Nutrient Loading, Temperature and Heat Wave Effects on Nutrients, Oxygen and Metabolism in Shallow Lake Mesocosms Pre-Adapted for 11 Years

 , , , , ,
, , , , , 
Abstract
:1. Introduction
2. Materials and Methods
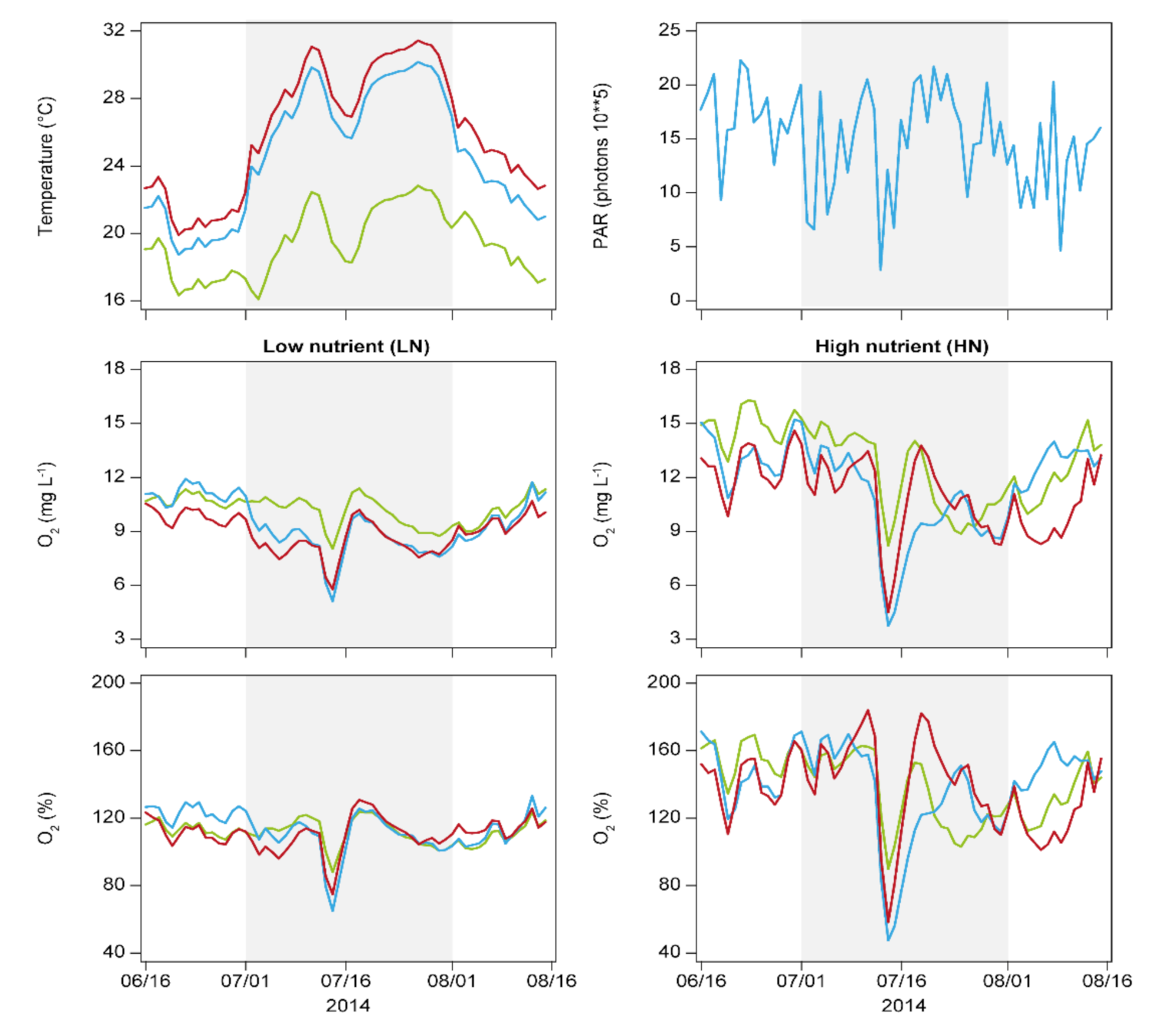
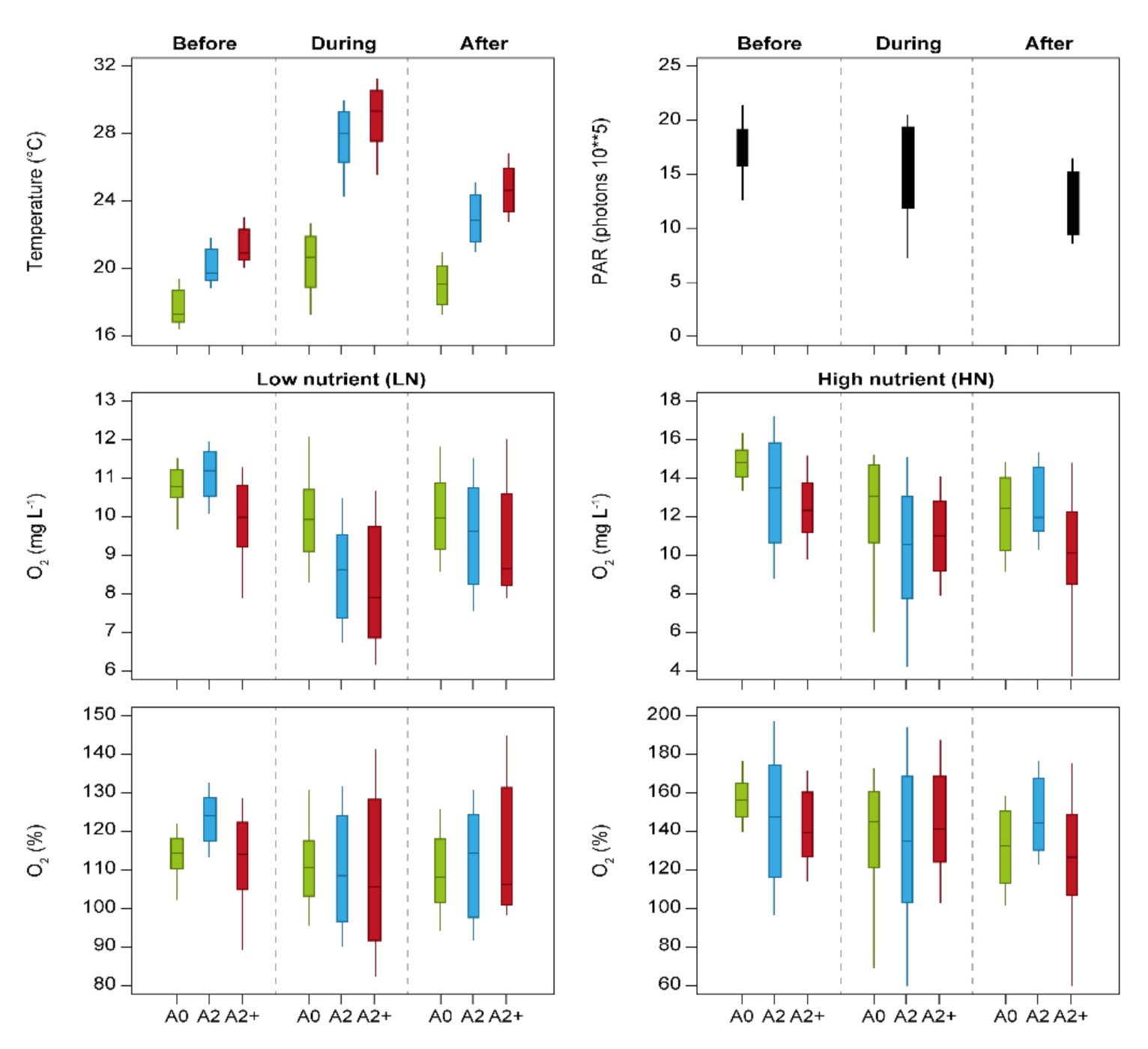
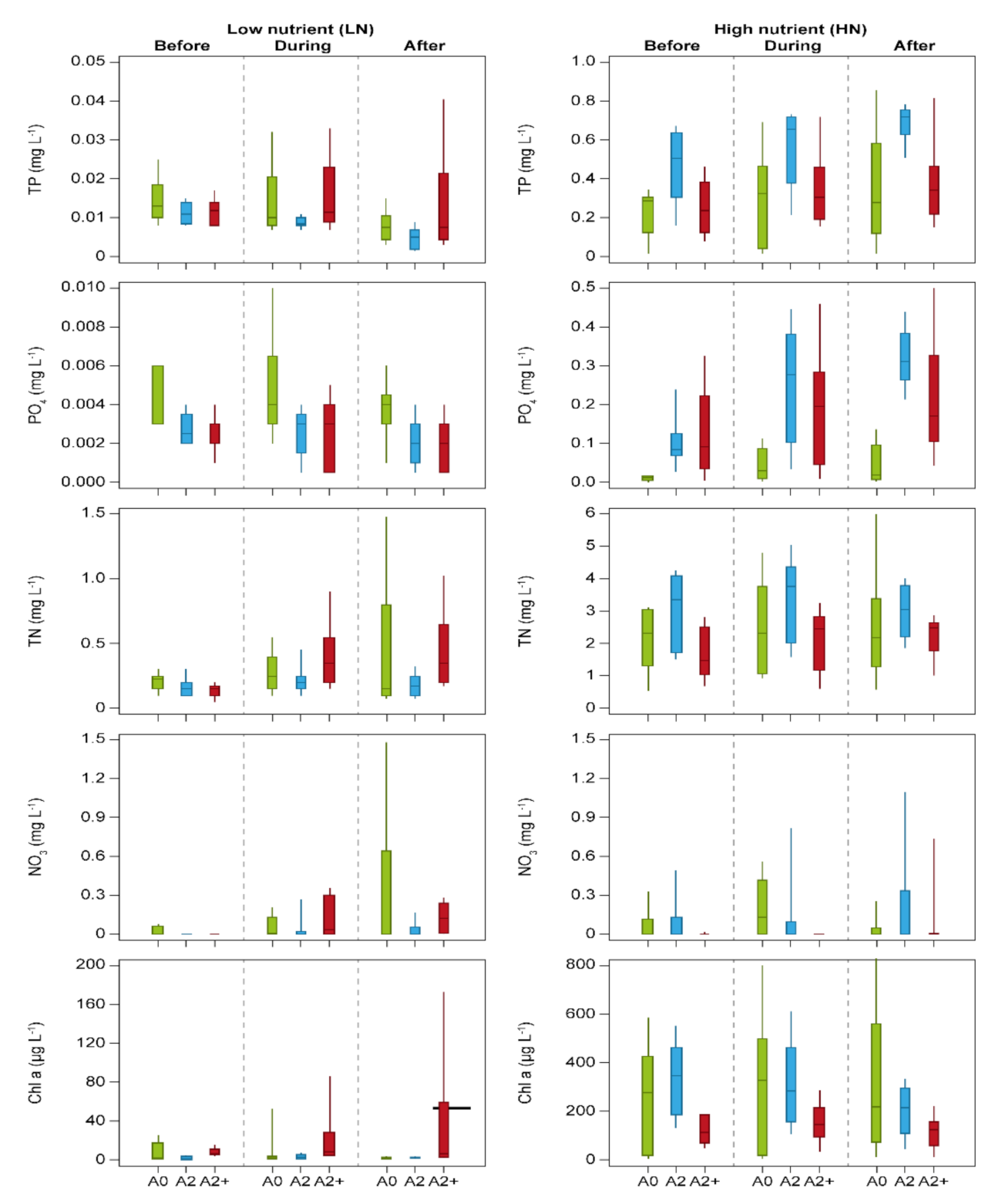
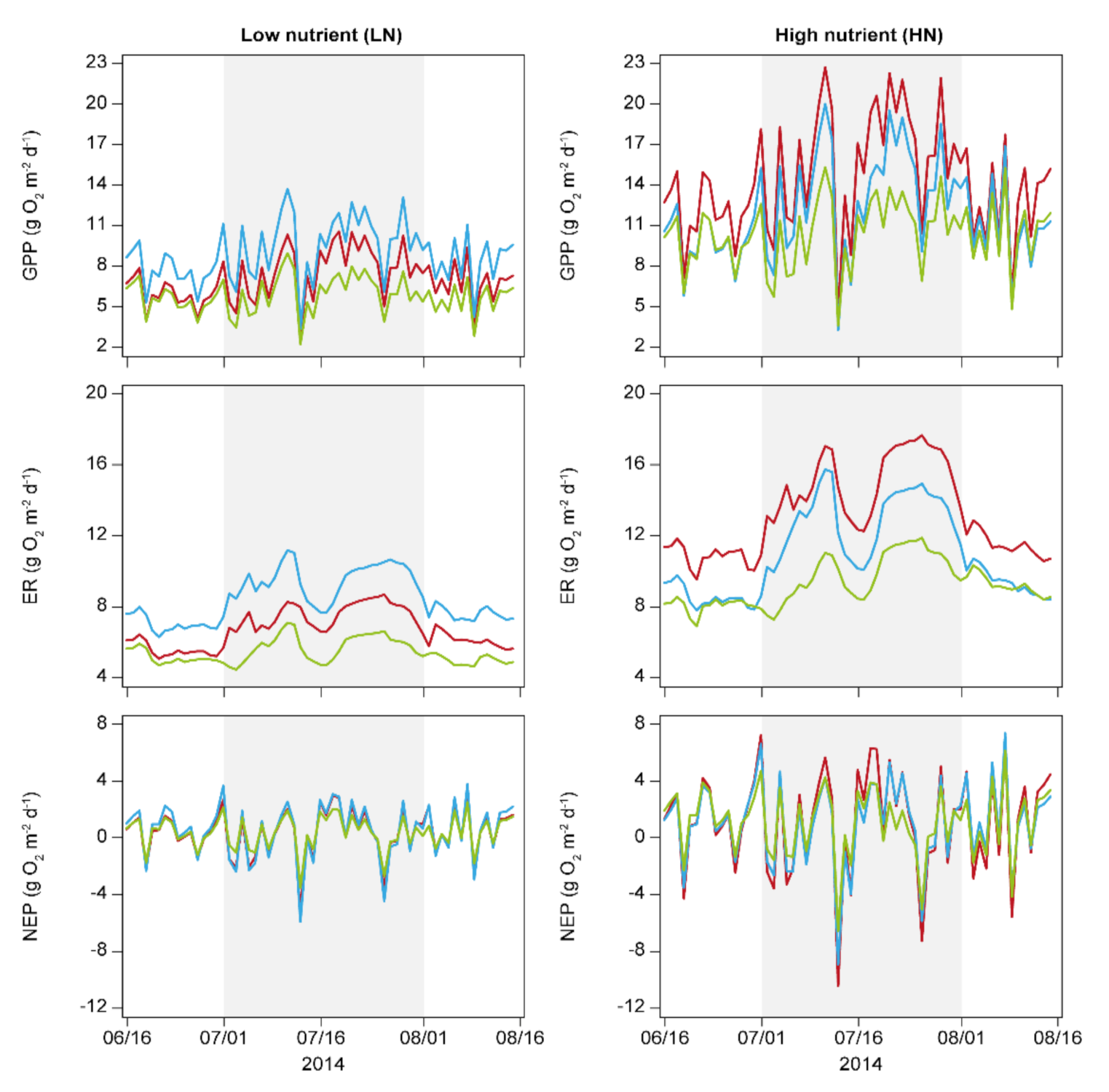
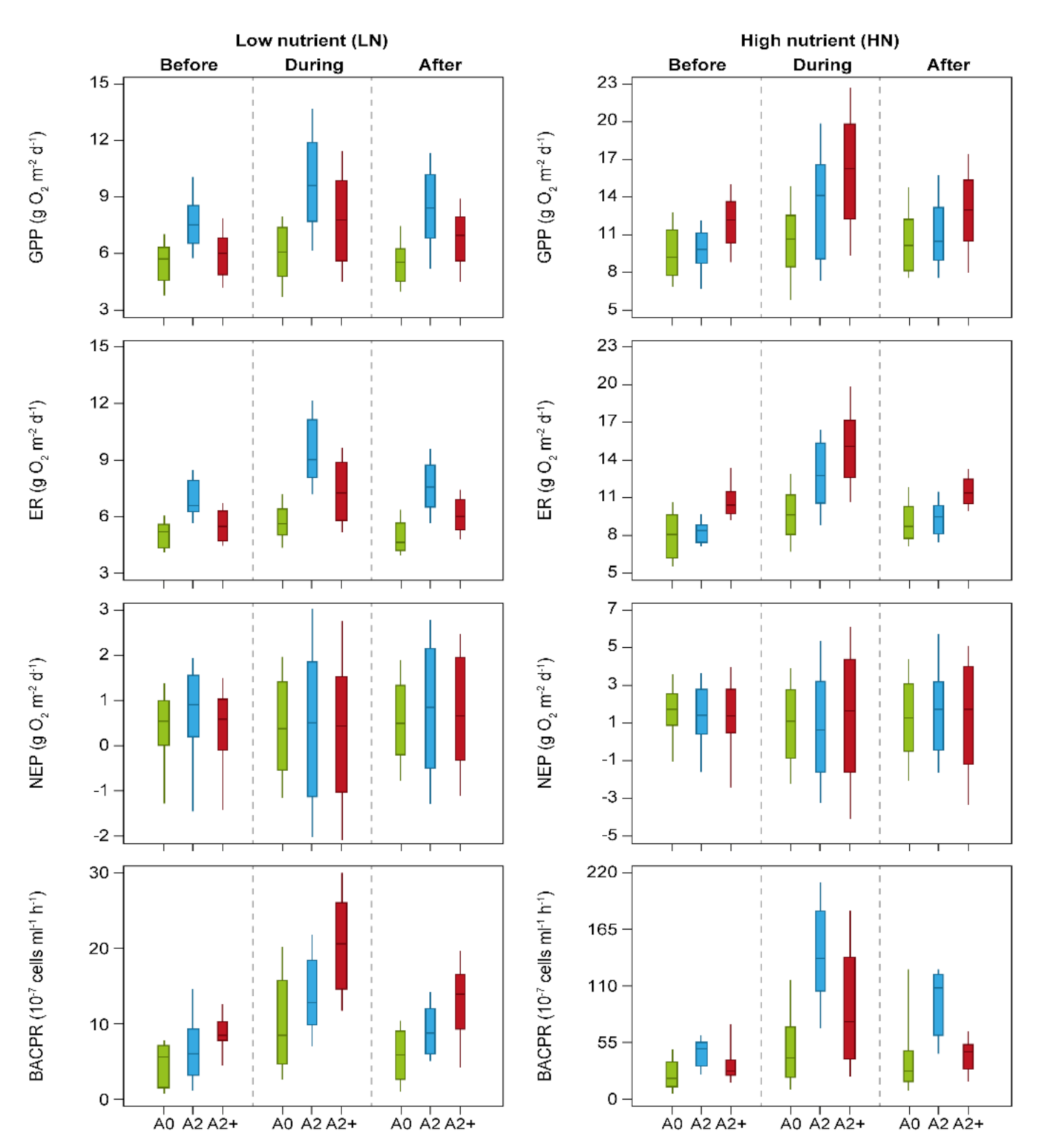
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- IPCC. Climate Change 2007: Synthesis Report, Contribution of Working Groups I, II and III to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Core Writing Team, Pachauri, R.K., Reisinger, A., Eds.; IPCC: Geneva, Switzerland, 2007; 104p. [Google Scholar]
- Jeppesen, E.; Kronvang, B.; Meerhoff, M.; Søndergaard, M.; Hansen, K.M.; Andersen, H.E.; Lauridsen, T.L.; Liboriussen, L.; Beklioglu, M.; Özen, A.; et al. Climate Change Effects on Runoff, Catchment Phosphorus Loading and Lake Ecological State, and Potential Adaptations. J. Environ. Qual. 2009, 38, 1930. [Google Scholar] [CrossRef] [PubMed]
- Jeppesen, E.; Meerhoff, M.; Davidson, T.A.; Trolle, D.; Søndergaard, M.; Lauridsen, T.L.; Beklioǧlu, M.; Brucet, S.; Volta, P.; González-Bergonzoni, I.; et al. Climate Change Impacts on Lakes: An Integrated Ecological Perspective Based on a Multi-Faceted Approach, with Special Focus on Shallow Lakes. J. Limnol. 2014, 73, 88–111. [Google Scholar] [CrossRef] [Green Version]
- Moss, B. Cogs in the Endless Machine: Lakes, Climate Change and Nutrient Cycles: A Review. Sci. Total Environ. 2012, 434, 130–142. [Google Scholar] [CrossRef] [PubMed]
- Kundzewicz, Z.W.; Mata, L.J.; Arnell, N.W.; DÖLL, P.; Jimenez, B.; Miller, K.; Oki, T.; ŞEN, Z.; Shiklomanov, I. The Implications of Projected Climate Change for Freshwater Resources and Their Management. Hydrol. Sci. J. 2008, 53, 3–10. [Google Scholar] [CrossRef]
