
Seasonal Variations in the Biochemical Compositions of Phytoplankton and Transparent Exopolymer Particles (TEPs) at Jang Bogo Station (Terra Nova Bay, Ross Sea), 2017–2018

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
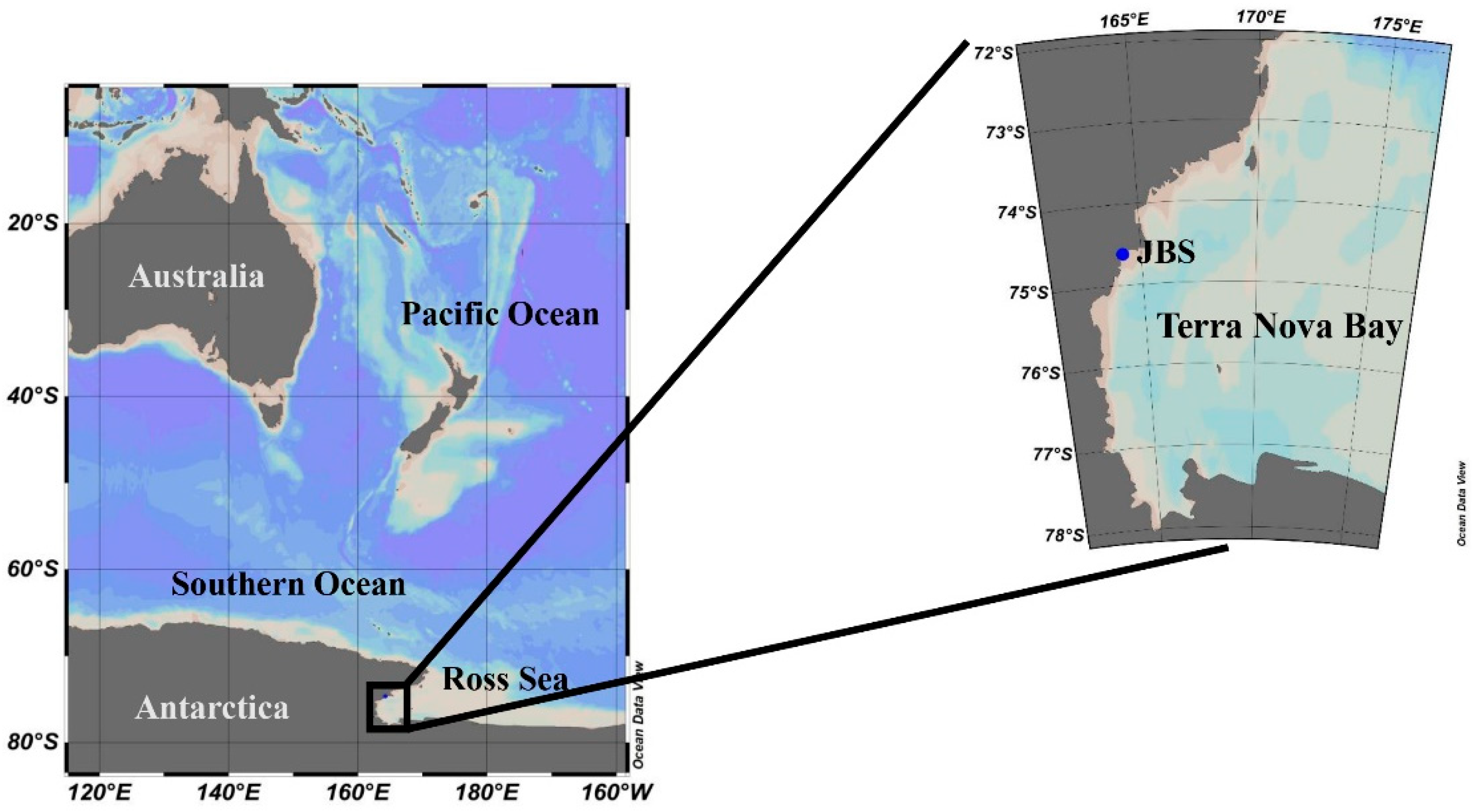
2.1. Study Area and Water Sampling
2.2. Nutrient Analysis
2.3. Chlorophyll-a Analysis
2.4. Particulate Organic Carbon and Nitrogen Analysis
2.5. Phytoplankton Community
2.6. Macromolecular Compositions
2.7. TEP Analysis
2.8. Data Treatment and Statistical Analyses
3. Results
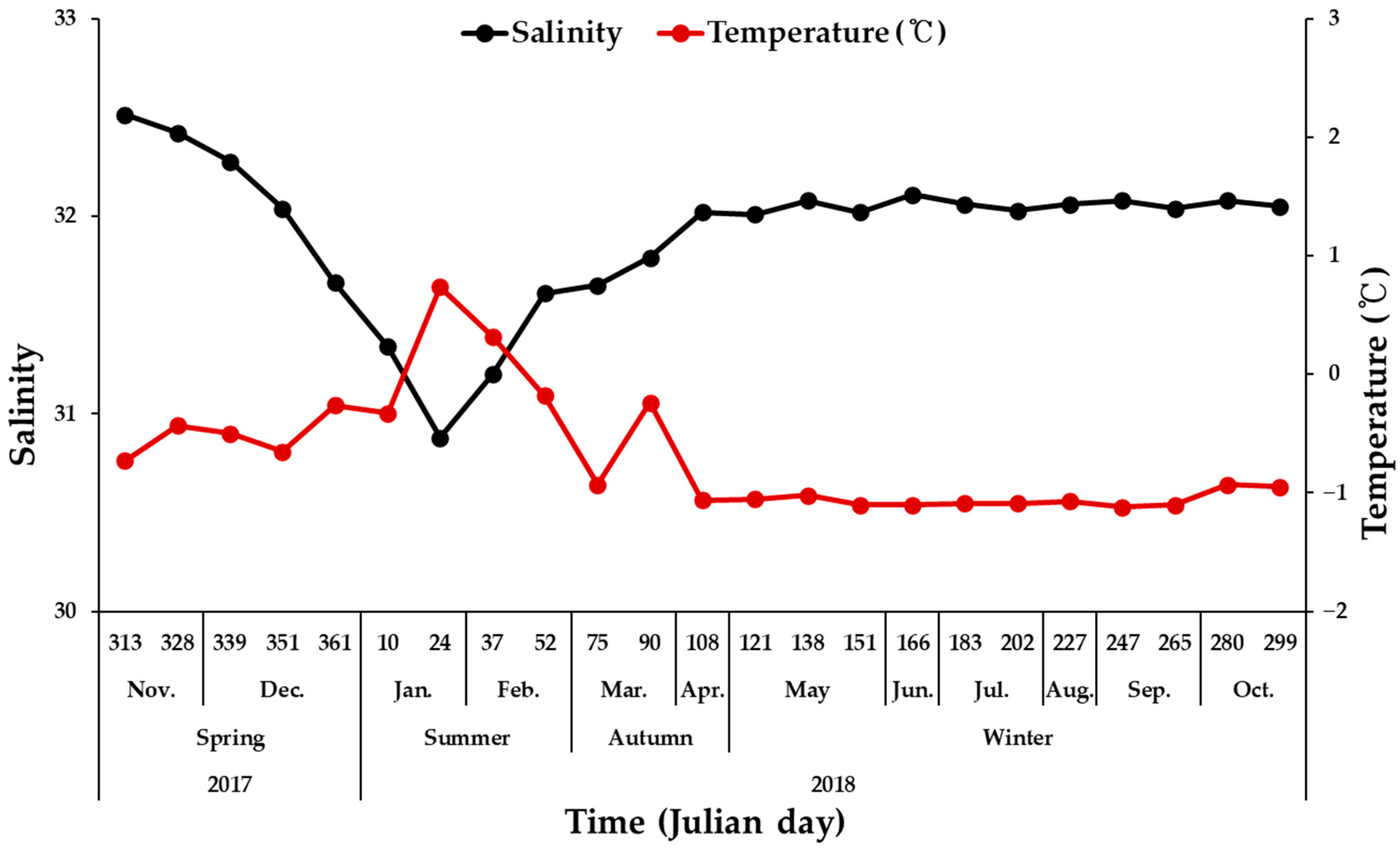
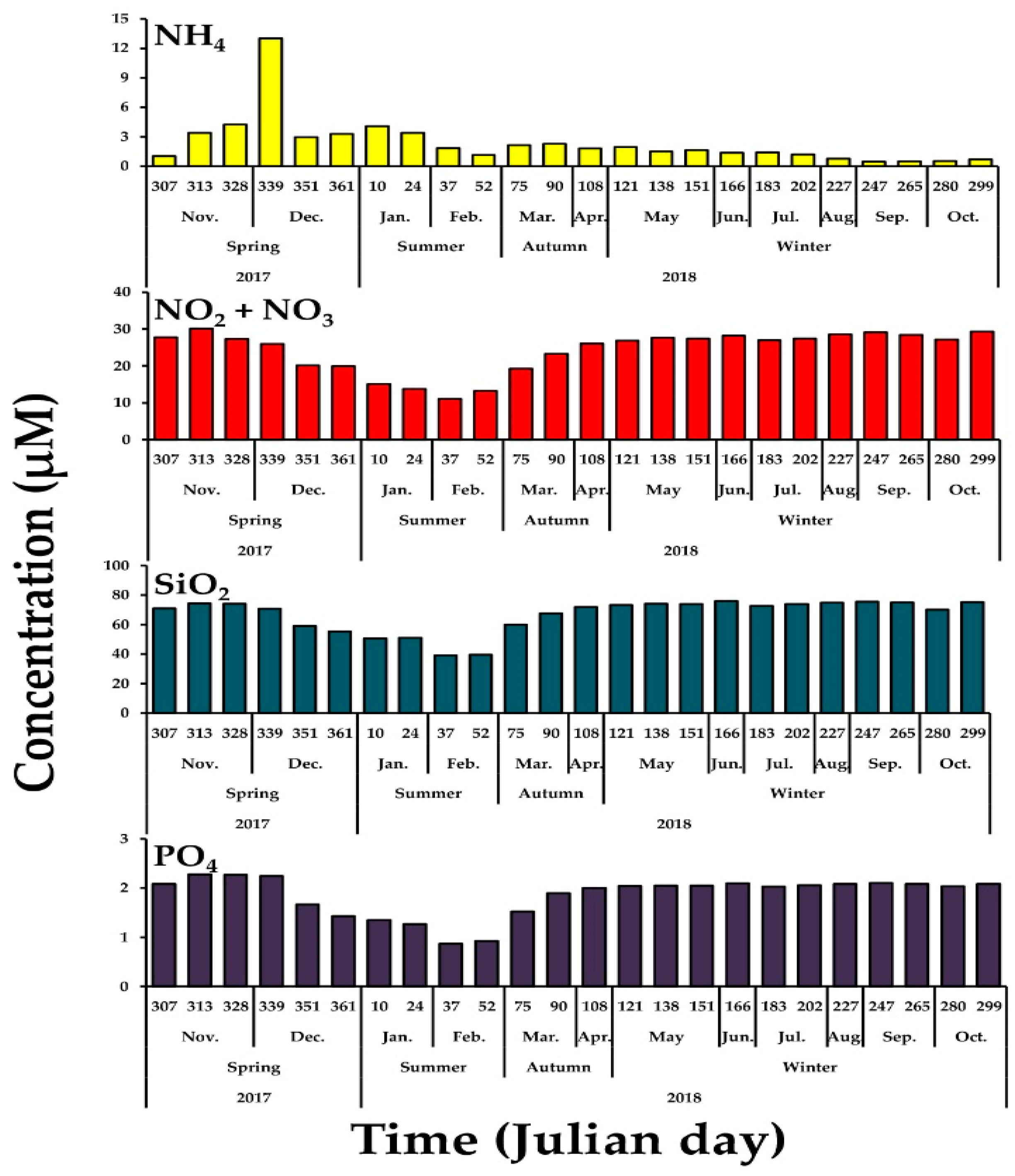
3.1. Physical and Chemical Conditions in the Study Area
3.2. Chl-a Concentrations
3.3. POC and PON
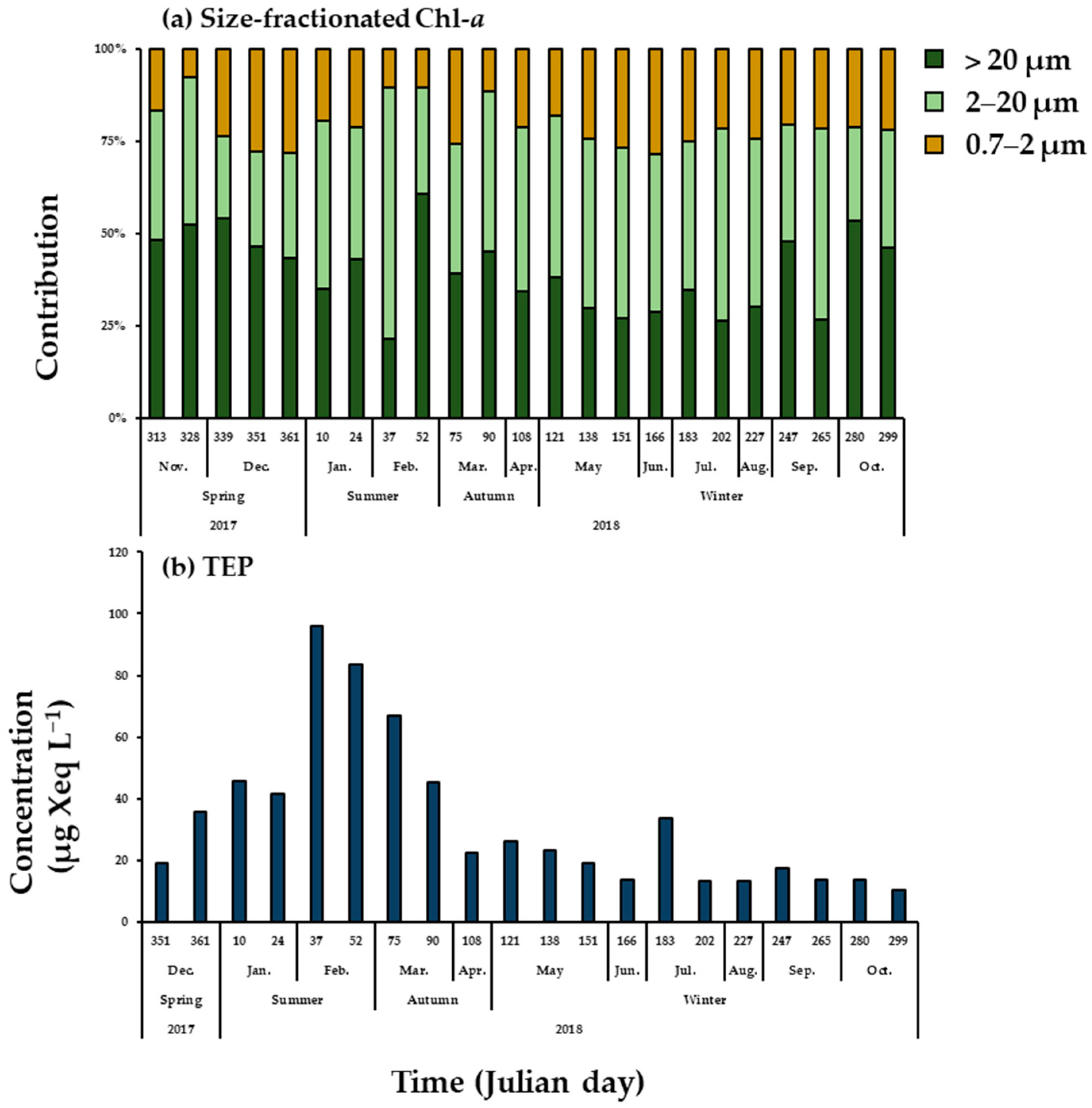
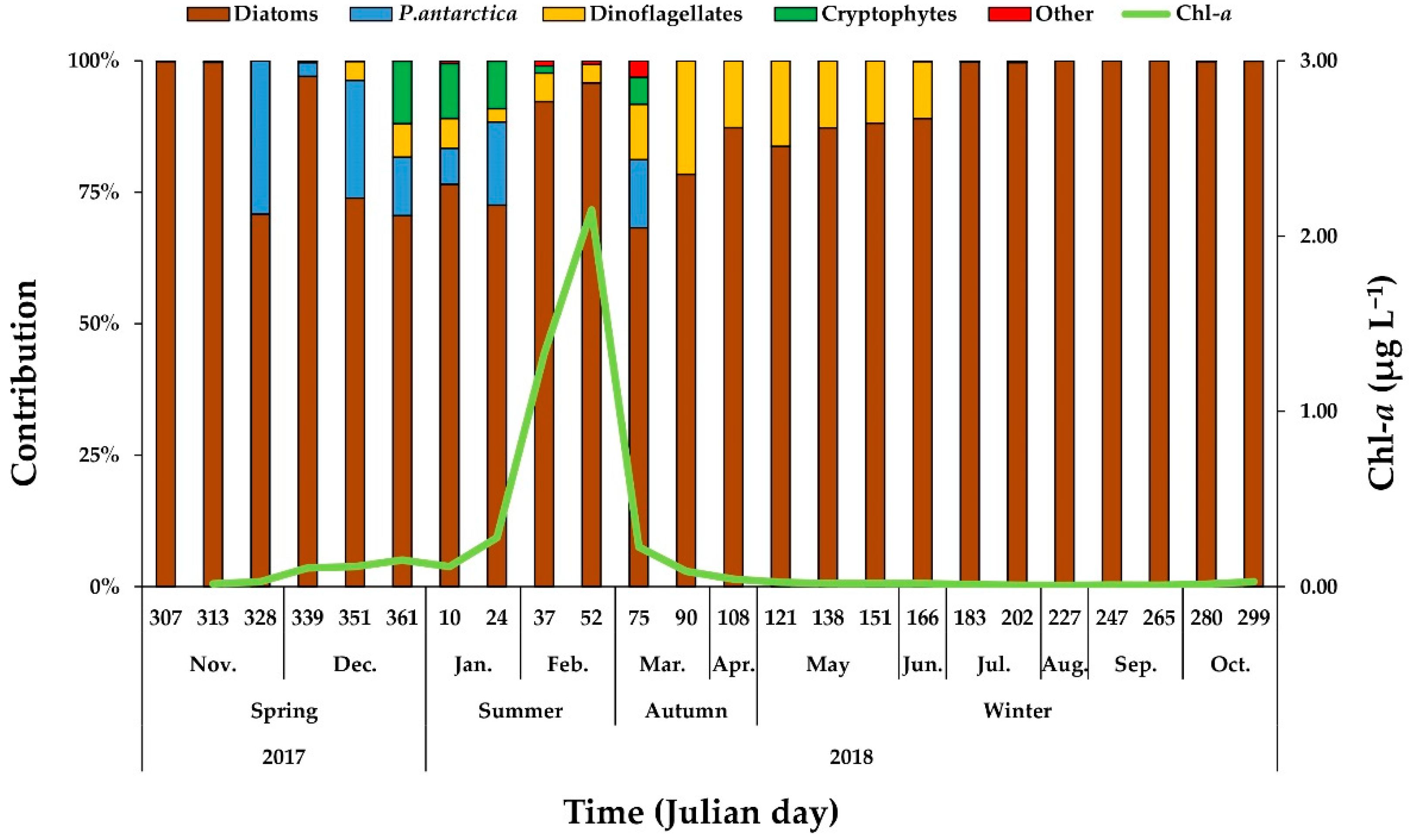
3.4. Phytoplankton Community Composition
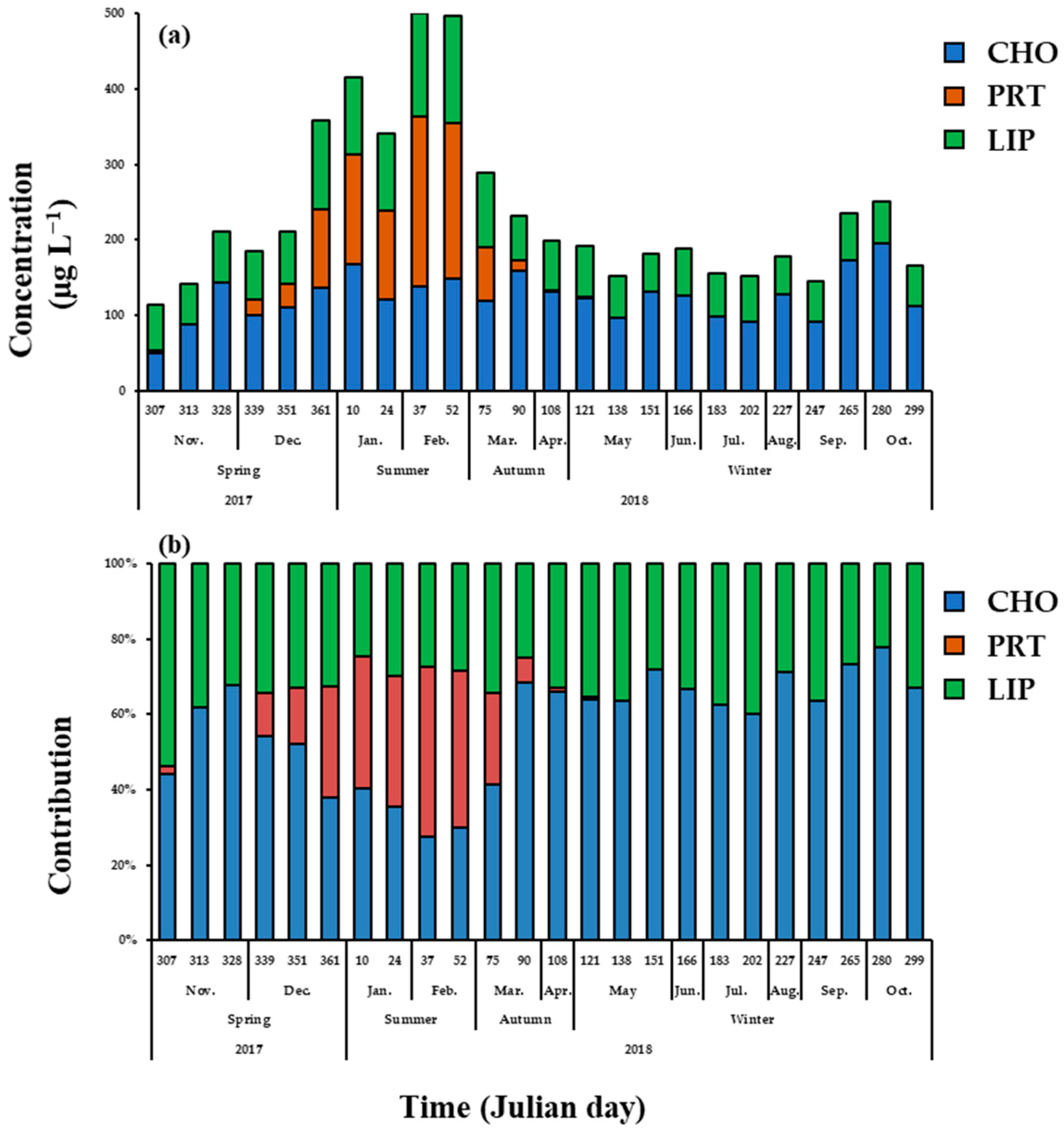
3.5. Macromolecular Composition of Phytoplankton
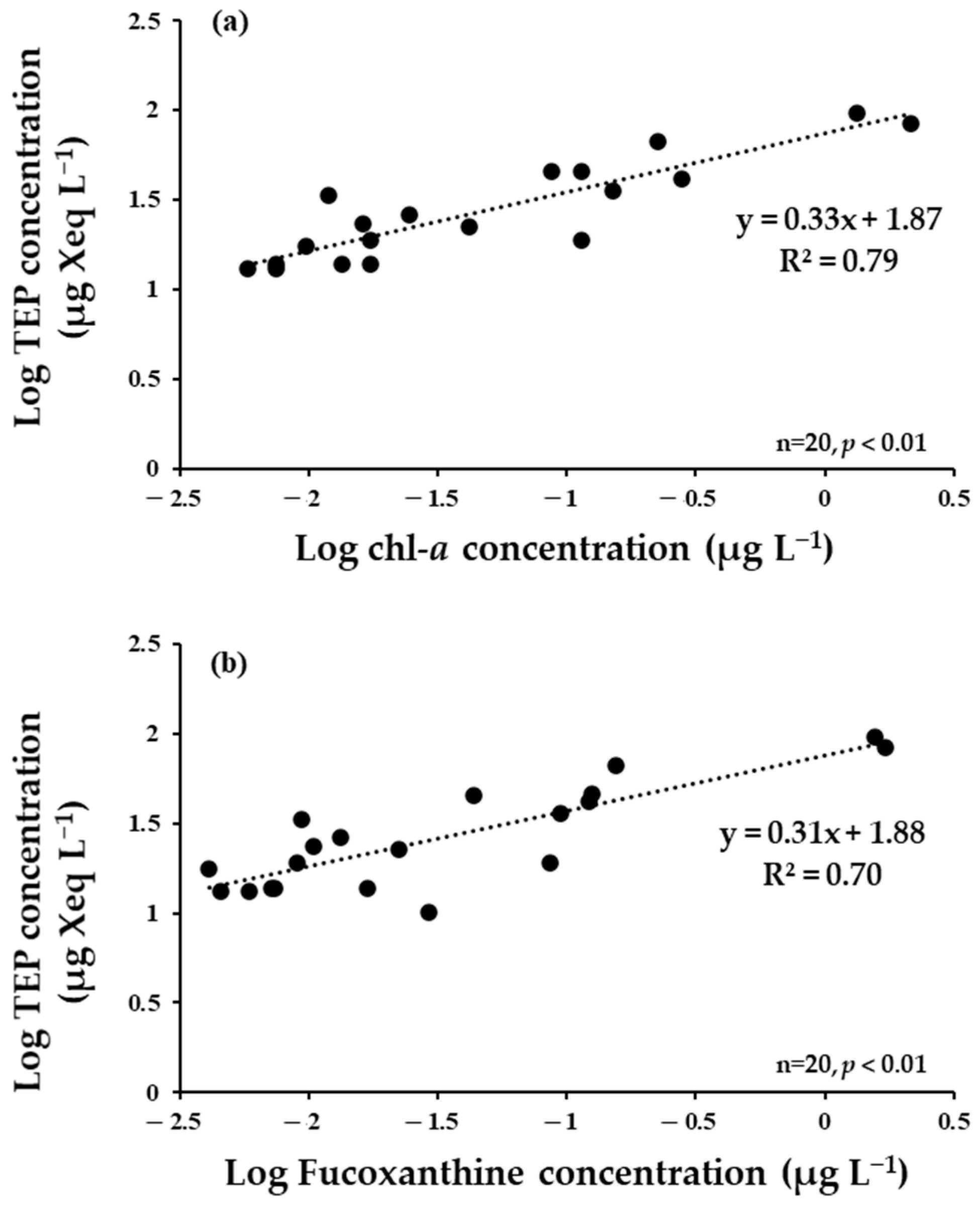
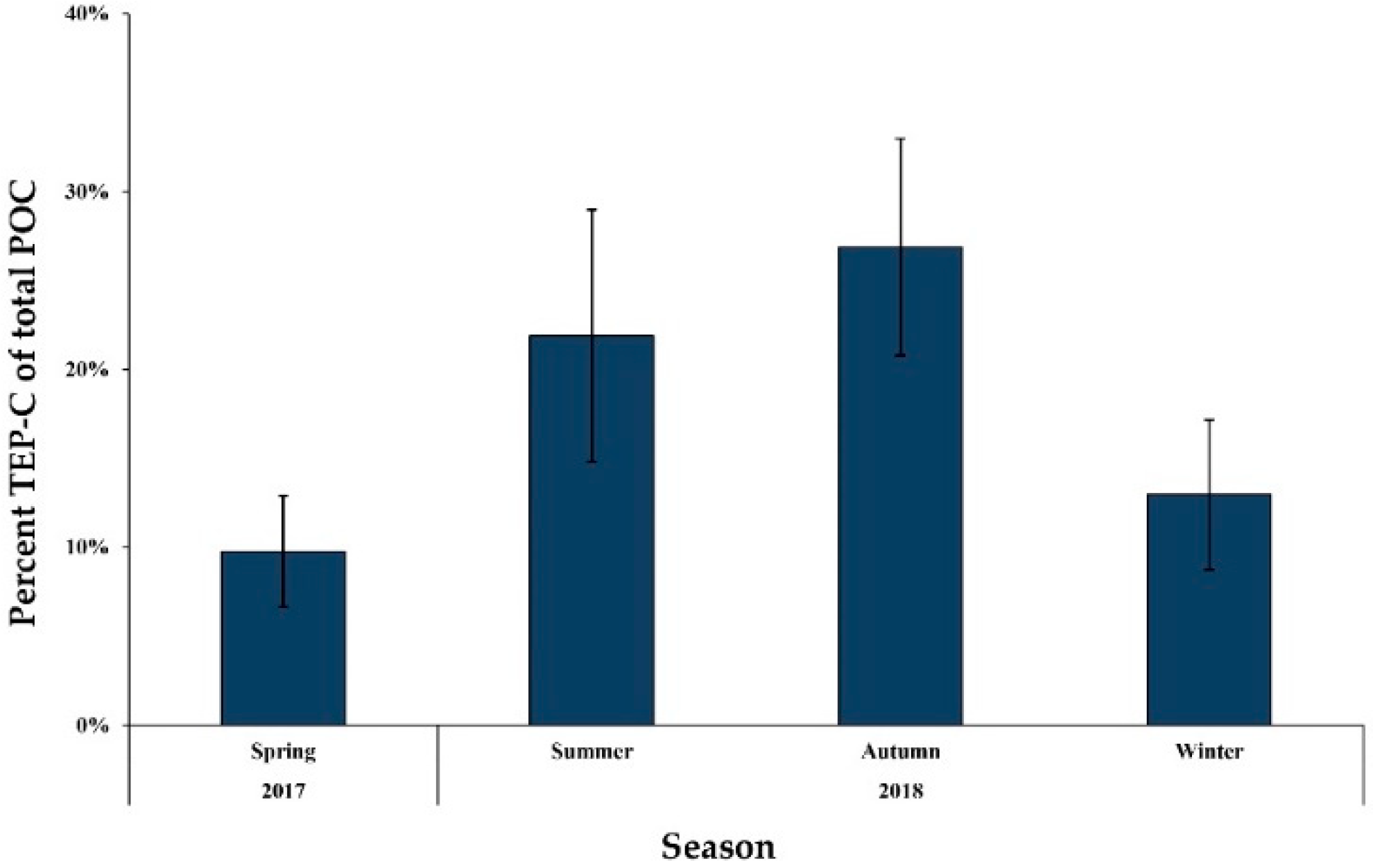
3.6. TEP Concentration
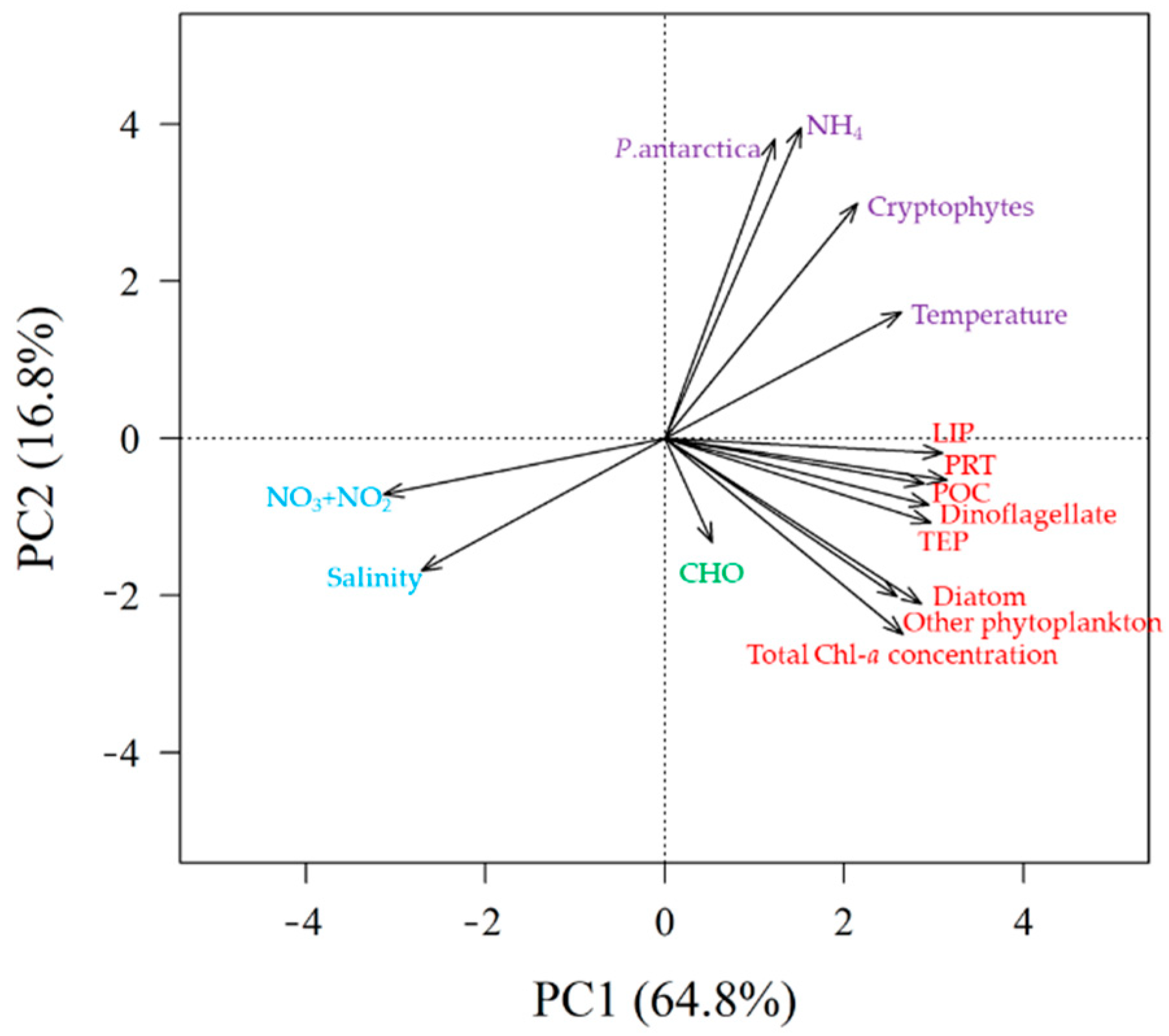
3.7. PCA
4. Discussion
4.1. Environmental Conditions
4.2. Seasonal Variations in Biomass and Major Species Compositions of Phytoplankton
4.3. Seasonal Variations of Macromolecular Composition
4.4. Seasonal Variations of TEPs
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nelson, D.M.; DeMaster, D.J.; Dunbar, R.B.; Smith, W.O. Cycling of organic carbon and biogenic silica in the Southern Ocean: Estimates of water-column and sedimentary fluxes on the Ross Sea continental shelf. J. Geophys. Res. C Ocean. 1996, 101, 18519–18532. [Google Scholar] [CrossRef]
- Smith, W.O.; Nelson, D.M. Phytoplankton bloom produced by a receding ice edge in the ross sea: Spatial coherence with the density field. Science 1985, 227, 163–166. [Google Scholar] [CrossRef]
- Sedwick, P.N.; Ditullio, G.R. Regulation of algal blooms in Antarctic shelf waters by the release of iron from melting sea ice. Geophys. Res. Lett. 1997, 24, 2515–2518. [Google Scholar] [CrossRef] [Green Version]
- Sedwick, P.N.; Di Tullio, G.R.; Mackey, D.J. Iron and manganese in the Ross Sea, Seasonal iron limitation in Antarctic: Seasonal iron limitation in Antarctic shelf waters. J. Geophys. Res. Ocean. 2000, 105, 11321–11336. [Google Scholar] [CrossRef]
- Krell, A.; Schnack-Schiel, S.B.; Thomas, D.N.; Kattner, G.; Zipan, W.; Dieckmann, G.S. Phytoplankton dynamics in relation to hydrography, nutrients and zooplankton at the onset of sea ice formation in the eastern Weddell Sea (Antarctica). Polar Biol. 2005, 28, 700–713. [Google Scholar] [CrossRef]
- Garrison, D.L.; Buck, K.R. Surface-layer sea ice assemblages in Antarctic pack ice during the austral spring: Environmental conditions, primary production and community structure. Mar. Ecol. Prog. Ser. 1991, 75, 161–172. [Google Scholar] [CrossRef]
- Smetacek, V.; Scharek, R.; Gordon, L.I.; Eicken, H.; Fahrbach, E.; Rohardt, G.; Moore, S. Early spring phytoplankton blooms in ice platelet layers of the southern Weddell Sea, Antarctica. Deep Sea Res. Part A Oceanogr. Res. Pap. 1992, 39, 153–168. [Google Scholar] [CrossRef]
- Brierley, A.S.; Thomas, D.N. Ecology of Southern Ocean pack ice. Adv. Mar. Biol. 2002, 43, 171–276. [Google Scholar]
- Parkinson, C.L.; Cavalieri, D.J. Antarctic sea ice variability and trends, 1979–2010. Cryosphere 2012, 6, 871–880. [Google Scholar] [CrossRef] [Green Version]
- Eayrs, C.; Holland, D.; Francis, D.; Wagner, T.; Kumar, R.; Li, X. Understanding the Seasonal Cycle of Antarctic Sea Ice Extent in the Context of Longer-Term Variability. Rev. Geophys. 2019, 57, 1037–1064. [Google Scholar] [CrossRef]
