
Environmental Drivers of Macroinvertebrate Assemblages within Peat Pool Habitat-Implication for Bioassessment

Department of Hydrobiology and Protection of Ecosystems, University of Life Sciences in Lublin, Dobrzańskiego 37, 20-262 Lublin, Poland
Water 2021, 13(17), 2369; https://doi.org/10.3390/w13172369
Submission received: 28 July 2021
/
Revised: 24 August 2021
/
Accepted: 26 August 2021
/
Published: 28 August 2021
(This article belongs to the Special Issue Abundance and Trophic Relationships in Freshwater Ecosystems)
Abstract
:Macroinvertebrates are a crucial component of wetland trophic webs. Many taxa are used as bioindicators of ecosystem change. However, relationships between macroinvertebrates and the environmental factors in peat pool habitats are still not well recognized. The present study shows the results of long-term studies during the years 2010–2020, on the responses of macroinvertebrates to the changes of environmental variables in a peat pool habitat formed as a result of peat exploitation on continental raised bog. The RDA analysis significantly explained 87.3% of the variance in macroinvertebrates abundances. Assemblages of most taxa (Anisoptera, Chironomidae, Ceratopogonidae, Coleoptera, Hydrachnidia and Tabanidae) showed a strong relationship with N-NO3 and pH. Moreover, densities of Chaoboridae larvae were explained by chlorophyll-a concentrations. Dominant taxon, Chironomidae, constituted from 48% to 87% of total faunal density. The highest proportions showed larvae of Psectrocladius sp. (gr. sordidellus) and Chironomus sp. Changes in the composition of macroinvertebrate fauna over a period of 10 years reflect the eutrophication process of the peat pool and deterioration of habitat conditions.
1. Introduction
Permanent pools are typical small water bodies for many northern bogs [1]. The pools are fed by rainwater and are usually dystrophic, characterized by a low pH, low primary production and nutrients, but high levels of dissolved organic matter and brown-colored water. These ponds make a vital contribution to biodiversity, by maintaining a high species diversity of macroinvertebrate in relation to other water bodies [2,3,4].
Aquatic macroinvertebrates play a crucial role in nutrient cycling and energy flow and are an essential link of aquatic trophic webs. Macroinvertebrates participate in the nutrients transfer from the sediments and the water column to higher trophic levels. Invertebrates are crucial for energy flow and sustainability of wetland ecosystems [5,6]. Aquatic invertebrates, particularly larvae of insects, are selectively consumed by ducks and other bird species [7]. These organisms varied between wetland types and may exhibit different responses to changes in the peatland environment [8,9].
Aquatic invertebrates have become common tools in the assessment of ecological status and water quality of ecosystems [6,10]. They are used in projects of wetland bioassessment because different macroinvertebrate taxa have shown specific habitat requirements and their community structure accurately reflects ecological conditions [11,12].
Despite their role as bioindicators, relationships between macroinvertebrates and the environmental variables controlling their diversity in peat pool habitats are still not sufficiently understood, particularly in the long-term perspective. Long-term study can be an effective method to track environmental changes in peat pool habitat. Changes in the structure of macroinvertebrates along this study led to recognizing the current condition of peat pool habitat. This paper presents a 10-year study, aimed at (1) analysis of species structure and density of macroinvertebrates associated with peat pool; (2) determination of significant environmental variables for macroinvertebrate diversity; and (3) evaluation of the potential role of macroinvertebrates as indicators of environmental conditions of peat pool.
2. Materials and Methods
2.1. Study Area
The study was conducted on the area of Krugłe Bagno peat bog (Polesie Lubelskie, eastern Poland, area ca. 6.5 ha, 51° N, 23° E). This is a former continental raised bog, almost completely exploited in the mid-20th century. At the present time its surface area is densely overgrown with Sphagnum mats, typical for transitional bogs. Krugłe Bagno is surrounded by a wide belt of mixed woodland, with domination of Pinus sylvestris L.; on the north-west side are buildings and arable fields. The studied peat pool is a shallow water body (max. depth 2.2 m) with a surface area of 0.5 ha. It was formed as a result of peat exploitation. Vegetation is dominated by Sphagnum angustifolium (C.C.O. Jensen ex Russow) and Sphagnum cuspidatum Ehrh. ex Hoffm.
2.2. Water Analysis
Three water samples for chemical analysis were taken at the same sites and dates as bottom fauna. pH, conductivity, and dissolved oxygen were recorded in situ by means of YSI 556 MPS electrode. TOC concentration was determined using PASTEL UV. Nutrients (N-NO3, TP, P-PO4) were analyzed at the laboratory, using spectrophotometric methods according to Hermanowicz et al. [13]. Chlorophyll-a was determined using spectrophotometric method after 24-h extraction with 90% acetone in the dark [14].
2.3. Macroinvertebrates Sampling
Macroinvertebrates were collected in spring (May), summer (July) and autumn (October) 2010, 2013, and 2020, using a tube apparatus (surface area 15.2 cm2). Each time and season three separate samples were taken (10 sediment cores per 1 sample). Each sample was sieved through the 250 µm mesh size, put into the plastic bag, and transported to the laboratory. Next, the macrofauna was selected from samples, preserved in 4% formaldehyde solution, counted, and identified according to Wiederholm [15] and Kołodziejczyk and Koperski [16]. Densities were calculated per m2 of the bottom surface.
2.4. Statistical Analysis
The effect of year and season for the structure of macroinvertebrate fauna was verified using a two-way analysis of variance (two-way ANOVA, year, season). Before the analysis, the Shapiro–Wilk test was used to check the normal distribution of data. The test of Kruskal–Wallis was used to compare significant (p < 0.05) differences between means. The analysis was performed using STATISTICA 13.0 software.