- Jeppesen, E.; Nõges, P.; Davidson, T.A.; Haberman, J.; Nõges, T.; Blank, K.; Lauridsen, T.L.; Søndergaard, M.; Sayer, C.; Laugaste, R.; et al. Zooplankton as Indicators in Lakes: A Scientific-Based Plea for Including Zooplankton in the Ecological Quality Assessment of Lakes According to the European Water Framework Directive (WFD). Hydrobiologia 2011, 676, 279–297. [Google Scholar] [CrossRef]
- Kosten, S.; Roland, F.; Da Motta Marques, D.M.L.; Van Nes, E.H.; Mazzeo, N.; Sternberg, L.D.S.L.; Scheffer, M.; Cole, J.J. Climate-Dependent CO2 Emissions from Lakes. Glob. Biogeochem. Cycles 2010, 24, GB2007. [Google Scholar] [CrossRef]
- Moss, B. Climate Change, Nutrient Pollution and the Bargain of Dr Faustus. Freshw. Biol. 2010, 55, 175–187. [Google Scholar] [CrossRef]
- Yvon-Durocher, G.; Jones, J.I.; Trimmer, M.; Woodward, G.; Montoya, J.M. Warming Alters the Metabolic Balance of Ecosystems. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 2117–2126. [Google Scholar] [CrossRef] [Green Version]
- Scharfenberger, U.; Jeppesen, E.; Beklioğlu, M.; Søndergaard, M.; Angeler, D.G.; Çakıroğlu, A.İ.; Drakare, S.; Hejzlar, J.; Mahdy, A.; Papastergiadou, E.; et al. Effects of Trophic Status, Water Level, and Temperature on Shallow Lake Metabolism and Metabolic Balance: A Standardized Pan-European Mesocosm Experiment. Limnol. Oceanogr. 2019, 64, 616–631. [Google Scholar] [CrossRef]
- Staehr, P.A.; Sand-Jensen, K.A.J. Seasonal Changes in Temperature and Nutrient Control of Photosynthesis, Respiration and Growth of Natural Phytoplankton Communities. Freshw. Biol. 2006, 51, 249–262. [Google Scholar] [CrossRef]
- Solomon, C.T.; Bruesewitz, D.A.; Richardson, D.C.; Rose, K.C.; Van de Bogert, M.C.; Hanson, P.C.; Kratz, T.K.; Larget, B.; Adrian, R.; Leroux Babin, B.; et al. Ecosystem Respiration: Drivers of Daily Variability and Background Respiration in Lakes around the Globe. Limnol. Oceanogr. 2013, 58, 849–866. [Google Scholar] [CrossRef] [Green Version]
