
Diversity of Multidrug-Resistant Bacteria in an Urbanized River: A Case Study of the Potential Risks from Combined Sewage Overflows


Abstract
:1. Introduction
2. Materials and Methods
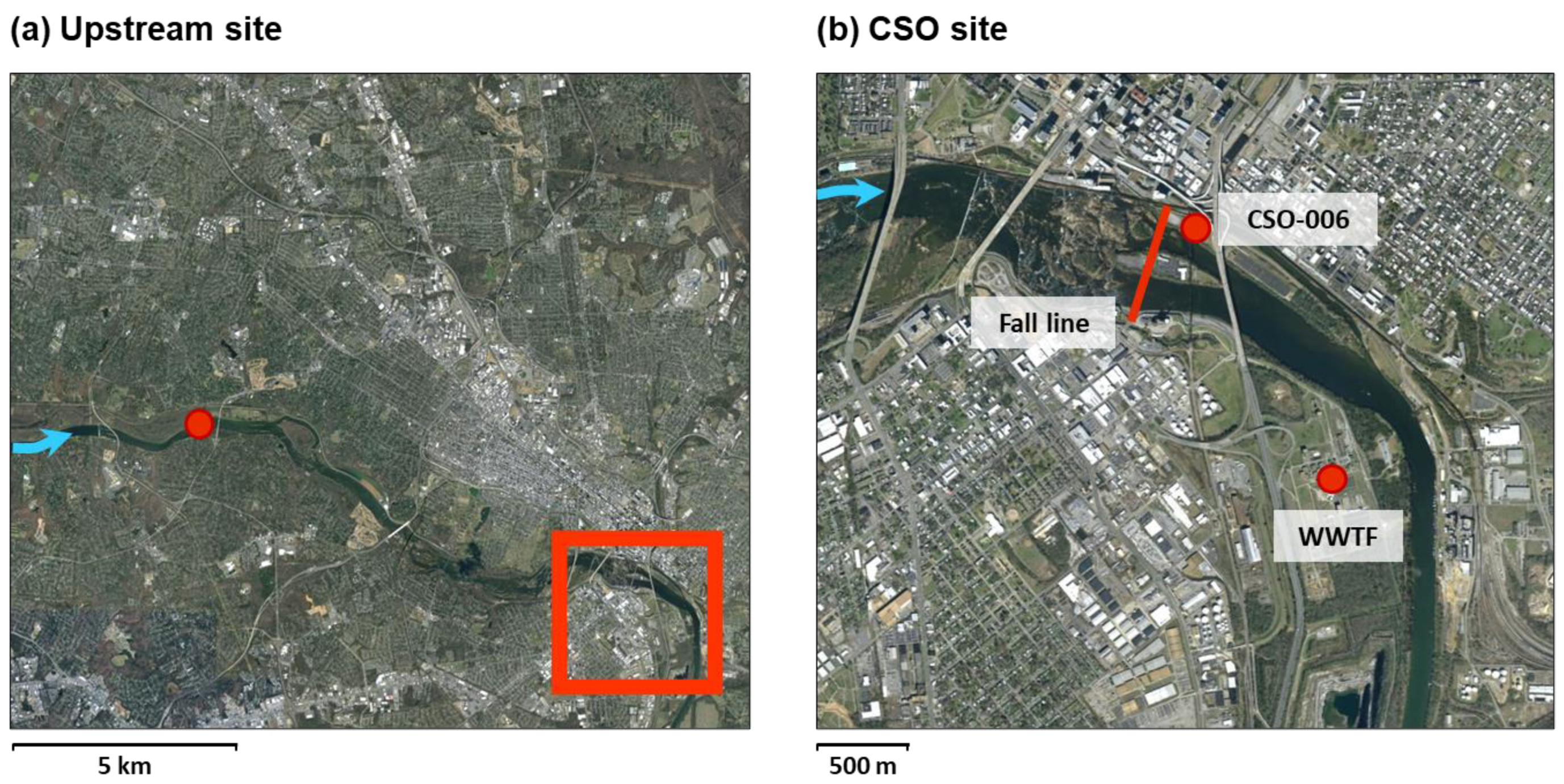
2.1. Study Sites
2.2. Sampling
2.3. Microbiological Analyses
2.4. Isolations and MDR Testing
2.5. DNA Extraction and Sequencing
2.6. Statistical Analyses
3. Results
3.1. Water Quality
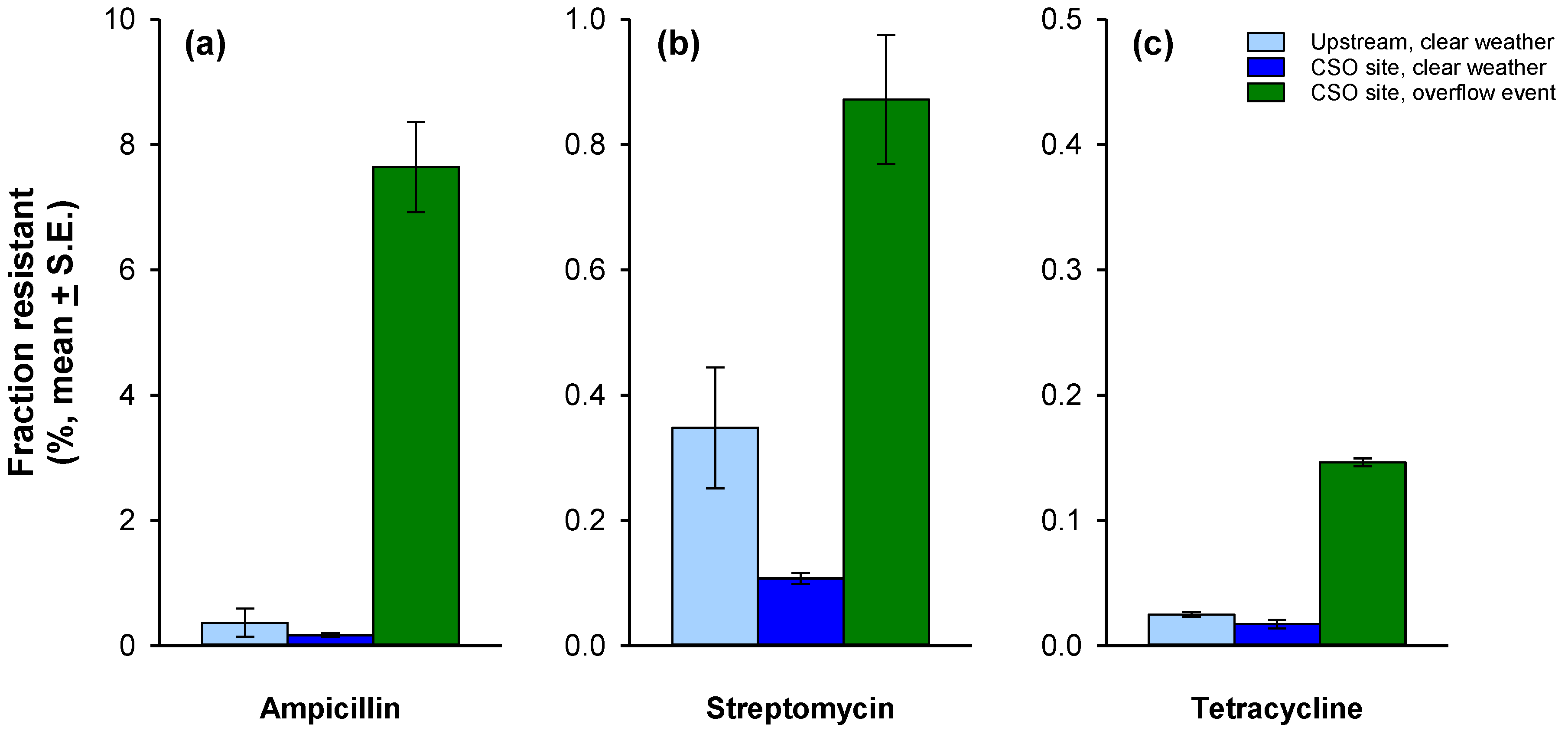
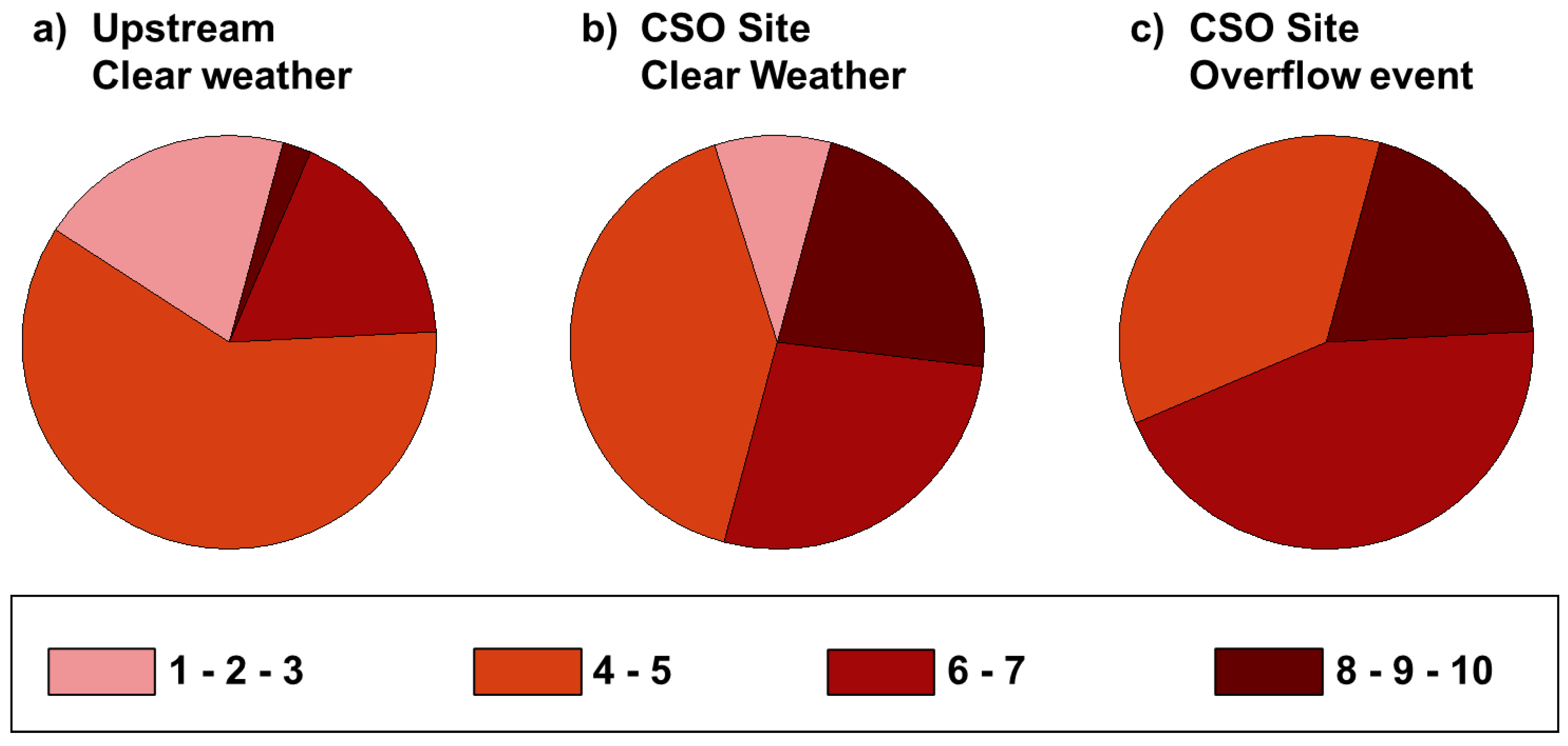
3.2. Antibiotic Resistance
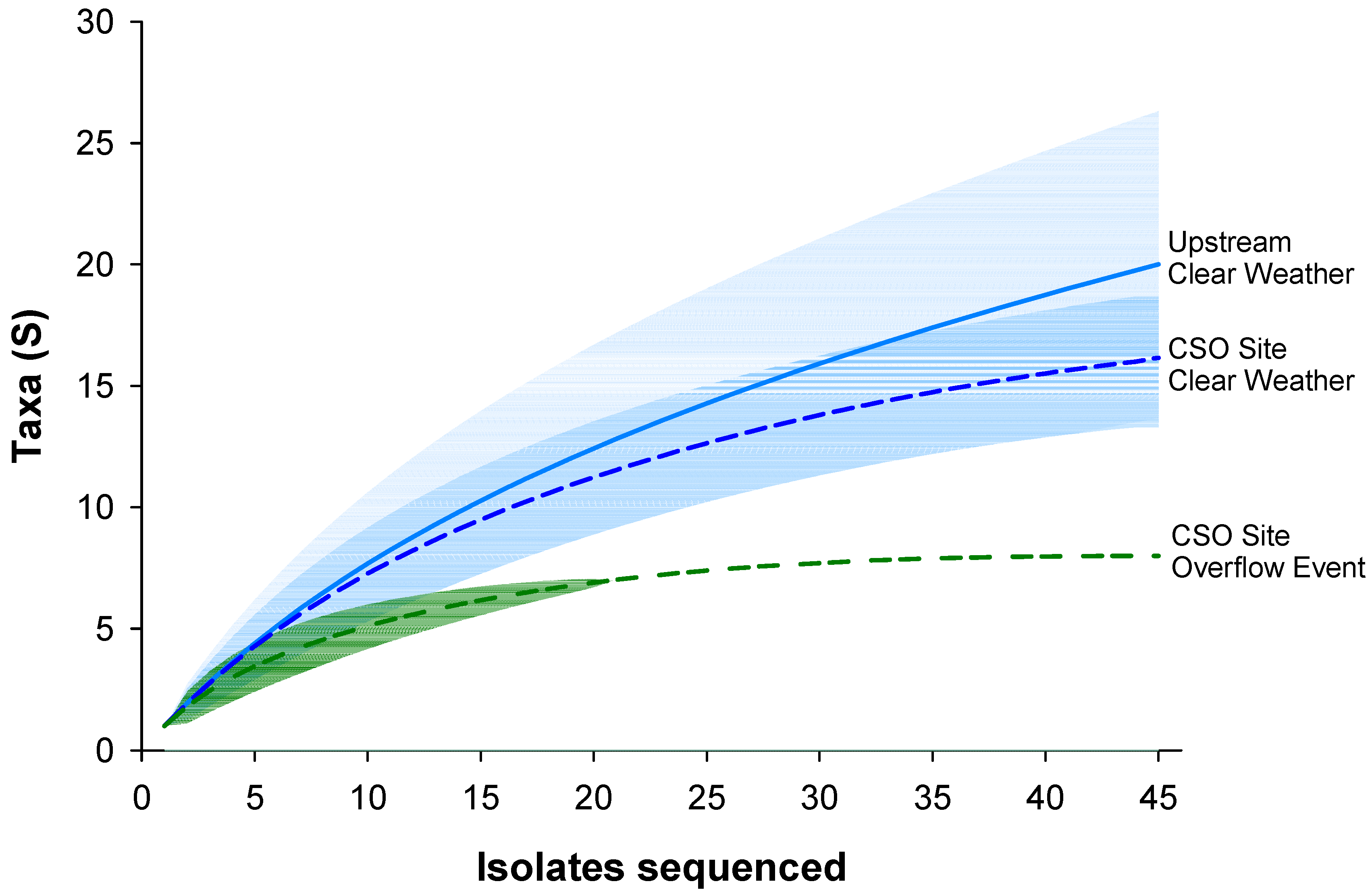
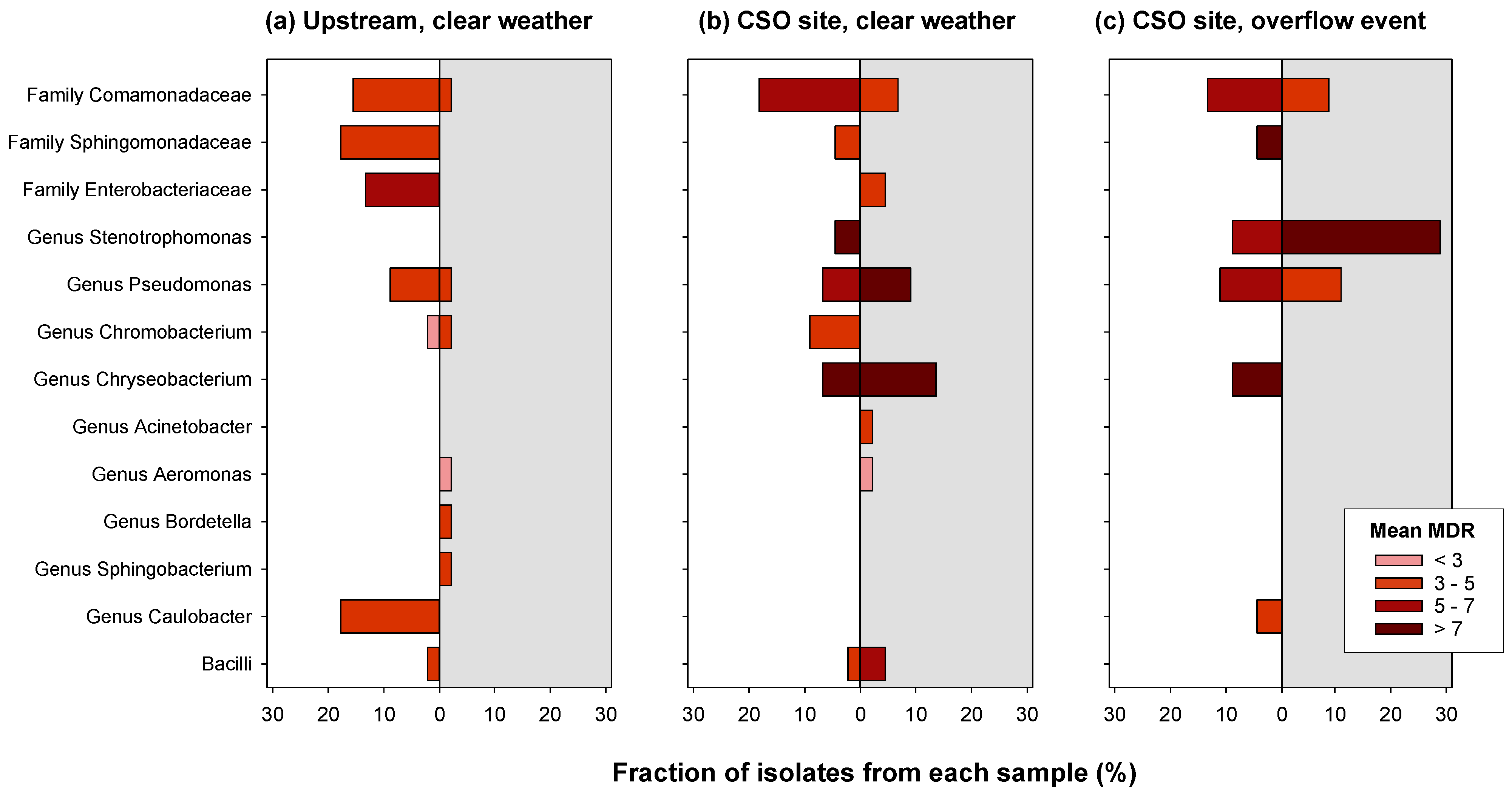
3.3. Identification of Bacterial Isolates
4. Discussion
4.1. Multidrug Resistance
4.2. Resistance to Individual Antibiotics
4.3. Identification and Diversity of Resistant Bacteria
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rizzo, L.; Manaia, C.; Merlin, C.; Schwartz, T.; Dagot, C.; Ploy, M.C.; Michael, I.; Fatta-Kassinos, D. Urban wastewater treatment plants as hotspots for antibiotic resistant bacteria and genes spread into the environment: A review. Sci. Total Environ. 2013, 447, 345–360. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chu, L.; Wojnárovits, L.; Takács, E. Occurrence and fate of antibiotics, antibiotic resistant genes (ARGs) and antibiotic resistant bacteria (ARB) in municipal wastewater treatment plant: An overview. Sci. Total Environ. 2020, 744, 140997. [Google Scholar] [CrossRef] [PubMed]
- Chu, B.T.T.; Petrovich, M.L.; Chaudhary, A.; Wright, D.; Murphy, B.; Wells, G.; Poretsky, R. Metagenomics reveals the impact of wastewater treatment plants on the dispersal of microorganisms and genes in aquatic sediments. Appl. Environ. Microbiol. 2018, 84, e02168-17. [Google Scholar]
- Michael, I.; Rizzo, L.; McArdell, C.S.; Manaia, C.M.; Merlin, C.; Schwartz, T.; Dagot, C.; Fatta-Kassinos, D. Urban wastewater treatment plants as hotspots for the release of antibiotics in the environment: A review. Water Res. 2013, 47, 957–995. [Google Scholar] [CrossRef]
- Raza, S.; Jo, H.; Kim, J.; Shin, H.; Hur, H.G.; Unno, T. Metagenomic exploration of antibiotic resistome in treated wastewater effluents and their receiving water. Sci. Total Environ. 2021, 765, 142755. [Google Scholar]
- Rodríguez-Mozaz, S.; Chamorro, S.; Marti, E.; Huerta, B.; Gros, M.; Sànchez-Melsió, A.; Borrego, C.; Barcelo, D.; Balcázar, J.L. Occurrence of antibiotics and antibiotic resistance genes in hospital and urban wastewaters and their impact on the receiving river. Water Res. 2015, 69, 234–242. [Google Scholar] [CrossRef]
- Singh, R.; Singh, A.P.; Kumar, S.; Giri, B.S.; Kim, K.H. Antibiotic resistance in major rivers in the world: A systematic review on occurrence, emergence, and management strategies. J. Clean. Prod. 2019, 234, 1484–1505. [Google Scholar] [CrossRef]
- Tibbetts, J. Combined sewer systems: Down, dirty, and out of date. Environ. Health Perspect. 2005, 113, A464–A467. [Google Scholar] [CrossRef]
- Honda, R.; Watanabe, T.; Sawaittayotin, V.; Masago, Y.; Chulasak, R.; Tanong, K.; Chaminda, G.; Wongsila, K.; Sienglum, C.; Sunthonwatthanaphong, V.; et al. Impacts of urbanization on the prevalence of antibiotic-resistant Escherichia coli in the Chaophraya River and its tributaries. Water Sci. Technol. 2015, 73, 362–374. [Google Scholar] [CrossRef]
- Port, J.A.; Cullen, A.C.; Wallace, J.C.; Smith, M.N.; Faustman, E.M. Metagenomic frameworks for monitoring antibiotic resistance in aquatic environments. Environ. Health Perspect. 2014, 122, 222–228. [Google Scholar] [CrossRef]
