
F-Specific RNA Bacteriophage Transport in Stream Water: Hydro-Meteorological Controls and Association with Suspended Solids


 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
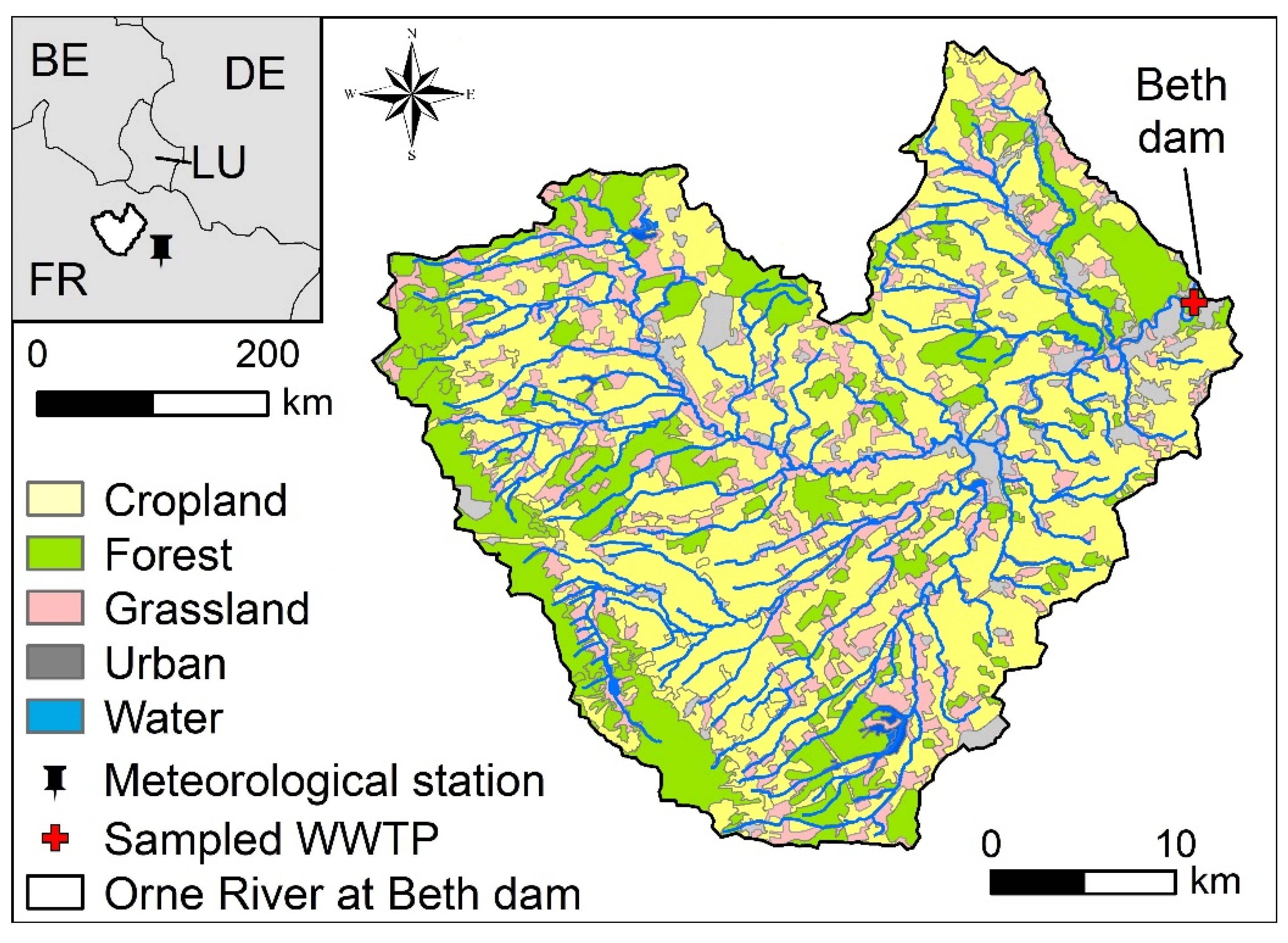
2.1. Study Area
2.2. Experimental Set-Up and Sampling
2.3. Laboratory Analysis
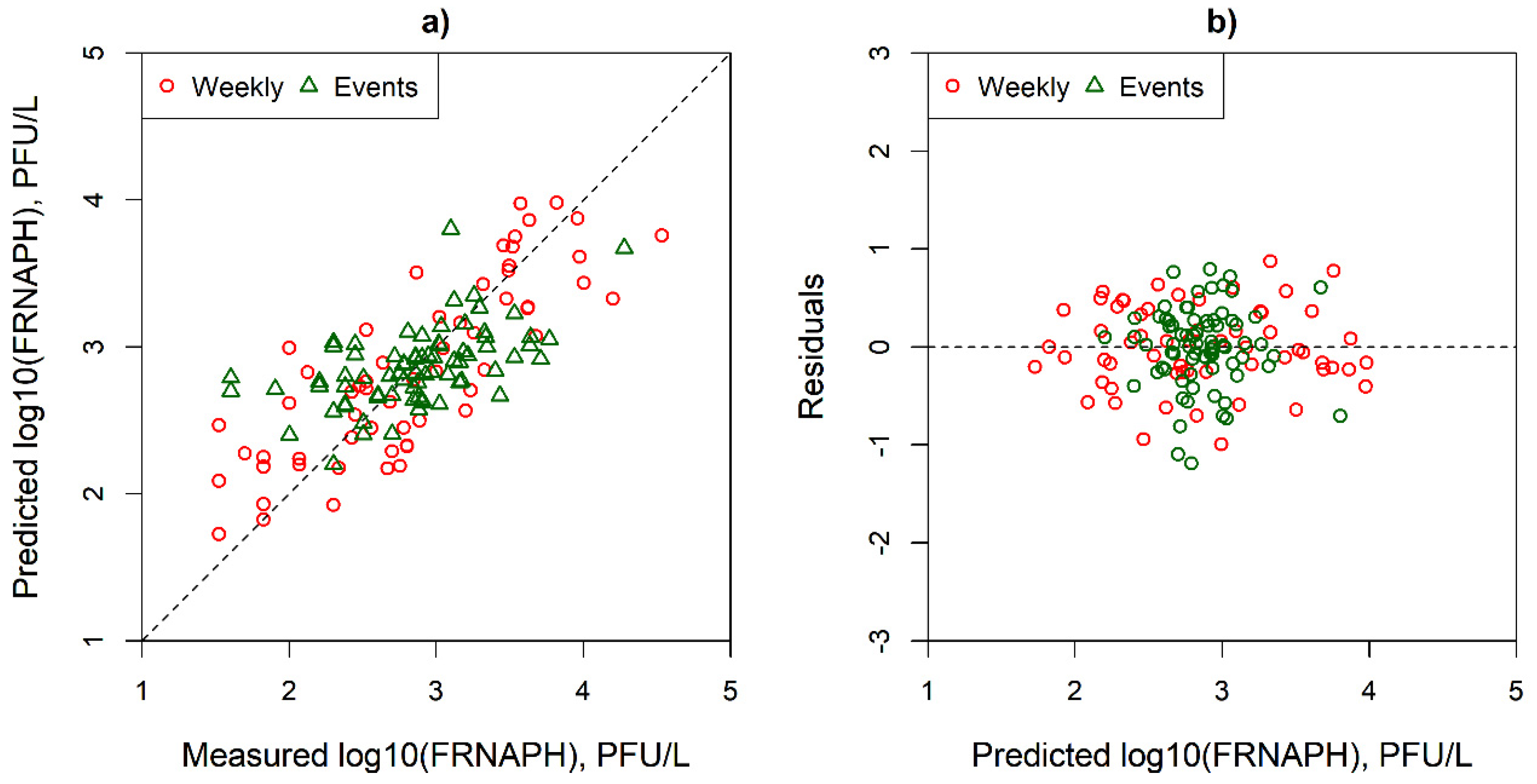
2.4. Statistical Analysis
3. Results
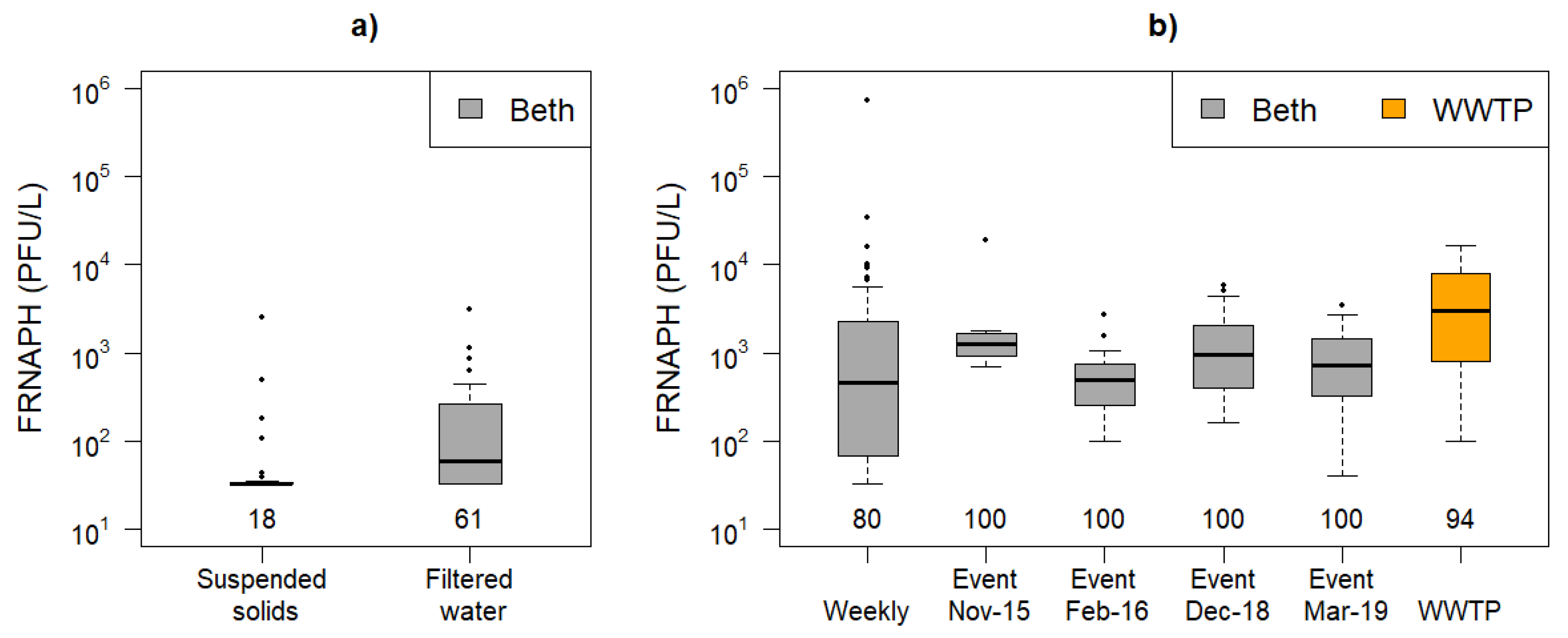
3.1. F-Specific RNA Bacteriophage Concentrations
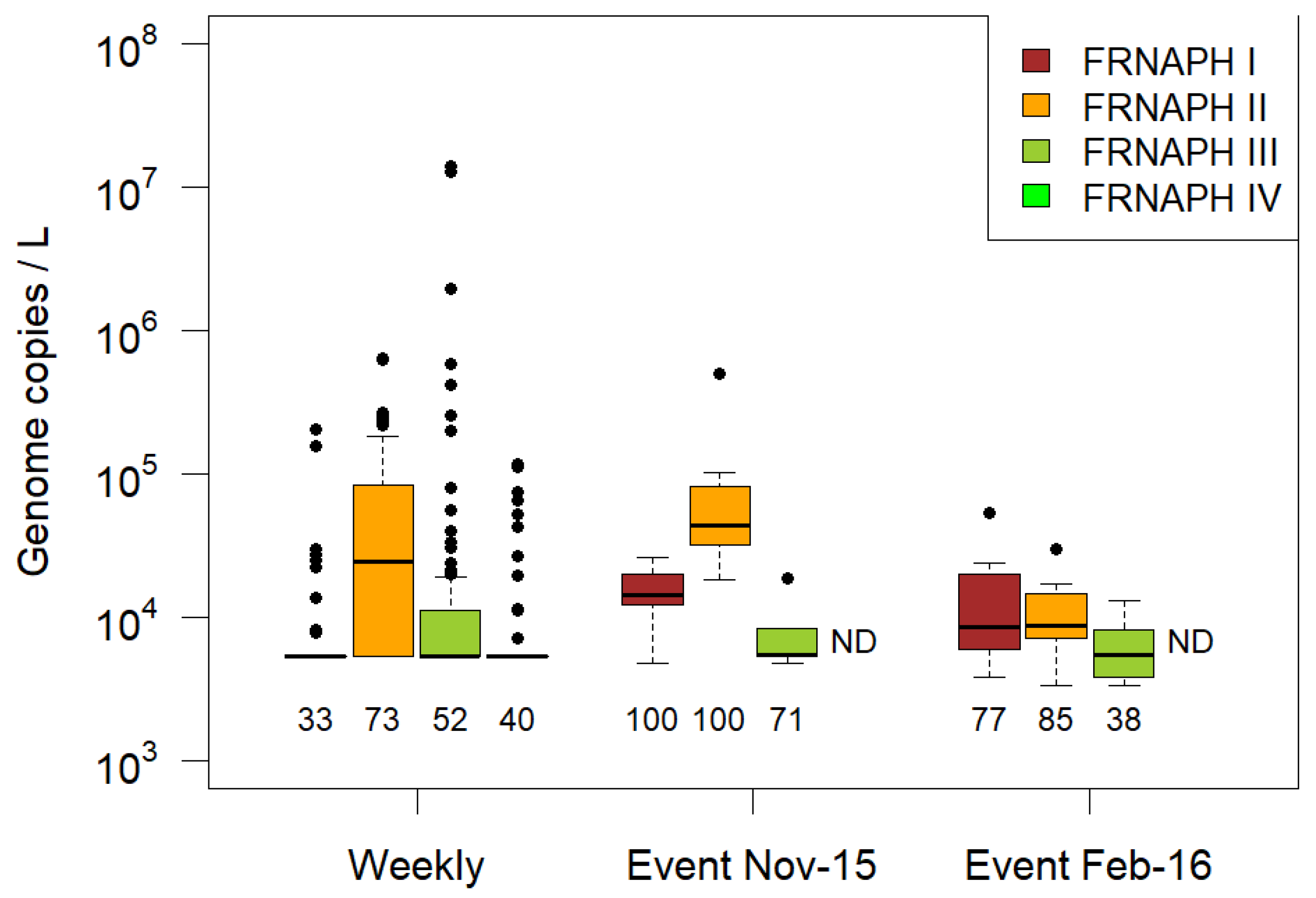
3.2. FRNAPH Genogroups
3.3. Simple and Multi Parameter Regression
4. Discussion
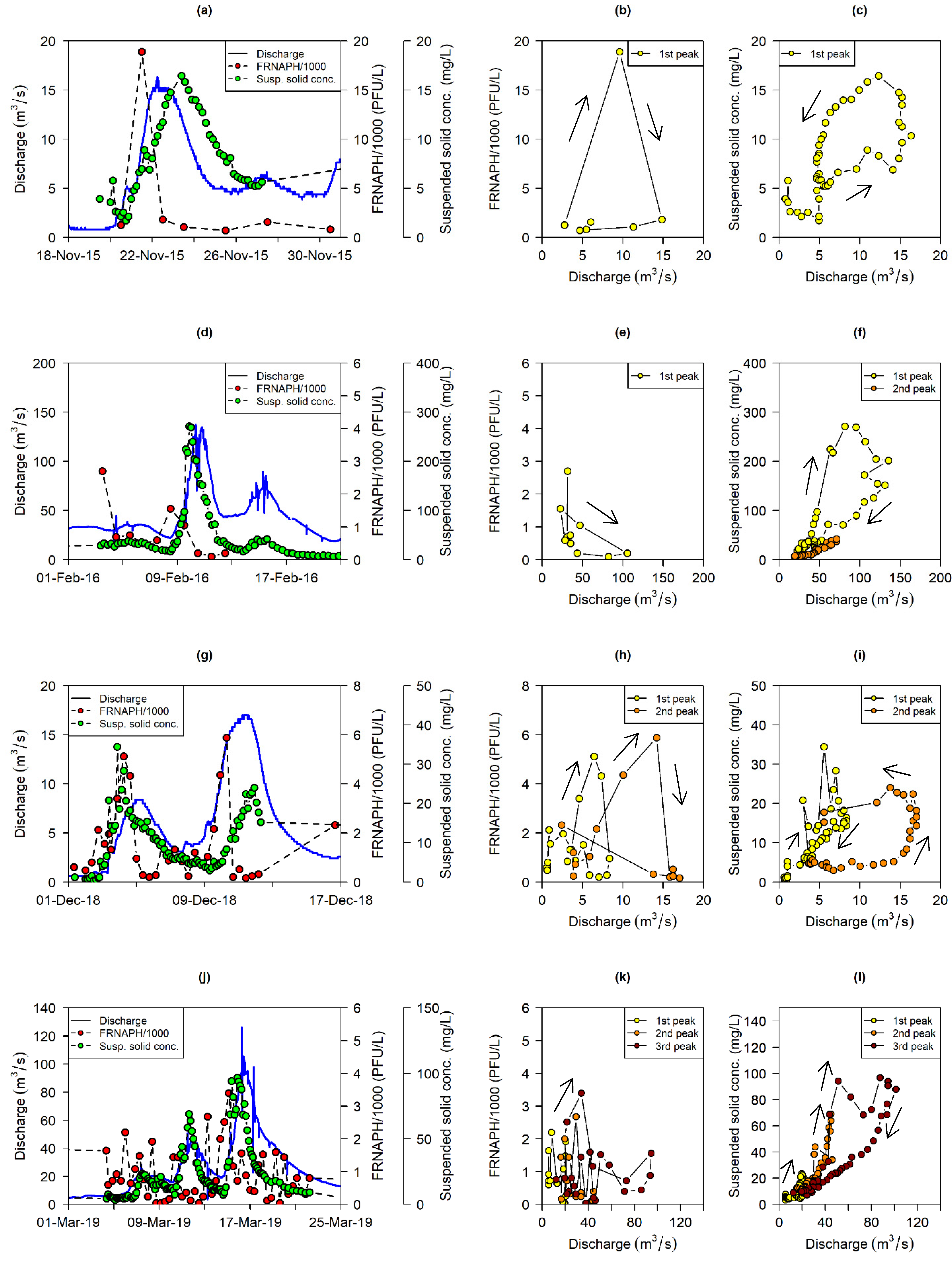
4.1. Interplay between FRNAPHs and Suspended Solids in the Water Column
4.2. FRNAPH Controlling Factors at Low Flows and during Storm Runoff Events
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sano, D.; Amarasiri, M.; Hata, A.; Watanabe, T.; Katayama, H. Risk management of viral infectious diseases in wastewater reclamation and reuse: Review. Environ. Int. 2016, 91, 220–229. [Google Scholar] [CrossRef] [Green Version]
- Fong, T.-T.; Lipp, E.K. Enteric Viruses of Humans and Animals in Aquatic Environments: Health Risks, Detection, and Potential Water Quality Assessment Tools. Microbiol. Mol. Biol. Rev. 2005, 69, 357–371. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, R.A.; Gundy, P.M.; Rijal, G.K.; Gerba, C.P. The Impact of Combined Sewage Overflows on the Viral Contamination of Receiving Waters. Food Environ. Virol. 2012, 4, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Fauvel, B.; Cauchie, H.M.; Gantzer, C.; Ogorzaly, L. Contribution of hydrological data to the understanding of the spatio-temporal dynamics of F-specific RNA bacteriophages in river water during rainfall-runoff events. Water Res. 2016, 94, 328–340. [Google Scholar] [CrossRef] [PubMed]