- del Giorgio, P.A.; Peters, R.H. Patterns in Planktonic P:R Ratios in Lakes: Influence of Lake Trophy and Dissolved Organic Carbon. Limnol. Oceanogr. 1994, 39, 772–787. [Google Scholar] [CrossRef]
- Hanson, P.C.; Bade, D.L.; Carpenter, S.R.; Kratz, T.K. Lake Metabolism: Relationships with Dissolved Organic Carbon and Phosphorus. Limnol. Oceanogr. 2003, 48, 1112–1119. [Google Scholar] [CrossRef] [Green Version]
- Anderson-Teixeira, K.J.; Vitousek, P.M. Ecosystems. In Metabolic Ecology; Sibly, R.M., Brown, J.H., Kodric-Brown, A., Eds.; Wiley-Blackwell: London, UK, 2012; pp. 99–111. [Google Scholar]
- Welter, J.R.; Benstead, J.P.; Cross, W.F.; Hood, J.M.; Huryn, A.D.; Johnson, P.W.; Williamson, T.J. Does N2 Fixation Amplify the Temperature Dependence of Ecosystem Metabolism? Ecology 2015, 96, 603–610. [Google Scholar] [CrossRef] [PubMed]
- Cole, J.; Findlay, S.; Pace, M. Bacterial Production in Fresh and Saltwater Ecosystems: A Cross-System Overview. Mar. Ecol. Prog. Ser. 1988, 43, 1–10. [Google Scholar] [CrossRef]
- Jeppesen, E.; Erlandsen, M.; Søndergaard, M. Can Simple Empirical Equations Describe the Seasonal Dynamics of Bacterioplankton in Lakes: An Eight-Year Study in Shallow Hypertrophic and Biologically Highly Dynamic Lake Sobygard, Denmark. Microb. Ecol. 1997, 34, 11–26. [Google Scholar] [CrossRef]
- Rae, R.; Vincent, W.F. Effects of Temperature and Ultraviolet Radiation on Microbial Foodweb Structure: Potential Responses to Global Change. Freshw. Biol. 1998, 40, 747–758. [Google Scholar] [CrossRef]
- Savage, C.; Leavitt, P.R.; Elmgren, R. Distribution and Retention of Effluent Nitrogen in Surface Sediments of a Coastal Bay. Limnol. Oceanogr. 2004, 49, 1503–1511. [Google Scholar] [CrossRef]
- Christoffersen, K.; Andersen, N.; Søndergaard, M.; Liboriussen, L.; Jeppesen, E. Implications of Climate-Enforced Temperature Increases on Freshwater Pico- and Nanoplankton Populations Studied in Artificial Ponds during 16 Months. Hydrobiologia 2006, 560, 259–266. [Google Scholar] [CrossRef]