Detrended Correspondence Analysis (DCA) was used to measure the variance gradient of macrofauna data and then Redundancy Analysis (RDA) was performed to recognize the relationships between the macroinvertebrates and the environmental data. Variance partitioning was used to calculate proportions of variance explained by environmental variables. The significance of the ordination analysis and explanatory variables used in the model was tested under 999 permutations. The analyses were performed using CANOCO 5.0 Software (Microcomputer Power, Ithaca, NY, USA).
3. Results
3.1. Environmental Variables
Environmental conditions in the peat pool habitat varied between studied years (Table 1). Temperature depended on the season and fluctuated from 8.2 to 14.8 °C in autumn to 21.4 to 26.7 °C in summer. pH was typically low for dystrophic waters and ranged from 4.2 to 5.3. Concentrations of chlorophyll-a in 2010 and 2013 were low and varied between 4.7 and 26.8 µg dm−3. In 2020, these values were several times higher and oscillated between 37.8 and 101.8 µg dm−3. The concentration of TP, PO4, and N-NO3 changed dependently on the season and showed no clear trends.
3.2. Abundance and Domination Structure of Macroinvertebrates
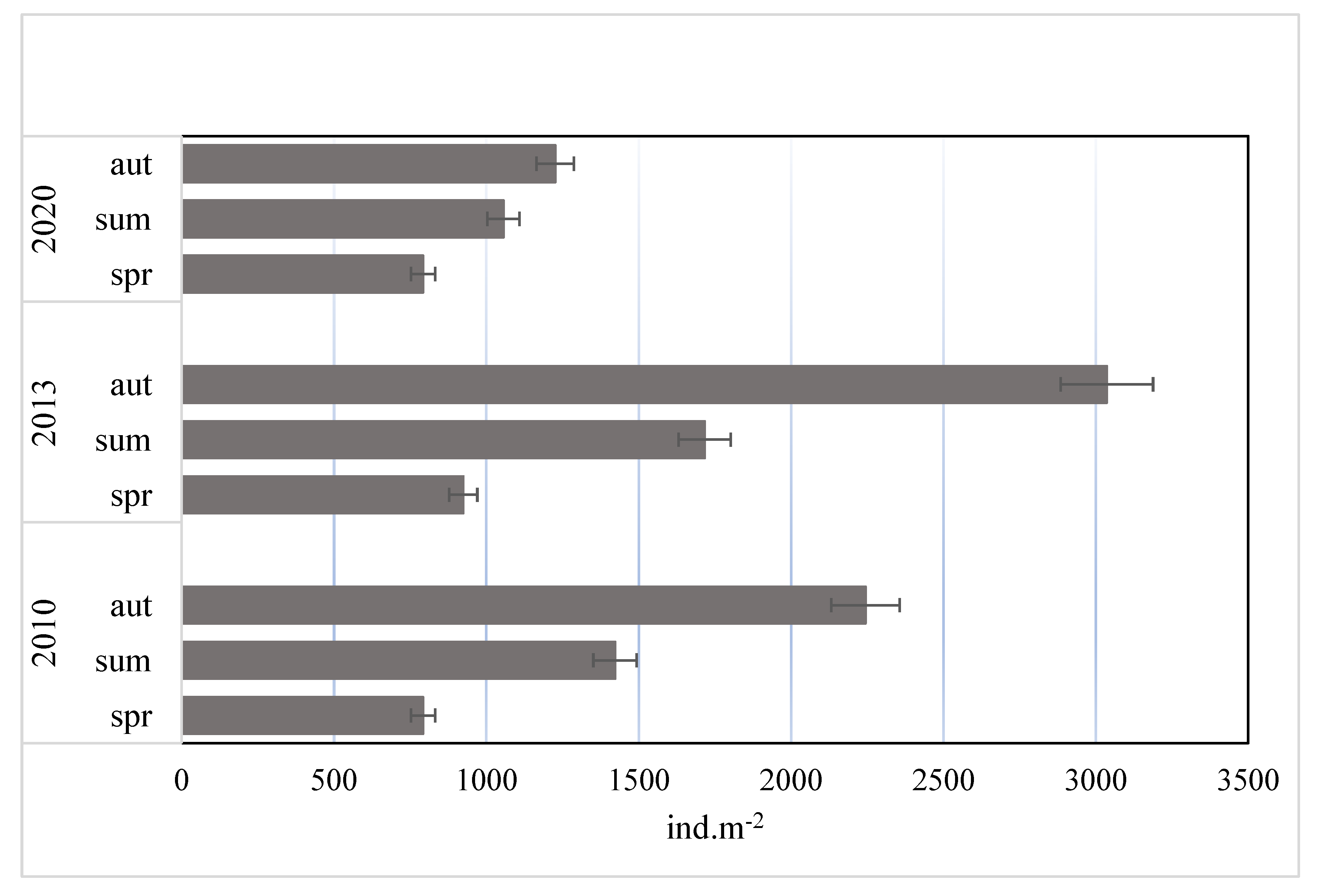
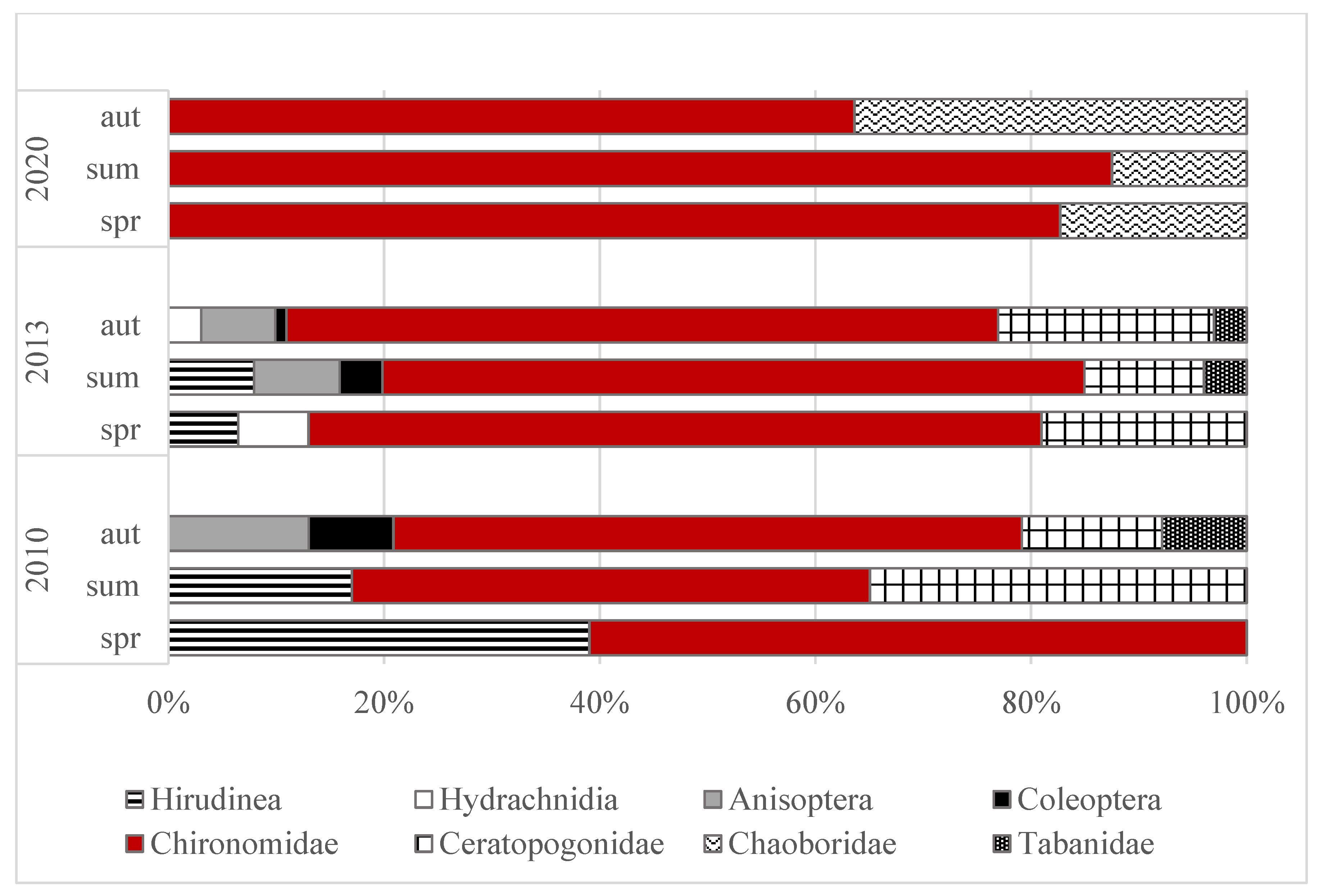
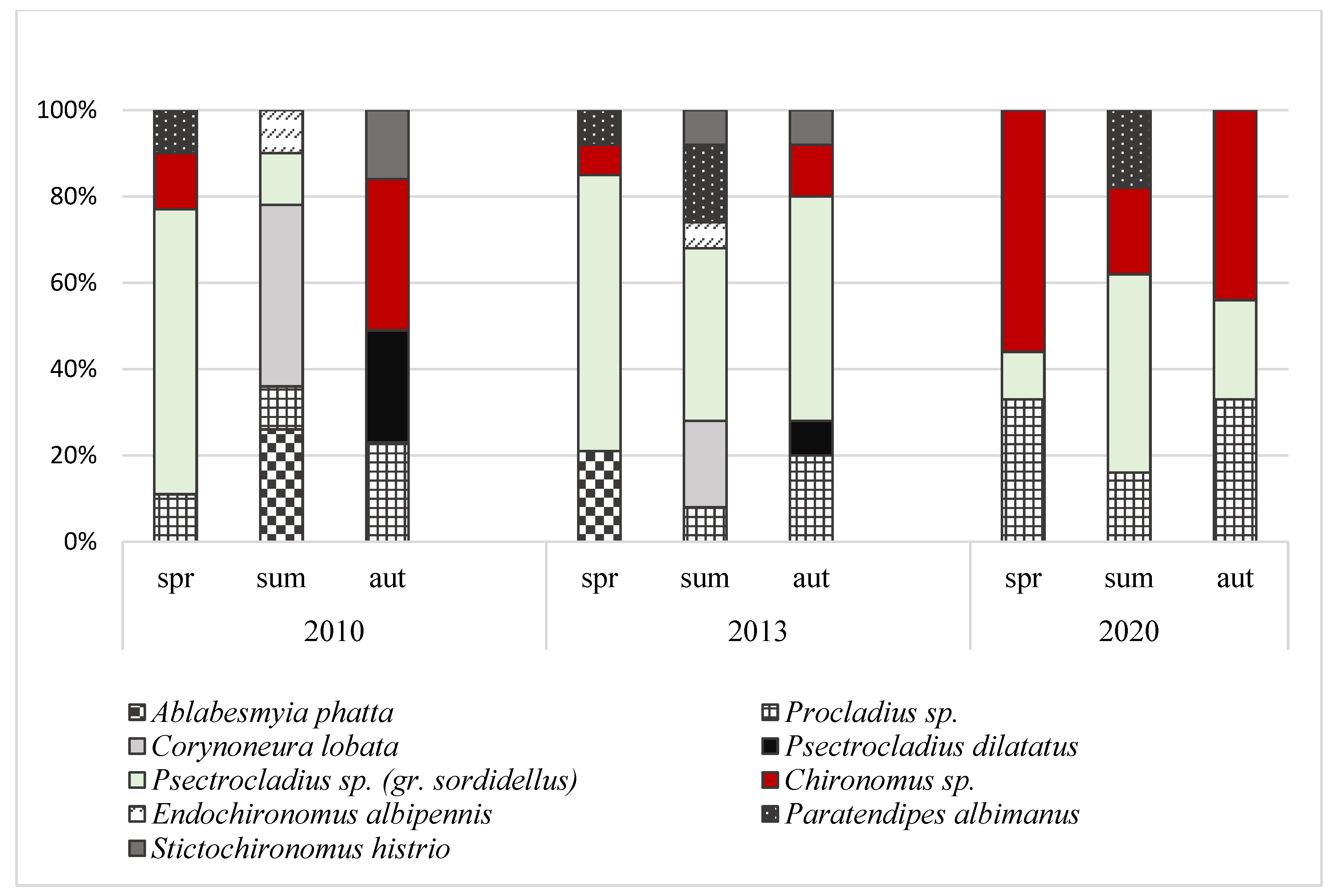
The abundances of macrofauna differed significantly between the years (ANOVA, F = 28.72; p = 0.004) and seasons (ANOVA, F = 11.38; p = 0.022) (Figure 1). In individual years of the study, the highest densities of macroinvertebrates were noted in autumn (1726–3036 ind. m−2) and the lowest in spring (764–992 ind. m−2). Generally there was a clear decline in the total density of bottom fauna in 2020 in comparison to the years 2010 and 2013, primarily in summer and autumn. In total, eight taxa of macrofauna were noted in the studied peat pool. Independently