- Ogorzaly, L.; Tissier, A.; Bertrand, I.; Maul, A.; Gantzer, C. Relationship between F-specific RNA phage genogroups, faecal pollution indicators and human adenoviruses in river water. Water Res. 2009, 43, 1257–1264. [Google Scholar] [CrossRef] [PubMed]
- Hassard, F.; Gwyther, C.L.; Farkas, K.; Andrews, A.; Jones, V.; Cox, B.; Brett, H.; Jones, D.L.; McDonald, J.E.; Malham, S.K. Abundance and Distribution of Enteric Bacteria and Viruses in Coastal and Estuarine Sediments—A Review. Front. Microbiol. 2016, 7, 1692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savichtcheva, O.; Okabe, S. Alternative indicators of fecal pollution: Relations with pathogens and conventional indicators, current methodologies for direct pathogen monitoring and future application perspectives. Water Res. 2006, 40, 2463–2476. [Google Scholar] [CrossRef] [PubMed]
- Jofre, J.; Lucena, F.; Blanch, A.R. Coliphages as a Complementary Tool to Improve the Management of Urban Wastewater Treatments and Minimize Health Risks in Receiving Waters. Water 2021, 13, 1110. [Google Scholar] [CrossRef]
- Directive 2020/2184. Directive (EU) 2020/2184 of the European Parliament and of the Council of 16 December 2020 on the quality of intended water for human consumption. Off. J. Eur. Communities 2020, 435, 1–62. [Google Scholar]
- 2020/741 Regulation (EU). European Parliament and of the Council of 25 May 2020 on minimum requirements for water reuse. Off. J. Eur. Union 2020, 177, 32–55. [Google Scholar]
- Field, K.G.; Samadpour, M. Fecal source tracking, the indicator paradigm, and managing water quality. Water Res. 2007, 41, 3517–3538. [Google Scholar] [CrossRef] [PubMed]
- Mauffret, A.; Caprais, M.-P.; Gourmelon, M. Relevance of Bacteroidales and F-Specific RNA Bacteriophages for Efficient Fecal Contamination Tracking at the Level of a Catchment in France. Appl. Environ. Microbiol. 2012, 78, 5143–5152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMinn, B.R.; Ashbolt, N.J.; Korajkic, A. Bacteriophages as indicators of faecal pollution and enteric virus removal. Lett. Appl. Microbiol. 