- Kumar, M.; Ram, B.; Honda, R.; Poopipattana, C.; Canh, V.; Chaminda, T.; Furumai, H. Concurrence of antibiotic resistant bacteria (ARB), viruses, pharmaceuticals and personal care products (PPCPs) in ambient waters of Guwahati, India: Urban vulnerability and resilience perspective. Sci. Total Environ. 2019, 693, 133640. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, T.; Kohnen, W.; Jansen, B.; Obst, U. Detection of antibiotic-resistant bacteria and their resistance genes in wastewater, surface water, and drinking water biofilms. FEMS Microbiol. Ecol. 2003, 43, 325–335. [Google Scholar] [CrossRef]
- Berendonk, T.U.; Manaia, C.M.; Merlin, C.; Fatta-Kassinos, D.; Cytryn, E.; Walsh, F.; Burgmann, H.; Sorum, H.; Norstrom, M.; Pons, M.N.; et al. Tackling antibiotic resistance: The environmental framework. Nat. Rev. Microbiol. 2015, 13, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Garner, E.; Benitez, R.; von Wagoner, E.; Sawyer, R.; Schaberg, E.; Hession, W.; Krometis, L.; Badgley, B.; Pruden, A. Stormwater loadings of antibiotic resistance genes in an urban stream. Water Res. 2017, 123, 144–152. [Google Scholar] [CrossRef]
- Brown, P.C.; Borowska, E.; Peschke, R.; Schwartz, T.; Horn, H. Decay of elevated antibiotic resistance genes in natural river sediments after sedimentation of wastewater particles. Sci. Total Environ. 2020, 705, 135861. [Google Scholar] [CrossRef]
- Tennstedt, T.; Szczepanowski, R.; Braun, S.; Pühler, A.; Schlüter, A. Occurrence of integron-associated resistance gene cassettes located on antibiotic resistance plasmids isolated from a wastewater treatment plant. FEMS Microbiol. Ecol. 2003, 45, 239–252. [Google Scholar] [CrossRef]
- Von Wintersdorff, C.J.H.; Penders, J.; van Niekerk, J.M.; Mills, N.D.; Majumder, S.; van Alphen, L.B.; Savelkoul, P.H.M.; Wolffs, P.F.G. Dissemination of antimicrobial resistance in microbial ecosystems through horizontal gene transfer. Front. Microbiol. 2016, 7, 173. [Google Scholar] [CrossRef]
- Kim, S.; Aga, D.S. Potential ecological and human health impacts of antibiotics and antibiotic-resistant bacteria from wastewater treatment plants. J. Toxicol. Environ. Health Part B Crit. Rev. 2007, 10, 559–573. [Google Scholar] [CrossRef]
- Alekshun, M.N.; Levy, S.B. Molecular mechanisms of antibacterial multidrug resistance. Cell 2007, 128, 1037–1050. [Google Scholar] [CrossRef]
- Chen, B.; Liang, X.; Huang, X.; Zhang, T.; Li, X. Differentiating anthropogenic impacts on ARGs in the Pearl River Estuary by using suitable gene indicators. Water Res. 2013, 47, 2811–2820. [Google Scholar] [CrossRef]
- Garner, E.; Wallace, J.; Argoty, G.; Wilkinson, C.; Fahrenfeld, N.; Heath, L.; Zhang, L.; Arabi, M.; Aga, D.; Pruden, A. Metagenomic profiling of historic Colorado Front Range flood impact on distribution of riverine antibiotic resistance genes. Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Imchen, M.; Kumavath, R. Shotgun metagenomics reveals a heterogeneous prokaryotic community and a wide array of antibiotic resistance genes in mangrove sediment. FEMS Microbiol. Ecol. 2020, 96, 173. [Google Scholar] [CrossRef] [PubMed]
- Reichert, G.; Hilgert, S.; Alexander, J.; de Azevedo, J.C.R.; Morck, T.; Fuchs, S.; Schwartz, T. Determination of antibiotic resistance genes in a WWTP-impacted river in surface water, sediment, and biofilm: Influence of seasonality and water quality. Sci. Total Environ. 2021, 768, 144526. [Google Scholar] [CrossRef]