- White, P.A.; Kalff, J.; Rasmussen, J.B.; Gasol, J.M. The Effect of Temperature and Algal Biomass on Bacterial Production and Specific Growth Rate in Freshwater and Marine Habitats. Microb. Ecol. 1991, 21, 99–118. [Google Scholar] [CrossRef]
- Meehl, G.A.; Tebaldi, C. More Intense, More Frequent, and Longer Lasting Heat Waves in the 21st Century. Science 2004, 305, 994–997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- IPCC. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2014. [Google Scholar]
- Christidis, N.; Jones, G.S.; Stott, P.A. Dramatically Increasing Chance of Extremely Hot Summers since the 2003 European Heatwave. Nat. Clim. Chang. 2015, 5, 46–50. [Google Scholar] [CrossRef]
- Mooij, W.M.; De Senerpont Domis, L.N.; Hülsmann, S. The Impact of Climate Warming on Water Temperature, Timing of Hatching and Young-of-the-Year Growth of Fish in Shallow Lakes in the Netherlands. J. Sea Res. 2008, 60, 32–43. [Google Scholar] [CrossRef]
- Kragh, T.; Martinsen, K.T.; Kristensen, E.; Sand-Jensen, K. From Drought to Flood: Sudden Carbon Inflow Causes Whole-Lake Anoxia and Massive Fish Kill in a Large Shallow Lake. Sci. Total Environ. 2020, 739, 140072. [Google Scholar] [CrossRef]
- Jeppesen, E.; Søndergaard, M.; Sortkjoær, O.; Mortensen, E.; Kristensen, P. Interactions between Phytoplankton, Zooplankton and Fish in a Shallow, Hypertrophic Lake: A Study of Phytoplankton Collapses in Lake Søbygård, Denmark. Hydrobiologia 1990, 191, 149–164. [Google Scholar] [CrossRef]
- Liboriussen, L.; Landkildehus, F.; Meerhoff, M.; Bramm, M.E.; Søndergaard, M.; Christoffersen, K.; Richardson, K.; Søndergaard, M.; Lauridsen, T.L.; Jeppesen, E. Global Warming: Design of a Flow-through Shallow Lake Mesocosm Climate Experiment. Limnol. Oceanogr. Methods 2005, 3, 1–9. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2001: The Scientific Basis, Contribution of Working Group I to the Third Assessment Report of the Intergovernmental Panel on Climate Change; Houghton, J.T., Ding, Y., Griggs, D.J., Noguer, M., van der Linden, P.J., Dai, X., Maskell, K., Johnson, C.A., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2001. [Google Scholar] [CrossRef]
- Scheffer, M.; Hosper, S.H.; Meijer, M.L.; Moss, B.; Jeppesen, E. Alternative Equilibria in Shallow Lakes. Trends Ecol. Evol. 1993, 8, 275–279. [Google Scholar] [CrossRef]
- Jespersen, A.; Christoffersen, K. Measurements of Chlorophyll―A from Phytoplankton Using Ethanol as Extraction Solvent. Arch. Hydrobiol. 1987, 109, 445–454. [Google Scholar]
- Søndergaard, M.; Jeppesen, E.; Jensen, J.P.; Amsinck, S.L. Water Framework Directive: Ecological Classification of Danish Lakes. J. Appl. Ecol. 2005, 42, 616–629. [Google Scholar] [CrossRef]
- Šorf, M.; Davidson, T.A.; Brucet, S.; Menezes, R.F.; Søndergaard, M.; Lauridsen, T.L.; Landkildehus, F.; Liboriussen, L.; Jeppesen, E. Zooplankton Response to Climate Warming: A Mesocosm Experiment at Contrasting Temperatures and Nutrient Levels. Hydrobiologia 2015, 742, 185–203. [Google Scholar] [CrossRef]
- Işkın, U.; Filiz, N.; Cao, Y.; Neif, É.M.; Öğlü, B.; Lauridsen, T.L.; Davidson, T.A.; Søndergaard, M.; Tavşanoğlu, Ü.N.; Beklioǧlu, M.; et al. Impact of Nutrients, Temperature and a Heat Wave on Zooplankton Community Structure: An Experimental Approach. Water 2020, 12, 3416. [Google Scholar] [CrossRef]
- Liboriussen, L.; Lauridsen, T.L.; Søndergaard, M.; Landkildehus, F.; Søndergaard, M.; Larsen, S.E.; Jeppesen, E. Effects of Warming and Nutrients on Sediment Community Respiration in Shallow Lakes: An Outdoor Mesocosm Experiment. Freshw. Biol. 2011, 56, 437–447. [Google Scholar] [CrossRef]
- Jähne, B.; Münnich, K.O.; Bösinger, R.; Dutzi, A.; Huber, W.; Libner, P. On the Parameters Influencing Air-Water Gas Exchange. J. Geophys. Res. Ocean. 1987, 92, 1937–1949. [Google Scholar] [CrossRef]