2017, 65, 11–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muniesa, M.; Jofre, J. Factors influencing the replication of somatic coliphages in the water environment. Antonie Van Leeuwenhoek 2004, 86, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Jofre, J. Is the replication of somatic coliphages in water environments significant? J. Appl. Microbiol. 2009, 106, 1059–1069. [Google Scholar] [CrossRef]
- Bollback, J.P.; Huelsenbeck, J.P. Phylogeny, Genome Evolution, and Host Specificity of Single-Stranded RNA Bacteriophage (Family Leviviridae). J. Mol. Evol. 2001, 52, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Cole, D.; Long, S.C.; Sobsey, M.D. Evaluation of F+ RNA and DNA Coliphages as Source-Specific Indicators of Fecal Contamination in Surface Waters. Appl. Environ. Microbiol. 2003, 69, 6507–6514. [Google Scholar] [CrossRef] [Green Version]
- Hartard, C.; Banas, S.; Loutreul, J.; Rincé, A.; Benoit, F.; Boudaud, N.; Gantzer, C. Relevance of F-Specific RNA Bacteriophages in Assessing Human Norovirus Risk in Shellfish and Environmental Waters. Appl. Environ. Microbiol. 2016, 82, 5709–5719. [Google Scholar] [CrossRef] [Green Version]
- Bertrand, I.; Schijven, J.F.; Sánchez, G.; Wyn-Jones, P.; Ottoson, J.; Morin, T.; Muscillo, M.; Verani, M.; Nasser, A.; de Roda Husman, A.M.; et al. The impact of temperature on the inactivation of enteric viruses in food and water: A review. J. Appl. Microbiol. 2012, 112, 1059–1074. [Google Scholar] [CrossRef]
- Boehm, A.B.; Silverman, A.I.; Schriewer, A.; Goodwin, K. Systematic review and meta-analysis of decay rates of waterborne mammalian viruses and coliphages in surface waters. Water Res. 2019, 164, 114898. [Google Scholar] [CrossRef]
- Rajala, R.L.; Heinonen-Tanski, H. Survival and transfer of faecal indicator organisms of wastewater effluents in receiving lake waters. Water Sci. Technol. 1998, 38, 191–194. [Google Scholar] [CrossRef]