- Goffart, A.; Catalano, G.; Hecq, J.H. Factors controlling the distribution of diatoms and Phaeocystis in the Ross Sea. J. Mar. Syst. 2000, 27, 161–175. [Google Scholar] [CrossRef]
- Grotti, M.; Soggia, F.; Ianni, C.; Frache, R. Trace metals distributions in coastal sea ice of Terra Nova Bay, Ross Sea, Antarctica. Antarct. Sci. 2005, 17, 289–300. [Google Scholar] [CrossRef]
- Smith, W.O.; Shields, A.R.; Peloquin, J.A.; Catalano, G.; Tozzi, S.; Dinniman, M.S.; Asper, V.A. Interannual variations in nutrients, net community production, and biogeochemical cycles in the Ross Sea. Deep Res. Part II Top. Stud. Oceanogr. 2006, 53, 815–833. [Google Scholar] [CrossRef]
- Finkel, Z.V.; Follows, M.J.; Liefer, J.D.; Brown, C.M.; Benner, I.; Irwin, A.J. Phylogenetic diversity in the macromolecular composition of microalgae. PLoS ONE 2016, 11, e0155977. [Google Scholar] [CrossRef] [Green Version]
- Ben-Amotz, A.; Tornabene, T.G.; Thomas, W.H. Chemical Profile of Selected Species of Microalgae With Emphasis on Lipids. J. Phycol. 1985, 21, 72–81. [Google Scholar] [CrossRef]
- Lee, S.H.; Kim, H.J.; Whitledge, T.E. High incorporation of carbon into proteins by the phytoplankton of the Bering Strait and Chukchi Sea. Cont. Shelf Res. 2009, 29, 1689–1696. [Google Scholar] [CrossRef]
- Smith, W.O.; Marra, J.; Hiscock, M.R.; Barber, R.T. The seasonal cycle of phytoplankton biomass and primary productivity in the Ross Sea, Antarctica. Deep Res. Part II Top. Stud. Oceanogr. 2000, 47, 3119–3140. [Google Scholar] [CrossRef]
- Olson, R.J.; Sosik, H.M.; Chekalyuk, A.M.; Shalapyonok, A. Effects of iron enrichment on phytoplankton in the Southern Ocean during late summer: Active fluorescence and flow cystometric analyses. Deep Res. Part II Top. Stud. Oceanogr. 2000, 47, 3181–3200. [Google Scholar] [CrossRef]
- Fabiano, M.; Povero, P.; Danovaro, R. Participate organic matter composition in Terra Nova Bay (Ross Sea, Antarctica) during summer 1990. Antarct. Sci. 1996, 8, 7–13. [Google Scholar] [CrossRef]
- Fabiano, M.; Pusceddu, A. Total and hydrolizable particulate organic matter (carbohydrates, proteins and lipids) at a coastal station in Terra Nova Bay (Ross Sea, Antarctica). Polar Biol. 1998, 19, 125–132. [Google Scholar] [CrossRef]
- Pusceddu, A.; Cattaneo-Vietti, R.; Albertelli, G.; Fabiano, M. Origin, biochemical composition and vertical flux of particulate organic matter under the pack ice in Terra Nova Bay (Ross Sea, Antarctica) during late summer 1995. Polar Biol. 1999, 22, 124–132. [Google Scholar] [CrossRef]
- Baldi, F.; Marchetto, D.; Pini, F.; Fani, R.; Michaud, L.; Lo Giudice, A.; Berto, D.; Giani, M. Biochemical and microbial features of shallow marine sediments along the Terra Nova Bay (Ross Sea, Antarctica). Cont. Shelf Res. 2010, 30, 1614–1625. [Google Scholar] [CrossRef]
- Kim, K.; Park, J.; Jo, N.; Park, S.; Yoo, H.; Kim, J.; Lee, S.H. Monthly Variation in the Macromolecular Composition of Phytoplankton Communities at Jang Bogo Station, Terra Nova Bay, Ross Sea. Front. Microbiol. 2021, 12, 618999. [Google Scholar] [CrossRef]
- Alldredge, A.L.; Passow, U.; Logan, B.E. The abundance and significance of a class of large, transparent organic particles in the ocean. Deep Sea Res. Part I Ocean. Res. Pap. 1993, 40, 1131–1140. [Google Scholar] [CrossRef]
- Passow, U. Production of transparent exopolymer particles (TEPS) by phyto- and bacterioplankton. Mar. Ecol. Prog. Ser. 2002, 236, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Passow, U. Formation of transparent exopolymer particles, TEPS, from dissolved precursor material. Mar. Ecol. Prog. Ser. 2000, 192, 1–11. [Google Scholar] [CrossRef]
