
Effects of Water Depth on the Growth of the Submerged Macrophytes Vallisneria natans and Hydrilla verticillata: Implications for Water Level Management


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Sampling and Analytical Methods
3. Results
3.1. Water Physical and Chemical Parameters
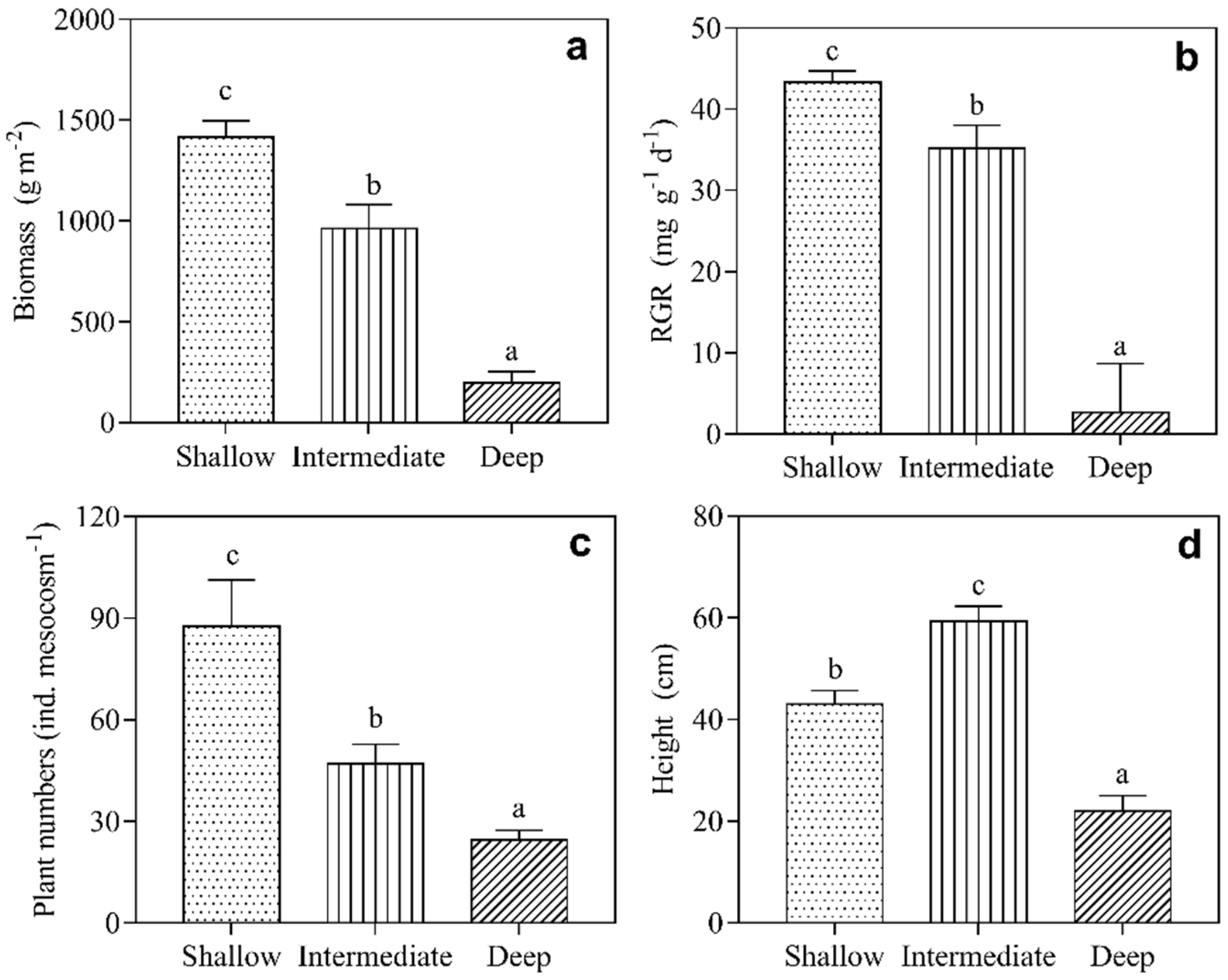
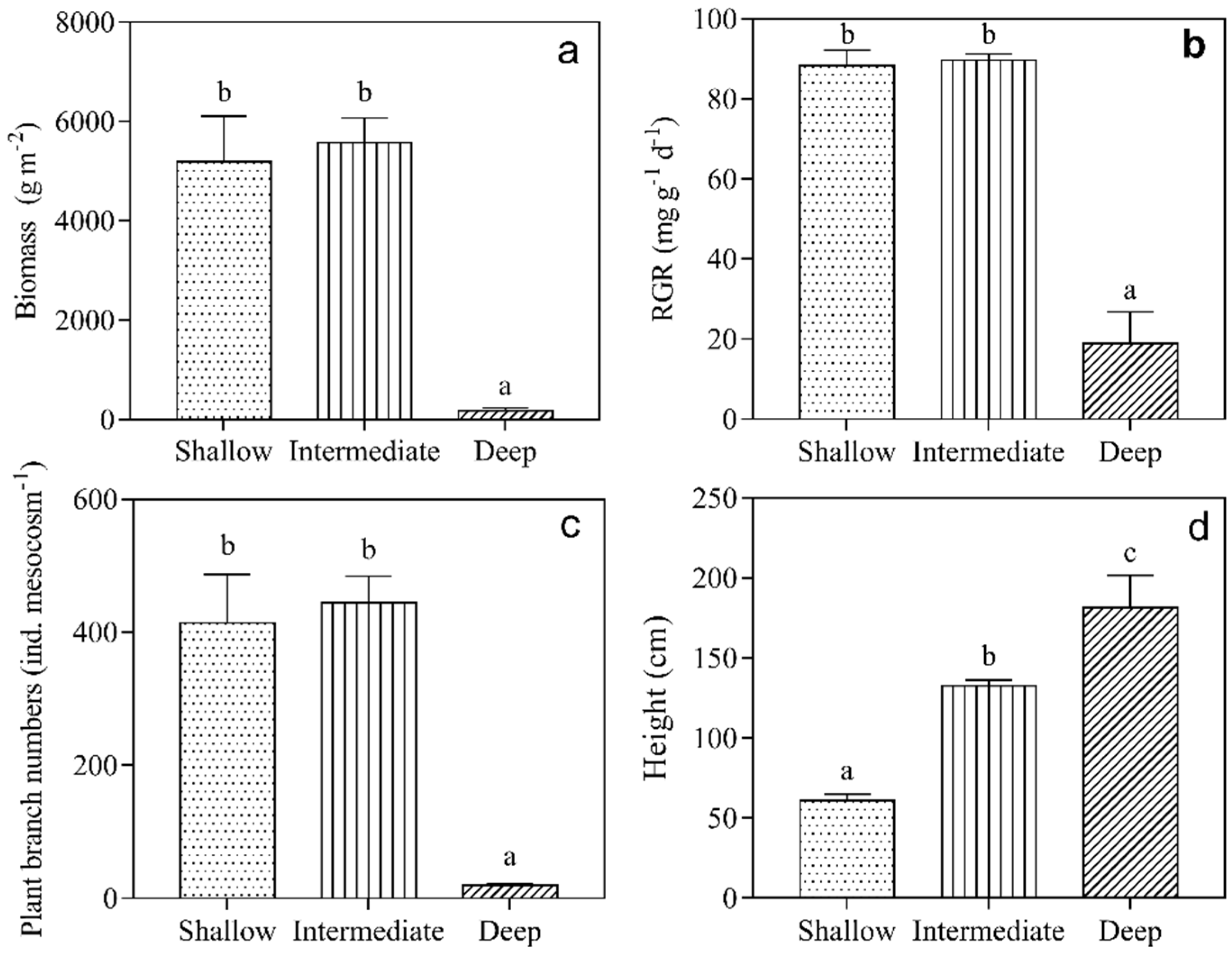
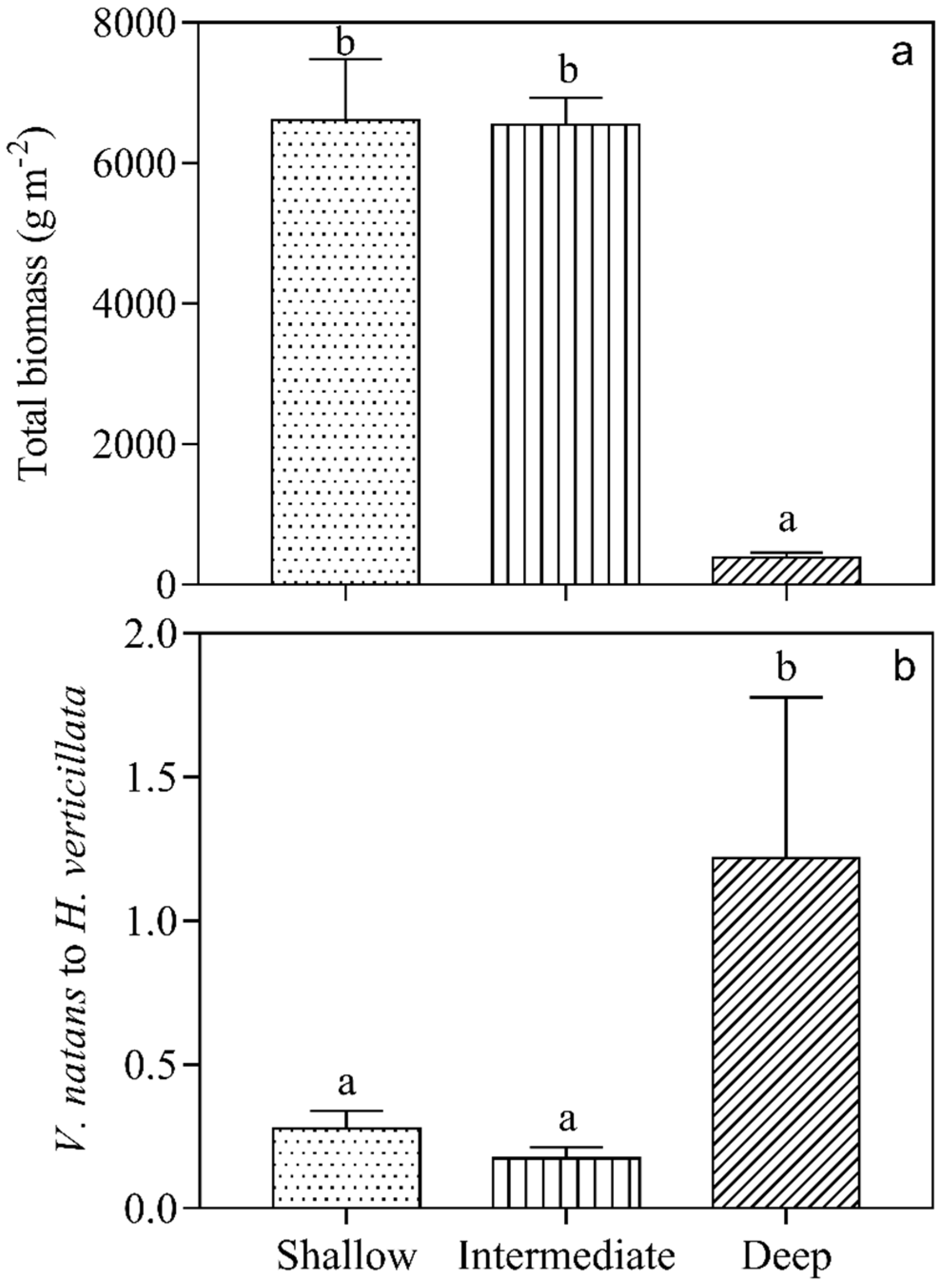
3.2. Growth of Submerged Macrophytes
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Scheffer, M.; Hosper, S.H.; Meijer, M.L.; Moss, B.; Jeppesen, E. Alternative equilibria in shallow lakes. Trends Ecol. Evol. 1993, 8, 275–279. [Google Scholar] [CrossRef]
- Jeppesen, E.; Søndergaard, M.; Søndergaard, M.; Christoersen, K. (Eds.) The Structuring Role of Submerged Macrophytes in Lakes; Springer-Verlag New York, Inc.: New York, NY, USA, 1998. [Google Scholar]
- Stansfield, J.H.; Perrow, M.R.; Tench, L.D.; Jowitt, A.; Taylor, A. Submerged macrophytes as refuges for grazing Cladocera against fish [-3pt] predation: Observations on seasonal changes in relation to macrophyte cover and predation pressure. Hydrobiologia 1997, 342, 229–240. [Google Scholar] [CrossRef]
- Maceina, M.; Bettoli, P.; Klussmann, W.; Betsill, R.K.; Noble, R.L. Effect of aquatic macrophyte removal on recruitment and growth of black crappies and white crappies in Lake Conroe, Texas. N. Am. J. Fish. Manag. 1991, 11, 556–563. [Google Scholar] [CrossRef]
- Moss, B. Engineering and biological approaches to the restoration from eutrophication of shallow lakes in which aquatic plant communities are important components. Hydrobiologia 1990, 200/201, 367–377. [Google Scholar] [CrossRef]