- Westrell, T.; Teunis, P.; van den Berg, H.; Lodder, W.; Ketelaars, H.; Stenström, T.A.; de Roda Husman, A.M. Short- and long-term variations of norovirus concentrations in the Meuse river during a 2-year study period. Water Res. 2006, 40, 2613–2620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fauvel, B.; Gantzer, C.; Cauchie, H.M.; Ogorzaly, L. In Situ Dynamics of F-Specific RNA Bacteriophages in a Small River: New Way to Assess Viral Propagation in Water Quality Studies. Food Environ. Virol. 2017, 9, 89–102. [Google Scholar] [CrossRef] [Green Version]
- Skraber, S.; Gassilloud, B.; Gantzer, C. Comparison of Coliforms and Coliphages as Tools for Assessment of Viral Contamination in River Water. Appl. Environ. Microbiol. 2004, 70, 3644–3649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skraber, S.; Schijven, J.; Italiaander, R.; Husman, A.M.d.R. Accumulation of enteric bacteriophage in fresh water sediments. J. Water Health 2009, 7, 372–379. [Google Scholar] [CrossRef] [Green Version]
- Fauvel, B.; Cauchie, H.-M.; Gantzer, C.; Ogorzaly, L. Influence of physico-chemical characteristics of sediment on the in situ spatial distribution of F-specific RNA phages in the riverbed. FEMS Microbiol. Ecol. 2019, 95, fiy240. [Google Scholar] [CrossRef]
- Mackowiak, M.; Leifels, M.; Hamza, I.A.; Jurzik, L.; Wingender, J. Distribution of Escherichia coli, coliphages and enteric viruses in water, epilithic biofilms and sediments of an urban river in Germany. Sci. Total Environ. 2018, 626, 650–659. [Google Scholar] [CrossRef]
- Templeton, M.R.; Andrews, R.C.; Hofmann, R. Inactivation of particle-associated viral surrogates by ultraviolet light. Water Res. 2005, 39, 3487–3500. [Google Scholar] [CrossRef]
- Bradford, S.A.; Tadassa, Y.F.; Jin, Y. Transport of Coliphage in the Presence and Absence of Manure Suspension. J. Environ. Qual. 2006, 35, 1692–1701. [Google Scholar] [CrossRef]
