
Vertical Boundary Mixing Events during Stratification Govern Heat and Nutrient Dynamics in a Windy Tropical Reservoir Lake with Important Water-Level Fluctuations: A Long-Term (2001–2021) Study

 , ,
, ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
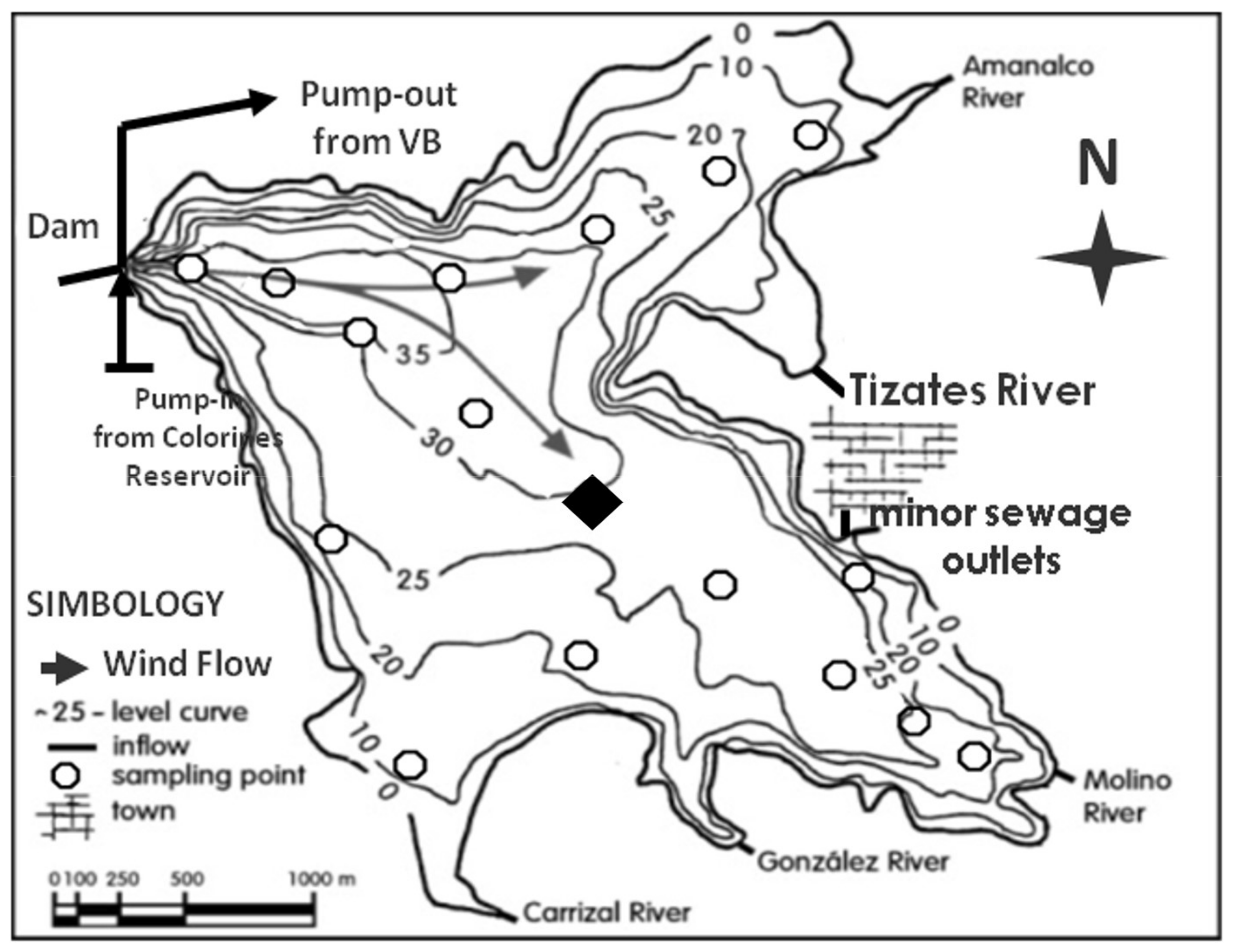
2.1. Study Area
2.2. Long-Term Monitoring of VB
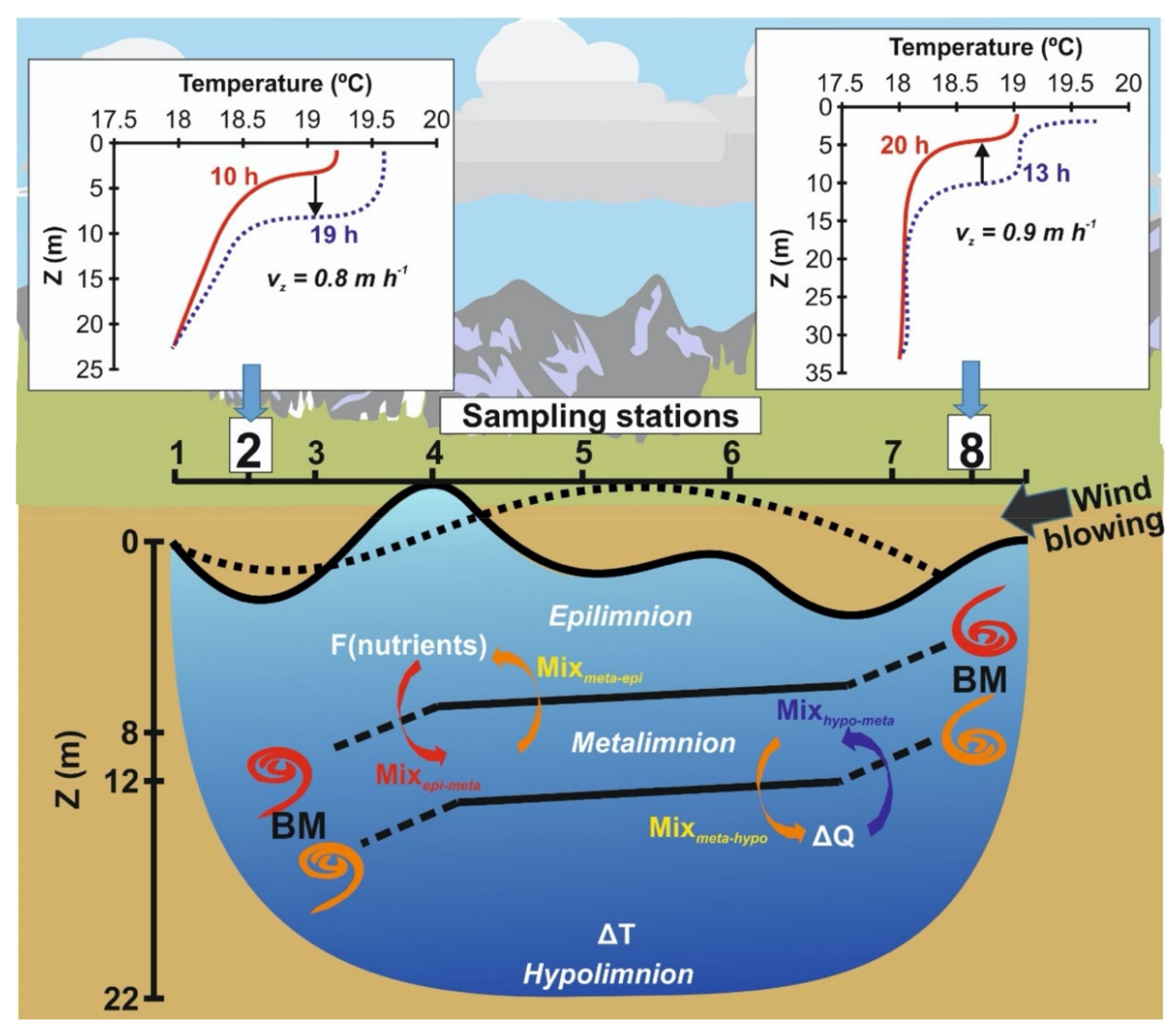
2.3. Vertical Mixing Model and Calculations
2.4. Hypolimnetic Heat Balance and Calculation of Vertical Water Mixing
2.5. Calculation of Vertical Nutrient Fluxes
2.6. Data Statistical Analysis
3. Results
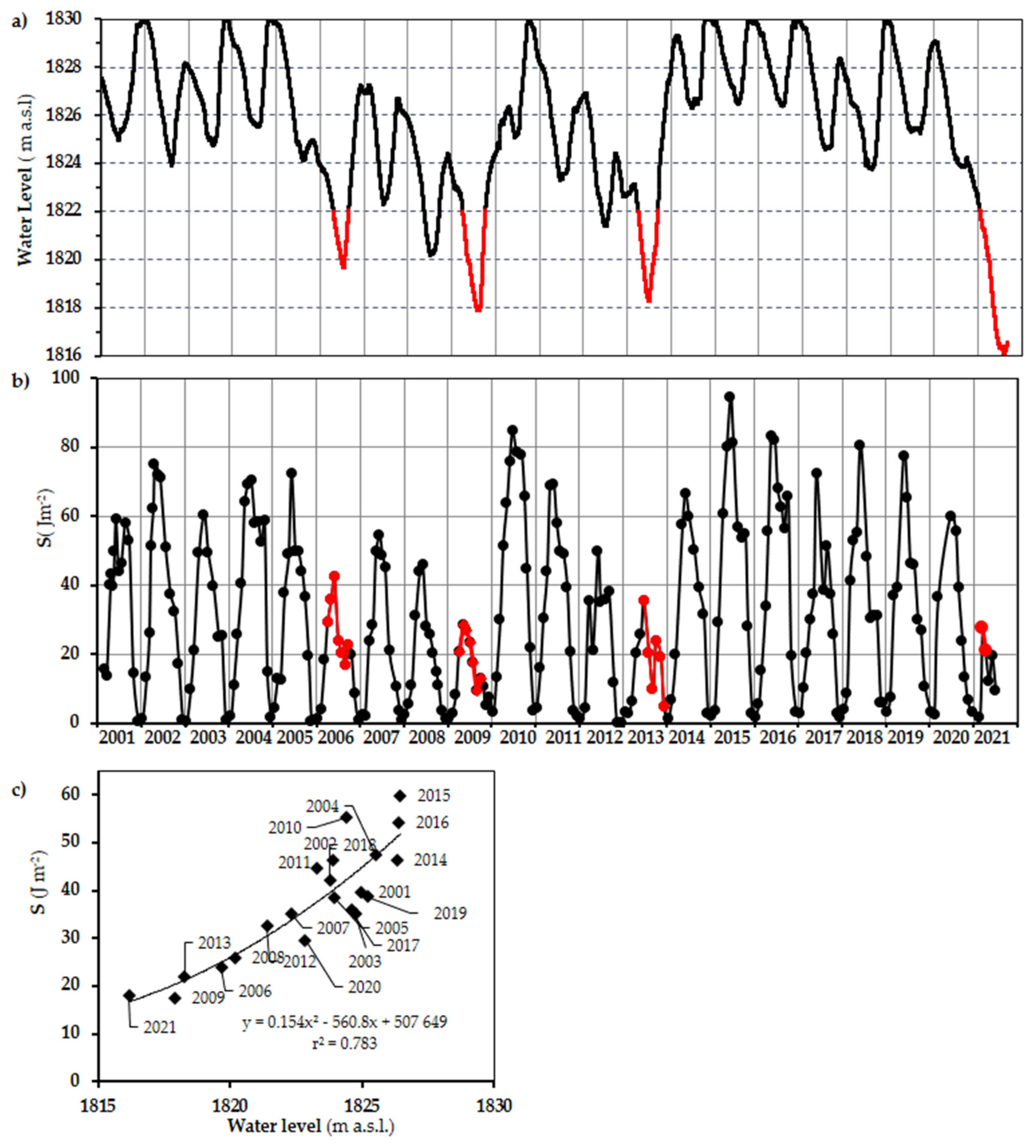
3.1. Water Level Fluctuations (WLFs) in VB during 2001–2021
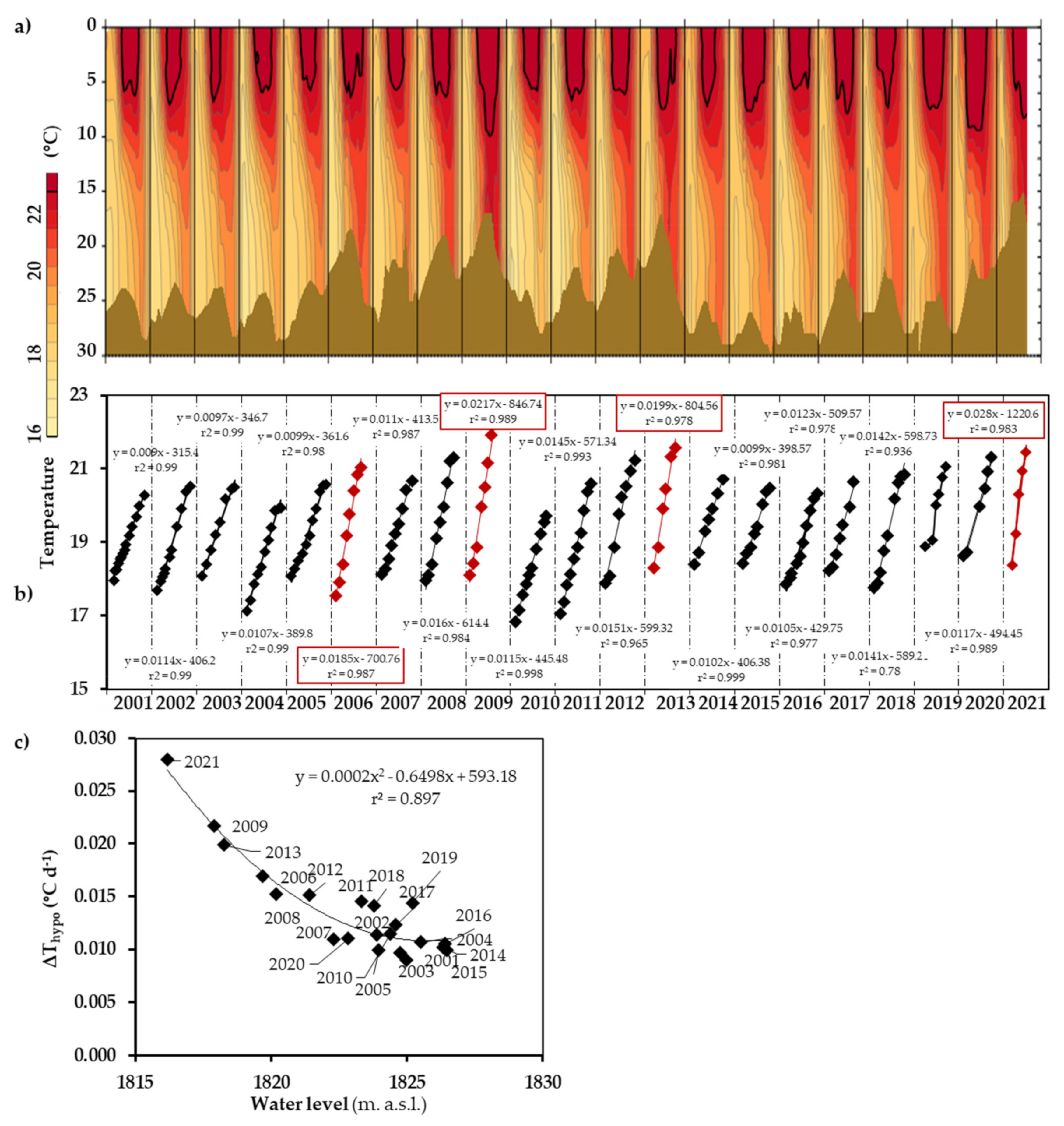
3.2. Thermal Regime and Stratification
3.3. Hypolimnetic Warming
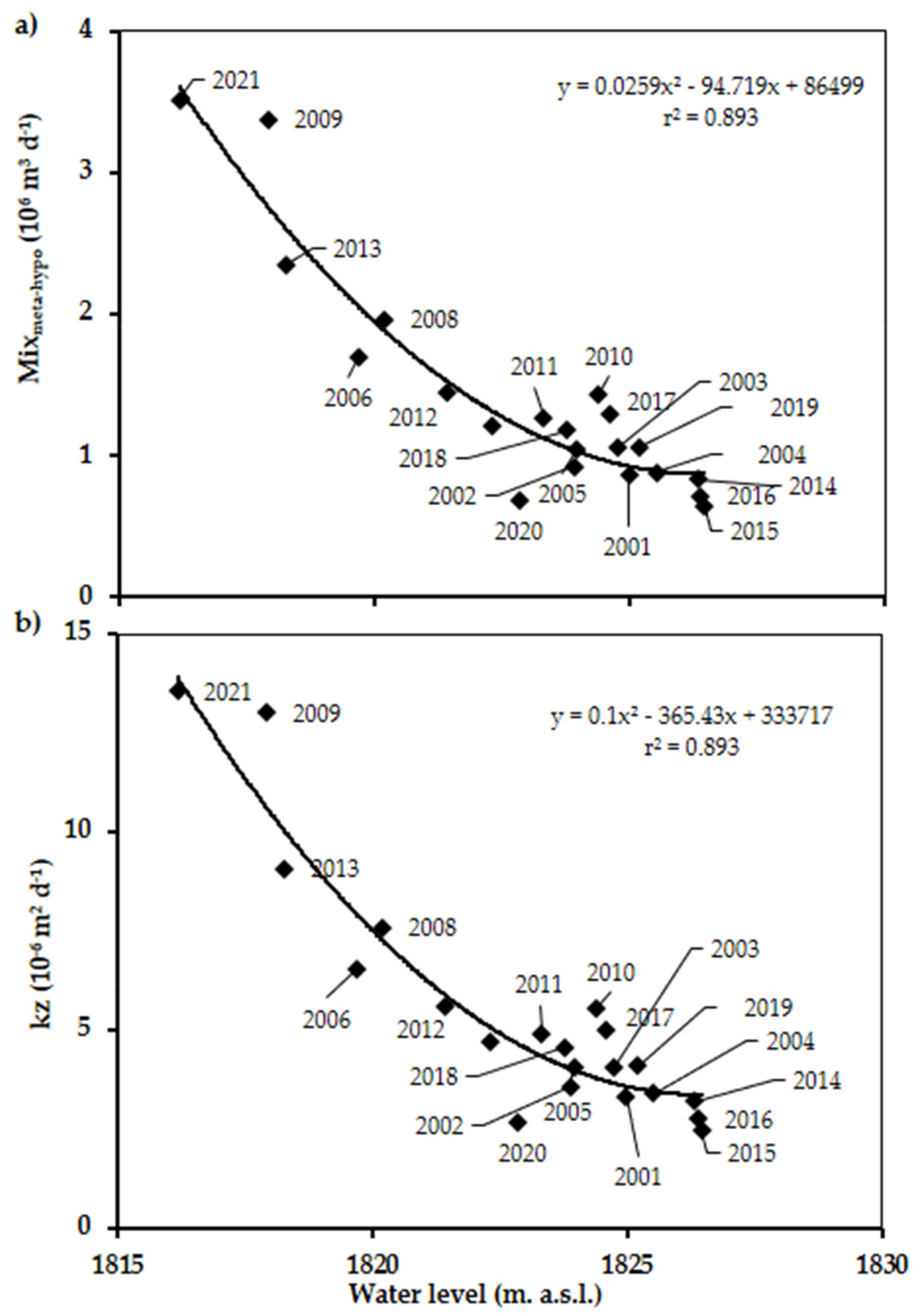
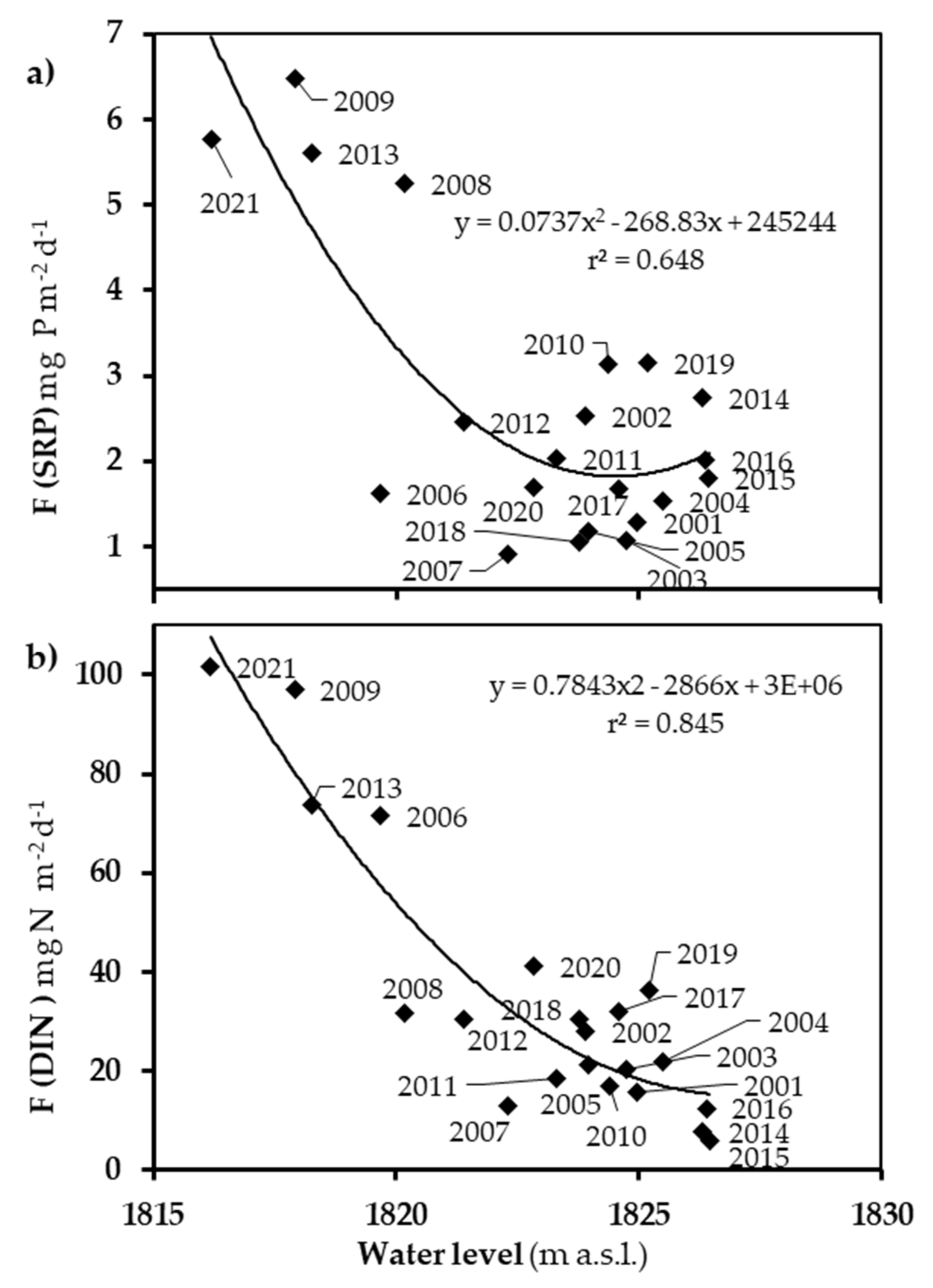
3.4. Mixmeta-hypo, Kz, and Nutrient Fluxes
4. Discussion
4.1. Hypolimnetic Warming Allows the Assessment of Vertical Mixing Rates during Stratification
4.2. Vertical Mixing Increases Up to Fivefold as the Water Level Decreases
4.3. The Contribution of Vertical Nutrient Fluxes to the Surface Layer Budget Can More than Duplicate as the Water Level Decreases
4.4. The Assessment of Vertical Nutrient Fluxes Also Helps Understanding Some Ecological Variations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Imberger, J. Flux paths in a stratified lake: A review. In Physical Processes in Lakes and Oceans; Coastal and Estuarine Studies; American Geophysical Union: Washington, DC, USA, 1998; pp. 1–17. ISBN 9781118665381. [Google Scholar]
- Kalff, J. Limnology: Inland Water Ecosystems; Prentice Hall: Upper Saddle River, NJ, USA, 2002; ISBN 0130337757. [Google Scholar]
- Kamarainen, A.M.; Penczykowski, R.M.; van de Bogert, M.C.; Hanson, P.C.; Carpenter, S.R. Phosphorus sources and demand during summer in a eutrophic lake. Aquat. Sci. 2009, 71, 214–227. [Google Scholar] [CrossRef]
- Lean, D.R.S.; Abbott, A.A.; Pick, F. Phosphorus deficiency of lake Ontario plankton. Can. J. Fish. Aquat. Sci. 1987, 44, 2069–2076. [Google Scholar] [CrossRef]
- Staehr, P.; Sand-Jensen, K. Temporal dynamics and regulation of lake metabolism. Limnol. Oceanogr. 2007, 52, 108–120. [Google Scholar] [CrossRef] [Green Version]
- MacIntyre, S.; Flynn, K.M.; Jellison, R.; Romero, J.R. Boundary mixing and nutrient fluxes in Mono Lake, California. Limnol. Oceanogr. 1999, 44, 512–529. [Google Scholar] [CrossRef]
