
Distribution Patterns of Benthic Foraminifera in Fish Farming Areas (Corsica, France): Implications for the Implementation of Biotic Indices in Biomonitoring Studies

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling Strategy
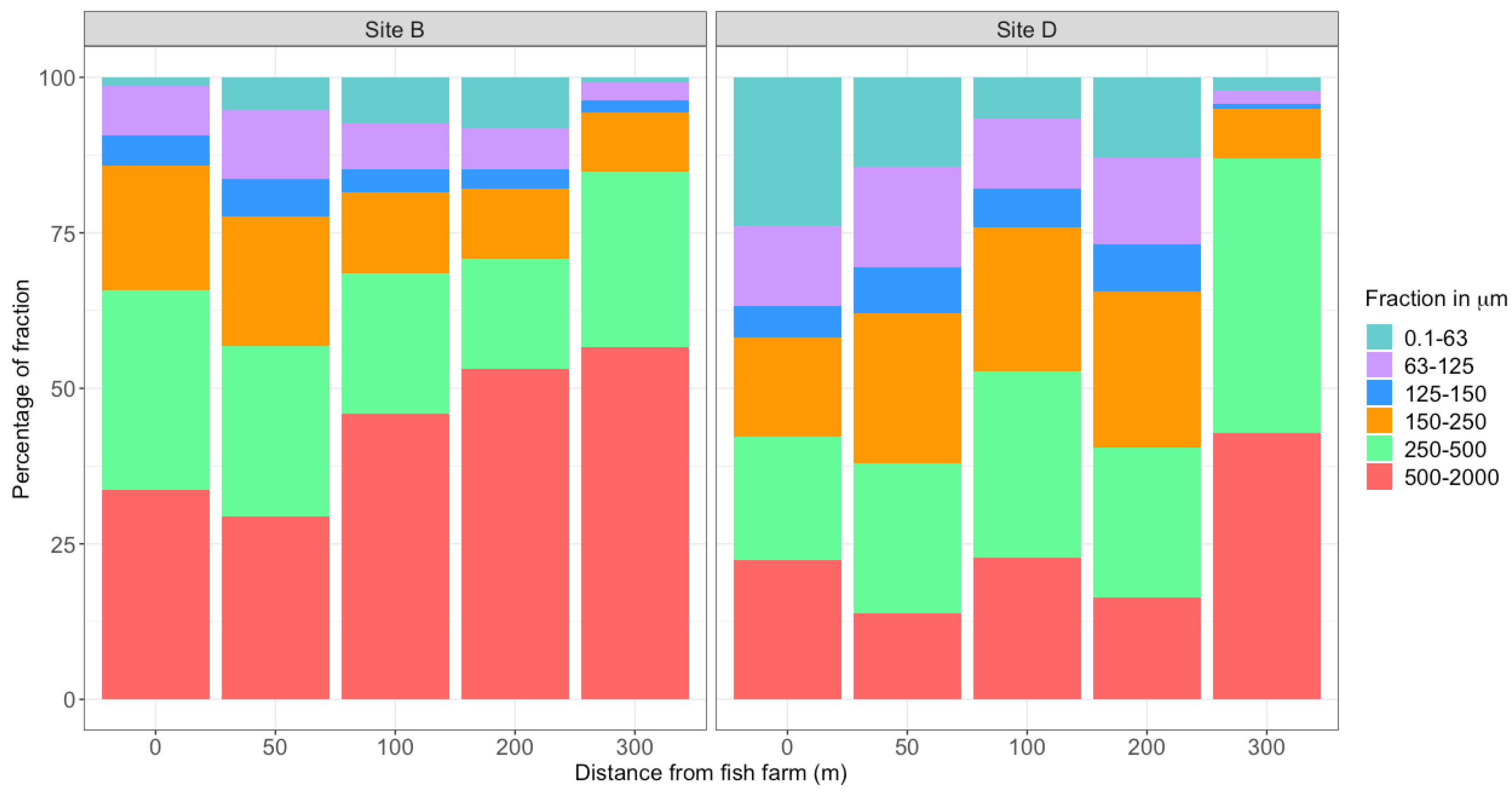
2.2. Grain Size Analyses
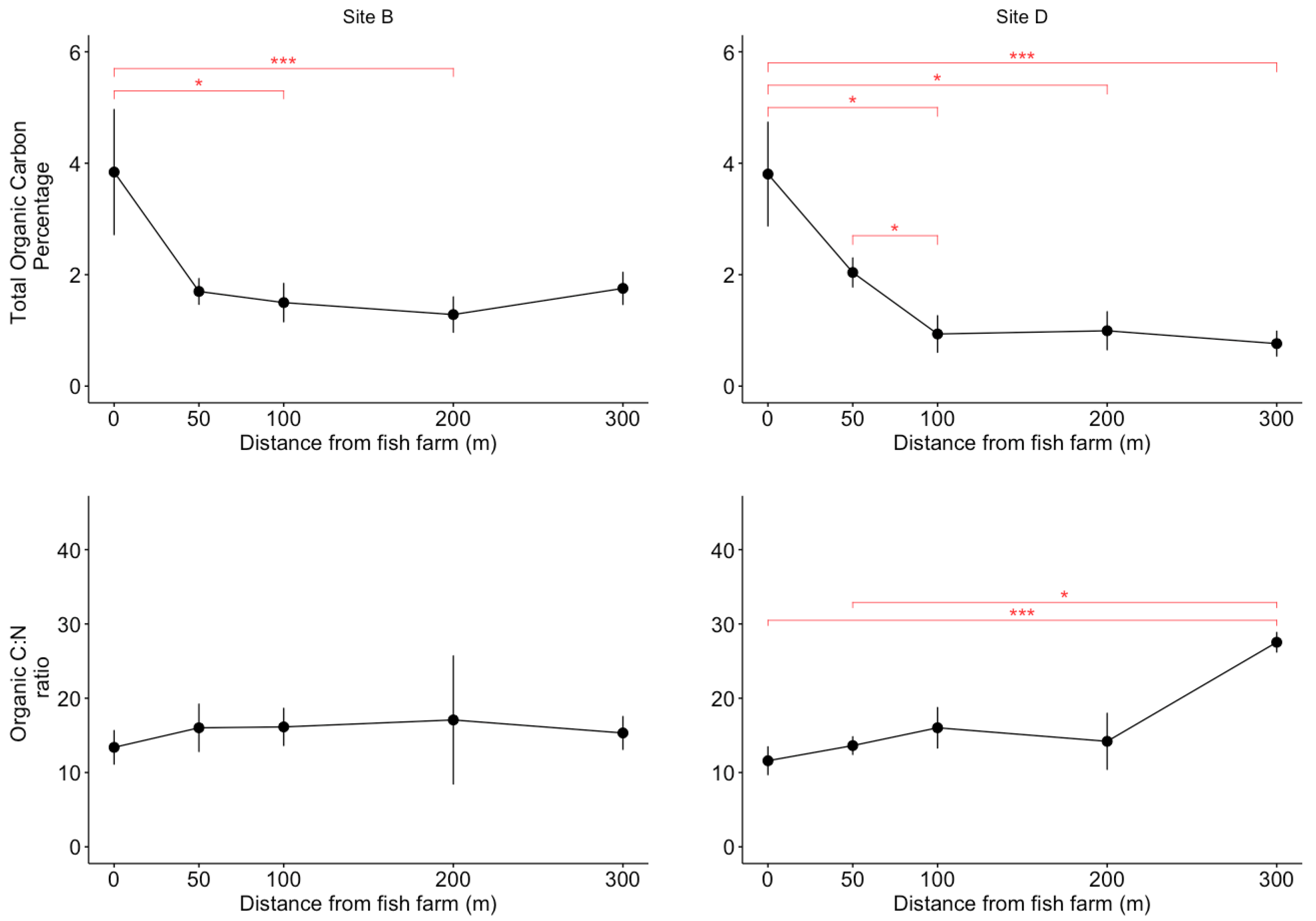
2.3. Organic Matter Analysis
2.4. Foraminiferal Analysis
2.5. Faunal Analyses
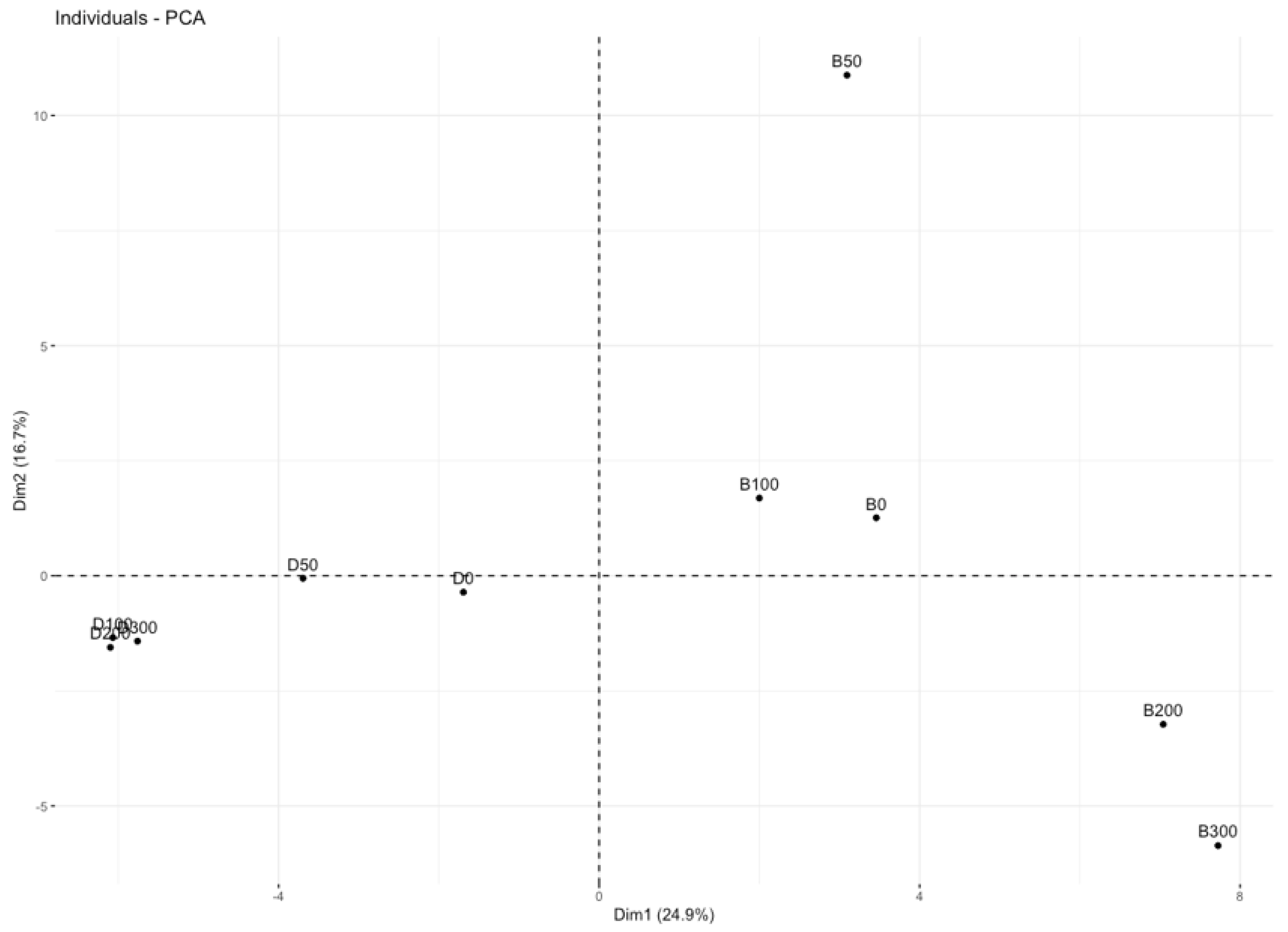
2.6. Numerical Analysis
3. Results
3.1. Environmental Parameters Analayses
3.2. Foraminiferal Analyses
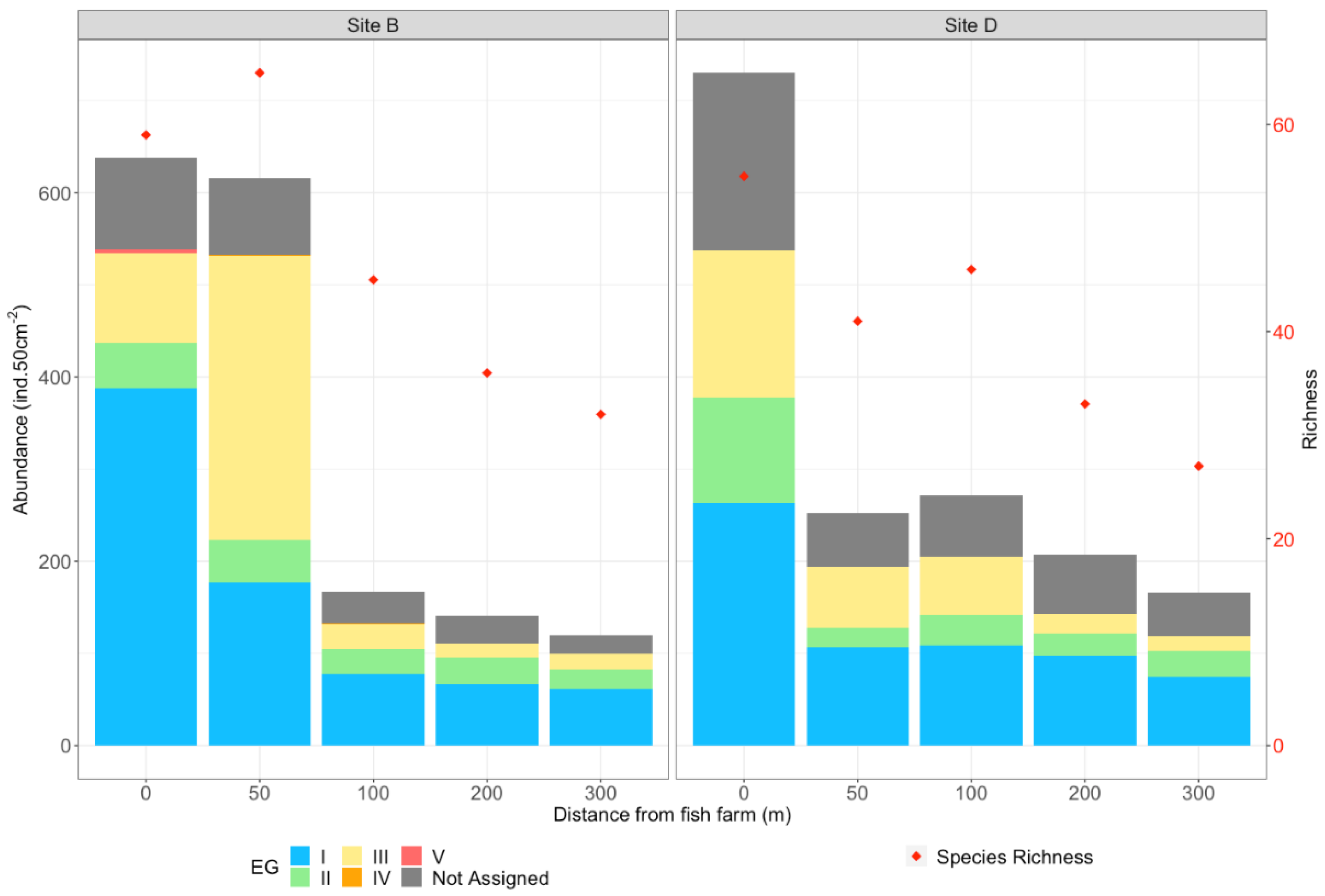
3.2.1. Abundance and Diversity
3.2.2. Faunal Composition
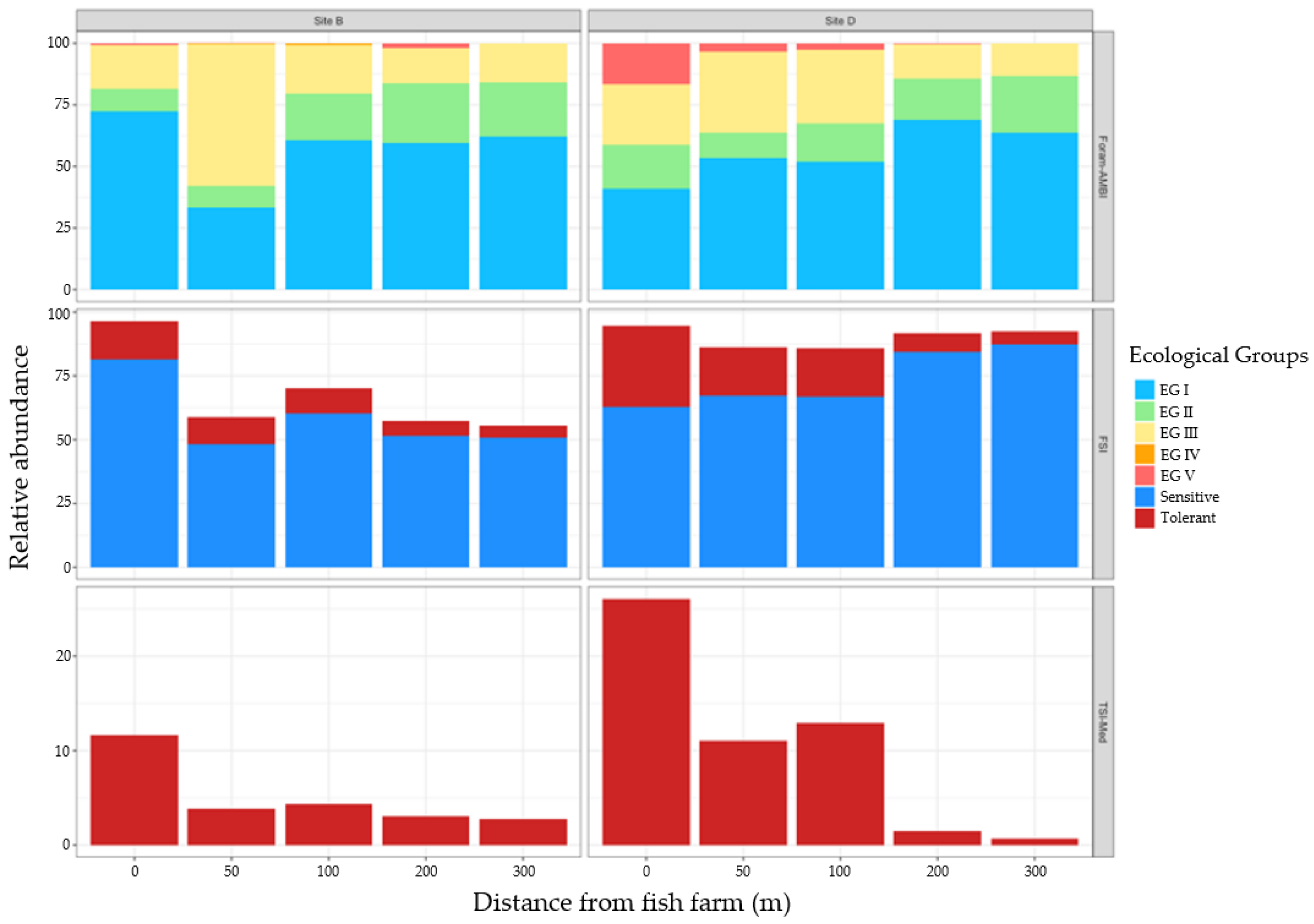
3.2.3. Proportions of Ecological Groups
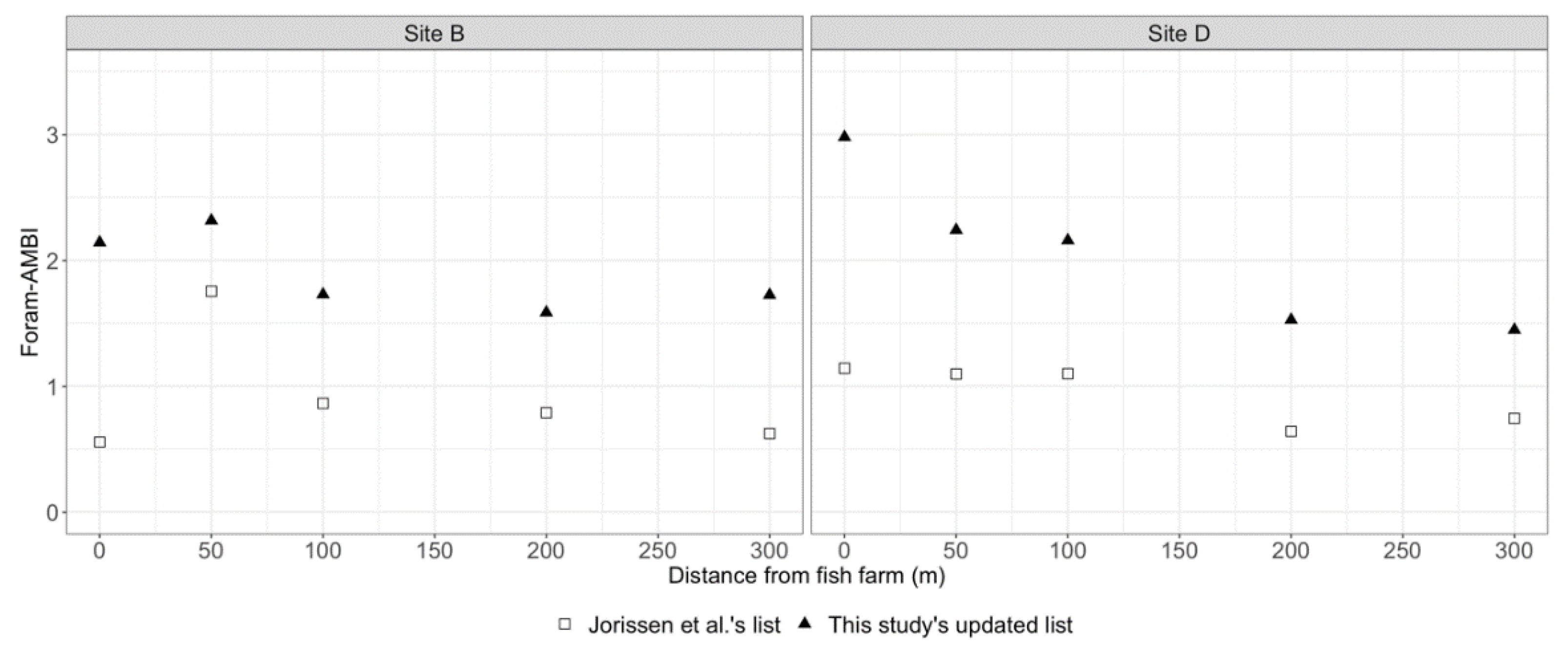
3.2.4. Biotic Indices and Environment Parameters
4. Discussion
4.1. A Limited Effect of Fish Farming on Habitat Features
4.2. Diversity-Based Indices May Not Be Adapted to Coastal Waters in the Mediterranean Sea
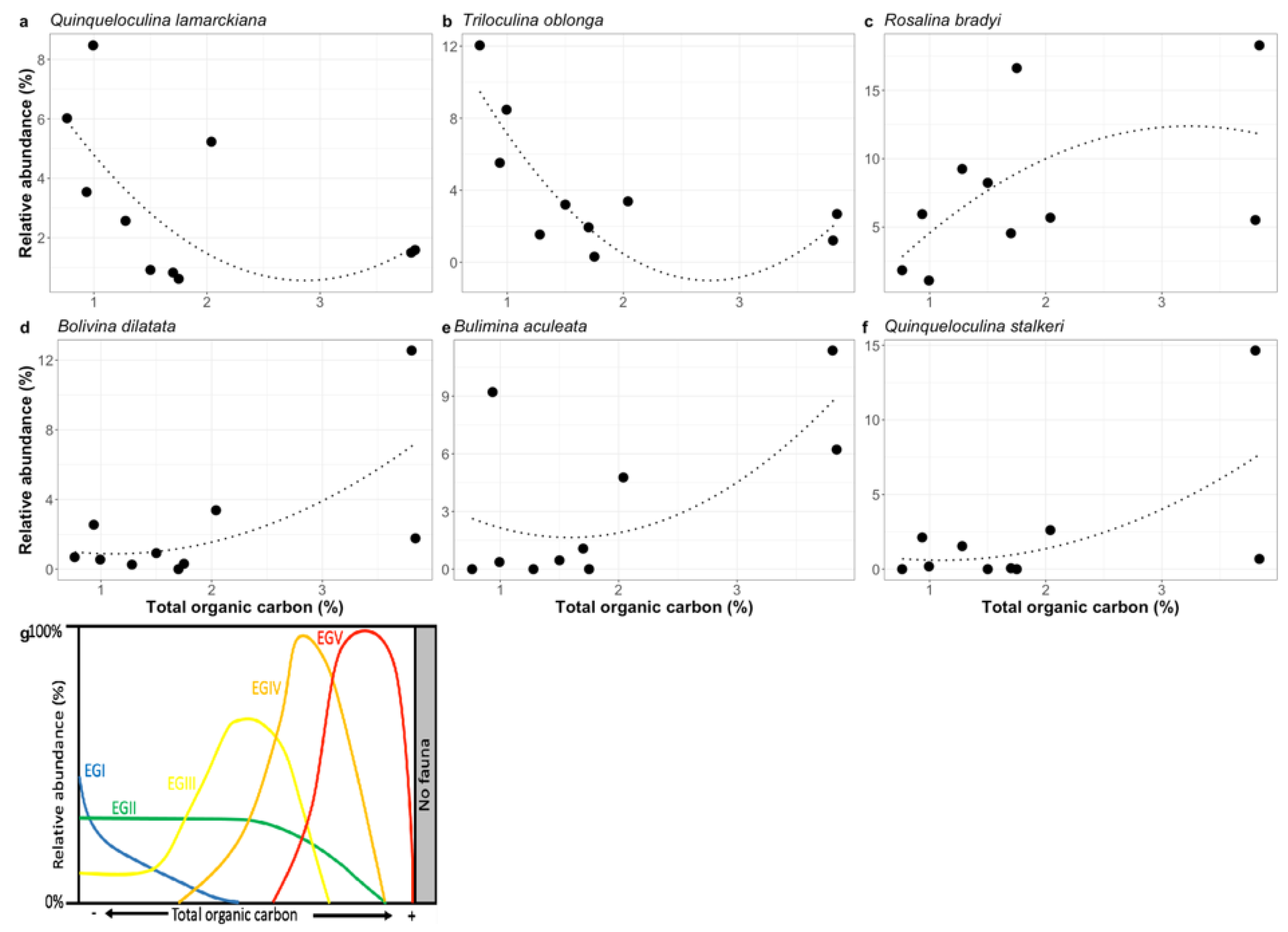
4.3. Species Response to Fish Farming Cages: Improving Foram-AMBI Original Species List Assignment to Ecological Group, the Foram-AMBI Updated
4.4. Comparison of Sensitivity-Based Indices’ Efficiency in Monitoring the Impact of Fish Farming
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations. The State of the World Fisheries and Aquaculture 2014: Opportunities and Challenges; Food and Agriculture Organization of the United Nations: Rome, Italy, 2014. [Google Scholar]
- Food and Agriculture Organization of the United Nations. The State of World Fisheries and Aquaculture 2016: Contributing to Food Security and Nutrition for All; Food and Agriculture Organization of the United Nations: Rome, Italy, 2016. [Google Scholar]
- Food and Agriculture Organization of the United Nations. National Aquaculture Sector Overview—France; Food and Agriculture Organization of the United Nations: Rome, Italy, 2012. [Google Scholar]
- Lensi, P. La Politique de Diversification de l’aquaculture En Corse. La Démarche “Nouvelles Espèces”. Aspects économiques de la production aquacole. Cah. Opt. Médit 1995, 14, 137–148. [Google Scholar]
- Yokoyama, H.; Abo, K.; Ishihi, Y. Quantifying Aquaculture-Derived Organic Matter in the Sediment in and around a Coastal Fish Farm Using Stable Carbon and Nitrogen Isotope Ratios. Aquaculture 2006, 254, 411–425. [Google Scholar] [CrossRef]