- Martín-Díaz, J.; Lucena, F.; Blanch, A.R.; Jofre, J. Review: Indicator bacteriophages in sludge, biosolids, sediments and soils. Environ. Res. 2020, 182, 109133. [Google Scholar] [CrossRef]
- Kay, D.; Edwards, A.C.; Ferrier, R.C.; Francis, C.; Kay, C.; Rushby, L.; Watkins, J.; McDonald, A.T.; Wyer, M.; Crowther, J.; et al. Catchment microbial dynamics: The emergence of a research agenda. Prog. Phys. Geogr. Earth Environ. 2007, 31, 59–76. [Google Scholar] [CrossRef]
- Cho, K.H.; Pachepsky, Y.A.; Oliver, D.M.; Muirhead, R.W.; Park, Y.; Quilliam, R.S.; Shelton, D.R. Modeling fate and transport of fecally-derived microorganisms at the watershed scale: State of the science and future opportunities. Water Res. 2016, 100, 38–56. [Google Scholar] [CrossRef]
- Kim, M.; Boithias, L.; Cho, K.H.; Silvera, N.; Thammahacksa, C.; Latsachack, K.; Rochelle-Newall, E.; Sengtaheuanghoung, O.; Pierret, A.; Pachepsky, Y.A.; et al. Hydrological modeling of Fecal Indicator Bacteria in a tropical mountain catchment. Water Res. 2017, 119, 102–113. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.M.M.; Sokolova, E.; Hofstra, N. Modelling of river faecal indicator bacteria dynamics as a basis for faecal contamination reduction. J. Hydrol. 2018, 563, 1000–1008. [Google Scholar] [CrossRef]
- Dorner, S.M.; Anderson, W.B.; Slawson, R.M.; Kouwen, N.; Huck, P.M. Hydrologic Modeling of Pathogen Fate and Transport. Environ. Sci. Technol. 2006, 40, 4746–4753. [Google Scholar] [CrossRef] [PubMed]
- Fauvel, B.; Ogorzaly, L.; Cauchie, H.M.; Gantzer, C. Interactions of infectious F-specific RNA bacteriophages with suspended matter and sediment: Towards an understanding of FRNAPH distribution in a river water system. Sci. Total Environ. 