
Complexity of the Relationship between Environmental Factors, Interspecific Competition, and Intrinsic Traits of the Species in Explaining the Invasive Success of Gobio lozanoi Doadrio & Madeira, 2004

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
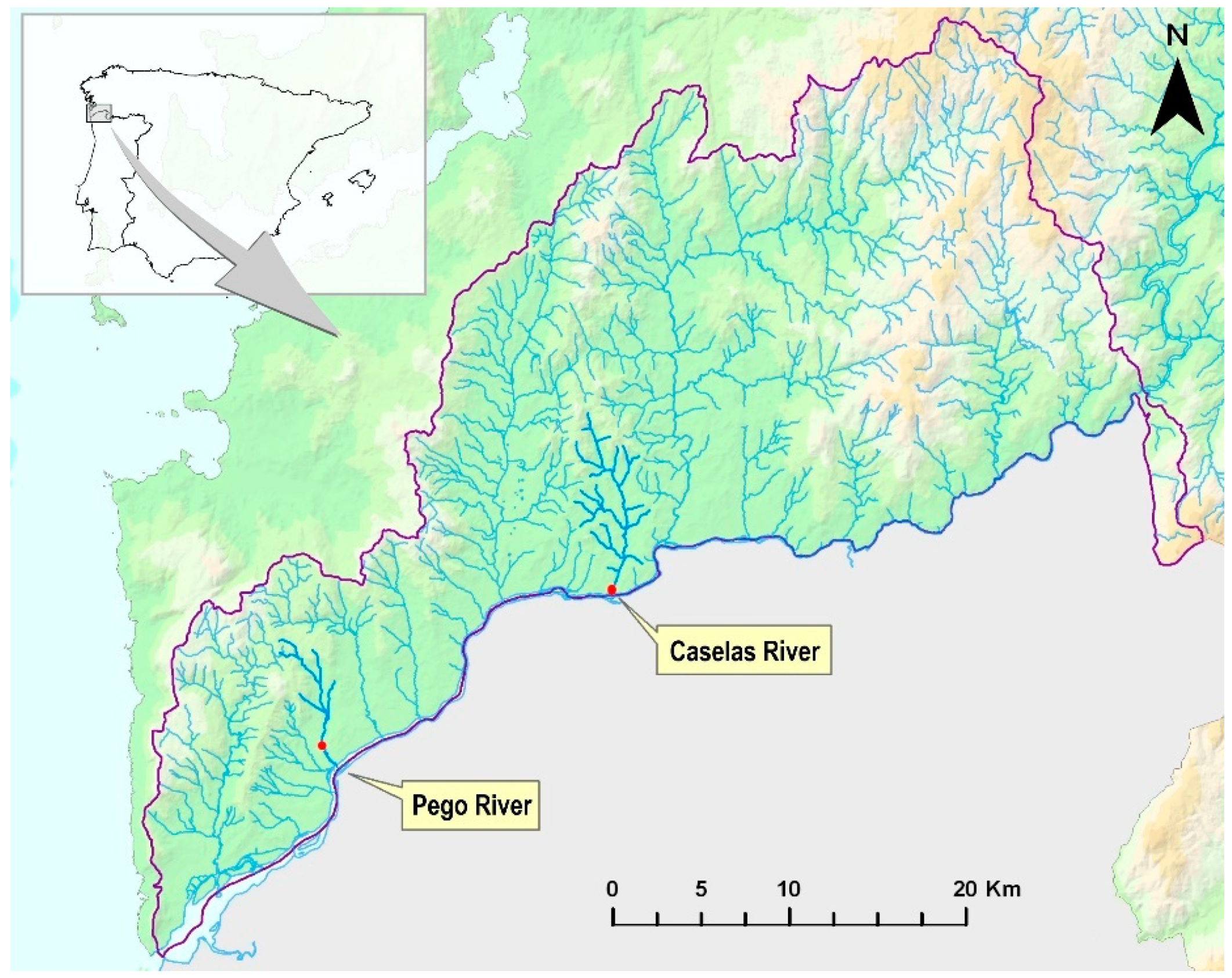
3.1. Characterization of Tributaries
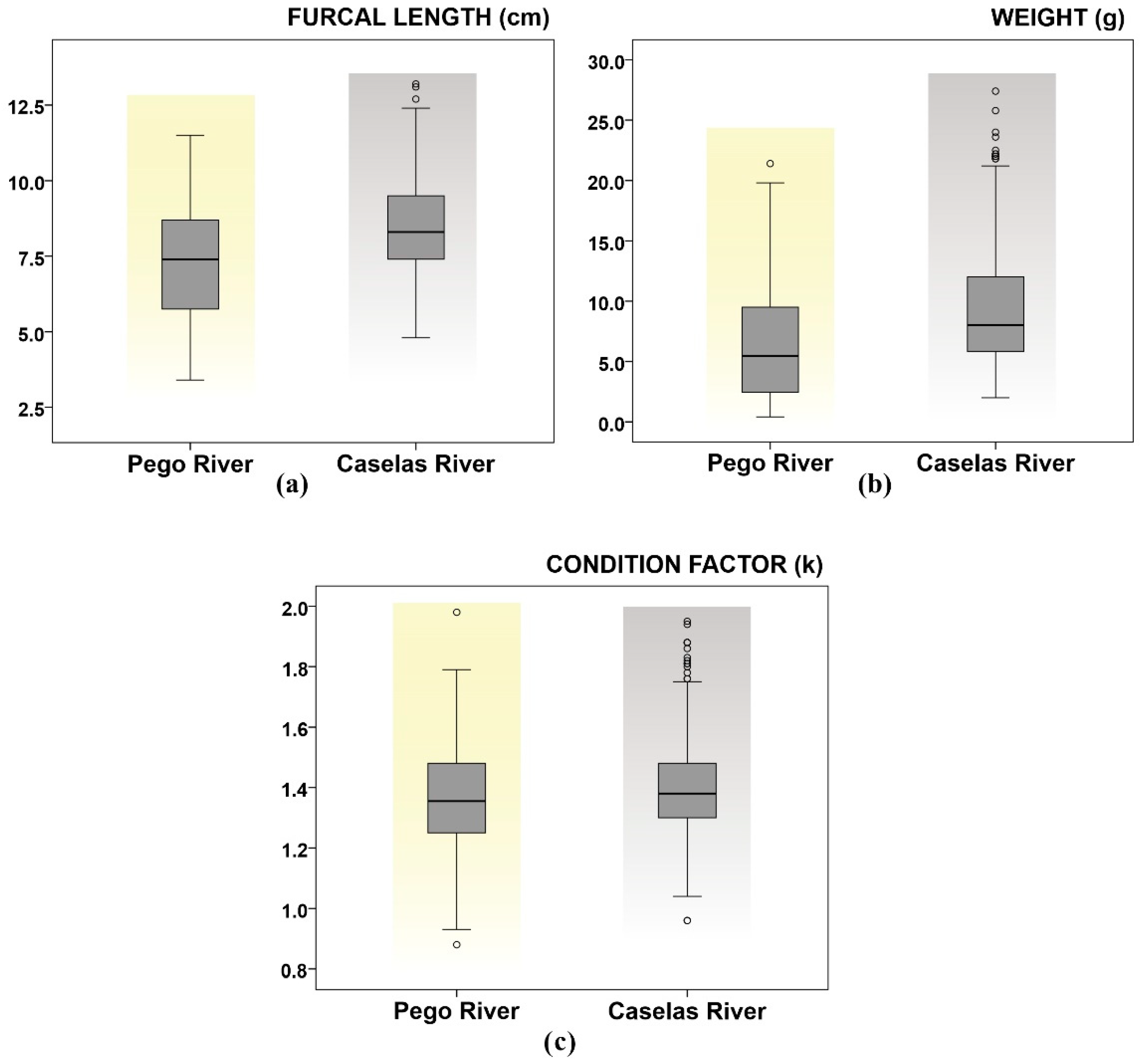
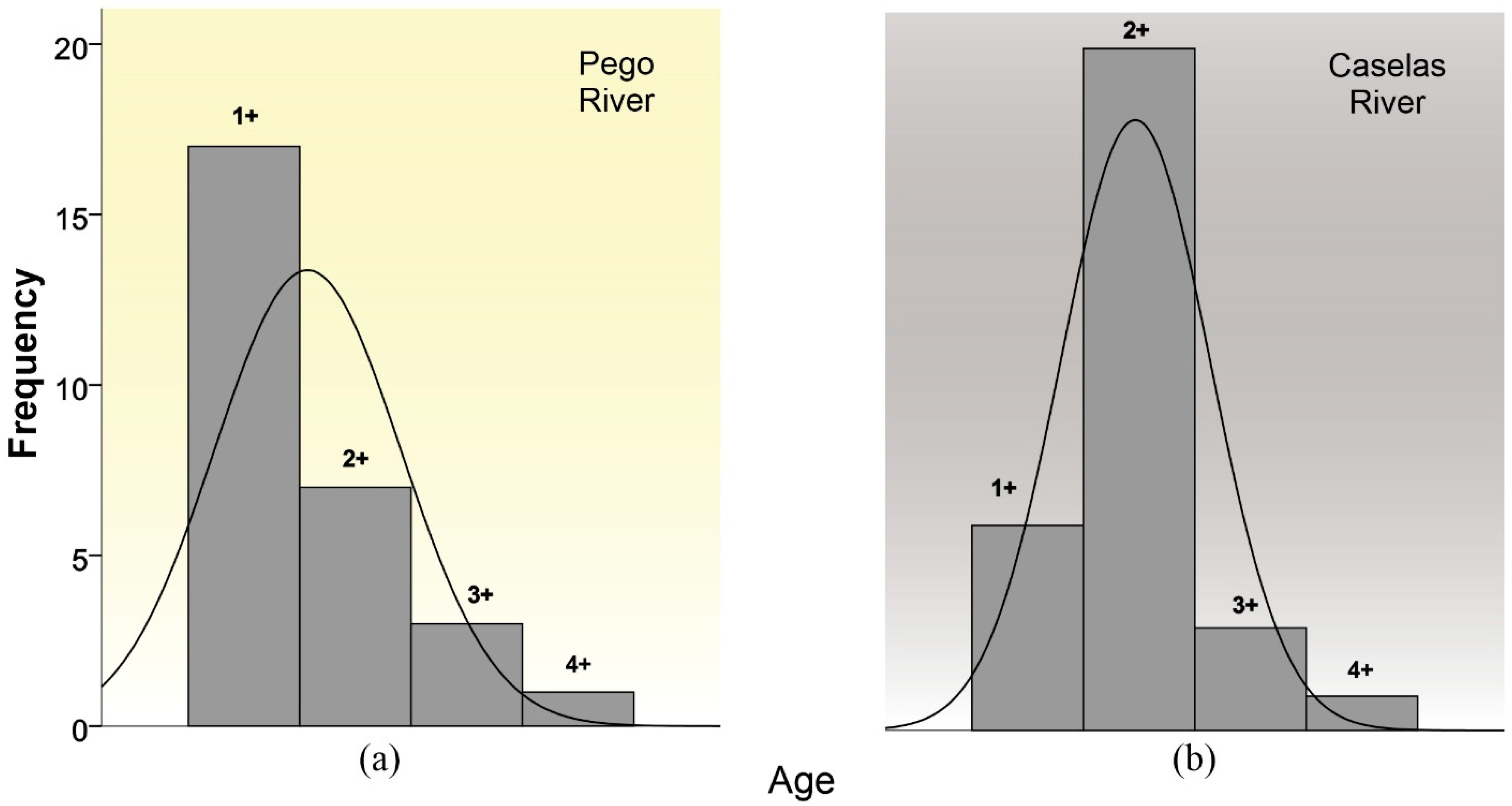
3.2. Characterization of the Two Populations of Gobio lozanoi
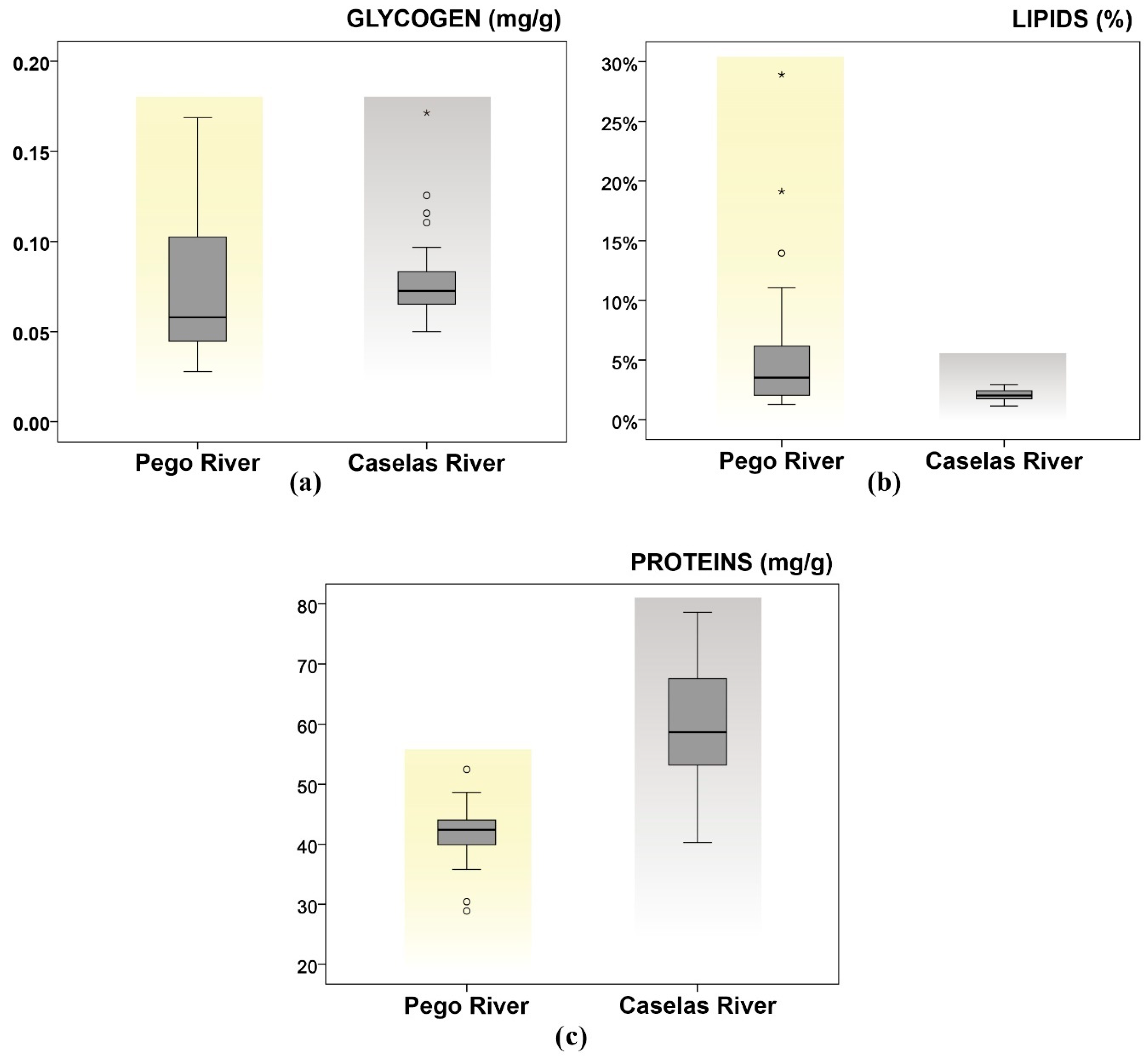
3.3. Biomarkers: Glycogen, Lipids and Proteins
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mack, R.N.; Simberloff, D.; Mark Lonsdale, W.; Evans, H.; Clout, M.; Bazzaz, F.A. Biotic invasions: Causes, epidemiology, global consequences, and control. Ecol. Appl. 2000, 10, 689–710. [Google Scholar] [CrossRef]
- Sala, O.E.; Chapin, F.S.; Armesto, J.J.; Berlow, E.; Bloomfield, J.; Dirzo, R.; Leemans, R. Global biodiversity scenarios for the year 2100. Science 2000, 287, 1770–1774. [Google Scholar] [CrossRef]
- Meyerson, L.A.; Mooney, H.A. Invasive Alien Species in an Era of Globalization. Front. Ecol. Environ. 2007, 5, 199–208. [Google Scholar] [CrossRef]
- Simberloff, D. Biological invasions: What’s worth fighting and what can be won? Ecol. Eng. 2014, 65, 112–121. [Google Scholar] [CrossRef]
- Ricciardi, A.; Cohen, J. The invasiveness of an introduced species does not predict its impact. Biol. Invasions 2007, 9, 309–315. [Google Scholar] [CrossRef]
- Hui, C.; Richardson, D.M.; Landi, P.; Minoarivelo, H.O.; Garnas, J.; Roy, H.E. Defining invasiveness and invasibility in ecological networks. Biol. Invasions 2016, 18, 971–983. [Google Scholar] [CrossRef] [Green Version]