- Korzeniewska, E.; Korzeniewska, A.; Harnisz, M. Antibiotic resistant Escherichia coli in hospital and municipal sewage and their emission to the environment. Ecotoxicol. Environ. Saf. 2013, 91, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Müller, H.; Sib, E.; Gajdiss, M.; Klanke, U.; Lenz-Plet, F.; Barabasch, V.; Albert, C.; Schallenberg, A.; Timm, C.; Zacharias, N.; et al. Dissemination of multi-resistant Gram-negative bacteria into German wastewater and surface waters. FEMS Microbiol. Ecol. 2018, 94, fiy057. [Google Scholar] [CrossRef]
- Vaz-Moreira, I.; Varela, A.R.; Pereira, T.V.; Fochat, R.C.; Manaia, C.M. Multidrug resistance in quinolone-resistant gram-negative bacteria isolated from hospital effluent and the municipal wastewater treatment plant. Microb. Drug Resist. 2016, 22, 155–163. [Google Scholar] [CrossRef]
- Osińska, A.; Harnisz, M.; Korzeniewska, E. Prevalence of plasmid-mediated multidrug resistance determinants in fluoroquinolone-resistant bacteria isolated from sewage and surface water. Environ. Sci. Pollut. Res. 2016, 23, 10818–10831. [Google Scholar] [CrossRef]
- Ho, J.Y.; Jong, M.C.; Acharya, K.; Liew, S.S.X.; Smith, D.R.; Noor, Z.Z.; Goodson, M.L.; Werner, D.; Graham, D.W.; Eswaran, J. Multidrug-resistant bacteria and microbial communities in a river estuary with fragmented suburban waste management. J. Hazard. Mater. 2021, 405, 124687. [Google Scholar] [CrossRef]
- Yewale, P.P.; Lokhande, K.B.; Sridhar, A.; Vaishnav, M.; Khan, F.A.; Mandal, A.; Swamy, K.V.; Jass, J.; Nawani, N. Molecular profiling of multidrug-resistant river water isolates: Insights into resistance mechanism and potential inhibitors. Environ. Sci. Pollut. Res. Int. 2020, 27, 27279–27292. [Google Scholar] [CrossRef]
- Levengood, E.S. Effect of combined sewer overflow on the abundance of antibiotic resistant bacteria in the James River. Master’s Thesis, Virginia Commonwealth University, Richmond, VA, USA, 2017. [Google Scholar]
- United States Environmental Protection Agency [EPA]. Method 1603: Escherichia Coli (E. coli) in Water by Membrane Filtration Using Modified Membrane-Thermotolerant Escherichia Coli Agar (Modified mTEC); United States Environmental Protection Agency: Washington, DC, USA, 2009. [Google Scholar]
- Sandegren, L. Selection of antibiotic resistance at very low antibiotic concentrations. Ups. J. Med. Sci. 2014, 119, 103–107. [Google Scholar] [CrossRef]
- Patel, J.B.; Cockerill, F.; Alder, J.; Bradford, P.; Eliopoulos, G.; Hardy, D.; Hindler, J.; Jenkins, S.; Lewis, J.; Miller, L.; et al. Performance Standards for Antimicrobial Susceptibility Testing: Twenty-Fourth Informational Supplement; CLSI: Wayne, PA, USA, 2014. [Google Scholar]
- Dhanda, G.; Sarkar, P.; Samaddar, S.; Haldar, J. Battle against vancomycin-resistant bacteria: Recent developments in chemical strategies. J. Med. Chem. 2019, 62, 3184–3205. [Google Scholar] [CrossRef] [PubMed]
- Werner, G.; Strommenger, B.; Witte, W. Acquired vancomycin resistance in clinically relevant pathogens. Future Microbiol. 2008, 3, 547–562. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.O.; Baptiste, K.E. Vancomycin-resistant Enterococci: A review of antimicrobial resistance mechanisms and perspectives of human and animal health. Microb. Drug Resist. 2018, 24, 590–606. [Google Scholar] [CrossRef] [PubMed]
- Ortez, J. Disk diffusion testing. In Manual of Antimicrobial Susceptibility Testing; Coyle, M.B., Ed.; American Society for Microbiology: Washington, DC, USA, 2005; pp. 39–52. [Google Scholar]