- Chamberlain, J. Impacts of Biodeposits from Suspended Mussel (Mytilus Edulis L.) Culture on the Surrounding Surficial Sediments. ICES J. Mar. Sci. 2001, 58, 411–416. [Google Scholar] [CrossRef] [Green Version]
- Dauvin, J.-C.; Pezy, J.-P.; Baffreau, A.; Bachelet, Q.; Baux, N.; Méar, Y.; Murat, A.; Poizot, E. Effects of a Salmon Fish Farm on Benthic Habitats in a High-Energy Hydrodynamic System: The Case of the Rade de Cherbourg (English Channel). Aquaculture 2020, 518, 734832. [Google Scholar] [CrossRef]
- Holmer, M.; Wildish, D.; Hargrave, B. Organic Enrichment from Marine Finfish Aquaculture and Effects on Sediment Biogeochemical Processes. In Environmental Effects of Marine Finfish Aquaculture; Handbook of Environmental Chemistry; Hargrave, B.T., Ed.; Springer: Berlin/Heidelberg, Germany, 2005; Volume 5M, pp. 181–206. [Google Scholar] [CrossRef]
- Johannessen, P.J.; Botnen, H.B.; Tvedten, O.F. Macrobenthos: Before, during and after a Fish Farm. Aquac. Res. 1994, 25, 55–66. [Google Scholar] [CrossRef]
- Pearson, T.H.; Rosenberg, R. Macrobenthic Succession in Relation to Organic Enrichment and Pollution of the Marine Environment. Oceanogr. Mar. Biol. 1978, 16, 229–311. [Google Scholar]
- Angel, D.L. Impact of a Net Cage Fish Farm on the Distribution of Benthic Foraminifera in the Northern Gulf of Eilat (Aqaba, Red Sea). J. Foraminifer. Res. 2000, 30, 54–65. [Google Scholar] [CrossRef]
- Apostolaki, E.T.; Tsagaraki, T.; Tsapakis, M.; Karakassis, I. Fish Farming Impact on Sediments and Macrofauna Associated with Seagrass Meadows in the Mediterranean. Estuar. Coast. Shelf Sci. 2007, 75, 408–416. [Google Scholar] [CrossRef]
- Delgado, O.; Ruiz, J.; Pérez, M.; Romero, J.; Ballesteros, E. Effects of Fish Farming on Seagrass (Posidonia Oceanica) in a Mediterranean Bay: Seagrass Decline after Organic Loading Cessation. Oceanol. Acta 1999, 22, 109–117. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, J.M. Effects of Fish Farm Loadings on Seagrass (Posidonia Oceanica) Distribution, Growth and Photosynthesis. Mar. Pollut. Bull. 2001, 42, 12. [Google Scholar] [CrossRef]
- Karakassis, I.; Tsapakis, M.; Smith, C.J.; Rumohr, H. Fish Farming Impacts in the Mediterranean Studied through Sediment Profiling Imagery. Mar. Ecol. Prog. Ser. 2002, 227, 125–133. [Google Scholar] [CrossRef]