- Smith, P.R.; Davey, S. Evidence for the competitive exclusion of Aeromonas salmonicida from fish with stress-inducible furunculosis by a fluorescent pseudomonad. J. Fish Dis. 1993, 16, 521–524. [Google Scholar] [CrossRef]
- Simon, K.S.; Townsend, C.R. Impacts of freshwater invaders at different levels of ecological organisation, with emphasis on salmonids and ecosystem consequences. Fresh. Biol. 2003, 48, 982–994. [Google Scholar] [CrossRef] [Green Version]
- Matsuzaki, S.I.S.; Sasaki, T.; Akasaka, M. Consequences of the introduction of exotic and translocated species and future extirpations on the functional diversity of freshwater fish assemblages. Glob. Ecol. Biogeogr. 2013, 22, 1071–1082. [Google Scholar] [CrossRef]
- Dorenbosch, M.; Bakker, E.S. Effects of contrasting omnivorous fish on submerged macrophyte biomass in temperate lakes: A mesocosm experiment. Fresh. Biol. 2012, 57, 1360–1372. [Google Scholar] [CrossRef]
- Hobbs, R.J. Invasive Species in a Changing World; Mooney, H.A., Hobbs, R.J., Eds.; Island Press: Washington, DC, USA, 2000; p. 457. [Google Scholar]
- McNeely, J. Invasive Species: A Costly Catastrophe for Native Biodiversity. Land Use Water Resour. Res. 2001, 2, 1–10. [Google Scholar]
- Andersen, M.C.; Adams, H.; Hope, B.; Powell, M. Risk assessment for invasive species. Risk Anal. Int. J. 2004, 24, 787–793. [Google Scholar] [CrossRef] [PubMed]
- Gurevitch, J.; Padilla, D.K. Are invasive species a major cause of extinctions? Trends. Ecol. Evol. 2004, 19, 470–474. [Google Scholar] [CrossRef] [PubMed]
- EEA (European Environment Agency). Halting the Loss of Biodiversity by 2010: Proposal for a First Set of Indicators to Monitor Progress in Europe; EEA Technical Report No. 11/2007; European Environment Agency: Copenhagen, Denmark, 2007; p. 186. [Google Scholar]