of the year and season, the highest relative abundances were found for larvae of Chironomidae (Figure 2). Their proportion varied from 48.2% (summer 2010) to 87.5% (summer 2020). A total of nine chironomid taxa were identified in this study (Figure 3). The highest proportions were registered to larvae of Psectrocladius sp. (gr. sordidellus) (11–66%), Chironomus sp. (7–56%), and Procladius sp. (8–33%).
3.3. Ordination Analysis
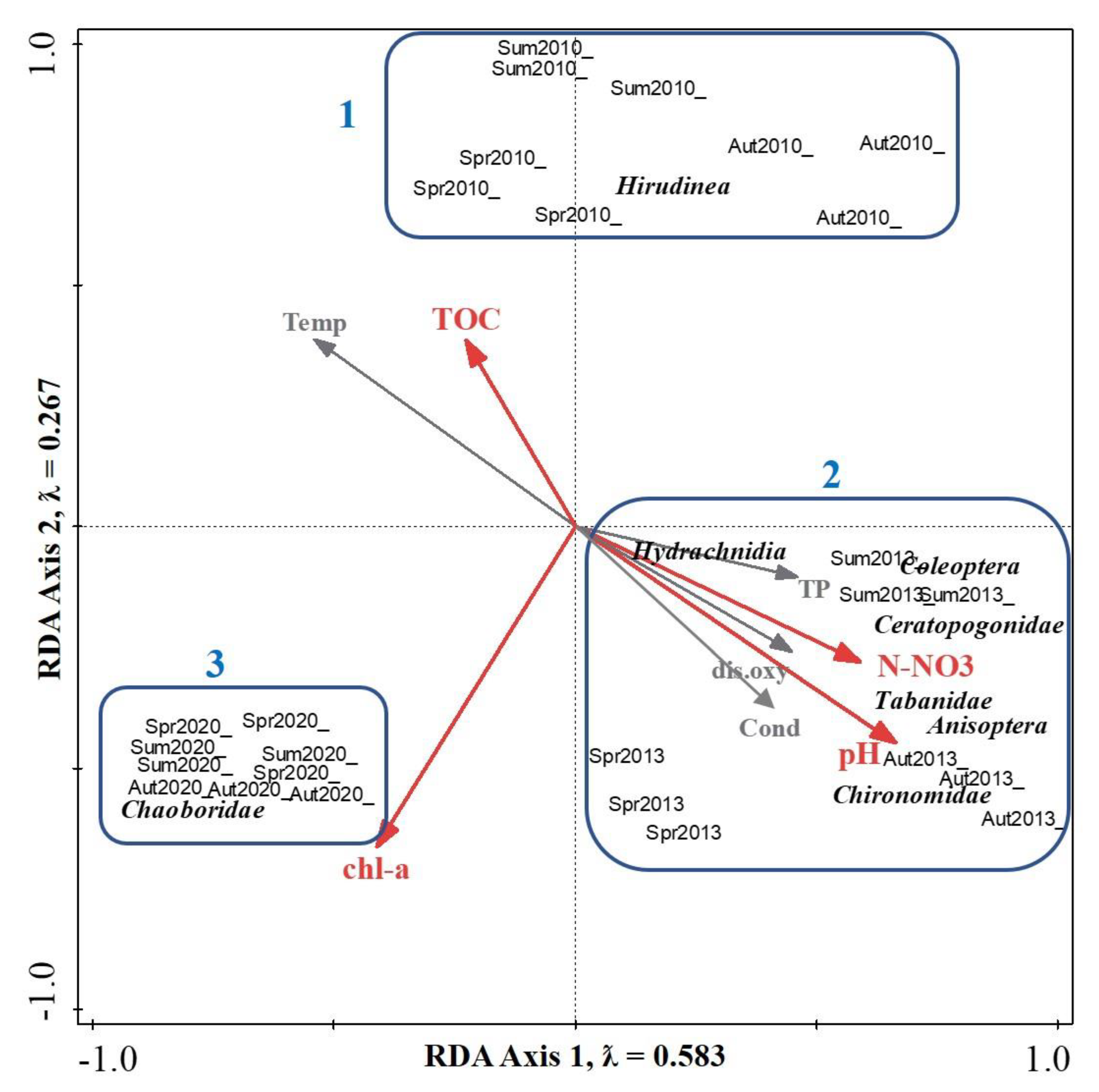
The RDA significantly explained 87.3% of the variance in macroinvertebrates abundances (p < 0.05). Results of variance partitioning showed that N-NO3 explained 44.3% (p = 0.002), pH 16.0% (p = 0.002), chlorophyll-a 15.1% (p = 0.014), and TOC 11.8% (p = 0.016) of the variance. Temperature, TP, P-PO4, dissolved oxygen, and conductivity were not significant (p > 0.05). On the ordination triplot, macroinvertebrate samples were separated, depending on the year of study, into three groups, 2010, 2013, and 2020 (Figure 4). The first group corresponds to abundance of Hirudinea; the second group to abundances of Anisoptera, Chironomidae, Ceratopogonidae, Coleoptera, Hydrachnidia, and Tabanidae; and the third group to relatively high abundances of Chaoboridae. The results of RDA indicate that N-NO3 (λ = 24.8; F = 19.9; p = 0.002) and pH (λ = 11.6; F = 9.7; p = 0.002) tend to affect the abundances of macroinvertebrate taxa belonging to the second group whereas chlorophyll-a (λ = 10.5; F = 14.1; p = 0.01) positively affects the abundance of the third group (Chaoboridae larvae). The position of TOC (λ = 9.8; F = 20.3; p = 0.01) gradient on the triplot suggests a negative relation of this variable to all macroinvertebrate taxa.
4. Discussion