- La Rosa, T.; Mirto, S.; Mazzola, A.; Maugeri, T.L. Benthic Microbial Indicators of Fish Farm Impact in a Coastal Area of the Tyrrhenian Sea. Aquaculture 2004, 230, 153–167. [Google Scholar] [CrossRef]
- Mazzola, A. Fish-Farming Effects on Benthic Community Structure in Coastal Sediments: Analysis of Meiofaunal Recovery. ICES J. Mar. Sci. 2000, 57, 1454–1461. [Google Scholar] [CrossRef] [Green Version]
- Muxika, I.; Borja, A.; Bonne, W. The Suitability of the Marine Biotic Index (AMBI) to New Impact Sources along European Coasts. Ecol. Indic. 2005, 5, 19–31. [Google Scholar] [CrossRef]
- Neofitou, N.; Vafidis, D.; Klaoudatos, S. Spatial and Temporal Effects of Fish Farming on Benthic Community Structure in a Semi-Enclosed Gulf of the Eastern Mediterranean. Aquacult. Environ. Interact. 2010, 1, 95–105. [Google Scholar] [CrossRef] [Green Version]
- Krause-Jensen, D.; Greve, T.M.; Nielsen, K. Eelgrass as a Bioindicator under the European Water Framework Directive. Water Resour. Manag. 2005, 19, 63–75. [Google Scholar] [CrossRef]
- Ar Gall, E.; Le Duff, M.; Sauriau, P.-G.; de Casamajor, M.-N.; Gevaert, F.; Poisson, E.; Hacquebart, P.; Joncourt, Y.; Barillé, A.-L.; Buchet, R.; et al. Implementation of a New Index to Assess Intertidal Seaweed Communities as Bioindicators for the European Water Framework Directory. Ecol. Indic. 2016, 60, 162–173. [Google Scholar] [CrossRef] [Green Version]
- Borja, A.; Franco, J.; Pérez, V. A Marine Biotic Index to Establish the Ecological Quality of Soft-Bottom Benthos within European Estuarine and Coastal Environments. Mar. Pollut. Bull. 2000, 40, 1100–1114. [Google Scholar] [CrossRef]
- Alve, E. Benthic Foraminiferal Responses to Estuarine Pollution; a Review. J. Foraminifer. Res. 1995, 25, 190–203. [Google Scholar] [CrossRef]
- Francescangeli, F.; Quijada, M.; Armynot du Châtelet, E.; Frontalini, F.; Trentesaux, A.; Billon, G.; Bouchet, V.M.P. Multidisciplinary Study to Monitor Consequences of Pollution on Intertidal Benthic Ecosystems (Hauts de France, English Channel, France): Comparison with Natural Areas. Mar. Environ. Res. 2020, 160, 105034. [Google Scholar] [CrossRef]
- Frontalini, F.; Coccioni, R. Benthic Foraminifera as Bioindicators of Pollution: A Review of Italian Research over the Last Three Decades. Rev. Micropaléontol. 2011, 54, 115–127. [Google Scholar] [CrossRef]