- Turner, S.; Pryer, K.M.; Miao, V.P.W.; Palmer, J.D. Investigating deep phylogenetic relationships among cyanobacteria and plastids by small subunit rRNA sequence analysis. J. Eukaryot. Microbiol. 1999, 46, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Young, S.; Juhl, A.; O’Mullan, G. Antibiotic-resistant bacteria in the Hudson River Estuary linked to wet weather sewage contamination. J. Water Health 2013, 11, 297–310. [Google Scholar] [CrossRef] [PubMed]
- Ash, R.J.; Mauck, B.; Morgan, M. Antibiotic resistance of gram-negative bacteria in rivers, United States. Emerg. Infect. Dis. 2002, 8, 713–716. [Google Scholar] [CrossRef]
- Guardabassi, L.; Lo Fo Wong, D.M.A.; Dalsgaard, A. The effects of tertiary wastewater treatment on the prevalence of antimicrobial resistant bacteria. Water Res. 2002, 36, 1955–1964. [Google Scholar] [CrossRef]
- West, B.M.; Liggit, P.; Clemans, D.L.; Francoeur, S.N. Antibiotic resistance, gene transfer, and water quality patterns observed in waterways near CAFO farms and wastewater treatment facilities. Water Air Soil Pollut. 2011, 217, 473–489. [Google Scholar] [CrossRef]
- Delgado-Gardea, M.; Tamez-Guerra, P.; Gomez-Flores, R.; Zavala-Díaz de la Serna, F.; Eroza-de la Vega, G.; Nevárez-Moorillón, G.; Pérez-Recoder, M.; Sánchez-Ramírez, B.; González-Horta, M.; Infante-Ramírez, R. Multidrug-resistant bacteria isolated from surface water in Bassaseachic Falls National Park, Mexico. Int. J. Environ. Res. Public Health 2016, 13, 597. [Google Scholar] [CrossRef] [PubMed]
- Odonkor, S.; Addo, K. Prevalence of multidrug-resistant Escherichia coli isolated from drinking water sources. Int. J. Microbiol. 2018, 2018, 7204013. [Google Scholar] [CrossRef] [PubMed]
- Auguet, O.; Pijuan, M.; Borrego, C.M.; Rodriguez-Mozaz, S.; Triadó-Margarit, X.; Giustina, S.V.D.; Gutierrez, O. Sewers as potential reservoirs of antibiotic resistance. Sci. Total Environ. 2017, 605, 1047–1054. [Google Scholar] [CrossRef] [PubMed]
- Almakki, A.; Jumas-Bilak, E.; Marchandin, H.; Licznar-Fajardo, P. Antibiotic resistance in urban runoff. Sci. Total Environ. 2019, 667, 64–76. [Google Scholar] [CrossRef]
- Karkman, A.; Do, T.T.; Walsh, F.; Virta, M.P.J. Antibiotic-resistance genes in waste water. Trends Microbiol. 2018, 3, 220–228. [Google Scholar] [CrossRef]
- Andersson, D.I.; Hughes, D. Persistence of antibiotic resistance in bacterial populations. FEMS Microbiol. Rev. 2011, 35, 901–911. [Google Scholar] [CrossRef]
- Barbosa, T.M.; Levy, S.B. The impact of antibiotic use on resistance development and persistence. Drug Resist. Updates 2000, 3, 303–311. [Google Scholar] [CrossRef]
- Martinez, J.L. Environmental pollution by antibiotics and by antibiotic resistance determinants. Environ. Pollut. 2009, 157, 2893–2902. [Google Scholar] [CrossRef]
- Barrón, M.D.L.C.; Merlin, C.; Guilloteau, H.; Montargès-Pelletier, E.; Bellanger, X. Suspended materials in river waters differentially enrich Class 1 Integron- and IncP-1 plasmid-carrying bacteria in sediments. Front. Microbiol. 2018, 9, 1443. [Google Scholar] [CrossRef]
- Kim, S.; Thiessen, P.A.; Bolton, E.E.; Chen, J.; Fu, G.; Gindulyte, A.; Han, L.; He, J.; He, S.; Shoemaker, B.A.; et al. PubChem Substance and Compound databases. Nucleic Acids Res. 2016, 44, D1202–D1213. [Google Scholar] [CrossRef]
- Shen, Y.; Zhao, W.; Zhang, C.; Shi, Y.S. Degradation of streptomycin in aquatic environment: Kinetics, pathway, and antibacterial activity analysis. Environ. Sci. Pollut. Res. 2017, 24, 14337. [Google Scholar] [CrossRef]