- MacIntyre, S.; Jellison, R. Nutrient fluxes from upwelling and enhanced turbulence at the top of the pycnocline in Mono Lake, California. Hydrobiologia 2001, 466, 13–29. [Google Scholar] [CrossRef]
- Auger, G.; Yamazaki, H.; Nagai, T.; Jiao, C.; Kumagai, M. Hypolimnetic turbulence generation associated with superposition of large-scale internal waves in a strongly stratified lake: Lake Biwa, Japan. Limnology 2013, 14, 229–238. [Google Scholar] [CrossRef]
- Zohary, T.; Ostrovsky, I. Ecological impacts of excessive water level fluctuations in stratified freshwater lakes. Inland Waters 2011, 1, 47–59. [Google Scholar] [CrossRef]
- Valdespino-Castillo, P.M.; Merino-Ibarra, M.; Ramírez-Zierold, J.A.; Castillo, F.S.; González-de-Zayas, R.; Carnero-Bravo, V. Hacia el inventario de flujos de carbono en aguas tropicales: Unificar métodos. Tecnol. Cienc. Agua 2019, 10, 234–252. [Google Scholar] [CrossRef]
- Merino-Ibarra, M.; Monroy-Ríos, E.; Vilaclara, G.; Castillo, F.S.; Gallegos, M.E.; Ramírez-Zierold, J. Physical and chemical limnology of a wind-swept tropical highland reservoir. Aquat. Ecol. 2007, 42, 335–345. [Google Scholar] [CrossRef]
- Merino, M.; Monroy, E.; Castillo, S.; Ramírez, J.; Gallegos, M.E.; Alcocer, J. Wind-driven upwelling indicated by hydrological variability in a tropical eutrophied reservoir, Valle de Bravo, Mexico. In Proceedings of the 4th International Conference on Reservoir Limnology and Water Quality, České Budějovice, Czech Republic, 12–16 August 2002; pp. 228–231. [Google Scholar]
- Valdespino-Castillo, P.M.; Merino-Ibarra, M.; Jiménez-Contreras, J.; Sandoval, F.S.C.; Ramírez-Zierold, J.A. Community metabolism in a deep (stratified) tropical reservoir during a period of high water-level fluctuations. Environ. Monit. Assess. 2014, 186, 6505–6520. [Google Scholar] [CrossRef] [Green Version]
- Guimarais-Bermejo, M.O.; Merino-Ibarra, M.; Valdespino-Castillo, P.; Sandoval, F.S.C.; Ramírez-Zierold, J.A. Metabolism in a deep hypertrophic aquatic ecosystem with high water-level fluctuations: A decade of records confirms sustained net heterotrophy. PeerJ 2018, 6, e5205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramírez-Zierold, J.A.; Merino-Ibarra, M.; Monroy-Ríos, E.; Olson, M.; Castillo, F.S.; Gallegos, M.E.; Vilaclara, G. Changing water, phosphorus and nitrogen budgets for Valle de Bravo reservoir, water supply for Mexico City Metropolitan Area. Lake Reserv. Manag. 2010, 26, 23–34. [Google Scholar] [CrossRef]