- Mari, X.; Passow, U.; Migon, C.; Burd, A.B.; Legendre, L. Transparent exopolymer particles: Effects on carbon cycling in the ocean. Prog. Oceanogr. 2017, 151, 13–37. [Google Scholar] [CrossRef] [Green Version]
- Passow, U.; Shipe, R.F.; Murray, A.; Pak, D.K.; Brzezinski, M.A.; Alldredge, A.L. The origin of transparent exopolymer particles (TEPS) and their role in the sedimentation of particulate matter. Cont. Shelf Res. 2001, 21, 327–346. [Google Scholar] [CrossRef] [Green Version]
- Ortega-Retuerta, E.; Reche, I.; Pulido-Villena, E.; Agustí, S.; Duarte, C.M. Uncoupled distributions of transparent exopolymer particles (TEPS) and dissolved carbohydrates in the Southern Ocean. Mar. Chem. 2009, 115, 59–65. [Google Scholar] [CrossRef]
- Mari, X.; Rassoulzadegan, F.; Brussaard, C.P.D.; Wassmann, P. Dynamics of transparent exopolymeric particles (TEPS) production by Phaeocystis globosa under N- or P-limitation: A controlling factor of the retention/export balance. Harmful Algae 2005, 4, 895–914. [Google Scholar] [CrossRef]
- Beauvais, S.; Pedrotti, M.L.; Egge, J.; Iversen, K.; Marrasé, C. Effects of turbulence on TEPS dynamics under contrasting nutrient conditions: Implications for aggregation and sedimentation processes. Mar. Ecol. Prog. Ser. 2006, 323, 47–57. [Google Scholar] [CrossRef] [Green Version]
- Hung, C.C.; Guo, L.; Santschi, P.H.; Alvarado-Quiroz, N.; Haye, J.M. Distributions of carbohydrate species in the Gulf of Mexico. Mar. Chem. 2003, 81, 119–135. [Google Scholar] [CrossRef]
- Passow, U. Transparent exopolymer particles (TEPS) in aquatic environments. Prog. Oceanogr. 2002, 55, 287–333. [Google Scholar] [CrossRef] [Green Version]
- Zamanillo, M.; Ortega-Retuerta, E.; Nunes, S.; Estrada, M.; Sala, M.M.; Royer, S.J.; López-Sandoval, D.C.; Emelianov, M.; Vaqué, D.; Marrasé, C.; et al. Distribution of transparent exopolymer particles (TEPS) in distinct regions of the Southern Ocean. Sci. Total Environ. 2019, 691, 736–748. [Google Scholar] [CrossRef] [Green Version]
- Passow, U.; Alldredge, A.L. A dye-binding assay for the spectrophotometric measurement of transparent exopolymer particles (TEPS). Limnol. Oceanogr. 1995, 40, 1326–1335. [Google Scholar] [CrossRef]
- Hong, Y.; Smith, W.O.; White, A.M. Studies on transparent exopolymer particles (TEPS) produced in the ross sea (Antarctica) and by Phaeocystis antarctica (Prymnesiophyceae). J. Phycol. 1997, 33, 368–376. [Google Scholar] [CrossRef]
- Sugimoto, K.; Fukuda, H.; Baki, M.A.; Koike, I. Bacterial contributions to formation of transparent exopolymer particles (TEPS) and seasonal trends in coastal waters of Sagami Bay, Japan. Aquat. Microb. Ecol. 2007, 46, 31–41. [Google Scholar] [CrossRef]
- Huertas, E.; Navarro, G.; Rodríguez-Gálvez, S.; Prieto, L. The influence of phytoplankton biomass on the spatial distribution of carbon dioxide in surface sea water of a coastal area of the Gulf of Cádiz (southwestern Spain). Can. J. Bot. 2005, 83, 929–940. [Google Scholar] [CrossRef]
- Parsons, T.R.; Maita, Y.; Lalli, C.M. A Manual of Chemical and Biological Methods for Seawater Analysis; Pergamon Press: New York, NY, USA, 1984. [Google Scholar]
- Mackey, M.D.; Mackey, D.J.; Higgins, H.W.; Wright, S.W. CHEMTAX—A program for estimating class abundances from chemical markers: Application to HPLC measurements of phytoplankton. Mar. Ecol. Prog. Ser. 1996, 144, 265–283. [Google Scholar] [CrossRef] [Green Version]