Environmental conditions (acidity, brown-colored water, low N concentration) of the studied peat pool were typical for ombrotrophic Sphagnum-dominated peatbogs [17,18] and shallow dystrophic lakes [19]. Dystrophic conditions result in the low heterogeneity of the habitat and the low species diversity of macroinvertebrates in the studied ecosystem [20]. In the studied peat pool, macrophytes are represented by a few Sphagnum species. It can provide much fewer potential niches for species and cause a decrease in species diversity of macrofauna. Similar observations were reported by Beadle et al. [1] during studies of biodiversity in British peat pools. Homogeneity of the studied habitat could negatively affect the presence of invertebrate taxa, such as Coleoptera. Taxa of aquatic beetles are able to use different mesohabitats such as water column, steep-sided pool edges, and Sphagnum carpets [21,22]. Dense homogenous vegetation may affect capture rates and swimming capability of particular species [23]. In the study, Coleoptera were observed in 2010 and 2013 and their proportions did not exceed 10% of the total faunal density. Dominant taxon, larvae of Chironomidae, are an ubiquitous component of peat bog fauna. Many Chironomidae larvae primarily feed on algae and detritus and they serve as food for fish and waterfowl [24,25]. Regulatory structures (anal papillae) make many chironomid taxa tolerant to a wide range of water chemistry conditions [26].
Ordination analysis indicated pronounced effects of environmental variables, N-NO3, pH, and chlorophyll-a on peat pool macroinvertebrates. The role of N-NO3 probably results from the structure of the peat pool vegetation. The macrophyte diversity is low, there are only a few Sphagnum species such as S. cuspidatum and S. angustifolium. Sphagnum spp. are characterized by low tissue nutrient concentration and very low nutrient demands. Moreover these plants produce phenolic metabolites, which may exhibit antimicrobial activity [27,28]. These factors determine low rates of plant debris decomposition in Sphagnum-dominated