- Cole, J.J.; Caraco, N.F. Atmospheric Exchange of Carbon Dioxide in a Low-Wind Oligotrophic Lake Measured by the Addition of SF6. Limnol. Oceanogr. 1998, 43, 647–656. [Google Scholar] [CrossRef] [Green Version]
- Erlandsen, M.; Thyssen, N. Modelling the Community Oxygen Production in Lowland Streams Dominated by Submerged Macrophytes. In Developments in Environmental Modelling; Elsevier: Amsterdam, The Netherlands, 1983. [Google Scholar] [CrossRef]
- Kelly, M.G.; Thyssen, N.; Moeslund, B. Light and the Annual Variation of Oxygen- and Carbon-based Measurements of Productivity in a Macrophyte-dominated River. Limnol. Oceanogr. 1983, 28, 503–515. [Google Scholar] [CrossRef]
- Jeppesen, E.; Søndergaard, M.; Lauridsen, T.L.; Davidson, T.A.; Liu, Z.; Mazzeo, N.; Trochine, C.; Özkan, K.; Jensen, H.S.; Trolle, D.; et al. Biomanipulation as a Restoration Tool to Combat Eutrophication. Recent Advances and Future Challenges. In Advances in Ecological Research; Academic Press: Cambridge, MA, USA, 2012. [Google Scholar] [CrossRef]
- McGraw-Hill. Water and Wastewater Calculations Manual; McGraw-Hill: New York, NY, USA, 2007. [Google Scholar]
- Fuhrman, J.A.; Azam, F. Thymidine Incorporation as a Measure of Heterotrophic Bacterioplankton Production in Marine Surface Waters: Evaluation and Field Results. Mar. Biol. 1982, 66, 109–120. [Google Scholar] [CrossRef]
- Riemann, B.; Sondergaard, M.; Schierup, H.H.; Bosselmann, S.; Christensen, G.; Hansen, J.; Nielsen, B. Carbon Metabolism during a Spring Diatom Bloom in the Eutrophic Lake Mossø. Int. Rev. Gesamten Hydrobiol. Hydrogr. 1982, 67, 145–185. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G.M. Statistics for Biology and Health, Mixed Effects Models and Extensions in Ecology with R. 2009. Available online: https://www.amazon.com/Effects-Extensions-Ecology-Statistics-Biology/dp/1441927646 (accessed on 29 December 2020).