- Li, P. Researches on the Relationships between Periphytic Algae, Phytoplankton and Vallisneria natans [Dissertation]. Master’s Thesis, Huazhong Agricultural University, Wuhan, China, 2012. [Google Scholar]
- Middelboe, A.L.; Markager, S. Depth limits and minimum light requirements of freshwater macrophytes. Freshw. Biol. 1997, 37, 553–568. [Google Scholar] [CrossRef]
- Chambers, P.A.; Kaiff, J. Depth distribution and biomass of submersed aquatic macrophyte communities in relation to Secchi depth. Can. J. Fish. Aquat. Sci. 1985, 42, 701–709. [Google Scholar] [CrossRef]
- Li, L.; Bonser, S.P.; Lan, Z.; Xu, L.; Chen, J.; Song, Z. Water depth affects reproductive allocation and reproductive allometry in the submerged macrophyte Vallisneria natans. Sci. Rep. 2017, 7, 16842. [Google Scholar] [CrossRef]
- Li, L.; Lan, Z.C.; Chen, J.K.; Song, Z.P. Allocation to clonal and sexual reproduction and its plasticity in Vallisneria spinulosa along a water-depth gradient. Ecosphere 2018, 9, e02070. [Google Scholar] [CrossRef] [Green Version]
- Beklioglu, M.; Altinayar, G.; Tan, C.O. Water level control over submerged macrophyte development in five shallow lakes of Mediterranean Turkey. Arch. fur Hydrobiol. 2006, 166, 535–556. [Google Scholar] [CrossRef]
- Havens, K.E.; Fox, D.; Gornak, S.; Hanlon, C. Aquatic vegetation and largemouth bass population responses to water-level variations in Lake Okeechobee, Florida (USA). Hydrobiologia 2005, 539, 225–237. [Google Scholar] [CrossRef]
- Geest, G.; Wolters, H.; Roozen, F.; Coops, H.; Roijacker, R.; Buijse, A. Water-level fluctuations affect macrophyte richness in floodplain lakes. Hydrobiologia 2005, 539, 239–248. [Google Scholar] [CrossRef]
- Li, D.H.; Yang, S.; Fang, T.; Liu, J.Y.; Liu, Y.D. Recovery of aquatic macrophytes by use of water level regulation method in eutrophicated lakes—A case study of Wuli Lake, Wuxi city. Environ. Sci. Technol. 2008, 31, 59–62. [Google Scholar]