- Wright, S.W.; Thomas, D.P.; Marchant, H.J.; Higgins, H.W.; Mackey, M.D.; Mackey, D.J. Analysis of phytoplankton of the Australian sector of the Southern Ocean: Comparisons of microscopy and size frequency data with interpretations of pigment HPLC data using the “CHEMTAX” matrix factorisation program. Mar. Ecol. Prog. Ser. 1996, 144, 285–298. [Google Scholar] [CrossRef]
- Wright, S.W.; Van den Enden, R.L. Phytoplankton community structure and stocks in the East Antarctic marginal ice zone (BROKE survey, January-March 1996) determined by CHEMTAX analysis of HPLC pigment signatures. Deep Res. Part II Top. Stud. Oceanogr. 2000, 47, 2363–2400. [Google Scholar] [CrossRef]
- DiTullio, G.R.; Geesey, M.E.; Jones, D.R.; Daly, K.L.; Campbell, L.; Smith, W.O. Phytoplankton assemblage structure and primary productivity along 170° W in the South Pacific Ocean. Mar. Ecol. Prog. Ser. 2003, 255, 55–80. [Google Scholar] [CrossRef] [Green Version]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [Green Version]
- Marsh, J.B.; Weinstein, D.B. Simple charring method for determination of lipids. J. Lipid Res. 1966, 7, 574–576. [Google Scholar] [CrossRef]
- Bhavya, P.S.; Kim, B.K.; Jo, N.; Kim, K.; Kang, J.J.; Lee, J.H.; Lee, D.; Lee, J.H.; Joo, H.T.; Ahn, S.H.; et al. A Review on the Macromolecular Compositions of Phytoplankton and the Implications for Aquatic Biogeochemistry. Ocean Sci. J. 2019, 54, 1–14. [Google Scholar] [CrossRef]
- Danovaro, R.; Dell’Anno, A.; Pusceddu, A.; Marrale, D.; Della Croce, N.; Fabiano, M.; Tselepides, A. Biochemical composition of pico-, nano- and micro-particulate organic matter and bacterioplankton biomass in the oligotrophic Cretan Sea (NE Mediterranean). Prog. Oceanogr. 2000, 46, 279–310. [Google Scholar] [CrossRef]
- Winberg, G.G. Symbols, Units and Conversion Factors in Study of Fresh Waters Productivity; International Biological Programme: London, UK, 1971. [Google Scholar]
- Engel, A.; Passow, U. Carbon and nitrogen content of transparent exopolymer particles (TEPS) in relation to their Alcian Blue adsorption. Mar. Ecol. Prog. Ser. 2001, 219, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Schloss, I.R.; Abele, D.; Ferreyra, G.A.; González, O.; Moreau, S.; Demers, S. Response of Potter Cove phytoplankton dynamics to long term climate trends. In Proceedings of the Open Science Conference XXXI SCAR, Buenos Aires, Argentina, 3–6 August 2010. [Google Scholar]
- Lee, S.H.; Joo, H.M.; Joo, H.T.; Kim, B.K.; Song, H.J.; Jeon, M.; Kang, S.H. Large contribution of small phytoplankton at Marian Cove, King George Island, Antarctica, based on long-term monitoring from 1996 to 2008. Polar Biol. 2015, 38, 207–220. [Google Scholar] [CrossRef]
- Schloss, I.R.; Ferreyra, G.A.; Ruiz-Pino, D. Phytoplankton biomass in Antarctic shelf zones: A conceptual model based on Potter Cove, King George Island. J. Mar. Syst. 2002, 36, 129–143. [Google Scholar] [CrossRef]
- Rivaro, P.; Luisa Abelmoschi, M.; Grotti, M.; Ianni, C.; Magi, E.; Margiotta, F.; Massolo, S.; Saggiomo, V. Combined effects of hydrographic structure and iron and copper availability on the phytoplankton growth in Terra Nova Bay Polynya (Ross Sea, Antarctica). Deep Res. Part I Oceanogr. Res. Pap. 2012, 62, 97–110. [Google Scholar] [CrossRef]
- Tremblay, J.E.; Smith, W.O. Chapter 8 Primary Production and Nutrient Dynamics in Polynyas. Elsevier Oceanogr. Ser. 2007, 74, 239–269. [Google Scholar] [CrossRef]
- Arrigo, K.R.; Weiss, A.M.; Smith, W.O. Physical forcing of phytoplankton dynamics in the southwestern Ross Sea. J. Geophys. Res. Ocean. 1998, 103, 1007–1021. [Google Scholar] [CrossRef]