- Fadel, A.; Lemaire, B.J.; Vinçon-Leite, B.; Atoui, A.; Slim, K.; Tassin, B. On the successful use of a simplified model to simulate the succession of toxic cyanobacteria in a hypereutrophic reservoir with a highly fluctuating water level. Environ. Sci. Pollut. Res. 2017, 24, 20934–20948. [Google Scholar] [CrossRef] [PubMed]
- Valeriano-Riveros, M.E.; Vilaclara, G.; Castillo-Sandoval, F.S.; Merino-Ibarra, M. Phytoplankton composition changes during water level fluctuations in a high-altitude, tropical reservoir. Inland Waters 2014, 4, 337–348. [Google Scholar] [CrossRef]
- May, L.; Aura, C.M.; Becker, V.; Briddon, C.L.; Carvalho, L.R.; Dobel, A.J.; Jamwal, P.; Kamphuis, B.; Marinho, M.M.; McGowan, S.; et al. Getting into hot water: Water quality in tropical lakes in relation to their utilisation. IOP Conf. Ser. Earth Environ. Sci. 2021, 789, 012021. [Google Scholar] [CrossRef]
- Bootsma, H.A.; Hecky, R.E. Water Quality Report, Lake Malawi/Nyasa Biodiversity Conservation Project; Southern African Development Community (SADC/GEF): Gaborone, Botswana; Global Environment Facility (GEF): Washington, DC, USA, 1999. [Google Scholar]
- Palter, J.; Lozier, M.S.; Barber, R.T. The effect of advection on the nutrient reservoir in the North Atlantic subtropical gyre. Nature 2005, 437, 687–692. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.-S.; Shen, S.-L.; Zhou, A.; Lyu, H.-M. Assessment and management of lake eutrophication: A case study in Lake Erhai, China. Sci. Total Environ. 2021, 751, 141618. [Google Scholar] [CrossRef]
- Díaz-Torres, O.; de Anda, J.; Lugo-Melchor, O.Y.; Pacheco, A.; Orozco-Nunnelly, D.A.; Shear, H.; Senés-Guerrero, C.; Gradilla-Hernández, M.S. Rapid changes in the phytoplankton community of a subtropical, shallow, hypereutrophic lake during the rainy season. Front. Microbiol. 2021, 12, 415. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, M.; Bleninger, T.; Lorke, A. Hydrodynamics and mixing mechanisms in a subtropical reservoir. Inland Waters 2021, 11, 1–16. [Google Scholar] [CrossRef]
- Alcocer, J.; Merino-Ibarra, M.; Ramírez-Zierold, J.A.; Oseguera, L.A.; Cortés-Guzmán, D.; Castillo-Sandoval, F.S.; Guzmán-Arias, A.P.; Pérez-Ramírez, M.G. Lake Alchichica limnology. In Lake Alchichica Limnology. The Uniqueness of a Tropical Maar Lake; Springer Nature: Berlin/Heidelberg, Germany, 2021; p. 491. ISBN 978-3-030-79095-0. in press. [Google Scholar]
- Nandini, S.; Merino-Ibarra, M.; Sarma, S.S.S. Seasonal changes in the zooplankton abundances of the reservoir Valle de Bravo (State of Mexico, Mexico). Lake Reserv. Manag. 2008, 24, 321–330. [Google Scholar] [CrossRef] [Green Version]