- Morris, D.; Galvin, S.; Boyle, F.; Hickey, P.; Mulligan, M.; Cormican, M. Enterococcus faecium of the vanA genotype in rural drinking water, effluent, and the aqueous environment. Appl. Environ. Microbiol. 2011, 78, 596–598. [Google Scholar] [CrossRef] [PubMed]
- Carsenti-Etesse, H.; Cavallo, J.; Roger, P.; Ziha-Zarifi, I.; Plesiat, P.; Garrabé, E.; Dellamonica, P. Effect of β-lactam antibiotics on the in vitro development of resistance in Pseudomonas aeruginosa. Clin. Microbiol. Infect. 2001, 7, 144–151. [Google Scholar] [CrossRef]
- Pezzella, C.; Ricci, A.; DiGiannatale, E.; Luzzi, I.; Carattoli, A. Tetracycline and streptomycin resistance genes, transposons, and plasmids in Salmonella enterica isolates from animals in Italy. Antimicrob. Agents Chemother. 2004, 48, 903–908. [Google Scholar] [CrossRef] [PubMed]
- Martins da Costa, P.M.; Vaz-Pires, P.M.; Bernardo, F.M. Antibiotic resistance of Enterococcus spp. isolated from wastewater and sludge of poultry slaughterhouses. J. Environ. Sci. Health Part B 2006, 41, 1393–1403. [Google Scholar] [CrossRef] [PubMed]
- Sanderson, H.; Ortega-Polo, R.; McDermott, K.; Hall, G.; Zaheer, R.; Brown, R.; Majury, A.; McAllister, T.; Liss, S. Quantification and multidrug resistance profiles of vancomycin-resistant Enterococci isolated from two wastewater treatment plants in the same municipality. Microorganisms 2019, 7, 626. [Google Scholar] [CrossRef]
- Martins da Costa, P.; Vaz-Pires, P.; Bernardo, F. Antimicrobial resistance in Enterococcus spp. isolated in inflow, effluent and sludge from municipal sewage water treatment plants. Water Res. 2006, 40, 1735–1740. [Google Scholar] [CrossRef]
- Tacão, M.; Moura, A.; Correia, A.; Henriques, I. Co-resistance to different classes of antibiotics among ESBL-producers from aquatic systems. Water Res. 2014, 48, 100–107. [Google Scholar] [CrossRef]
- Baindara, P. Mechanism of Bacterial Co-resistance. In Bacterial Adaptation to Co-Resistance; Mandal, S., Paul, D., Eds.; Springer: Singapore, 2019; pp. 191–210. [Google Scholar]
- Grenni, P.; Corno, G. Knowledge gaps and research needs in bacterial co-resistance in the environment. In Bacterial Adaptation to Co-Resistance; Mandal, S., Paul, D., Eds.; Springer: Singapore, 2019; pp. 39–59. [Google Scholar]
- Squadrone, S. Water environments: Metal-tolerant and antibiotic-resistant bacteria. Environ. Monit. Assess. 2020, 192, 1–12. [Google Scholar] [CrossRef]
- Marathe, N.P.; Shetty, S.A.; Shouche, Y.S.; Larsson, D.G.J. Limited bacterial diversity within a treatment plant receiving antibiotic-containing waste from bulk drug production. PLoS ONE 2016, 11, e0165914. [Google Scholar] [CrossRef] [PubMed]
- Calderon, O.; Porter-Morgan, H.; Jacob, J.; Elkins, W. Bacterial diversity impacts as a result of combined sewer overflow in a polluted waterway. Glob. J. Environ. Sci. Manag. 2017, 3, 437–446. [Google Scholar]
- Wang, X.; Gu, J.; Gao, H.; Qian, X.; Li, H. Abundances of clinically relevant antibiotic resistance genes and bacterial community diversity in the Weihe River, China. Int. J. Environ. Res. Public Health 2018, 15, 708. [Google Scholar] [CrossRef]
- Vaz-Moreira, I.; Nunes, O.C.; Manaia, C.M. Bacterial diversity and antibiotic resistance in water habitats: Searching the links with the human microbiome. FEMS Microbiol. Rev. 2014, 38, 761–778. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Qi, W.; Liang, J.; Qu, J. Using high-throughput sequencing to assess the impacts of treated and untreated wastewater discharge on prokaryotic communities in an urban river. Appl. Microbiol. Biotechnol. 2013, 98, 1841–1851. [Google Scholar] [CrossRef] [PubMed]