- Peloquin, J.A.; Smith, W.O. Phytoplankton blooms in the Ross Sea, Antarctica: Interannual variability in magnitude, temporal patterns, and composition. J. Geophys. Res. Ocean. 2007, 112, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Trimborn, S.; Hoppe, C.J.M.; Taylor, B.B.; Bracher, A.; Hassler, C. Physiological characteristics of open ocean and coastal phytoplankton communities of Western Antarctic Peninsula and Drake Passage waters. Deep Res. Part I Oceanogr. Res. Pap. 2015, 98, 115–124. [Google Scholar] [CrossRef]
- Kim, H.; Ducklow, H.W.; Abele, D.; Barlett, E.M.R.; Buma, A.G.J.; Meredith, M.P.; Rozema, P.D.; Schofield, O.M.; Venables, H.J.; Schloss, I.R. Inter-decadal variability of phytoplankton biomass along the coastalWest Antarctic Peninsula. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2018, 376, 20170174. [Google Scholar] [CrossRef]
- Fabiano, M.; Povero, P.; Danovaro, R. Distribution and composition of particulate organic matter in the Ross Sea (Antarctica). Polar Biol. 1993, 13, 525–533. [Google Scholar] [CrossRef]
- Mangoni, O.; Saggiomo, V.; Bolinesi, F.; Margiotta, F.; Budillon, G.; Cotroneo, Y.; Misic, C.; Rivaro, P.; Saggiomo, M. Phytoplankton blooms during austral summer in the Ross Sea, Antarctica: Driving factors and trophic implications. PLoS ONE 2017, 12, e0176033. [Google Scholar] [CrossRef] [Green Version]
- Pusceddu, A.; Fabiano, M. Changes in the biochemical composition of Tetraselmis suecia and Isochrysis galbana during growth and decay. Chem. Ecol. 1996, 12, 199–212. [Google Scholar] [CrossRef]
- Smith, W.O.; Asper, V.L. The influence of phytoplankton assemblage composition on biogeochemical characteristics and cycles in the southern Ross Sea, Antarctica. Deep Res. Part I Oceanogr. Res. Pap. 2001, 48, 137–161. [Google Scholar] [CrossRef]
- Mangoni, O.; Modigh, M.; Conversano, F.; Carrada, G.C.; Saggiomo, V. Effects of summer ice coverage on phytoplankton assemblages in the Ross Sea, Antarctica. Deep Res. Part I Oceanogr. Res. Pap. 2004, 51, 1601–1617. [Google Scholar] [CrossRef]
- Andreoli, C.; Tolomio, C.; Moro, I.; Radice, M.; Moschin, E.; Bellato, S. Diatoms and dinoflagellates in Terra Nova Bay (Ross Sea-Antarctica) during austral summer 1990. Polar Biol. 1995, 15, 465–475. [Google Scholar] [CrossRef]
- Arrigo, K.R.; Robinson, D.H.; Worthen, D.L.; Dunbar, R.B.; DiTullio, G.R.; VanWoert, M.; Lizotte, M.P. Phytoplankton community structure and the drawdown of nutrients and CO2 in the Southern Ocean. Science 1999, 283, 365–367. [Google Scholar] [CrossRef] [Green Version]
- Saggiomo, V.; Catalano, G.; Mangoni, O.; Budillon, G.; Carrada, G.C. Primary production processes in ice-free waters of the Ross Sea (Antarctica) during the austral summer 1996. Deep Res. Part II Top. Stud. Oceanogr. 2002, 49, 1787–1801. [Google Scholar] [CrossRef]
- Kilham, S.S.; Kreeger, D.A.; Goulden, C.E.; Lynn, S.G. Effects of nutrient limitation on biochemical constituents of Ankistrodesmus falcatus. Freshw. Biol. 1997, 38, 591–596. [Google Scholar] [CrossRef]
- Van Leeuwe, M.A.; Scharek, R.; De Baar, H.J.W.; De Jong, J.T.M.; Goeyens, L. Iron enrichment experiments in the Southern Ocean: Physiological responses of plankton communities. Deep Res. Part II Top. Stud. Oceanogr. 1997, 44, 189–207. [Google Scholar] [CrossRef]
- Morris, I.; Glover, H.E.; Yentsch, C.S. Products of photosynthesis by marine phytoplankton: The effect of environmental factors on the relative rates of protein synthesis. Mar. Biol. 1974, 27, 1–9. [Google Scholar] [CrossRef]
- Liebezeit, G. Particulate carbohydrates in relation to phytoplankton in the euphotic zone of the Bransfield Strait. Polar Biol. 1984, 24, 225–228. [Google Scholar] [CrossRef]