- Kirkwood, D.S. Sanplus Segmented Flow Analyzer and its Applications. Seawater Analysis; Skalar: Amsterdam, The Netherlands, 1994. [Google Scholar]
- Thorpe, S.A. Statically unstable layers produced by overturning internal gravity waves. J. Fluid Mech. 1994, 260, 333–350. [Google Scholar] [CrossRef]
- MacIntyre, S.; Melack, J.M. Vertical and horizontal transport in lakes: Linking littoral, benthic, and pelagic habitats. J. N. Am. Benthol. Soc. 1995, 14, 599–615. [Google Scholar] [CrossRef]
- Ostrovsky, I.; Yacobi, Y.Z.; Walline, P.; Kalikhman, I. Seiche-induced mixing: Its impact on lake productivity. Limnol. Oceanogr. 1996, 41, 323–332. [Google Scholar] [CrossRef]
- Idso, S.B. On the concept of lake stability 1. Limnol. Oceanogr. 1973, 18, 681–683. [Google Scholar] [CrossRef]
- MacIntyre, S. Vertical mixing in a shallow, eutrophic lake: Possible consequences for the light climate of phytoplankton. Limnol. Oceanogr. 1993, 38, 798–817. [Google Scholar] [CrossRef]
- Brookes, J. Effects of diurnal vertical mixing and stratification on phytoplankton productivity in geothermal Lake Rotowhero, New Zealand. Inland Waters 2013, 3, 369–376. [Google Scholar] [CrossRef] [Green Version]
- Lewis, W.M. Vertical eddy diffusivities in a large tropical lake 1. Limnol. Oceanogr. 1982, 27, 161–163. [Google Scholar] [CrossRef]
- Yang, P.; Fong, D.A.; Lo, E.; Monismith, S.G. Vertical mixing in a shallow tropical reservoir. Limnology 2019, 20, 279–296. [Google Scholar] [CrossRef]
- Stauffer, R.E. Vertical nutrient transport and its effects on epilimnetic phosphorus in four calcareous lakes. Hydrobiologia 1987, 154, 87–102. [Google Scholar] [CrossRef]
- Saggio, A.; Imberger, J. Mixing and turbulent fluxes in the metalimnion of a stratified lake. Limnol. Oceanogr. 2001, 46, 392–409. [Google Scholar] [CrossRef]
- Bormans, M.; Ford, P.W.; Fabbro, L.; Hancock, G. Onset and persistence of cyanobacterial blooms in a large impounded tropical river, Australia. Mar. Freshw. Res. 2004, 55, 1–15. [Google Scholar] [CrossRef]
- Preusse, M.; Peeters, F.; Lorke, A. Internal waves and the generation of turbulence in the thermocline of a large lake. Limnol. Oceanogr. 2010, 55, 2353–2365. [Google Scholar] [CrossRef] [Green Version]
- Lewis, W.M.J. Temperature, heat, and mixing in Lake Valencia, Venezuela 1. Limnol. Oceanogr. 1983, 28, 273–286. [Google Scholar] [CrossRef] [Green Version]
- Robarts, R.D.; Ward, P.R.B. Vertical diffusion and nutrient transport in a tropical lake (Lake McIlwaine, Rhodesia). Hydrobiologia 1978, 59, 213–221. [Google Scholar] [CrossRef]