2017, 574, 960–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demeter, K.; Derx, J.; Komma, J.; Parajka, J.; Schijven, J.; Sommer, R.; Cervero-Aragó, S.; Linder, G.; Zoufal-Hruza, C.M.; Linke, R.; et al. Modelling the interplay of future changes and wastewater management measures on the microbiological river water quality considering safe drinking water production. Sci. Total Environ. 2021, 768, 144278. [Google Scholar] [CrossRef] [PubMed]
- de Brauwere, A.; Ouattara, N.K.; Servais, P. Modeling Fecal Indicator Bacteria Concentrations in Natural Surface Waters: A Review. Crit. Rev. Environ. Sci. Technol. 2014, 44, 380–2453. [Google Scholar] [CrossRef] [Green Version]
- Kanbar, H.J.; Montargès-Pelletier, E.; Losson, B.; Bihannic, I.; Gley, R.; Bauer, A.; Villieras, F.; Manceau, L.; El Samrani, A.G.; Kazpard, V.; et al. Iron mineralogy as a fingerprint of former steelmaking activities in river sediments. Sci. Total. Environ. 2017, 599–600, 540–553. [Google Scholar] [CrossRef]
- Garcier, R.J. Rivers we can’t bring ourselves to clean–historical insights into the pollution of the Moselle River (France), 1850–2000. Hydrol. Earth Syst. Sci. 2007, 11, 1731–1745. [Google Scholar] [CrossRef] [Green Version]
- Losson, B.; Manceau, L.; Kanbar, H.J.; Waldvogel, Y.; Delus, C.; Mansuy-Huault, L.; Hissler, C.; Montargès-Pelletier, E. Orne River hydrodynamic and effect on sediment transport in the backwater zone of the Beth Dam (Lorraine, France). Géomorphol. Relief Process. Environ. 2020, 1, 3–17. [Google Scholar] [CrossRef]
- Lepesqueur, J.; Hostache, R.; Martínez-Carreras, N.; Montargès-Pelletier, E.; Hissler, C. Sediment transport modelling in riverine environments: On the importance of grain-size distribution, sediment density, and suspended sediment concentrations at the upstream boundary. Hydrol. Earth Syst. Sci. 2019, 23, 3901–3915. [Google Scholar] [CrossRef] [Green Version]
- The Water Information System Rhine-Meuse. Available online: https://rhin-meuse.eaufrance.fr (accessed on 30 June 2021).