- Yu, J.L.; Liu, Z.W.; Li, K.Y.; Chen, F.Z.; Guan, B.H.; Hu, Y.H.; Zhong, P.; Tang, Y.L.; Zhao, X.F.; He, H.; et al. Restoration of shallow lakes in subtropical and tropical China: Response of nutrients and water clarity to biomanipulation by fish removal and submerged plant transplantation. Water 2016, 8, 438. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.W.; Hu, J.R.; Zhong, P.; Zhang, X.F.; Ning, J.J.; Larsen, S.E.; Chen, D.Y.; Gao, Y.M.; He, H.; Jeppesen, E. Successful restoration of a tropical shallow eutrophic lake: Strong bottom-up but weak top-down effects recorded. Water Res. 2018, 146, 88–97. [Google Scholar] [CrossRef]
- Chambers, P.A.; Prepas, E.E. Competition and coexistence in submerged aquatic plant communities: The effects of species interactions versus abiotic factors. Freshw. Biol. 2010, 23, 541–550. [Google Scholar] [CrossRef]
- Gao, F.; Zhang, Y.M.; Fei, Y.; Ma, M.J.; Ding, Y.R. Growth and photosynthetic fluorescence characteristics responses of four submersed macrophytes to rising water level. J. Ecol. Rural. Environ. 2017, 33, 341–348. [Google Scholar] [CrossRef]
- Ge, F.J.; Liu, B.Y.; Lu, Z.Y.; Gao, Y.N.; Wu, Z.B. Effects of light intensity on growth and phenolic contents of Myriophyllum spicatum. Environ. Sci. Technol. 2012, 35, 30–34. [Google Scholar] [CrossRef]
- Liu, W.L.; Hu, W.P.; Chen, Y.G.; Gu, X.H.; Hu, Z.X.; Chen, Y.W.; Ji, J. Temporal and spatial variation of aquatic macrophytes in west Taihu Lake. Acta Ecol. Sin. 2007, 27, 159–170. [Google Scholar]
- Jin, X.; Tu, Q. The Standard Methods for Obervation and Analysis in Lake Eutrophication; Environmental Science Press: Beijing, China, 1990. (In Chinese) [Google Scholar]