- Katsev, S.; Crowe, S.A.; Mucci, A.; Sundby, B.; Nomosatryo, S.; Haffner, G.D.; Fowle, D.A. Mixing and its effects on biogeochemistry in the persistently stratified, deep, tropical Lake Matano, Indonesia. Limnol. Oceanogr. 2010, 55, 763–776. [Google Scholar] [CrossRef]
- Jellison, R.; Melack, J.M. Meromixis in hypersaline Mono Lake, California. 1. Stratification and vertical mixing during the onset, persistence, and breakdown of meromixis. Limnol. Oceanogr. 1993, 38, 1008–1019. [Google Scholar] [CrossRef]
- Goudsmit, G.-H.; Peeters, F.; Gloor, M.; Wüest, A. Boundary versus internal diapycnal mixing in stratified natural waters. J. Geophys. Res. Space Phys. 1997, 102, 27903–27914. [Google Scholar] [CrossRef] [Green Version]
- Jassby, A.; Powell, T. Vertical patterns of eddy diffusion during stratification in Castle Lake, California 1. Limnol. Oceanogr. 1975, 20, 530–543. [Google Scholar] [CrossRef]
- Lin, S.; Boegman, L.; Rao, Y. Characterizing spatial and temporal distributions of turbulent mixing and dissipation in Lake Erie. J. Great Lakes Res. 2021, 47, 168–179. [Google Scholar] [CrossRef]
- Pasche, N.; Dinkel, C.; MuxFCller, B.; Schmid, M.; Est, A.W.; Wehrli, B. Physical and biogeochemical limits to internal nutrient loading of meromictic Lake Kivu. Limnol. Oceanogr. 2009, 54, 1863–1873. [Google Scholar] [CrossRef] [Green Version]
- Kamarainen, A.M.; Yuan, H.; Wu, C.H.; Carpenter, S.R. Estimates of phosphorus entrainment in Lake Mendota: A comparison of one-dimensional and three-dimensional approaches. Limnol. Oceanogr. Methods 2009, 7, 553–567. [Google Scholar] [CrossRef]
- Hamblin, P.F.; Bootsma, H.A.; Hecky, R.E. Modeling nutrient upwelling in Lake Malawi/Nyasa. J. Great Lakes Res. 2003, 29, 34–47. [Google Scholar] [CrossRef]
- Duan, S.; Newcomer-Johnson, T.; Mayer, P.; Kaushal, S. Phosphorus retention in stormwater control structures across streamflow in urban and suburban watersheds. Water 2016, 8, 390. [Google Scholar] [CrossRef] [Green Version]
- Lau, M.P.; Valerio, G.; Pilotti, M.; Hupfer, M. Intermittent meromixis controls the trophic state of warming deep lakes. Sci. Rep. 2020, 10, 1–16. [Google Scholar] [CrossRef]
- Edwards, R.L.; Priscu, J.C. McMurdo LTER: Relationships between vertical nutrient flux and phytoplankton biomass and productivity in lakes of the Taylor Valley, Antarctica. Antarct. J. US 1995, 30, 294–295. [Google Scholar]