- Ibekwe, A.M.; Leddy, M.; Murinda, S.E. Potential human pathogenic bacteria in a mixed urban watershed as revealed by pyrosequencing. PLoS ONE 2013, 8, e79490. [Google Scholar] [CrossRef]
- Dantas, G.; Sommer, M.O.A.; Oluwasegun, R.D.; Church, G.M. Bacteria subsisting on antibiotics. Science 2008, 320, 100–103. [Google Scholar] [CrossRef] [PubMed]
- Forsberg, K.J.; Reyes, A.; Wang, B.; Selleck, E.M.; Sommer, M.O.A.; Dantas, G. The shared antibiotic resistome of soil bacteria and human pathogens. Science 2012, 337, 1107–1111. [Google Scholar] [CrossRef]
- D’costa, V.M.; Mcgrann, K.M.; Hughes, D.W.; Wright, G.D. Sampling the antibiotic resistome. Science 2006, 311, 374–377. [Google Scholar] [CrossRef] [PubMed]
- Riesenfeld, C.S.; Goodman, R.M.; Handelsman, J. Uncultured soil bacteria are a reservoir of new antibiotic resistance genes. Environ. Microbiol. 2004, 6, 981–989. [Google Scholar] [CrossRef]
- Hugon, P.; Dufour, J.-C.; Colson, P.; Fournier, P.-E.; Sallah, K.; Raoult, D. A comprehensive repertoire of prokaryotic species identified in human beings. Lancet Infect. Dis. 2015, 15, 1211–1219. [Google Scholar] [CrossRef]
- Glaeser, S.P.; Kämpfer, P. The Family Sphingomonadaceae. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 641–707. [Google Scholar]
- Abraham, W.R.; Rohde, M.; Bennasar, A. The Family Caulobacteraceae. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 179–205. [Google Scholar]
- Ariskina, E.V.; Chernousova, E.Y.; Lapteva, N.A.; Akimov, V.N. Evaluation of the taxonomic diversity of prosthecate bacteria belonging to the genera Brevundimonas and Caulobacter isolated from various Eurasian ecosystems by analysis of the 16S rRNA genes. Microbiology 2011, 80, 403–410. [Google Scholar] [CrossRef]
- Gensberger, E.; Gössl, E.; Antonielli, L.; Sessitsch, A.; Kostić, T. Effect of different heterotrophic plate count methods on the estimation of the composition of the culturable microbial community. PeerJ 2015, 3, 862. [Google Scholar] [CrossRef]
- Reasoner, D.J. Heterotrophic plate count methodology in the United States. Int. J. Food Microbiol. 2004, 92, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Coutinho, F.H.; Silveira, C.B.; Pinto, L.H.; Salloto, G.R.B.; Cardoso, A.M.; Martins, O.B.; Vieira, R.P.; Clementino, M.M. Antibiotic resistance is widespread in urban aquatic environments of Rio de Janeiro, Brazil. Microb. Ecol. 2014, 68, 441–452. [Google Scholar] [CrossRef]
- Brooke, J.S. Stenotrophomonas maltophilia: An emerging global opportunistic pathogen. Clin. Microbiol. Rev. 2012, 25, 2–41. [Google Scholar] [CrossRef] [PubMed]
- Leneveu-Jenvrin, C.; Madi, A.; Bouffartigues, E.; Biaggini, K.; Feuilloley, M.; Chevalier, S.; Connil, N. Cytotoxicity and inflammatory potential of two Pseudomonas mosselii strains isolated from clinical samples of hospitalized patients. BMC Microbiol. 2013, 13, 123. [Google Scholar] [CrossRef] [PubMed]
- Wong, V.; Levi, K.; Baddal, B.; Turton, J.; Boswell, T.C. Spread of Pseudomonas fluorescens due to contaminated drinking water in a bone marrow transplant unit. J. Clin. Microbiol. 2011, 49, 2093–2096. [Google Scholar] [CrossRef] [PubMed]
- Abdalhamid, B.; Elhadi, N.; Alsamman, K.; Aljindan, R. Chryseobacterium gleum pneumonia in an infant with nephrotic syndrome. IDCases 2016, 5, 34–36. [Google Scholar] [CrossRef] [PubMed]
- Loch, T.P.; Faisal, M. Emerging flavobacterial infections in fish: A review. J. Adv. Res. 2015, 6, 283–300. [Google Scholar] [CrossRef] [PubMed]