- Moal, J.; Jezequel, V.M.; Harris, R.P.; Samain, J.F.; Poulet, S.A. Interspecific and intraspecific variability of the chemical composition of marine phytoplankton. Oceanol. Acta 1987, 10, 339–346. [Google Scholar]
- Fabregas, J.; Herrero, C.; Cabezas, B.; Abalde, J. Mass culture and biochemical variability of the marine microalga Tetraselmis suecica Kylin (Butch) with high nutrient concentrations. Aquaculture 1985, 49, 231–244. [Google Scholar] [CrossRef] [Green Version]
- Ahn, S.H.; Whitledge, T.E.; Stockwell, D.A.; Lee, J.H.; Lee, H.w.; Lee, S.H. The biochemical composition of phytoplankton in the Laptev and East Siberian seas during the summer of 2013. Polar Biol. 2019, 42, 133–148. [Google Scholar] [CrossRef]
- Fitzwater, S.E.; Johnson, K.S.; Gordon, R.M.; Coale, K.H.; Smith, W.O. Trace metal concentrations in the Ross Sea and their relationship with nutrients and phytoplankton growth. Deep Res. Part II Top. Stud. Oceanogr. 2000, 47, 3159–3179. [Google Scholar] [CrossRef]
- Smith, W.O.; Dinniman, M.S.; Klinck, J.M.; Hofmann, E. Biogeochemical climatologies in the Ross Sea, Antarctica: Seasonal patterns of nutrients and biomass. Deep Res. Part II Top. Stud. Oceanogr. 2003, 50, 3083–3101. [Google Scholar] [CrossRef]
- Arrigo, K.R.; van Dijken, G.L. Phytoplankton dynamics within 37 Antarctic coastal polynya systems. J. Geophys. Res. Ocean. 2003, 108. [Google Scholar] [CrossRef]
- Ryan-Keogh, T.J.; DeLizo, L.M.; Smith, W.O.; Sedwick, P.N.; McGillicuddy, D.J.; Moore, C.M.; Bibby, T.S. Temporal progression of photosynthetic-strategy in phytoplankton in the Ross Sea, Antarctica. J. Mar. Syst. 2017, 166, 87–96. [Google Scholar] [CrossRef] [Green Version]
- McKnight, D.M.; Howes, B.L.; Taylor, C.D.; Goehringer, D.D. Phytoplankton dynamics in a stably stratified antarctic lake during winter darkness. J. Phycol. 2000, 36, 852–861. [Google Scholar] [CrossRef]
- Brown, P.C.; Painting, S.J.; Cochrane, K.L. Estimates of phytoplankton and bacterial biomass and production in the northern and southern benguela ecosystems. S. Afr. J. Mar. Sci. 1991, 11, 537–564. [Google Scholar] [CrossRef]
- Lee, J.H.; Lee, D.; Kang, J.J.; Joo, H.T.; Lee, J.H.; Lee, H.W.; Ahn, S.H.; Kang, C.K.; Lee, S.H. The effects of different environmental factors on the biochemical composition of particulate organic matter in Gwangyang Bay, South Korea. Biogeosciences 2017, 14, 1903–1917. [Google Scholar] [CrossRef] [Green Version]
- Alderkamp, A.C.; Buma, A.G.J.; Van Rijssel, M. The carbohydrates of Phaeocystis and their degradation in the microbial food web. Biogeochemistry 2007, 83, 99–118. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.K.; Lee, S.H.; Ha, S.Y.; Jung, J.; Kim, T.W.; Yang, E.J.; Jo, N.; Lim, Y.J.; Park, J.; Lee, S.H. Vertical Distributions of Macromolecular Composition of Particulate Organic Matter in the Water Column of the Amundsen Sea Polynya During the Summer in 2014. J. Geophys. Res. Ocean. 2018, 123, 1393–1405. [Google Scholar] [CrossRef]
- DiTullio, G.R.; Smith, W.O. Spatial patterns in phytoplankton biomass and pigment distributions in the Ross Sea. J. Geophys. Res. C Ocean. 1996, 101, 18467–18477. [Google Scholar] [CrossRef]
- Fabiano, M.; Danovaro, R.; Chiantore, M.; Pusceddu, A. Bacteria, Protozoa and Organic Matter Composition in the Sediments of Terra Nova Bay (Ross Sea); Ross Sea ecology; Faranda, F., Guglielmo, L., Ianora, A., Eds.; Springer: Berlin/Heidelberg, Germany; Springer: New York, NY, USA, 2000; pp. 159–169. [Google Scholar]