- Martins, V.A.; Frontalini, F.; Tramonte, K.M.; Figueira, R.C.L.; Miranda, P.; Sequeira, C.; Fernández-Fernández, S.; Dias, J.A.; Yamashita, C.; Renó, R.; et al. Assessment of the Health Quality of Ria de Aveiro (Portugal): Heavy Metals and Benthic Foraminifera. Mar. Pollut. Bull. 2013, 70, 18–33. [Google Scholar] [CrossRef]
- Martins, V.A.; Zaaboub, N.; Aleya, L.; Frontalini, F.; Pereira, E.; Miranda, P.; Mane, M.; Rocha, F.; Laut, L.; El Bour, M. Environmental Quality Assessment of Bizerte Lagoon (Tunisia) Using Living Foraminifera Assemblages and a Multiproxy Approach. PLoS ONE 2015, 10, e0137250. [Google Scholar] [CrossRef]
- Mojtahid, M.; Jorissen, F.; Durrieu, J.; Galgani, F.; Howa, H.; Redois, F.; Camps, R. Benthic Foraminifera as Bio-Indicators of Drill Cutting Disposal in Tropical East Atlantic Outer Shelf Environments. Mar. Micropaleontol. 2006, 61, 58–75. [Google Scholar] [CrossRef] [Green Version]
- Mojtahid, M.; Jorissen, F.; Pearson, T.H. Comparison of Benthic Foraminiferal and Macrofaunal Responses to Organic Pollution in the Firth of Clyde (Scotland). Mar. Pollut. Bull. 2008, 56, 42–76. [Google Scholar] [CrossRef] [Green Version]
- Morvan, M.; Le Cadre, V.; Jorissen, F.; Debenay, J.-P. Foraminifera as Potential Bio- Indicators of the “Erika” Oil Spill in the Bay of Bourgneuf: Field and Experimental Studies. Aquat. Living Resour. 2004, 17, 317–322. [Google Scholar] [CrossRef]
- Parent, B.; Hyams-Kaphzan, O.; Barras, C.; Lubinevsky, H.; Jorissen, F. Testing Foraminiferal Environmental Quality Indices along a Well-Defined Organic Matter Gradient in the Eastern Mediterranean. Ecol. Indic. 2021, 125, 107498. [Google Scholar] [CrossRef]
- Alve, E.; Lepland, A.; Magnusson, J.; Backer-Owe, K. Monitoring Strategies for Re-Establishment of Ecological Reference Conditions: Possibilities and Limitations. Mar. Pollut. Bull. 2009, 59, 297–310. [Google Scholar] [CrossRef] [PubMed]
- Dolven, J.K.; Alve, E.; Rygg, B.; Magnusson, J. Defining Past Ecological Status and In Situ Reference Conditions Using Benthic Foraminifera: A Case Study from the Oslofjord, Norway. Ecol. Indic. 2013, 29, 219–233. [Google Scholar] [CrossRef]
- Polovodova Asteman, I.; Hanslik, D.; Nordberg, K. An Almost Completed Pollution-Recovery Cycle Reflected by Sediment Geochemistry and Benthic Foraminiferal Assemblages in a Swedish–Norwegian Skagerrak Fjord. Mar. Pollut. Bull. 2015, 95, 126–140. [Google Scholar] [CrossRef] [PubMed]