- Cobo, F.; Vieira-Lanero, R.; Rego, E.; Servia, M.J. Temporal trends in non-indigenous freshwater species records during the 20th century: A case study in the Iberian Peninsula. Biodivers. Conserv. 2010, 19, 3471–3487. [Google Scholar] [CrossRef]
- Doadrio, I.; Madeira, M.J. A new species of the genus Gobio Cuvier, 1816 (Actinopterygii, Cyprinidae) from the Iberian Peninsula and southwestern France. Graellsia 2004, 6, 107–116. [Google Scholar] [CrossRef]
- Alpert, P.; Bone, E.; Holzapfel, C. Invasiveness, invasibility and the role of environmental stress in the spread of non-native plants. PPEES 2000, 3, 52–66. [Google Scholar] [CrossRef] [Green Version]
- Byers, J.E.; Noonburg, E.G. Scale dependent effects of biotic resistance to biological invasion. Ecology 2003, 84, 1428–1433. [Google Scholar] [CrossRef]
- Brown, J.H.; Lomolino, M.V. Biogeography; Sinauer: Sunderland, MA, USA, 1998; p. 691. [Google Scholar]
- Ríos, H.F.; Vargas, O. Ecología de las especies invasoras. Pérez-Arbelaezia 2003, 14, 119–148. [Google Scholar]
- Lonsdale, W.M. Global patterns of plant invasions and the concept of invasibility. Ecology 1999, 80, 1522–1536. [Google Scholar] [CrossRef]
- Beckman, B.; Larsen, D.; Sharpe, C.; Lee-Pawlak, B.; Schreck, C.; Dickhoff, W. Physiological status of naturally reared juvenile spring Chinook salmon in the Yakima river: Seasonal dynamics and changes associated with smolting. Trans. Am. Fish. Soc. 2000, 129, 727–753. [Google Scholar] [CrossRef]
- Smutná, M.; Vorlová, L.; Svobodová, Z. Pathobiochemestry of ammoniain the internal environment of fish. Acta Vet. Brno 2002, 71, 169–181. [Google Scholar] [CrossRef] [Green Version]
- Quintana, C.F. Respuestas neuroendocrinas al estrés en peces teleósteos. Rev. Ictiol. 2002, 10, 57–78. [Google Scholar]