- Skraber, S.; Langlet, J.; Kremer, J.R.; Mossong, J.; De Landtsheer, S.; Even, J.; Muller, C.P.; Hoffmann, L.; Cauchie, H.-M. Concentration and Diversity of Noroviruses Detected in Luxembourg Wastewaters in 2008–2009. Appl. Environ. Microbiol. 2011, 77, 5566–5568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogorzaly, L.; Gantzer, C. Development of real-time RT-PCR methods for specific detection of F-specific RNA bacteriophage genogroups: Application to urban raw wastewater. J. Virol. Methods 2006, 138, 131–139. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Williams, G.P. Sediment concentration versus water discharge during single hydrologic events in rivers. J. Hydrol. 1989, 111, 89–106. [Google Scholar] [CrossRef]
- Reimann, C.; Filzmoser, P.; Garrett, R.G.; Dutter, R. Statistical Data Analysis Explained: Applied Environmental Statistics with R; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2008. [Google Scholar]
- Kuhn, M. Building Predictive Models in R Using the caret Package. J. Stat. Softw. 2008, 28, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Knappett, P.S.K.; Emelko, M.B.; Zhuang, J.; McKay, L.D. Transport and retention of a bacteriophage and microspheres in saturated, angular porous media: Effects of ionic strength and grain size. Water Res. 2008, 42, 4368–4378. [Google Scholar] [CrossRef] [PubMed]
- Treumann, S.; Torkzaban, S.; Bradford, S.A.; Visalakshan, R.M.; Page, D. An explanation for differences in the process of colloid adsorption in batch and column studies. J. Contam. Hydrol. 2014, 164, 219–229. [Google Scholar] [CrossRef]
- Chu, Y.; Jin, Y.; Yates, M.V. Virus Transport through Saturated Sand Columns as Affected by Different Buffer Solutions. J. Environ. Qual. 2000, 29, 1103–1110. [Google Scholar] [CrossRef]
- Schijven, J.F.; Hassanizadeh, S.M. Removal of Viruses by Soil Passage: Overview of Modeling, Processes, and Parameters. Crit. Rev. Environ. Sci. Technol. 2000, 30, 49–127. [Google Scholar] [CrossRef]
- Zhuang, J.; Jin, Y. Interactions between viruses and goethite during saturated flow: Effects of solution pH, carbonate, and phosphate. J. Contam. Hydrol. 2008, 98, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Kanbar, H.J. What the Orne River Tells about the Former Steelmaking Activities: Chemical and Mineralogical Investigations on Sediment. Ph.D. Thesis, Université de Lorraine, Vandoeuvre-les-Nancy, France, en Cotutelle avec l’Université Libanaise, Beirut, Liban, 2017. [Google Scholar]
- Dika, C.; Ly-Chatain, M.H.; Francius, G.; Duval, J.F.L.; Gantzer, C. Non-DLVO adhesion of F-specific RNA bacteriophages to abiotic surfaces: Importance of surface roughness, hydrophobic and electrostatic interactions. Colloids Surf. A Physicochem. Eng. Asp. 2013, 435, 178–187. [Google Scholar] [CrossRef]
- Langlet, J.; Gaboriaud, F.; Duval, J.F.L.; Gantzer, C. Aggregation and surface properties of F-specific RNA phages: Implication for membrane filtration processes. Water Res. 2008, 42, 2769–2777. [Google Scholar] [CrossRef] [PubMed]
- Maranger, R.; Bird, D.F. High concentrations of viruses in the sediments of Lac Gilbert, Québec. Microb. Ecol. 1996, 31, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Droppo, I.G. Rethinking what constitutes suspended sediment. Hydrol. Process. 2001, 15, 1551–1564. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Variable Description | Units |
|---|---|---|
| Hydrological variables | Log10 transformed turbidity | NTU |