- Schönfeld, J.; Alve, E.; Geslin, E.; Jorissen, F.; Korsun, S.; Spezzaferri, S. The FOBIMO (FOraminiferal BIo-MOnitoring) Initiative—Towards a Standardised Protocol for Soft-Bottom Benthic Foraminiferal Monitoring Studies. Mar. Micropaleontol. 2012, 94–95, 1–13. [Google Scholar] [CrossRef]
- Bouchet, V.M.P.; Deldicq, N.; Baux, N.; Dauvin, J.-C.; Pezy, J.-P.; Seuront, L.; Méar, Y. Benthic Foraminifera to Assess Ecological Quality Statuses: The Case of Salmon Fish Farming. Ecol. Indic. 2020, 117, 106607. [Google Scholar] [CrossRef]
- Damak, M.; Fourati, R.; Elleuch, B.; Kallel, M. Environmental Quality Assessment of the Fish Farms’ Impact in the Monastir Bay (Eastern of Tunisia, Central Mediterranean): A Benthic Foraminiferal Perspective. Environ. Sci. Pollut. Res. 2020, 27, 9059–9074. [Google Scholar] [CrossRef]
- Schafer, C.T.; Winters, G.V.; Scott, D.B.; Pocklington, P.; Cole, F.E.; Honig, C. Survey of Living Foraminifera and Polychaete Populations at Some Canadian Aquaculture Sites; Potential for Impact Mapping and Monitoring. J. Foraminifer. Res. 1995, 25, 236–259. [Google Scholar] [CrossRef]
- Vidović, J.; Dolenec, M.; Dolenec, T.; Karamarko, V.; Žvab Rožič, P. Benthic Foraminifera Assemblages as Elemental Pollution Bioindicator in Marine Sediments around Fish Farm (Vrgada Island, Central Adriatic, Croatia). Mar. Pollut. Bull. 2014, 83, 198–213. [Google Scholar] [CrossRef] [PubMed]
- Oron, S.; Angel, D.; Goodman-Tchernov, B.; Merkado, G.; Kiflawi, M.; Abramovich, S. Benthic Foraminiferal Response to the Removal of Aquaculture Fish Cages in the Gulf of Aqaba-Eilat, Red Sea. Mar. Micropaleontol. 2014, 107, 8–17. [Google Scholar] [CrossRef]
- Bouchet, V.M.P.; Frontalini, F.; Francescangeli, F.; Sauriau, P.-G.; Geslin, E.; Martins, M.V.A.; Almogi-Labin, A.; Avnaim-Katav, S.; Di Bella, L.; Cearreta, A.; et al. Indicative Value of Benthic Foraminifera for Biomonitoring: Assignment to Ecological Groups of Sensitivity to Total Organic Carbon of Species from European Intertidal Areas and Transitional Waters. Mar. Pollut. Bull. 2021, 164, 112071. [Google Scholar] [CrossRef]