- Pinheiro, J.C.; Bates, D.M. Mixed-Effects Models in S and S-Plus: Statistics and Computing. 2000. Available online: https://www.springer.com/gp/book/9780387989570 (accessed on 29 December 2020).
- Audet, J.; Neif, É.M.; Cao, Y.; Hoffmann, C.C.; Lauridsen, T.L.; Larsen, S.E.; Søndergaard, M.; Jeppesen, E.; Davidson, T.A. Heat-Wave Effects on Greenhouse Gas Emissions from Shallow Lake Mesocosms. Freshw. Biol. 2017, 62, 1130–1142. [Google Scholar] [CrossRef]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Core Team. Nlme: Linear and Nonlinear Mixed Effects Models. R Package Version 3.1–117. 2014. Available online: http//cran.r-project.org/web/packages/nlme/index.html (accessed on 29 December 2020).
- R Core Team. R: A Language and Environment for Statistical Computing. R Found. Stat. Comput. Vienna, Austria. 2014. Available online: http//www.R-project.org/ (accessed on 29 December 2020).
- Søndergaard, M.; Lauridsen, T.L.; Johansson, L.S.; Jeppesen, E. Nitrogen or Phosphorus Limitation in Lakes and Its Impact on Phytoplankton Biomass and Submerged Macrophyte Cover. Hydrobiologia 2017, 795, 35–48. [Google Scholar] [CrossRef]
- Jensen, H.S.; Andersen, F.O. Importance of Temperature, Nitrate, and PH for Phosphate Release from Aerobic Sediments of Four Shallow, Eutrophic Lakes. Limnol. Oceanogr. 1992, 37, 577–589. [Google Scholar] [CrossRef]
- Søndergaard, M.; Bjerring, R.; Jeppesen, E. Persistent Internal Phosphorus Loading during Summer in Shallow Eutrophic Lakes. Hydrobiologia 2013, 710, 95–107. [Google Scholar] [CrossRef]
- Moss, B. Engineering and Biological Approaches to the Restoration from Eutrophication of Shallow Lakes in Which Aquatic Plant Communities Are Important Components. Hydrobiologia 1990, 200–201, 367–377. [Google Scholar] [CrossRef]
- Brothers, S.M.; Hilt, S.; Attermeyer, K.; Grossart, H.P.; Kosten, S.; Lischke, B.; Mehner, T.; Meyer, N.; Scharnweber, K.; Kohler, J. A Regime Shift from Macrophyte to Phytoplankton Dominance Enhances Carbon Burial in a Shallow, Eutrophic Lake. Ecosphere 2013, 4, 1–17. [Google Scholar] [CrossRef]
- Filiz, N.; Işkın, U.; Beklioǧlu, M.; Öğlü, B.; Cao, Y.; Davidson, T.A.; Søndergaard, M.; Lauridsen, T.L.; Jeppesen, E. Phytoplankton Community Response to Nutrients, Temperature, and a Heat Wave in Shallow Lakes: An Experimental Approach. Water 2020, 12, 3394. [Google Scholar] [CrossRef]
- Zingel, P.; Cremona, F.; Nõges, T.; Cao, Y.; Neif, É.M.; Coppens, J.; Işkın, U.; Lauridsen, T.L.; Davidson, T.A.; Søndergaard, M.; et al. Effects of Warming and Nutrients on the Microbial Food Web in Shallow Lake Mesocosms. Eur. J. Protistol. 2018, 64, 1–12. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation | Definition | Abbreviation | Definition |
|---|---|---|---|
| A0 | Ambient treatments | HN | High nutrient treatments |
| A2 | Ambient + 2–4 °C treatments | IPCC | Int. Panel on Climate Change |
| A2+ | Ambient + 3–6° C treatments | LN | Low nutrient treatments |
| BACPR | Bacterioplankton Production | NEP | Net Ecosystem Production |
| Chl-a | Chlorophyll-a | NEP20 | NEP standardized to 20 °C |
| DO | Dissolved Oxygen | NO3 | Nitrite+nitrate |
| DO% | DO percent saturation | PO4 | Orthophosphate |
| ER | Ecosystem Respiration | O2 | Oxygen concentration |
| ER20 | ER standardized to 20 °C | O2% | Oxygen percent saturation |
| GPP | Gross Primary Production | PAR | Photosynthetic Active Radiation |
| GPP20 | GPP standardized to 20 °C | PVI | Plant Volume Inhabited |
| HW | Heat wave | TCA | Trichloroacetic acid |
| HW0 | pre- and post- HW period | TEMP | Temperature in the water |
| HW1 | HW period | TP | Total phosphorus |
| HW2 | Ambient summer period | TN | Total nitrogen |