- Sánchez, J.; Barca, S.; Vieira-Lanero, R.; Servia, M.J.; Cobo, F. Cambios en la alimentación y en los niveles de glucógeno de la trucha común (Salmo trutta Linné, 1758) por efecto de la contaminación orgánica. AEMS-Ríos Con Vida 2008, 82, 14–16. [Google Scholar]
- Cobo, F.; Sánchez, J.; Vieira, R.; Servia, M. Organic pollution induces domestication like characteristics in feral populations of brown trout (Salmo trutta). Hydrobiologia 2013, 705, 119–134. [Google Scholar] [CrossRef]
- Post, J.; Parkinson, E. Energy allocation strategy in age-0 fish: Allometry and survival. Ecology 2001, 82, 1040–1051. [Google Scholar] [CrossRef]
- Barca, S. Biomarcadores Metabólicos y de Reserva Energética en Lamprea Marina (Petromyzon marinus Linnaeus, 1758). Ph.D. Thesis, Universidade de Santiago de Compostela, A Coruña, Spain, 2016. [Google Scholar]
- Villarroel, M.J.; Sancho, E.; Andreu-Moliner, E.; Ferrando, M.D. Biochemical stress response in tetradifon exposed Daphnia magna and its relationship to individual growth and reproduction. Sci. Total Environ. 2009, 407, 5537–5542. [Google Scholar] [CrossRef]
- Vieira-Lanero, R.; Servia, M.J.; Barca, S.; Couto, M.T.; Rivas, S.; Sánchez, J.; Nachón, D.; Silva, S.; Gómez-Sande, P.; Morquecho, C.; et al. Índices de Calidad de la Vegetación de Ribera y del Hábitat Fluvial en Los Afluentes de la Margen Española del Baixo Miño. In V Simposio Ibérico Sobre a Bacia Hidrográfica do Rio Minho; Câmara Municipal de Vila Nova de Cerveira: Vila Nova de Cerveira, Portugal, 2010; pp. 79–88. [Google Scholar]
- Barca, S.; Vieira-Lanero, R.; Servia, M.J.; Couto, M.T.; Rivas, S.; Sánchez, J.; Nachón, D.; Silva, S.; Gómez-Sande, P.; Morquecho, C.; et al. Nuevos datos sobre las características fisicoquímicas del río Miño y sus afluentes de la margen española. In V Simposio Ibérico Sobre a Bacia Hidrográfica do Rio Minho; Câmara Municipal de Vila Nova de Cerveira: Vila Nova de Cerveira, Portugal, 2010; pp. 131–137. [Google Scholar]