- Pawlowski, J.; Esling, P.; Lejzerowicz, F.; Cedhagen, T.; Wilding, T.A. Environmental Monitoring through Protist Next-Generation Sequencing Metabarcoding: Assessing the Impact of Fish Farming on Benthic Foraminifera Communities. Mol. Ecol. Resour. 2014, 14, 1129–1140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pochon, X.; Wood, S.A.; Keeley, N.B.; Lejzerowicz, F.; Esling, P.; Drew, J.; Pawlowski, J. Accurate Assessment of the Impact of Salmon Farming on Benthic Sediment Enrichment Using Foraminiferal Metabarcoding. Mar. Pollut. Bull. 2015, 100, 370–382. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, P.; Polovodova Asteman, I.; Bouchet, V.M.P. Benthic Foraminiferal Indices and Environmental Quality Assessment of Transitional Waters: A Review of Current Challenges and Future Research Perspectives. Water 2021, 13, 1898. [Google Scholar] [CrossRef]
- Bouchet, V.M.P.; Alve, E.; Rygg, B.; Telford, R.J. Benthic Foraminifera Provide a Promising Tool for Ecological Quality Assessment of Marine Waters. Ecol. Indic. 2012, 23, 66–75. [Google Scholar] [CrossRef]
- Alve, E.; Korsun, S.; Schönfeld, J.; Dijkstra, N.; Golikova, E.; Hess, S.; Husum, K.; Panieri, G. Foram-AMBI: A Sensitivity Index Based on Benthic Foraminiferal Faunas from North-East Atlantic and Arctic Fjords, Continental Shelves and Slopes. Mar. Micropaleontol. 2016, 122, 1–12. [Google Scholar] [CrossRef]
- Jorissen, F.; Nardelli, M.P.; Almogi-Labin, A.; Barras, C.; Bergamin, L.; Bicchi, E.; El Kateb, A.; Ferraro, L.; McGann, M.; Morigi, C.; et al. Developing Foram-AMBI for Biomonitoring in the Mediterranean: Species Assignments to Ecological Categories. Mar. Micropaleontol. 2018, 140, 33–45. [Google Scholar] [CrossRef] [Green Version]
- Barras, C.; Jorissen, F.J.; Labrune, C.; Andral, B.; Boissery, P. Live Benthic Foraminiferal Faunas from the French Mediterranean Coast: Towards a New Biotic Index of Environmental Quality. Ecol. Indic. 2014, 36, 719–743. [Google Scholar] [CrossRef] [Green Version]
- Dimiza, M.D.; Triantaphyllou, M.V.; Koukousioura, O.; Hallock, P.; Simboura, N.; Karageorgis, A.P.; Papathanasiou, E. The Foram Stress Index: A New Tool for Environmental Assessment of Soft-Bottom Environments Using Benthic Foraminifera. A Case Study from the Saronikos Gulf, Greece, Eastern Mediterranean. Ecol. Indic. 2016, 60, 611–621. [Google Scholar] [CrossRef]
- Bouchet, V.M.P.; Goberville, E.; Frontalini, F. Benthic Foraminifera to Assess Ecological Quality Statuses in Italian Transitional Waters. Ecol. Indic. 2018, 84, 130–139. [Google Scholar] [CrossRef]