- Wen, H.; Wang, K.; Liu, Y.; Tay, M.; Lauro, F.M.; Huang, H.; Wu, H.; Liang, H.; Ding, Y.; Givskov, M.; et al. Population dynamics of an Acinetobacter baumannii clonal complex during colonization of patients. J. Clin. Microbiol. 2014, 52, 3200–3208. [Google Scholar] [CrossRef]
- Igbinosa, I.H.; Igumbor, E.U.; Aghdasi, F.; Tom, M.; Okoh, A.I. Emerging Aeromonas species infections and their significance in public health. Sci. World J. 2012, 2012, 625023. [Google Scholar] [CrossRef]
- Finger, H.; von Koenig, C.H.W. Chapter 31: Bordetella. In Medical Microbiology, 4th ed.; Baron, S., Ed.; University of Texas Medical Branch at Galveston: Galveston, TX, USA, 1996. [Google Scholar]
- Ouyang, J.; Pei, Z.; Lutwick, L.; Dalal, S.; Yang, L.; Cassai, N.; Sandhu, K.; Hanna, B.; Wieczorek, R.; Bluth, M.; et al. Case report: Paenibacillus thiaminolyticus: A new cause of human infection, inducing bacteremia in a patient on hemodialysis. Ann. Clin. Lab. Sci. 2008, 38, 393–400. [Google Scholar] [PubMed]
- Batista, J.H.; Neto, J.F.D.S. Chromobacterium violaceum pathogenicity: Updates and insights from genome sequencing of novel Chromobacterium species. Front. Microbiol. 2017, 8, 2213. [Google Scholar] [CrossRef] [PubMed]
- Kämpfer, P.; Denner, E.B.M.; Witzenberger, R.; Neef, A.; Busse, H.-J. Sphingopyxis witflariensis sp. nov., isolated from activated sludge. Int. J. Syst. Evol. Microbiol. 2002, 52, 2029–2034. [Google Scholar]
- Bilgin, H.; Sarmis, A.; Tigen, E.; Soyletir, G.; Mulazimoglu, L. Delftia Acidovorans: A rare pathogen in immunocompetent and immunocompromised patients. Can. J. Infect. Dis. Med. Microbiol. 2015, 26, 277–279. [Google Scholar] [CrossRef]
- Bayhan, G.I.; Tanir, G.; Karaman, I.; Ozkan, S. Comamonas testosteroni: An unusual bacteria associated with acute appendicitis. Balk. Med. J. 2013, 30, 447–448. [Google Scholar] [CrossRef] [PubMed]
- Haynes, J.; Hawkey, P.M. Providencia alcalifaciens and travellers diarrhoea. BMJ 1989, 299, 94. [Google Scholar] [CrossRef]
- Xi, C.; Zhang, Y.; Marrs, C.F.; Ye, W.; Simon, C.; Foxman, B.; Nriagu, J. Prevalence of antibiotic resistance in drinking water treatment and distribution systems. Appl. Environ. Microbiol. 2009, 75, 5714–5718. [Google Scholar] [CrossRef]
- Vaz-Moreira, I.; Nunes, O.C.; Manaia, C.M. Diversity and antibiotic resistance patterns of Sphingomonadaceae isolates from drinking water. Appl. Environ. Microbiol. 2011, 77, 5697–5706. [Google Scholar] [CrossRef] [PubMed]
- Vaz-Moreira, I.; Nunes, O.C.; Manaia, C.M. Diversity and antibiotic resistance in Pseudomonas spp. from drinking water. Sci. Total Environ. 2012, 426, 366–374. [Google Scholar] [CrossRef]
- Figueira, V.; Serra, E.A.; Vaz-Moreira, I.; Brandão, T.R.S.; Manaia, C.M. Comparison of ubiquitous antibiotic-resistant Enterobacteriaceae populations isolated from wastewaters, surface waters and drinking waters. J. Water Health 2011, 10, 1–10. [Google Scholar] [CrossRef]
- Narciso-Da-Rocha, C.; Vaz-Moreira, I.; Svensson-Stadler, L.; Moore, E.R.B.; Manaia, C.M. Diversity and antibiotic resistance of Acinetobacter spp. in water from the source to the tap. Appl. Microbiol. Biotechnol. 2012, 97, 329–340. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Suits, M.; Wituszynski, D.; Winston, R.; Martin, J.; Lee, J. Residential urban stormwater runoff: A comprehensive profile of microbiome and antibiotic resistance. Sci. Total Environ. 2020, 723, 138033. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, T.; Vaz-Moreira, I.; Manaia, C.M. Neighbor urban wastewater treatment plants display distinct profiles of bacterial community and antibiotic resistance genes. Environ. Sci. Pollut. Res. 2019, 26, 11269–11278. [Google Scholar] [CrossRef] [PubMed]