wetlands [29] and may limit the abundance of detritus-feeding macroinvertebrates [30,31]. Chlorophyll-a is a primary indicator of algae biomass. Periphytic algae are the most important food source for macroinvertebrate grazers, such as chironomids. Availability of food resources is a key regulator of chironomid structure and distribution [32,33]. High chlorophyll-a content is associated with low water transparency. It may be related to the appearance of high numbers of Chaoboridae larvae in 2020. These larvae prefer cloudy water, thus avoiding the fish predation [34] and are recognized as being very tolerant to low oxygen concentrations [35]. Peat pool macroinvertebrates were affected by pH. Changes in pH may affect the birth rate and survival of macroinvertebrates [36], resulting in a significant decrease in their diversity. As it was observed by Woodcock et al. [26], pH has a direct effect on the species structure of chironomid communities. Under low pH, chironomids are typically dominated by fewer species [37]. Low species richness in the peat pool habitat is typical for eutrophic shallow lakes and small oxbow ponds [38,39,40].
5. Conclusions
The study confirmed the role of chironomids in bioassessment. Chironomid larvae are useful bioindicators of water quality such as pH and nutrients [41,42,43]. The most abundant chironomid taxa in the peat pool habitat, pelophilous Chironomus sp. and phytophilous larvae of Psectrocladius sp. (gr. sordidellus), are commonly reported as indicators of eutrophication processes of water bodies. Changes in the structure and abundance of invertebrate fauna over a period of 10 years clearly indicate the eutrophication process of the peat pool habitat. High concentrations of chlorophyll-a in 2020 suggest the possibility of cyanobacterial blooms and further deterioration of this peat pool habitat.
Funding
This research received no external funding.