- Cavaliere, M.; Barrenechea Angeles, I.; Montresor, M.; Bucci, C.; Brocani, L.; Balassi, E.; Margiotta, F.; Francescangeli, F.; Bouchet, V.M.P.; Pawlowski, J.; et al. Assessing the Ecological Quality Status of the Highly Polluted Bagnoli Area (Tyrrhenian Sea, Italy) Using Foraminiferal EDNA Metabarcoding. Sci. Total Environ. 2021, 790, 147871. [Google Scholar] [CrossRef] [PubMed]
- El Kateb, A.; Stalder, C.; Martínez-Colón, M.; Mateu-Vicens, G.; Francescangeli, F.; Coletti, G.; Stainbank, S.; Spezzaferri, S. Foraminiferal-Based Biotic Indices to Assess the Ecological Quality Status of the Gulf of Gabes (Tunisia): Present Limitations and Future Perspectives. Ecol. Indic. 2020, 111, 105962. [Google Scholar] [CrossRef]
- Fossile, E.; Sabbatini, A.; Spagnoli, F.; Caridi, F.; Dell’Anno, A.; De Marco, R.; Dinelli, E.; Droghini, E.; Tramontana, M.; Negri, A. Sensitivity of Foraminiferal-Based Indices to Evaluate the Ecological Quality Status of Marine Coastal Benthic Systems: A Case Study of the Gulf of Manfredonia (Southern Adriatic Sea). Mar. Pollut. Bull. 2021, 163, 111933. [Google Scholar] [CrossRef]
- Melis, R.; Celio, M.; Bouchet, V.M.P.; Varagona, G.; Bazzaro, M.; Crosera, M.; Pugliese, N. Seasonal Response of Benthic Foraminifera to Anthropogenic Pressure in Two Stations of the Gulf of Trieste (Northern Adriatic Sea, Italy): The Marine Protected Area of Miramare versus the Servola Water Sewage Outfall. Mediterr. Mar. Sci. 2019, 20, 120–141. [Google Scholar] [CrossRef]
- Parent, B.; Barras, C.; Jorissen, F. An Optimised Method to Concentrate Living (Rose Bengal-Stained) Benthic Foraminifera from Sandy Sediments by High Density Liquids. Mar. Micropaleontol. 2018, 144, 1–13. [Google Scholar] [CrossRef]
- Cimerman, F.; Langer, M.R. Mediterranean Foraminifera; Dela/Slovenska Akademija Znanosti in Umetnosti, Razred za Naravoslovne Vede = Opera/Academia Scientiarum et Artium Slovenica, Classis IV—Historia Naturalis, Znanstvenoraziskovalni Center SAZU, Paleontološki inštitut Ivana Rakovca; Slovenska Akademija Znanosti in Umetnosti: Ljubljana, Slovenia, 1991. [Google Scholar]
- Jones, R.W.; Brady, H.B. The Challenger Foraminifera; Oxford University Press: Oxford, MS, USA, 1994. [Google Scholar]
- World Register of Marine Species. Available online: http://www.marinespecies.org (accessed on 7 October 2021).