- Saunders, A.M.; Albertsen, M.; Vollertsen, J.; Nielsen, P.H. The activated sludge ecosystem contains a core community of abundant organisms. ISME J. 2016, 10, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Wan, Y.; Mao, Y.; Zhang, T. Partnership of Arthrobacter and Pimelobacter in aerobic degradation of sulfadiazine revealed by metagenomics analysis and isolation. Environ. Sci. Technol. 2018, 52, 2963–2972. [Google Scholar] [CrossRef]
- Curriero, F.C.; Patz, J.A.; Rose, J.B.; Lele, S. The association between extreme precipitation and waterborne disease outbreaks in the United States, 1948–1994. Am. J. Public Health 2001, 91, 1194–1199. [Google Scholar] [CrossRef] [PubMed]
- Nichols, G.; Lane, C.; Asgari, N.; Verlander, N.Q.; Charlett, A. Rainfall and outbreaks of drinking water related disease and in England and Wales. J. Water Health 2009, 7, 1–8. [Google Scholar] [CrossRef]
- Rose, J.; Daeschner, S.; Easterling, D.; Curriero, F.; Lele, S.; Patz, J.A. Climate and waterborne disease outbreaks. J. Am. Water Work. Assoc. 2000, 92, 77–87. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Upstream Clear Weather | CSO Site Clear Weather | CSO Site Overflow Event | |
|---|---|---|---|
| River discharge (m3 s−1) | 160 | 160 | 400 |
| Rainfall during 12 h prior to sampling (cm) | <0.1 | <0.1 | 2.5 |
| Water temperature (°C) | 25.6 | 26.5 | 24.7 |
| pH | 7.3 | 7.7 | 8.1 |
| Dissolved oxygen (mg L−1) | 8.6 | 8.4 | 8.5 |
| Specific Conductance (µS cm−1) | 140.0 | 139.0 | 112.0 |
| Turbidity (NTU) | 7.6 | 14.3 | 230.0 |
| Bacteria abundance (CFU mL−1) | 1.5 × 105 | 6.4 × 105 | 4.2 × 105 |
| E. coli abundance (CFU 100-mL−1) | 103 | 188 | 370 |
| Classification | Upstream Clear Weather | CSO Site Clear Weather | CSO Site Overflow Event |
|---|---|---|---|
| Genus Bordetella | 2.1 (5.0) | ||
| Chromobacterium violaceum | 2.1 (4.0) | ||
| Sphingobacterium multivorum | 2.1 (4.0) | ||
| Aeromonas hydrophila | 2.1 (1.0) | 2.1 (3.0) | |
| Genus Pseudomonas † | 2.1 (4.0) | 8.5 (7.8) | 10.6 (4.6) |
| Genus Chryseobacterium ‡ | 12.8 (7.6) | ||
| Bacillus cereus F837/76 | 4.3 (6.0) | ||
| Providencia alcalifaciens | 4.3 (4.0) | ||
| Acinetobacter baumannii | 2.1 (4.0) | ||
| Comamonas testosteroni | 2.1 (3.0) | ||
| Acidovorax temperans | 4.3 (5.5) | ||
| Stenotrophomonas maltophilia | 27.7 (7.7) | ||
| Delftia acidovorans | 2.1 (4.0) | 8.5 (6.0) | |
| Total | 12.6 (3.6) | 40.5 (6.2) | 46.8 (6.7) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balasa, G.; Levengood, E.S.; Battistelli, J.M.; Franklin, R.B. Diversity of Multidrug-Resistant Bacteria in an Urbanized River: A Case Study of the Potential Risks from Combined Sewage Overflows. Water 2021, 13, 2122. https://doi.org/10.3390/w13152122
Balasa G, Levengood ES, Battistelli JM, Franklin RB. Diversity of Multidrug-Resistant Bacteria in an Urbanized River: A Case Study of the Potential Risks from Combined Sewage Overflows. Water. 2021; 13(15):2122. https://doi.org/10.3390/w13152122
Chicago/Turabian StyleBalasa, Gabriella, Enjolie S. Levengood, Joseph M. Battistelli, and Rima B. Franklin. 2021. "Diversity of Multidrug-Resistant Bacteria in an Urbanized River: A Case Study of the Potential Risks from Combined Sewage Overflows" Water 13, no. 15: 2122. https://doi.org/10.3390/w13152122