Acknowledgments
The study was partially funded by the Polish National Science Foundation under Grant 2012/05/B/NZ8/01263 “Influence of “bottom-up” and “top-down” control mechanisms on the dynamics and diversity of protozoa in peat bog ecosystems (mesosystem experiments).
Conflicts of Interest
The author declare no conflict of interest.
References
- Beadle, J.M.; Brown, L.E.; Holden, J. Biodiversity and ecosystem functioning in natural bog pools and those created by rewetting schemes. WIREs Water 2015, 2, 65–84. [Google Scholar] [CrossRef]
- Maitland, P.S. New horizons—new species? The invertebrate fauna of unexplored aquatic habitats in Scotland. Aquat. Conserv. Mar. Freshw. Ecosyst. 1999, 9, 529–534. [Google Scholar] [CrossRef]
- Williams, P.; Whitfield, M.; Biggs, J.; Bray, S.; Fox, G.; Nicolet, P.; Sear, D.A. Comparative biodiversity of rivers, streams, ditches and ponds in an agricultural landscape in Southern England. Biol. Conserv. 2003, 115, 329–341. [Google Scholar] [CrossRef]
- Drinan, T.J.; Foster, G.N.; Nelson, B.H.; O’Halloran, J.; Harrison, S.S.C. Macroinvertebrate assemblages of peatland lakes: Assessment of conservation value with respect to anthropogenic land-cover change. Biol. Conserv. 2013, 158, 175–187. [Google Scholar] [CrossRef]
- Streever, W.J.; Portier, K.M.; Crisman, T.L. A comparison of dipterans from ten created and ten natural wetlands. Wetlands 1996, 16, 416–428. [Google Scholar] [CrossRef]
- Nelson, S.M.; Roline, R.A.; Thullen, J.S.; Sartoris, J.J.; Boutwell, J.E. Invertebrate assemblages and trace element bioaccumulation associated with constructed wetlands. Wetlands 2000, 20, 406–415. [Google Scholar] [CrossRef]
- King, T.S.; Wrubleski, D.A. Spatial and diel availability of flying insects as potential duckling food in prairie wetlands. Wetlands 1998, 18, 100–114. [Google Scholar] [CrossRef]
- Hájková, P.; Bojková, J.; Fránková, M.; Opravilová, V. Disentangling the effects of water chemistry and substratum structure on moss-dwelling unicellular and multicellular microorganisms in spring-fens. J. Limnol. 2011, 70, 54–64. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Langdon, P.G.; Yan, Y.; Wang, S.; Zheng, Z.; Zhang, Z. Chironomid communities from subalpine peatlands in subtropical China as indicators of environmental change. J. Paleolimnol. 2019, 62, 165–179. [Google Scholar] [CrossRef]
- Galbrand, C.; Lemieux, I.G.; Ghaly, A.E.; Cote, A.; Verma, M. Assessment of wetland biological integrity using aquatic macroinvertebrates. J. Biol. Sci. 2007, 7, 52–65. [Google Scholar] [CrossRef] [Green Version]
- Spieles, D.J.; Mitsch, W.J. Macroinvetrebrate community structure in high- and low- nutrient constructed wetlands. Wetlands 2000, 20, 716–729. [Google Scholar] [CrossRef]
- Stewart, T.W.; Downing, J.A. Macroinvertebrate communities and environmental conditions in recently constructed wetlands. Wetlands 2008, 28, 141–150. [Google Scholar] [CrossRef]
- Hermanowicz, W.; Dojlido, J.; Dożańska, W.; Kosiorowski, B.; Zerze, J. Fizycznochemiczne Badanie Wody i Ścieków; Arkady: Warszawa, Poland, 1999. [Google Scholar]
- Golterman, H.L. Methods for chemical analysis of freshwaters. In IBP Handbook No. 8; Blackwell Scientific Publications: Oxford, UK, 1969. [Google Scholar]
- Wiederholm, T. Chironomidae of the holartic region. Keys and diagnoses. Part 1. Larvae. Entomol. Scand. 1986, 19, 1–457. [Google Scholar]
- Kołodziejczyk, A.; Koperski, P. Bezkręgowce Słodkowodne Polski. Klucz do Oznaczania oraz Podstawy Biologii I Ekologii Makrofauny; Wydawnictwo Uniwersytetu Warszawskiego: Warszawa, Poland, 2000; pp. 1–250. [Google Scholar]
- Mataloni, G.G.; González-Garraza, G.; Vinocur, A. Landscape-driven environmental variability largely determines abiotic characteristics and phytoplankton patterns in peat bog pools (Tierra del Fuego, Argentina). Hydrobiologia 2015, 751, 105–125. [Google Scholar] [CrossRef]
- Küppers, G.C.; González-Garraza, G.C.; Quiroga, M.V.; Lombardo, R.; Marinone, M.C.; Vinocur, A.; Mataloni, G. Drivers of highly diverse planktonic ciliate assemblages in peat bog pools from Tierra del Fuego (Argentina). Hydrobiologia 2016, 773, 117–134. [Google Scholar] [CrossRef]
- Górniak, A.; Jekaterynczuk-Rudczyk, E.; Dobrzy, P. Hydrochemistry of three dystrophic lakes in Northeastern Poland (in Polish). Acta Hydrohichim. Hydrobiol. 1999, 27, 12–18. [Google Scholar] [CrossRef]
- Luoto, T.P. Dystrophy in determining midge community composition in boreal lakes. Écoscience 2013, 20, 391–398. [Google Scholar] [CrossRef]
- Foster, G.N.; Bilton, D.T. The aquatic coleoptera associated with patterned mires. In Patterned Mires and Mire Pools: Origin and Development: Flora and Fauna: Proceedings, University of Durham, 6–7 April 1998; Standen, V., Tallis, J.H., Meade, R., Eds.; British Ecological Society: London, UK, 1998; pp. 65–84. [Google Scholar]