- SEPA. Analytical Methods for Water and Waster Water Monitor, 4th ed.; Chinese Environmental Science Press: Beijing, China, 2002. (In Chinese) [Google Scholar]
- Langhans, R.W.; Tibbitts, T.W. Plant Growth Chamber Handbook; Iowa Agriculture and Home Economics Experimental Station: Ames, IA, America, 1997. [Google Scholar] [CrossRef]
- Li, Q.S.; Huang, Q.; Li, Y.J.; Han, Y.Q.; Jin, H.; He, H.; Li, K.Y. Effects of water depth on growth of submerged macrophytes Vallisneria natans and Myriophyllum spicatum. J. Lake Sci. 2019, 31, 1045–1054. [Google Scholar] [CrossRef] [Green Version]
- Jin, S.; Ibrahim, M.; Muhammad, S.; Khan, S.; Li, G. Light intensity effects on the growth and biomass production of submerged macrophytes in different water strata. Arab. J. Geosci. 2020, 13, 948. [Google Scholar] [CrossRef]
- Dong, B.L.; Zhou, Y.Q.; Jeppesen, E.; Shi, K.; Qin, B.Q. Response of community composition and biomass of submerged macrophytes to variation in underwater light, wind and trophic status in a large eutrophic shallow lake. J. Environ. Sci. 2021, 103, 298–310. [Google Scholar] [CrossRef] [PubMed]
- Su, W.H.; Zhang, G.F.; Zhang, Y.S.; Xiao, H.; Xia, F. Photosynthetic characteristics of five submerged macrophytes. Acta Hydrobiol. Sin. 2004, 28, 391–395. [Google Scholar]
- Diao, R.X.; Xu, Z.A.; Wu, D.H. In situ enclosure study on growth of Vallisneria natans at different water depths in Taihu Lake. J. Hydroecology 2017, 38, 43–47. [Google Scholar] [CrossRef]
- Fu, H.; Yuan, G.X.; Cao, T.; Ni, L.Y.; Zhang, X.L. Clonal growth and foraging behavior of a submerged macrophyte Vallisneria natans in response to water depth gradient. J. Lake Sci. 2012, 24, 705–711. [Google Scholar] [CrossRef] [Green Version]
- Xiao, K.Y.; Yu, D.; Wu, Z. Differential effects of water depth and sediment type on clonal growth of the submersed macrophyte Vallisneria natans. Hydrobiologia 2007, 589, 265–272. [Google Scholar] [CrossRef]