- Cendejas, J.C.; Ramírez, L.M.; Zierold, J.R.; Valenzuela, J.D.; Ibarra, M.M.; de Tagle, S.M.S.; Téllez, A.C. Evaluation of the impacts of land use in water quality and the role of nature-based solutions: A citizen science-based study. Sustainability 2021, 13, 10519. [Google Scholar] [CrossRef]
- Pomati, F.; Matthews, B.; Jokela, J.; Schildknecht, A.; Ibelings, B.W. Effects of re-oligotrophication and climate warming on plankton richness and community stability in a deep mesotrophic lake. Oikos 2012, 121, 1317–1327. [Google Scholar] [CrossRef]
- Matzinger, A.; Schmid, M.; Veljanoska-Sarafiloska, E.; Patceva, S.; Guseska, D.; Wagner, B.; Müller, B.; Sturm, M.; Wüest, A. Eutrophication of ancient Lake Ohrid: Global warming amplifies detrimental effects of increased nutrient inputs. Limnol. Oceanogr. 2007, 52, 338–353. [Google Scholar] [CrossRef]
- Rabalais, N.N.; Turner, R.E.; Díaz, R.J.; Justić, D. Global change and eutrophication of coastal waters. ICES J. Mar. Sci. 2009, 66, 1528–1537. [Google Scholar] [CrossRef]
- Boyce, D.G.; Lewis, M.R.; Worm, B. Global phytoplankton decline over the past century. Nature 2010, 466, 591–596. [Google Scholar] [CrossRef]
- Bartosiewicz, M.; Przytulska, A.; Deshpande, B.N.; Antoniades, D.; Cortes, A.; MacIntyre, S.; Lehmann, M.; Laurion, I. Effects of climate change and episodic heat events on cyanobacteria in a eutrophic polymictic lake. Sci. Total Environ. 2019, 693, 133414. [Google Scholar] [CrossRef]
- Xu, H.; Yan, M.; Long, L.; Ma, J.; Ji, D.; Liu, D.; Yang, Z. Modeling the effects of hydrodynamics on thermal stratification and algal blooms in the Xiangxi Bay of Three Gorges Reservoir. Front. Ecol. Evol. 2021, 8, 453. [Google Scholar] [CrossRef]
- De Crop, W.; Verschuren, D. Determining patterns of stratification and mixing in tropical crater lakes through intermittent water-column profiling: A case study in western Uganda. J. Afr. Earth Sci. 2019, 153, 17–30. [Google Scholar] [CrossRef]
- Gutiérrez, S.G.; Sarma, S.S.S.; Nandini, S. Seasonal variations of rotifers from a high altitude urban shallow water body, La Cantera Oriente (Mexico City, Mexico). Chin. J. Oceanol. Limnol. 2016, 35, 1387–1397. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stratification Period | Mixmeta-hypo (106 m3 day−1) | Kz (m2 s−1) | F (SRP) (mg P m−2 day−1) | F (DIN) (mg N m−2 day−1) |
|---|---|---|---|---|
| 2001 | 0.861 | 3.3 × 10−6 | 0.79 | 15.9 |
| 2002 | 0.924 | 3.6 × 10−6 | 2.03 | 28.0 |
| 2003 | 1.057 | 4.1 × 10−6 | 0.57 | 20.3 |
| 2004 | 0.880 | 3.4 × 10−6 | 1.03 | 21.7 |
| 2005 | 1.046 | 4.0 × 10−6 | 0.68 | 21.1 |