- Fairchild, G.W.; Faulds, A.M.; Matta, J.F. Beetle assemblages in ponds: Effects of habitat and site age. Freshw. Biol. 2000, 44, 523–534. [Google Scholar] [CrossRef]
- Ribera, I.; Foster, G.N.; Holt, W.V. Functional types of diving beetle (Coleoptera: Hygrobiidae and Dytiscidae), as identified by comparative swimming behaviour. Biol. J. Linn. Soc. 1997, 61, 537–558. [Google Scholar] [CrossRef]
- Batzer, D.P.; Wissinger, S.A. Ecology of insect communities in nontidal wetlands. Annu. Rev. Entomol. 1996, 41, 75–100. [Google Scholar] [CrossRef]
- Richardson, W.B.; Ziegler, S.J.; Dewey, M.R. Bioenergetic relations in submerged aquatic vegetation: An experimental test of prey use by juvenile bluegills. Ecol. Freshw. Fish 1998, 7, 1–12. [Google Scholar] [CrossRef]
- Woodcock, T.; Longcore, J.; McAuley, D.; Mingo, T.; Bennatti, C.R.; Stromborg, K. The role of pH in structuring communities of maine wetland macrophytes and chironomid larvae (Diptera). Wetlands 2005, 25, 306–316. [Google Scholar] [CrossRef] [Green Version]
- Rudolph, H.J.; Samland, J. Occurrence and metabolism of sphagnum acid in the cell walls of bryophytes. Phytochemistry 1985, 24, 745–749. [Google Scholar] [CrossRef]
- Jassey, V.E.; Chiapusio, G.; Binet, P.; Buttler, A.; Laggoun, F.; Delarue, F.; Bernard, N.; Mitchell, E.A.; Toussaint, M.; Francez, A.; et al. Above- and belowground linkages in Sphagnum peatland: Climate warming affects plant–microbial interactions. Glob. Change Biol. 2013, 19, 811–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aerts, R.; Wallen, B.; Malmer, N. Growth-limiting nutrients in Sphagnum-dominated bogs subject to low a high atmospheric nitrogen supply. J. Ecol. 1992, 80, 131–140. [Google Scholar] [CrossRef]
- Limpens, J.; Berendse, F.; Klees, H. N deposition affects N availability in interstitial water, growth of Sphagnum and invasion of vascular plants in bog vegetation. New Phytol. 2003, 157, 339–347. [Google Scholar] [CrossRef]
- van Duinen, G.A.; Vermonden, K.; Brock, A.M.T.; Leuven, R.S.E.W.; Smolders, A.J.P.; van der Velde, G.; Verberk, W.C.E.P.; Esselink, H. Basal food sources for the invertebrate food web in nutrient poor and nutrient enriched raised bog pools. Proc. Neth. Entomol. Soc. Meet. 2006, 17, 37–44. [Google Scholar]
- Taylor, A.N.; Batzer, D.P. Spatial and temporal variation in invertebrate consumer diets in forested and herbaceous wetlands. Hydrobiologia 2010, 651, 145–159. [Google Scholar] [CrossRef]
- Tarkowska-Kukuryk, M. Periphytic algae as food source for grazing chironomids in a shallow phytoplankton dominated lake. Limnologica 2013, 43, 254–264. [Google Scholar] [CrossRef]
- Wissel, B.; Yan, N.D.; Ramcharan, C.W. Predation and refugia: Implications for Chaoborus abundance and species composition. Freshw. Biol. 2003, 48, 1421–1431. [Google Scholar] [CrossRef]
- Jäger, I.S.; Walz, N. Chaoborus flavicans (Diptera) is an oxy-regulator. Arch. Hydrobiol. 2002, 155, 401–411. [Google Scholar] [CrossRef]
- Walag, A.M.P.; Canencia, M.O.P. Physico-chemical parameters and macrobenthic invertebrates of the intertidal zone of Gusa, Cagayan de Oro city, Philippines. Adv. Environ. Sci. 2016, 8, 71–82. [Google Scholar]
- Tokeshi, M. Species abundance patterns and community structure. Adv. Ecol. Res. 1993, 24, 111–186. [Google Scholar]
- Toth, M.; Arva, D.; Sandor, A.N.; Specziar, A. Species diversity and abundance of plant-dwelling chironomids across hierarchical habitat and seasonal scales in the oxbow lakes of River Tisza, Hungary. Fundam. Appl. Limnol. 2013, 182, 309–321. [Google Scholar] [CrossRef]
- Wazbinski, K.E.; Quinlan, R. Midge (Chironomidae, Chaoboridae, Ceratopogonidae) assemblages and their relationship with biological and physicochemical variables in shallow, polymictic lakes. Freshw. Biol. 2013, 58, 2464–2480. [Google Scholar] [CrossRef]
- Tarkowska-Kukuryk, M.; Mieczan, T. Distribution and environmental determinants of chironomids(Diptera: Chironomidae) in Sphagnum microhabitats. Pol. J. Environ. Stud. 2014, 23, 483–490. [Google Scholar]
- Brodersen, K.P.; Quinlan, R. Midges as palaeoindicators of lake productivity eutrophication and hypolimnetic oxygen. Quat. Sci. Rev. 2006, 25, 1995–2012. [Google Scholar] [CrossRef]
- Weller, M.O.; Bossart, J.L. Insect community diversity tracks degradation and recovery of a wastewater assimilation marsh in southeast Louisiana. Wetlands 2017, 37, 1–13. [Google Scholar] [CrossRef]
- Jyväsjärvi, J.; Virtanen, R.; Ilmonen, J.; Paasivirta, L.; Muotka, T. Identifying taxonomic and functional surrogates for spring biodiversity conservation. Conserv. Biol. 2018, 32, 883–893. [Google Scholar] [CrossRef]
Figure 1.