- Gu, Y.F.; Wang, J.; Wang, J.; Fang, G.S.; Han, L. Morphological response and growth strategy of the submerged macrophyte Vallisneria natans under different water depths. J. Lake Sci. 2017, 29, 654–661. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Wang, G.X. Influence of silts on growth and development of Hydrilla verticillata seeding in silt waters. Acta Ecol. Sin. 2010, 30, 995–1002. [Google Scholar]
- Wu, X.D.; Wang, G.X.; Cheng, Z.Y.; Wei, H.N.; Li, Z.G.; Wang, L.Z. Response of Hydrilla verticillata in growth to water depth gradient. J. Ecol. Rural. Environ. 2011, 27, 40–45. [Google Scholar]
- Li, K.Y.; Liu, Z.W.; Wang, C.Z.; Wang, C.H.; Shi, J.D.; Hu, Y.H. Effect of low dissolved oxygen on the growth of Vallisneria spiralis. Resour. Environ. Yangtze Basin 2006, 15, 670–673. [Google Scholar]
- Wang, Q.; Zhou, X.D.; Luo, J.H.; Chen, C. Remote sensing monitoring and analysis of dominant species of submerged vegetation in Taihu Lake over last 30 years. Water Resour. Prot. 2016, 32, 123–129. [Google Scholar] [CrossRef]
- Xiong, F.; Li, W.C.; Pan, J.Z.; Xia, T.X.; Li, A.Q. Distribution and community structure characteristics of submerged macrophytes in Lake Fuxian, Yunnan Province. Acta Bot. Yunnanica 2006, 28, 277–282. [Google Scholar]
- Regmi, T.; Shah, D.N.; Doody, T.M.; Cuddy, S.; Shah, R. Hydrological alteration induced changes on macrophyte community composition in sub-tropical floodplain wetlands of Nepal. Aquat. Bot. 2021, 103413. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Shallow | Intermediate | Deep | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Max | Min | Average | Max | Min | Average | Max | Min | Average | |