- Alba-Tercedor, J.; Sánchez-Ortega, A. Un método rápido y simple para evaluar la calidad biológica de las aguas corrientes basado en el de Hellawell (1978). Limnetica 1988, 4, 51–56. [Google Scholar]
- González, M.; Cobo, F. Macroinvertebrados de las Aguas Dulces de Galicia; Hércules, Ed.; Hércules Ediciones: A Coruña, Spain, 2006; 175p. [Google Scholar]
- Amat-Trigo, F.; Oliva-Paterna, F.J.; Verdiell-Cubedo, D.; Ruiz-Navarro, A.; Torralva, M. Edad y crecimiento de Gobio lozanoi Doadrio y Madeira, 2004 (Cypriniformes: Cyprinidae) en gradientes longitudinales de la cuenca del río Segura (SE Península Ibérica). An. Biol. 2013, 35, 109–121. [Google Scholar]
- Van Handel, E. Estimation of glycogen in small amounts of tissue. Anal. Biochem. 1965, 11, 256–265. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G.H. Simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elton, C. The Ecology of Invasions by Animals and Plants; Methuen Publisher Ltd.: London, UK, 1958; p. 196. [Google Scholar]
- Hutchinson, G. Concluding remarks. Cold Spring Harb. Symp. Quant. Biol. 1957, 22, 415–427. [Google Scholar] [CrossRef]
- Grinnell, J. Presence and absence of animals. Chron. Univ. Calif. 1928, 30, 429–450. [Google Scholar]
- Hutchinson, G. The paradox of the plankton. Am. Nat. 1961, 95, 137–145. [Google Scholar] [CrossRef] [Green Version]
- Connell, J. Diversity in tropical rain forests and coral reefs. Science 1978, 199, 1302–1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Townsend, C.; Scarsbrook, M.; Dolédec, S. The Intermediate Disturbance Hypothesis, Refugia, and Biodiversity in Streams. Limnol. Oceanogr. 1997, 42, 938–949. [Google Scholar] [CrossRef] [Green Version]