- Chao, A.; Shen, T.-J. Non-Parametric Estimation of Shannon’s Index of Diversity When There Are Unseen Species in Sample. Environ. Ecol. Stat. 2003, 10, 429–443. [Google Scholar] [CrossRef]
- Beck, J.; Schwanghart, W. Comparing Measures of Species Diversity from Incomplete Inventories: An Update. Methods Ecol. Evol. 2010, 1, 38–44. [Google Scholar] [CrossRef]
- Hayek, L.-A.; Buzas, M.A. Surveying Natural Populations: Quantitative Tools for Assessing Biodiversity, 2nd ed.; Columbia University Press: New York, NY, USA, 1997. [Google Scholar]
- Jost, L. Entropy and Diversity. Oikos 2006, 113, 363–375. [Google Scholar] [CrossRef]
- Hill, M.O. Diversity and Evenness: A Unifying Notation and Its Consequences. Ecology 1973, 54, 427–432. [Google Scholar] [CrossRef] [Green Version]
- Sokal, R.F.; Rolf, S.W. Biometry: The Principles and Practice of Statistics in Biological Research; MacMillan: New York, NY, USA, 1995. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Compunting: Vienna, Austria, 2020. [Google Scholar]
- Souissi, R.; Turki, I.; Souissi, F. Effect of Submarine Morphology on Environment Quality: Case of Monastir Bay (Eastern Tunisia). Carpathian J. Earth Environ. Sci. 2014, 9, 231–239. [Google Scholar]
- Vizzini, S.; Sarà, G.; Michener, R.; Mazzola, A. The Role and Contribution of the Seagrass Posidonia oceanica (L.) Delile Organic Matter for Secondary Consumers as Revealed by Carbon and Nitrogen Stable Isotope Analysis. Acta Oeacologica 2002, 23, 277–285. [Google Scholar] [CrossRef]
- Ellis, A.M.; Culver, S.J.; Mallinson, D.J.; Corbett, D.R.; Leorri, E.; Buzas, M.A.; Shazili, N.A.M. The Influence of Aquaculture of Modern Foraminifera and Sediments in the Setiu Estuary and Lagoon, Terengganu, Malaysia: A Spatial Investigation. J. Foraminifer. Res. 2014, 44, 390–415. [Google Scholar] [CrossRef]
- Hyams-Kaphzan, O.; Almogi-Labin, A.; Benjamini, C.; Herut, B. Natural Oligotrophy vs. Pollution-Induced Eutrophy on the SE Mediterranean Shallow Shelf (Israel): Environmental Parameters and Benthic Foraminifera. Mar. Pollut. Bull. 2009, 58, 1888–1902. [Google Scholar] [CrossRef] [PubMed]
- Pearl, R.; Reed, L.J. On the Rate of Growth of the Population of the United States since 1790 and Its Mathematical Representation. Proc. Natl. Acad. Sci. USA 1920, 6, 275–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langer, M.R. Epiphytic Foraminifera. Mar. Micropaleontol. 1993, 20, 235–265. [Google Scholar] [CrossRef]
- Choi, Y.H.; Jeong, D.U.; Lee, Y.G.; Choi, J.U.; Kim, S.; Lee, J.S. Opportunistic Species and Seasonal Variation in Epiphytic Foraminiferal Assemblages in Abalone Farm Nets. J. Coast. Res. 2019, 35, 559–573. [Google Scholar] [CrossRef]
- El Kateb, A.; Stalder, C.; Neururer, C.; Fentimen, R.; Spangenberg, J.E.; Spezzaferri, S. Distribution of Benthic Foraminiferal Assemblages in the Transitional Environment of the Djerba Lagoon (Tunisia). Swiss J. Geosci. 2018, 111, 589–606. [Google Scholar] [CrossRef] [Green Version]
- Buosi, C.; Armynot du Châtelet, E.; Cherchi, A. Benthic Foraminiferal Assemblages in the Current-Dominated Strait of Bonifacio (Mediterranean Sea). J. Foraminifer. Res. 2012, 42, 39–55. [Google Scholar] [CrossRef]
- Mojtahid, M.; Jorissen, F.; Lansard, B.; Fontanier, C.; Bombled, B.; Rabouille, C. Spatial Distribution of Live Benthic Foraminifera in the Rhône Prodelta: Faunal Response to a Continental–Marine Organic Matter Gradient. Mar. Micropaleontol. 2009, 70, 177–200. [Google Scholar] [CrossRef] [Green Version]
- Donnici, S.; Barbero, R. The benthic foraminiferal communities of the northern Adriatic continental shelf. Mar. Micropaleontol. 2002, 44, 93–123. [Google Scholar] [CrossRef]
- De Rijk, S.; Jorissen, F.J.; Rohling, E.J.; Troelstra, S.R. Organic flux control on bathymetric zonation of Mediterranean benthic foraminifera. Mar. Micropaleontol. 2000, 40, 151–166. [Google Scholar] [CrossRef]
- Murray, J.W. Ecology and Paleoecology of Benthic Foraminifera; Longman Scientific & Technical: London, UK, 1991; 397p. [Google Scholar]
- Dauvin, J.-C.; Bellan, G.; Bellan-Santini, D. Benthic Indicators: From Subjectivity to Objectivity—Where Is the Line? Mar. Pollut. Bull. 2010, 60, 947–953. [Google Scholar] [CrossRef] [PubMed]
- Spilmont, N. The Future of Benthic Indicators: Moving up to the Intertidal. Open J. Mar. Sci. 2013, 3, 76–86. [Google Scholar] [CrossRef]
- Zettler, M.L.; Proffitt, C.E.; Darr, A.; Degraer, S.; Devriese, L.; Greathead, C.; Kotta, J.; Magni, P.; Martin, G.; Reiss, H.; et al. On the Myths of Indicator Species: Issues and Further Consideration in the Use of Static Concepts for Ecological Applications. PLoS ONE 2013, 8, e78219. [Google Scholar] [CrossRef] [Green Version]
- Pearson, T.H.; Black, K.D. The Environmental Impacts of Marine Fish Cage Culture. In Environmental Impacts of Aquaculture; Sheffield Academic Press: London, UK, 2001; pp. 1–31. [Google Scholar]
- Vidović, J.; Ćosović, V.; Juračić, M.; Petricioli, D. Impact of Fish Farming on Foraminiferal Community, Drvenik Veliki Island, Adriatic Sea, Croatia. Mar. Pollut. Bull. 2009, 58, 1297–1309. [Google Scholar] [CrossRef]
- Alve, E.; Hess, S.; Bouchet, V.M.P.; Dolven, J.K.; Rygg, B. Intercalibration of benthic foraminiferal and macrofaunal biotic indices: An example from the Norwegian Skagerrak coast (NE North Sea). Ecol. Indic. 2019, 96, 107–115. [Google Scholar] [CrossRef]
- de Jesus, M.S.D.S.; Frontalini, F.; Bouchet, V.M.; Yamashita, C.; Sartoretto, J.R.; Figueira, R.C.; e Sousa, S.H.D.M. Reconstruction of the Palaeo-Ecological Quality Status in an Impacted Estuary Using Benthic Foraminifera: The Santos Estuary (São Paulo State, SE Brazil). Mar. Environ. Res. 2020, 162, 105121. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Originally Used List [47] | This Study |
|---|---|---|
| Bolivina dilatata | EGII | EGIV |
| Bulimina aculeata | EGIII | EGIV |
| Quinqueloculina lamarckiana | NA | EGI |
| Quinqueloculina stalkeri | NA | EGIV |
| Rosalina bradyi | EGI | EGIII |
| Triloculina oblonga | NA | EG I |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dubois, A.; Barras, C.; Pavard, J.-C.; Donnay, A.; Béatrix, M.; Bouchet, V.M.P. Distribution Patterns of Benthic Foraminifera in Fish Farming Areas (Corsica, France): Implications for the Implementation of Biotic Indices in Biomonitoring Studies. Water 2021, 13, 2821. https://doi.org/10.3390/w13202821
Dubois A, Barras C, Pavard J-C, Donnay A, Béatrix M, Bouchet VMP. Distribution Patterns of Benthic Foraminifera in Fish Farming Areas (Corsica, France): Implications for the Implementation of Biotic Indices in Biomonitoring Studies. Water. 2021; 13(20):2821. https://doi.org/10.3390/w13202821
Chicago/Turabian StyleDubois, Antoine, Christine Barras, Jean-Charles Pavard, Annick Donnay, Melvin Béatrix, and Vincent M. P. Bouchet. 2021. "Distribution Patterns of Benthic Foraminifera in Fish Farming Areas (Corsica, France): Implications for the Implementation of Biotic Indices in Biomonitoring Studies" Water 13, no. 20: 2821. https://doi.org/10.3390/w13202821