| Light (target depth: surface) | 0.437 | 0.181 | 0.304 | 0.132 | 0.029 | 0.075 | 0.048 | 0.005 | 0.023 |
| TN (mg L−1) | 1.60 | 1.04 | 1.26 | 1.36 | 1.06 | 1.21 | 1.41 | 1.04 | 1.21 |
| TP (μg L−1) | 70 | 42 | 56 | 67 | 44 | 57 | 76 | 47 | 60 |
| TDN (mg L−1) | 0.96 | 0.78 | 0.87 | 0.95 | 0.79 | 0.88 | 0.94 | 0.78 | 0.87 |
| TDP (μg L−1) | 37 | 20 | 28 | 37 | 21 | 28 | 39 | 20 | 28 |
| Chl-a (μg L−1) | 28 | 5 | 17 | 25 | 6 | 15 | 27 | 6 | 17 |
| Variables | F | Df | p | |
|---|---|---|---|---|
| V. natans | Biomass | 134.48 | 2.00 | <0.001 |
| RGR | 28.76 | 2.00 | <0.001 | |
| Numbers | 88.94 | 2.00 | <0.001 | |
| Height | 124.57 | 2.00 | <0.001 | |
| H. verticillata | Biomass | 256.56 | 2.00 | <0.001 |
| RGR | 31.93 | 2.00 | <0.001 | |
| Numbers | 657.42 | 2.00 | <0.001 | |
| Height | 224.94 | 2.00 | <0.001 | |
| Total | Total biomass | 505.88 | 2.00 | <0.001 |
| V. natans: H. verticillata | 35.21 | 2.00 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Q.; Han, Y.; Chen, K.; Huang, X.; Li, K.; He, H. Effects of Water Depth on the Growth of the Submerged Macrophytes Vallisneria natans and Hydrilla verticillata: Implications for Water Level Management. Water 2021, 13, 2590. https://doi.org/10.3390/w13182590
Li Q, Han Y, Chen K, Huang X, Li K, He H. Effects of Water Depth on the Growth of the Submerged Macrophytes Vallisneria natans and Hydrilla verticillata: Implications for Water Level Management. Water. 2021; 13(18):2590. https://doi.org/10.3390/w13182590
Chicago/Turabian StyleLi, Qisheng, Yanqing Han, Kunquan Chen, Xiaolong Huang, Kuanyi Li, and Hu He. 2021. "Effects of Water Depth on the Growth of the Submerged Macrophytes Vallisneria natans and Hydrilla verticillata: Implications for Water Level Management" Water 13, no. 18: 2590. https://doi.org/10.3390/w13182590