- Adams, S.M.; Ham, K.D.; Greeley, M.S.; LeHew, D.E.; Hinton, R.F.; Saylor, C.F. Downstream gradients in bioindicator responses: Point source contaminant effects on fish health. Can. J. Fish. Aquat. Sci. 1996, 53, 2177–2187. [Google Scholar] [CrossRef]
- Porter, C.; Janz, D.M. Treated municipal sewage discharge affects multiple levels of biological organization in fish. Ecotoxicol. Environ. Saf. 2003, 54, 199–206. [Google Scholar] [CrossRef]
- Speranza, E.D.; Colombo, J.C. Biochemical composition of a dominant detritivorous fish Prochilodus lineatus along pollution gradients in the Paraná-Río de la Plata Basin. J. Fish Biol. 2009, 74, 1226–1244. [Google Scholar] [CrossRef]
- Wood, C.M. Dogmas and controversies in the handling of nitrogenous wastes: Is exogenous ammonia a growth stimulant in fish? J. Exp. Biol. 2004, 207, 2043–2054. [Google Scholar] [CrossRef] [Green Version]
- Kühn, I.; Klotz, S. Urbanization and homogenization—Comparing the floras of urban and rural areas in Germany. Biol. Conserv. 2006, 127, 292–300. [Google Scholar] [CrossRef]
- Cobo, F.; Mera, A.; González, M.A. Análisis químico y valor energético de algunas familias de insectos heterometábolos dulceacuícolas. Boletín AeE 1999, 23, 213–221. [Google Scholar]
- Cobo, F.; Mera, A.; González, M.A. Análisis químico y contenido energético de algunas familias de insectos holometábolos dulceacuícolas. NACC 2000, 10, 1–12. [Google Scholar]
- Shingles, A.; McKenzie, D.J.; Taylor, E.W.; Moretti, A.; Butler, P.J.; Ceradini, S. Effects of sublethal ammonia exposure on swimming performance in rainbow trout (Oncorhynchus mykiss). J. Exp. Biol. 2001, 204, 2691–2698. [Google Scholar] [CrossRef] [PubMed]
- Tudorache, C.; Blust, R.; De Boeck, G. Social interactions, predation behaviour and fast start performance are affected by ammonia exposure in brown trout (Salmo trutta L.). Aquat. Toxicol. 2008, 90, 145–153. [Google Scholar] [CrossRef] [PubMed]
