobia | Galassi, 1997 | SB | 0 | 10 | 0 | 0 | 2 | 8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Cicco, M.; Di Lorenzo, T.; Iannella, M.; Vaccarelli, I.; Galassi, D.M.P.; Fiasca, B. Linking Hydrogeology and Ecology in Karst Landscapes: The Response of Epigean and Obligate Groundwater Copepods (Crustacea: Copepoda). Water 2021, 13, 2106. https://doi.org/10.3390/w13152106
Di Cicco M, Di Lorenzo T, Iannella M, Vaccarelli I, Galassi DMP, Fiasca B. Linking Hydrogeology and Ecology in Karst Landscapes: The Response of Epigean and Obligate Groundwater Copepods (Crustacea: Copepoda). Water. 2021; 13(15):2106. https://doi.org/10.3390/w13152106
Chicago/Turabian StyleDi Cicco, Mattia, Tiziana Di Lorenzo, Mattia Iannella, Ilaria Vaccarelli, Diana Maria Paola Galassi, and Barbara Fiasca. 2021. "Linking Hydrogeology and Ecology in Karst Landscapes: The Response of Epigean and Obligate Groundwater Copepods (Crustacea: Copepoda)" Water 13, no. 15: 2106. https://doi.org/10.3390/w13152106