- Kim, B.K.; Lee, J.H.; Yun, M.S.; Joo, H.T.; Song, H.J.; Yang, E.J.; Chung, K.H.; Kang, S.H.; Lee, S.H. High lipid composition of particulate organic matter in the northern Chukchi Sea, 2011. Deep Res. Part II Top. Stud. Oceanogr. 2015, 120, 72–81. [Google Scholar] [CrossRef]
- Jo, N.; La, H.S.; Kim, J.H.; Kim, K.; Kim, B.K.; Kim, M.J.; Son, W.; Lee, S.H. Different Biochemical Compositions of Particulate Organic Matter Driven by Major Phytoplankton Communities in the Northwestern Ross Sea. Front. Microbiol. 2021, 12, 5. [Google Scholar] [CrossRef]
- Young, J.N.; Goldman, J.A.L.; Kranz, S.A.; Tortell, P.D.; Morel, F.M.M. Slow carboxylation of Rubisco constrains the rate of carbon fixation during Antarctic phytoplankton blooms. New Phytol. 2015, 205, 172–181. [Google Scholar] [CrossRef]
- Hitchcock, G.L. A comparative study of the size-dependent organic composition of marine diatoms and dinoflagellates. J. Plankton Res. 1982, 4, 363–377. [Google Scholar] [CrossRef]
- Berdalet, E.; Latasa, M.; Estrada, M. Effects of nitrogen and phosphorus starvation on nucleic acid and protein content of Heterocapsa sp. J. Plankton Res. 1994, 16, 303–316. [Google Scholar] [CrossRef] [Green Version]
- Myklestad, S.; Haug, A. Production of carbohydrates by the marine diatom Chaetoceros affinis var. willei (Gran) Hustedt. I. Effect of the concentration of nutrients in the culture medium. J. Exp. Mar. Bio. Ecol. 1972, 9, 125–136. [Google Scholar] [CrossRef]
- Lindqvist, K.; Lignell, R. Intracellular partitioning of 14CO2 in phytoplankton during a growth season in the northern Baltic. Mar. Ecol. Prog. Ser. 1997, 152, 41–50. [Google Scholar] [CrossRef]
- Handa, N. Carbohydrate metabolism in the marine diatom Skeletonema costatum. Mar. Biol. 1969, 4, 208–214. [Google Scholar] [CrossRef]
- Parrish, C.C. Time series of particulate and dissolved lipid classes during spring phytoplankton blooms in Bedford Basin, a marine inlet. Mar. Ecol. Prog. Ser. 1987, 35, 129–139. [Google Scholar] [CrossRef]
- Taipale, S.J.; Galloway, A.W.E.; Aalto, S.L.; Kahilainen, K.K.; Strandberg, U.; Kankaala, P. Terrestrial carbohydrates support freshwater zooplankton during phytoplankton deficiency. Sci. Rep. 2016, 6, 30897. [Google Scholar] [CrossRef] [Green Version]
- Corzo, A.; Rodríguez-Gálvez, S.; Lubian, L.; Sangrá, P.; Martínez, A.; Morillo, J.A. Spatial distribution of transparent exopolymer particles in the Bransfield Strait, Antarctica. J. Plankton Res. 2005, 27, 635–646. [Google Scholar] [CrossRef] [Green Version]
- Ramaiah, N.; Yoshikawa, T.; Furuya, K. Temporal variations in transparent exopolymer particles (TEPS) associated with a diatom spring bloom in a subarctic ria in Japan. Mar. Ecol. Prog. Ser. 2001, 212, 79–88. [Google Scholar] [CrossRef]
- Ortega-Retuerta, E.; Duarte, C.M.; Reche, I. Significance of bacterial activity for the distribution and dynamics of transparent exopolymer particles in the Mediterranean Sea. Microb. Ecol. 2010, 59, 808–818. [Google Scholar] [CrossRef] [Green Version]
- Malpezzi, M.A.; Sanford, L.P.; Crump, B.C. Abundance and distribution of transparent exopolymer particles in the estuarine turbidity maximum of Chesapeake Bay. Mar. Ecol. Prog. Ser. 2013, 486, 23–35. [Google Scholar] [CrossRef]
- Yamada, Y.; Yokokawa, T.; Uchimiya, M.; Nishino, S.; Fukuda, H.; Ogawa, H.; Nagata, T. Transparent exopolymer particles (TEPS) in the deep ocean: Full-depth distribution patterns and contribution to the organic carbon pool. Mar. Ecol. Prog. Ser. 2017, 583, 81–93. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Lee, W.C.; Kim, H.C.; Jo, N.; Jang, H.K.; Kang, J.J.; Lee, D.; Kim, K.; Lee, S.H. Transparent exopolymer particle (TEPs) dynamics and contribution to particulate organic carbon (POC) in Jaran bay, Korea. Water 2020, 12, 1057. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1. Salinity | 1 | −0.93 ** | 0.88 ** | −0.58 ** | −0.61 ** | −0.52 * | −0.72 ** | −0.60 ** | −0.68 ** | - | −0.82 ** | −0.50 * | - | −0.81 ** | −0.77 ** |