- McKenzie, D.J.; Shingles, A.; Claireaux, G.; Domenici, P. Sublethal concentrations of ammonia impair performance of the teleost fast-start escape response. Physiol. Biochem. Zool. 2009, 82, 353–362. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Pego River | Caselas River |
|---|---|---|
| pH | 5.94 | 6.33 |
| Temperature (°C) | 17.6 | 14.1 |
| Conductivity (μS/cm) | 80.7 | 103.7 |
| TDS (mg/L) | 51.7 | 66.3 |
| Oxygen Saturation (%) | 87.0 | 88.5 |
| Dissolved Oxygen (mg/L) | 8.3 | 9.0 |
| Turbidity (UNT) | 0.50 | 1.19 |
| Ammonium (mg/L) | 0.141 | 0.035 |
| Nitrites NO2− (mg/L) | 0.009 * | 0.003 * |
| Nitrates NO3− (mg/L) | 7.33 | 11.54 |
| Sulphates (mg/l) | 5.45 | 8.73 |
| Orthophosphates (mg/L) | 0.02 * | 0.01 * |
| Chlorides (mg/L) | 13.83 | 19.50 |
| Calcium Ca++ (mg/L) | 2.20 | 3.21 |
| Magnesium Mg++ (mg/L) | 0.97 | 2.19 |
| Hardness (mg/L) | 9.5 | 17 |
| Suspended solids (mg/L) | 10.9 | 20.4 |
| Parameters | Pego River | Caselas River |
|---|---|---|
| Taxonomic richness (S) | 26 | 23 |
| Absolute abundances | 919 | 443 |
| Density (individuals/m2) | 2756 | 1328 |
| Shannon-Wienner Index (H’) | 2.66 | 1.22 |
| Simpson Index (D’) | 0.28 | 0.17 |
| Fisher Index (a) | 4.79 | 2.38 |
| Pielou Index (J’) | 0.58 | 0.27 |
| Biotic Index (IBMWP) | 81 | 120 |
| Species | Pego River (Indiv./m2) | Caselas River (Indiv./m2) |
|---|---|---|
| Gobio lozanoi * | 0.22 | 0.12 |
| Anguilla anguilla | 0.16 | 0.19 |
| Pseudochondrostoma duriense | 0.06 | 0.06 |
| Salmo salar | 0.06 | 0.04 |
| Salmo trutta | 0.2 | 0.11 |
| Petromyzon marinus | 0.02 | 0.02 |
| Achondrostoma arcasii | 0.23 | 0.07 |
| Cobitis paludica | 0.14 | 0.02 |
| Platichthys flesus | 0.010 | 0.002 |
| P. duriense x A. arcasii | 0.001 | 0.005 |