Seasonal changes of mean densities (±SD) of macroinvertebrates in peat pool habitat during 10 years of study.
Figure 1.
Seasonal changes of mean densities (±SD) of macroinvertebrates in peat pool habitat during 10 years of study.

Figure 2.
Changes of relative abundances of macroinvertebrate fauna in peat pool habitat during 10 years of study.
Figure 2.
Changes of relative abundances of macroinvertebrate fauna in peat pool habitat during 10 years of study.

Figure 3.
Domination structure of Chironomidae larvae in peat pool habitat during 10 years of study.
Figure 3.
Domination structure of Chironomidae larvae in peat pool habitat during 10 years of study.

Figure 4.
RDA triplot showing macroinvertebrate taxa, environmental variables, and samples. Solid red arrows indicate significant variables based on Monte Carlo permutation test (p < 0.05). Spr2010, Sum2010, Aut2010-samples collected in spring, summer, and autumn 2010; Spr2013, Sum2013, Aut2013-samples collected in spring, summer, and autumn 2013; Spr2020, Sum2020, Aut2020-samples collected in spring, summer, and autumn 2020.
Figure 4.
RDA triplot showing macroinvertebrate taxa, environmental variables, and samples. Solid red arrows indicate significant variables based on Monte Carlo permutation test (p < 0.05). Spr2010, Sum2010, Aut2010-samples collected in spring, summer, and autumn 2010; Spr2013, Sum2013, Aut2013-samples collected in spring, summer, and autumn 2013; Spr2020, Sum2020, Aut2020-samples collected in spring, summer, and autumn 2020.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Mean values (±SD) of physical and chemical water parameters of peat pool habitat.
| 2010 | 2013 | 2020 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Spr | Sum | Aut | Spr | Sum | Aut | Spr | Sum | Aut | |
| Temp (°C) | 17.8 ± 3.4 | 24.8 ± 5.3 | 8.2 ± 0.8 | 20.2 ± 2.5 | 21.4 ± 0.1 | 14.8 ± 0.2 | 20.3 ± 0.9 | 26.7 ± 0.5 | 18.8 ± 0.5 |
| pH | 4.7 ± 0.1 | 3.8 ± 1.2 | 5.3 ± 0.9 | 4.2 ± 0.5 | 5.2 ± 0.3 | 4.9 ± 0.4 | 4.2 ± 0.1 | 4.5 ± 0.3 | 4.3 ± 0.2 |
| Cond (µS cm−1) | 43 ± 9 | 50 ± 5 | 62 ± 2 | 45 ± 3 | 49 ± 2 | 54 ± 9 | 35 ± 13 | 56 ± 3 | 44 ± 9 |
| Dis.oxy (mg dm−3) | 5.0 ± 2.5 | 5.4 ± 1.9 | 6.8 ± 1.1 | 6.2 ± 1.9 | 6.7 ± 1.8 | 5.4 ± 2.3 | 6.0 ± 1.6 | 5.2 ± 2.6 | 5.9 ± 2.5 |
| N-NO3 (mg dm−3) | 0.234 ± 0.09 | 0.734 ± 0.24 | 1.276 ± 0.21 | 0.540 ± 0.04 | 0.5904 ± 0.07 | 0.670 ± 0.09 | 0.019 ± 0.02 | 0.013 ± 0.01 | 0.024 ± 0.05 |
| P-PO4 (mg dm−3) | 0.031 ± 0.014 | 0.182 ± 0.01 | 0.017 ± 0.01 | 0.077 ± 0.04 | 0.074 ± 0.48 | 0.058 ± 0.48 | 0.017 ± 0.04 | 0.105 ± 0.001 | 0.014 ± 0.002 |
| TP (mg dm−3) | 0.129 ± 0.031 | 0.547 ± 0.026 | 0.497 ± 0.123 | 0.098 ± 0.154 | 0.103 ± 0.163 | 0.521 ± 0.162 | 0.046 ± 0.011 | 0.429 ± 0.041 | 0.039 ± 0.009 |
| TOC (mg dm−3) | 51.5 ± 9.1 | 63.3 ± 11.2 | 60.3 ± 6.4 | 48.5 ± 8.5 | 30.4 ± 2.3 | 10.6 ± 0.4 | 27.3 ± 1.5 | 55.7 ± 3.4 | 41.9 ± 2.2 |
| Chl-a (µg dm−3) | 4.8 ± 1.3 | 26.8 ± 1.9 | 69.7 ± 10.4 | 6.1 ± 71.8 | 6.6 ± 0.5 | 16.3 ± 1.3 | 37.8 ± 4.6 | 74.6 ± 7.1 | 101.8 ± 17.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tarkowska-Kukuryk, M. Environmental Drivers of Macroinvertebrate Assemblages within Peat Pool Habitat-Implication for Bioassessment. Water 2021, 13, 2369. https://doi.org/10.3390/w13172369
AMA Style
Tarkowska-Kukuryk M. Environmental Drivers of Macroinvertebrate Assemblages within Peat Pool Habitat-Implication for Bioassessment. Water. 2021; 13(17):2369. https://doi.org/10.3390/w13172369
Chicago/Turabian StyleTarkowska-Kukuryk, Monika. 2021. "Environmental Drivers of Macroinvertebrate Assemblages within Peat Pool Habitat-Implication for Bioassessment" Water 13, no. 17: 2369. https://doi.org/10.3390/w13172369
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.