| Log10 transformed suspended solid concentration | mg/L | |
| Log10 transformed discharge | m3/s | |
| Conductivity | µS/cm | |
| Water temperature | °C | |
| Meteorological variables | Antecedent precipitation: 5, 10-days | mm |
| Air temperature | °C | |
| Solar radiation | W/m2 |
| Weekly/Events | a. | b. | c. | d. | e. | f. | g. | h. | i. | j. |
|---|---|---|---|---|---|---|---|---|---|---|
| a. FRNAPH | 1 | |||||||||
| b. Conductivity | −0.52/0.37 | 1 | ||||||||
| c. Water temp. | −0.72/0.16 | 0.45/0.26 | 1 | |||||||
| d. Turbidity | 0.39/−0.39 | −0.61/−0.78 | −0.28/−0.09 | 1 | ||||||
| e. Susp. solid conc. | 0.26/−0.24 | −0.43/−0.77 | −0.05/−0.25 | 0.74/0.73 | 1 | |||||
| f. Air temperature | −0.64/0.13 | 0.37/0.21 | 0.90/0.21 | −0.17/−0.14 | 0.07/−0.08 | 1 | ||||
| g. Solar radiation | −0.45/−0.17 | 0.25/−0.18 | 0.59/−0.24 | −0.15/0.00 | 0.09/0.14 | 0.63/0.10 | 1 | |||
| h. 5-day ant. rainfall | 0.26/0.18 | −0.34/−0.02 | −0.08/0.18 | 0.44/0.15 | 0.32/0.34 | −0.08/0.07 | −0.07/−0.03 | 1 | ||
| i. 10-day ant. rainfall | 0.17/−0.03 | −0.57/−0.32 | 0.01/0.13 | 0.48/0.20 | 0.36/0.25 | 0.04/−0.30 | 0.02/−0.02 | 0.71/0.44 | 1 | |
| j. Discharge | 0.60/−0.30 | −0.51/−0.95 | −0.51/−0.30 | 0.57/0.73 | 0.33/0.78 | −0.39/−0.21 | −0.09/0.22 | 0.37/0.08 | 0.30/0.25 | 1 |
| Data | Weekly 1 | Events 2 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Variable | Coef. | Std. Error | t-Value | Pr (>|t|) | Sign. 3 | Coef. | Std. Error | t-Value | Pr (>|t|) | Sign. 3 |
| (Intercept) | 2.72 | 0.76 | 3.57 | 0.00 | *** | 1.22 | 1.34 | 0.91 | 0.37 | |
| Log10 transformed turbidity | −0.77 | 0.46 | −1.66 | 0.10 | −0.46 | 0.33 | −1.41 | 0.16 | ||
| Log10 transformed suspended solid concentration | 0.89 | 0.35 | 2.52 | 0.01 | * | −0.07 | 0.25 | −0.27 | 0.79 | |
| Log10 transformed discharge | 0.51 | 0.21 | 2.40 | 0.02 | * | 0.44 | 0.28 | 1.58 | 0.12 | |
| Conductivity | 0.00 | 0.00 | 1.18 | 0.25 | 0.00 | 0.00 | 1.48 | 0.14 | ||
| Water temperature | −0.08 | 0.03 | −3.12 | 0.00 | ** | 0.02 | 0.05 | 0.41 | 0.69 | |
| Antecedent precipitation: 5-days | 0.01 | 0.01 | 1.08 | 0.29 | 0.01 | 0.01 | 1.33 | 0.19 | ||
| Antecedent precipitation: 10-days | 0.00 | 0.01 | 0.34 | 0.73 | 0.01 | 0.01 | 0.92 | 0.36 | ||
| Air temperature | −0.01 | 0.02 | −0.35 | 0.73 | 0.03 | 0.02 | 1.49 | 0.14 | ||
| Solar radiation | 0.00 | 0.00 | −0.20 | 0.84 | 0.00 | 0.00 | −0.92 | 0.36 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez-Carreras, N.; Ogorzaly, L.; Walczak, C.; Merlin, C.; Montargès-Pelletier, E.; Gantzer, C.; Iffly, J.-F.; Cauchie, H.-M.; Hissler, C. F-Specific RNA Bacteriophage Transport in Stream Water: Hydro-Meteorological Controls and Association with Suspended Solids. Water 2021, 13, 2250. https://doi.org/10.3390/w13162250
Martínez-Carreras N, Ogorzaly L, Walczak C, Merlin C, Montargès-Pelletier E, Gantzer C, Iffly J-F, Cauchie H-M, Hissler C. F-Specific RNA Bacteriophage Transport in Stream Water: Hydro-Meteorological Controls and Association with Suspended Solids. Water. 2021; 13(16):2250. https://doi.org/10.3390/w13162250
Chicago/Turabian StyleMartínez-Carreras, Núria, Leslie Ogorzaly, Cécile Walczak, Christophe Merlin, Emmanuelle Montargès-Pelletier, Christophe Gantzer, Jean-François Iffly, Henry-Michel Cauchie, and Christophe Hissler. 2021. "F-Specific RNA Bacteriophage Transport in Stream Water: Hydro-Meteorological Controls and Association with Suspended Solids" Water 13, no. 16: 2250. https://doi.org/10.3390/w13162250