| Gambusia holbrooki * | 0.001 | 0.002 |
| Lepomis gibbosus * | 0.01 | 0.002 |
| Atherina presbyter | ND | 0.0004 |
| River | N | Furcal Length (cm) | Weight (g) | Condition Factor (K) |
|---|---|---|---|---|
| Pego | 288 | 7.30 ± 0.11 (3.4–11.5) | 6.50 ± 0.28 (0.4–21.4) | 1.36 ± 0.01 (0.88–1.98) |
| Caselas | 399 | 8.50 ± 0.08 (4.8–13.2) | 9.30 ± 0.24 (2.0–27.4) | 1.390 ± 0.008 (0.96–1.95) |
| River | N | Glycogen (mg/g) | Lipids (%) | Proteins (mg/g) |
|---|---|---|---|---|
| Pego | 30 | 0.07 ± 0.01 (0.03–0.17) | 5.4 ± 1.1 (1.30–28.90) | 43.95 ± 1.61 (28.88–75.82) |
| Caselas | 30 | 0.080 ± 0.005 (0.05–0.17) | 2.1 ± 0.1 (1.10–2.90) | 58.92 ± 1.92 (40.28–78.62) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barca, S.; Vieira-Lanero, R.; Nachón, D.J.; Sánchez-Hernández, J.; Cobo, M.d.C.; Cobo, F. Complexity of the Relationship between Environmental Factors, Interspecific Competition, and Intrinsic Traits of the Species in Explaining the Invasive Success of Gobio lozanoi Doadrio & Madeira, 2004. Water 2021, 13, 3043. https://doi.org/10.3390/w13213043
Barca S, Vieira-Lanero R, Nachón DJ, Sánchez-Hernández J, Cobo MdC, Cobo F. Complexity of the Relationship between Environmental Factors, Interspecific Competition, and Intrinsic Traits of the Species in Explaining the Invasive Success of Gobio lozanoi Doadrio & Madeira, 2004. Water. 2021; 13(21):3043. https://doi.org/10.3390/w13213043
Chicago/Turabian StyleBarca, Sandra, Rufino Vieira-Lanero, David José Nachón, Javier Sánchez-Hernández, María del Carmen Cobo, and Fernando Cobo. 2021. "Complexity of the Relationship between Environmental Factors, Interspecific Competition, and Intrinsic Traits of the Species in Explaining the Invasive Success of Gobio lozanoi Doadrio & Madeira, 2004" Water 13, no. 21: 3043. https://doi.org/10.3390/w13213043