| 2. Temperature | 1 | −0.88 ** | 0.61 ** | 0.63 ** | 0.55 * | 0.66 ** | 0.62 ** | 0.64 ** | - | 0.71 ** | - | - | 0.80 ** | 0.73 ** | |
| 3. NO3− + NO2− | 1 | −0.59 ** | −0.86 ** | −0.76 ** | −0.87 ** | −0.82 ** | −0.86 ** | −0.50 * | −0.68 ** | −0.71 ** | - | −0.94 ** | −0.92 ** | ||
| 4. NH4+ | 1 | - | - | - | - | - | 0.76 ** | 0.68 ** | - | - | - | - | |||
| 5. POC | 1 | 0.87 ** | 0.80 ** | 0.87 ** | 0.80 ** | - | 0.47 * | 0.76 ** | - | 0.90 ** | 0.92 ** | ||||
| 6. Chl-a | 1 | 0.83 ** | 0.97 ** | 0.77 ** | - | - | 0.80 ** | - | 0.88 ** | 0.83 ** | |||||
| 7. TEPs | 1 | 0.89 ** | 0.95 ** | - | 0.49 * | 0.90 ** | - | 0.89 ** | 0.87 ** | ||||||
| 8. Diatoms | 1 | 0.87 ** | - | - | 0.85 ** | - | 0.94 ** | 0.87 ** | |||||||
| 9. Dinoflagellates | 1 | - | 0.54 * | 0.90 ** | - | 0.88 ** | 0.86 ** | ||||||||
| 10. P. antarctica | 1 | 0.62 ** | - | - | - | - | |||||||||
| 11. Cryptophytes | 1 | - | - | 0.61 ** | 0.67 ** | ||||||||||
| 12. Other phytoplankton | 1 | - | 0.78 ** | 0.78 ** | |||||||||||
| 13. CHO | 1 | - | - | ||||||||||||
| 14. PRT | 1 | 0.97 ** | |||||||||||||
| 15. LIP | 1 |
| Region | Season | Depth (m) | TEP Range (Mean) | TEPs/Chl-a | TEPs–Chl-a Relationship | References |
|---|---|---|---|---|---|---|
| (µg Xeq. L−1) | (Mean) | (Log Converted) | ||||
| Anvers Island | summer | Surface | 10–407 (207) | 12–708 (123) | Not related | Passow et al. (1995) [35] |
| Ross Sea | summer | 0–150 | 0–2800 (308) | 89.1 | y = 3.63x + 1.01 | Hong et al. (1997) [36] |
| Bransfield Strait | summer | 0–100 | 0–346 (57) | 51 | y = 0.32x + 1.63 | Corzo et al. (2005) [97] |
| Gerlache Strait | summer | 0–100 | 0–283 | 32.7 | y = 0.67x + 1.52 | Corzo et al. (2005) [97] |
| Northern Weddell Sea | summer | Surface | 39.2–177.6 (102.3) | 79.3 | y = 0.35x + 1.90 | Zamanillo et al. (2019) [34] |
| Antarctic Peninsula | summer | 0–200 | 0–48.9 (15.4) | 0–1492 (40.9) | y = 0.38x + 1.08 | Ortega-Retuerta et al. (2009) [29] |
| Jang Bogo Station | summer | Surface | 45.15–96.1 (73.0) | 38.9–404.3 (165.9) | y = 0.33x + 1.87 | This Study |
| Jang Bogo Station | winter | Surface | 10–33.5 (17.9) | 371.1–2807.5 (1477.9) | Not related | This Study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, S.; Park, J.; Yoo, K.-C.; Yoo, J.; Kim, K.; Jo, N.; Jang, H.-K.; Kim, J.; Kim, J.; Kim, J.; et al. Seasonal Variations in the Biochemical Compositions of Phytoplankton and Transparent Exopolymer Particles (TEPs) at Jang Bogo Station (Terra Nova Bay, Ross Sea), 2017–2018. Water 2021, 13, 2173. https://doi.org/10.3390/w13162173
Park S, Park J, Yoo K-C, Yoo J, Kim K, Jo N, Jang H-K, Kim J, Kim J, Kim J, et al. Seasonal Variations in the Biochemical Compositions of Phytoplankton and Transparent Exopolymer Particles (TEPs) at Jang Bogo Station (Terra Nova Bay, Ross Sea), 2017–2018. Water. 2021; 13(16):2173. https://doi.org/10.3390/w13162173
Chicago/Turabian StylePark, Sanghoon, Jisoo Park, Kyu-Cheul Yoo, Jaeill Yoo, Kwanwoo Kim, Naeun Jo, Hyo-Keun Jang, Jaehong Kim, Jaesoon Kim, Joonmin Kim, and et al. 2021. "Seasonal Variations in the Biochemical Compositions of Phytoplankton and Transparent Exopolymer Particles (TEPs) at Jang Bogo Station (Terra Nova Bay, Ross Sea), 2017–2018" Water 13, no. 16: 2173. https://doi.org/10.3390/w13162173