| 2006 | 1.691 | 6.5 × 10−6 | 1.12 | 71.5 |
| 2007 | 1.212 | 4.7 × 10−6 | 0.42 | 13.0 |
| 2008 | 1.963 | 7.6 × 10−6 | 4.75 | 31.6 |
| 2009 | 3.373 | 13.0 × 10−6 | 5.99 | 97.1 |
| 2010 | 1.439 | 5.6 × 10−6 | 2.64 | 17.0 |
| 2011 | 1.271 | 4.9 × 10−6 | 1.54 | 18.6 |
| 2012 | 1.446 | 6.6 × 10−6 | 1.95 | 30.4 |
| 2013 | 2.349 | 9.1 × 10−6 | 5.11 | 73.8 |
| 2014 | 0.832 | 3.2 × 10−6 | 2.24 | 7.6 |
| 2015 | 0.639 | 2.5 × 10−6 | 1.30 | 5.8 |
| 2016 | 0.712 | 2.7 × 10−6 | 1.52 | 12.5 |
| 2017 | 1.296 | 5.0 × 10−6 | 1.17 | 32.0 |
| 2018 | 1.180 | 4.6 × 10−6 | 0.56 | 30.4 |
| 2019 | 1.059 | 4.1 × 10−6 | 2.65 | 36.3 |
| 2020 | 0.691 | 2.6 × 10−6 | 1.20 | 41.0 |
| 2021 | 3.515 | 13.6 × 10−6 | 5.27 | 101.7 |
| 2001–2021 Mean | 1.289 | 3.1 × 10−6 | 2.12 | 34.6 |
| Kz | Location | Reference | |
|---|---|---|---|
| Mean | (Range) | ||
| 9.9 × 10−6 | (1.2 × 10−7–5.0 × 10−5) | VB (monthly) | This study |
| 3.1 × 10−6 | (2.5 × 10−6–13.6 × 10−6) | VB (annual mean) | This study |
| 5.0 × 10−4 | (1.0 × 10−5–1.0 × 10−3) | North Lake, Australia | [31] MacIntyre 1993 |
| 1.0 × 10−4 | (8.0 × 10−5–1.3 × 10−4) | Lake Rotowhero, New Zealand | [32] Brookes et al. 2013 |
| 6.5 × 10−5 | (2.2 × 10−5–1.1 × 10−4) | Lake Lanao, Philippines | [33] Lewis 1982 |
| 5.1 × 10−5 | (1.0 × 10−6–1.0 × 10−4) | Lake Biwa, Japan | [8] Auger et al. 2014 |
| 5.1 × 10−5 | (1.0 × 10−6–1.0 × 10−4) | Kranji Reservoir, Singapore | [34] Yang et al. 2019 |
| 4.5 × 10−5 | (1.0 × 10−6–9.0 × 10−5) | Green Lake, | [35] Stauffer 1987 |
| 4.5 × 10−5 | (8.0 × 10−5–1.0 × 10−5) | Lake Kinneret, Israel | [36] Saggio and Imberger 2001 |
| 1.8 × 10−5 | (1.6 × 10−5–2.1 × 10−5) | Fitzroy River Barrage, Australia | [37] Bormans et al. 2004 |
| 1.2 × 10−5 | Lake Constance, Germany-Switzerland | [38] Preusse et al. 2010 | |
| 9.9 × 10−6 | (4.0 × 10−8–5.0 × 10−5) | Valle de Bravo (monthly) | This study |
| 6.0 × 10−6 | (9.0 × 10−6–1.1 × 10−5) | Lake Valencia, Venezuela | [39] Lewis 1983 |
| 6.0 × 10−6 | (6.0 × 10−6–5.0 × 10−5) | Lake McIlwaine, Rhodesia | [40] Robarts and Ward 1978 |
| 5.0 × 10−6 | Lake Matano, Indonesia | [41] Katsev et al. 2010 | |
| 3.4 × 10−6 | (1.0 × 10−7–2.6 × 10−5) | Mono Lake, California | [42] Jellison and Melack 1993 |
| 3.1 × 10−6 | (2.5 × 10−6–13.6 × 10−6) | Valle de Bravo (annual mean) | This study |
| 2.2 × 10−6 | (3.0 × 10−6–8.0 × 10−5) | Lake Alpnach (central Switzerland) | [43] Goudsmith et al. 1997 |
| 1.3 × 10−6 | (2.0 × 10−7–2.5 × 10−6) | Castle Lake, California | [44] Jassby and Powell 1975 |