| Data Set | Treatment | Chl-a | TP | PO4 | TN | NO3 | O2 | O2% |
|---|---|---|---|---|---|---|---|---|
| Full | Nutrient | *** (+) | ***(+) | *** (+) | *** (+) | ns | *** (+) | *** (+) |
| Temperature | ns | ns | ns | ns | ns | ns | ns | |
| Heat wave | * (+)0 | ns | *** (+)0 | ** (+)0 | ns | ns | ns | |
| Low Nutrient | Temperature | * (+) | ns | * (-) | ns | ns | ns | ns |
| Heat wave | *** (+)0 | ns | ns | ns | ns | ** (+)0 | ns | |
| High Nutrient | Temperature | ns | ns | ** (+) | ns | ns | ns | ns |
| Heat wave | ns | ns | ns | ns | ns | ns | ns |
| Data Set | Treatment | GPP | ER | NEP | BACPR |
|---|---|---|---|---|---|
| Full | Nutrient | *** (+) | *** (+) | * (+) | *** (+) |
| Temperature | ns | * (+) | ns | ** (+) | |
| Heat wave | *** (+)1 | *** (+)1 | *** (−)0 | *** (+)0 | |
| LN | Temperature | * (+) | *** (+) | ns | ** (+) |
| Heat wave | *** (+)1 | *** (+)1 | ns | *** (+)0 | |
| HN | Temperature | * (+) | * (+) | ns | * (+) |
| Heat wave | *** (+)0 | *** (+)0 | ns | *** (+)0 |
| Data Set | With Chl-a TP and TN |
|---|---|
| Full | Log(GPP20) = 2.50(0.04) + 0.022(0.009) Log (Chl-a), p < 0.0001, n = 337 Log(ER20) = 2.40(0.04) + 0.019(0.005) Log (Chl-a) − 0.026(0.007) Log(TN), p < 0.0001, n = 337 Log(NEP20) = 2.43(0.01) + 0.031(0.009) Log(TN), p < 0.02, n = 337 |
| LN | Log(GPP20) = 2.25(0.07) + 0.029(0.013) Log (TN) − 0.044(0.013) Log(TP), p < 0.0001, n = 168 Log(ER20) = 2.28(0.03) − 0.033(0.008) Log (TN), p < 0.0001, n = 168 Log(NEP20) = 2.27(0.07) + 0.055(0.013) Log(TN) − 0.041(0.013) Log(TP), p < 0.0003, n = 168 |
| HN | Log(ER20) = 2.40(0.06) + 0.027(0.009) Log (Chl-a) − 0.058(0.016) Log (TP), p < 0.0001, n = 169 GPP20 and NEP20 were not significantly related to these variables. |
| With PVI, Chl-a TP and TN, and PVI Must Be Included | |
| Full and HN | GPP20, ER20 and NEP20 were not significantly related to PVI. |
| LN | Log(GPP20) = 2.23(0.03) + 0.011(0.02) Log (PVI + 1), p < 0.02, n = 83 Log(ER20) = 2.21(0.04) − 0.04(0.01) Log (TN) + 0.05(0.02) Log(PVI + 1), p < 0.0001, n = 83 NEP20 was not significantly related to PVI. |
| Data Set | With Chl-a TP and TN |
|---|---|
| Full | Log(BACPR) = −2.62(0.35) + 0.14(0.01) Log (Chl-a) + 1.03(0.11) Log (TEMP), p < 0.0001, n = 328 |
| LN | Log(BACPR) =−4.97(0.80) − 0.16(0.04) Log (Chl-a) − 0.15(0.06) Log (TP) + 1.51(0.22) Log (TEMP), p < 0.0001, n = 160 |
| HN | Log(BACPR) = −0.66(0.19) − 0.024(0.011) Log(Chl-a) + 0.13(0.02) Log (TP) + 0.66(0.05) Log (TEMP), p < 0.0001, n = 168 |
| With PVI, Chl-a TP, TN, and PVI Must Be Included | |
| No significant relationships |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeppesen, E.; Audet, J.; Davidson, T.A.; Neif, É.M.; Cao, Y.; Filiz, N.; Lauridsen, T.L.; Larsen, S.E.; Beklioğlu, M.; Sh, T.; et al. Nutrient Loading, Temperature and Heat Wave Effects on Nutrients, Oxygen and Metabolism in Shallow Lake Mesocosms Pre-Adapted for 11 Years. Water 2021, 13, 127. https://doi.org/10.3390/w13020127
Jeppesen E, Audet J, Davidson TA, Neif ÉM, Cao Y, Filiz N, Lauridsen TL, Larsen SE, Beklioğlu M, Sh T, et al. Nutrient Loading, Temperature and Heat Wave Effects on Nutrients, Oxygen and Metabolism in Shallow Lake Mesocosms Pre-Adapted for 11 Years. Water. 2021; 13(2):127. https://doi.org/10.3390/w13020127
Chicago/Turabian StyleJeppesen, Erik, Joachim Audet, Thomas A. Davidson, Érika M. Neif, Yu Cao, Nur Filiz, Torben L. Lauridsen, Søren E. Larsen, Meryem Beklioğlu, Tserenpil Sh, and et al. 2021. "Nutrient Loading, Temperature and Heat Wave Effects on Nutrients, Oxygen and Metabolism in Shallow Lake Mesocosms Pre-Adapted for 11 Years" Water 13, no. 2: 127. https://doi.org/10.3390/w13020127