| 1.0 × 10−6 | (1.0 × 10−8–1.0 × 10−4) | Mono Lake, California | [6] MacIntyre et al. 1999 |
| 1.0 × 10−8 | Molecular conductivity | [6] MacIntyre et al. 1999 | |
| SRP (mg P m−2 day−1) | Ammonia (mg N m−2 day−1) | DIN (mg N m−2 day−1) | Lake | Reference |
|---|---|---|---|---|
| 0.10 | Lake Fish, Wisconsin, USA | [35] Stauffer 1987 | ||
| 1.60 | 6.2 | Lake McIlwaine, Rhodesia | [40] Robarts and Ward 1978 | |
| 2.12 | 13.9 | 34.6 | Valle de Bravo (overall mean) | This study |
| 2.71 | Lake Kivu, Africa | [46] Pasche et al. 2009 | ||
| 2.59 | Lake Mendota, Wisconsin, USA | [47] Kamarainen et al. 2009 | ||
| ~3.00 | ~20.0 | Lake Malawi/Nyasa, Africa | [48] Hamblin et al. 2003 | |
| 3.20 | Lakes of Taylor Valley, Antarctica | [51] Edwards and Priscu 1995 | ||
| 5.50 | Lake Green, Wisconsin, USA | [35] Stauffer 1987 | ||
| 5.99 | 66.0 | 101.7 | Valle de Bravo (lowest water level) | This study |
| 15.20 | Lake Delavan, Wisconsin, USA | [35] Stauffer 1987 | ||
| 39.2 | Mono Lake, California, USA | [6] MacIntyre et al. 1999 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Merino-Ibarra, M.; Ramírez-Zierold, J.A.; Valdespino-Castillo, P.M.; Castillo-Sandoval, F.S.; Guzmán-Arias, A.P.; Barjau-Aguilar, M.; Monroy-Ríos, E.; López-Gómez, L.M.; Sacristán-Ramírez, A.; Quintanilla-Terminel, J.G.; et al. Vertical Boundary Mixing Events during Stratification Govern Heat and Nutrient Dynamics in a Windy Tropical Reservoir Lake with Important Water-Level Fluctuations: A Long-Term (2001–2021) Study. Water 2021, 13, 3011. https://doi.org/10.3390/w13213011
Merino-Ibarra M, Ramírez-Zierold JA, Valdespino-Castillo PM, Castillo-Sandoval FS, Guzmán-Arias AP, Barjau-Aguilar M, Monroy-Ríos E, López-Gómez LM, Sacristán-Ramírez A, Quintanilla-Terminel JG, et al. Vertical Boundary Mixing Events during Stratification Govern Heat and Nutrient Dynamics in a Windy Tropical Reservoir Lake with Important Water-Level Fluctuations: A Long-Term (2001–2021) Study. Water. 2021; 13(21):3011. https://doi.org/10.3390/w13213011
Chicago/Turabian StyleMerino-Ibarra, Martín, Jorge A. Ramírez-Zierold, Patricia M. Valdespino-Castillo, Fermin S. Castillo-Sandoval, Andrea P. Guzmán-Arias, Mariel Barjau-Aguilar, Emiliano Monroy-Ríos, Luz M. López-Gómez, Arantxa Sacristán-Ramírez, José G. Quintanilla-Terminel, and et al. 2021. "Vertical Boundary Mixing Events during Stratification Govern Heat and Nutrient Dynamics in a Windy Tropical Reservoir Lake with Important Water-Level Fluctuations: A Long-Term (2001–2021) Study" Water 13, no. 21: 3011. https://doi.org/10.3390/w13213011