
Impact of Temperature on the Performance and Character of the Methanogenic Community of a Fixed-Bed Anaerobic Reactor at Psychrophilic Temperature

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bioreactor
2.2. Feeding Solution and Seed Sludge
2.3. Experimental Protocol
2.4. DNA Extraction and Conventional PCR
2.5. Microbial Community Analysis with PCR-DGGE
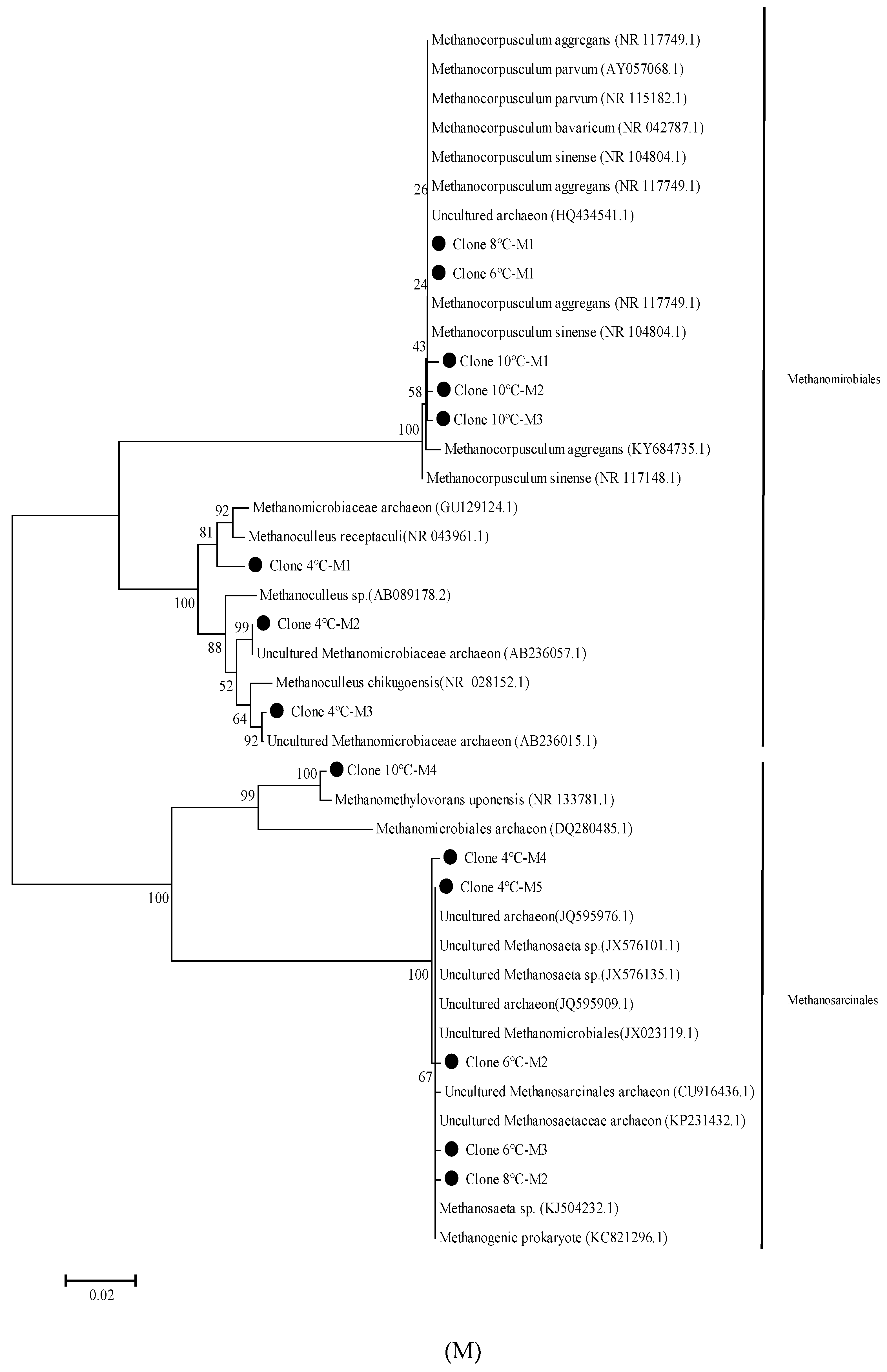
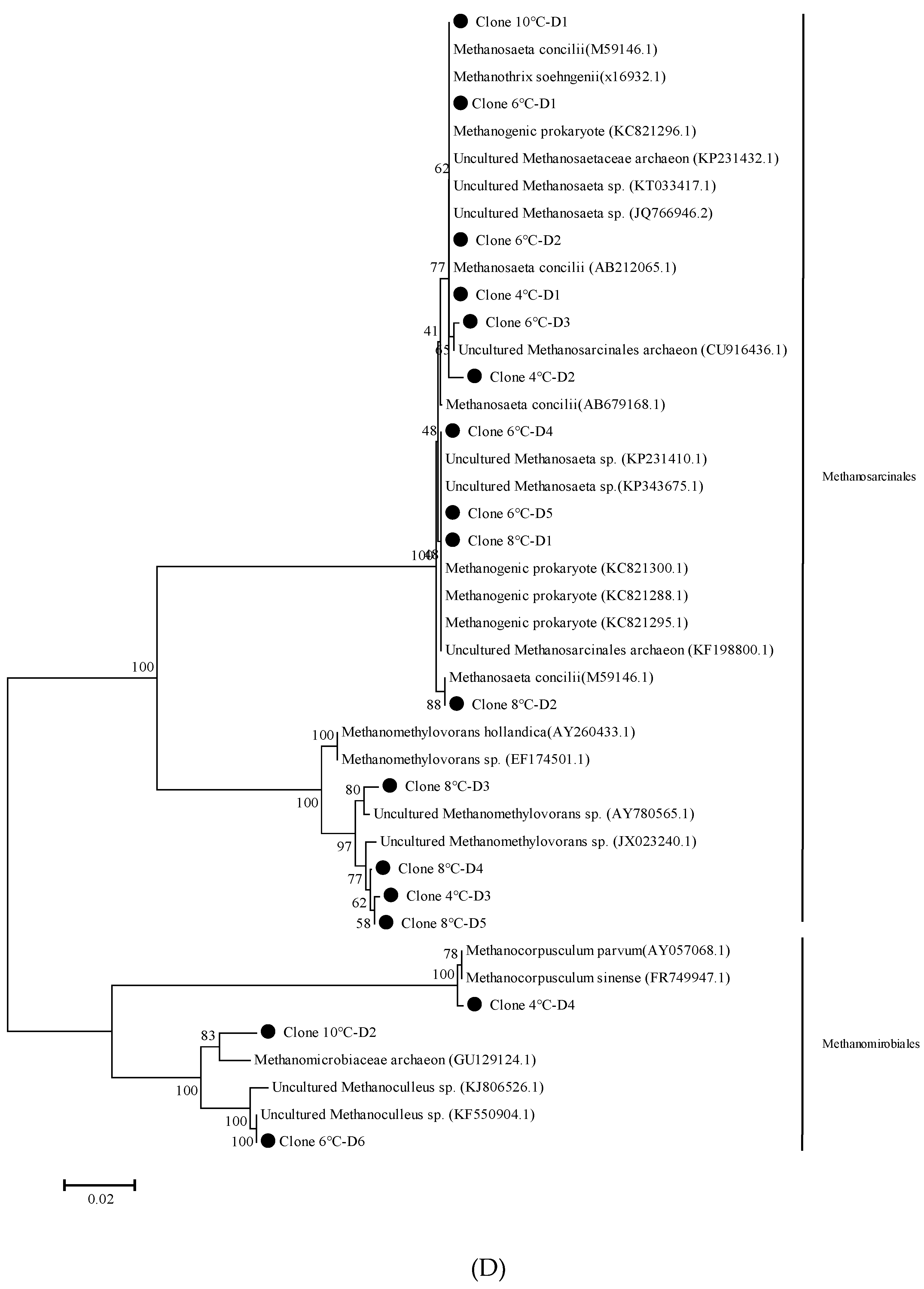
2.6. 16S rRNA Gene Clone Library and Phylogenetic Tree Analysis
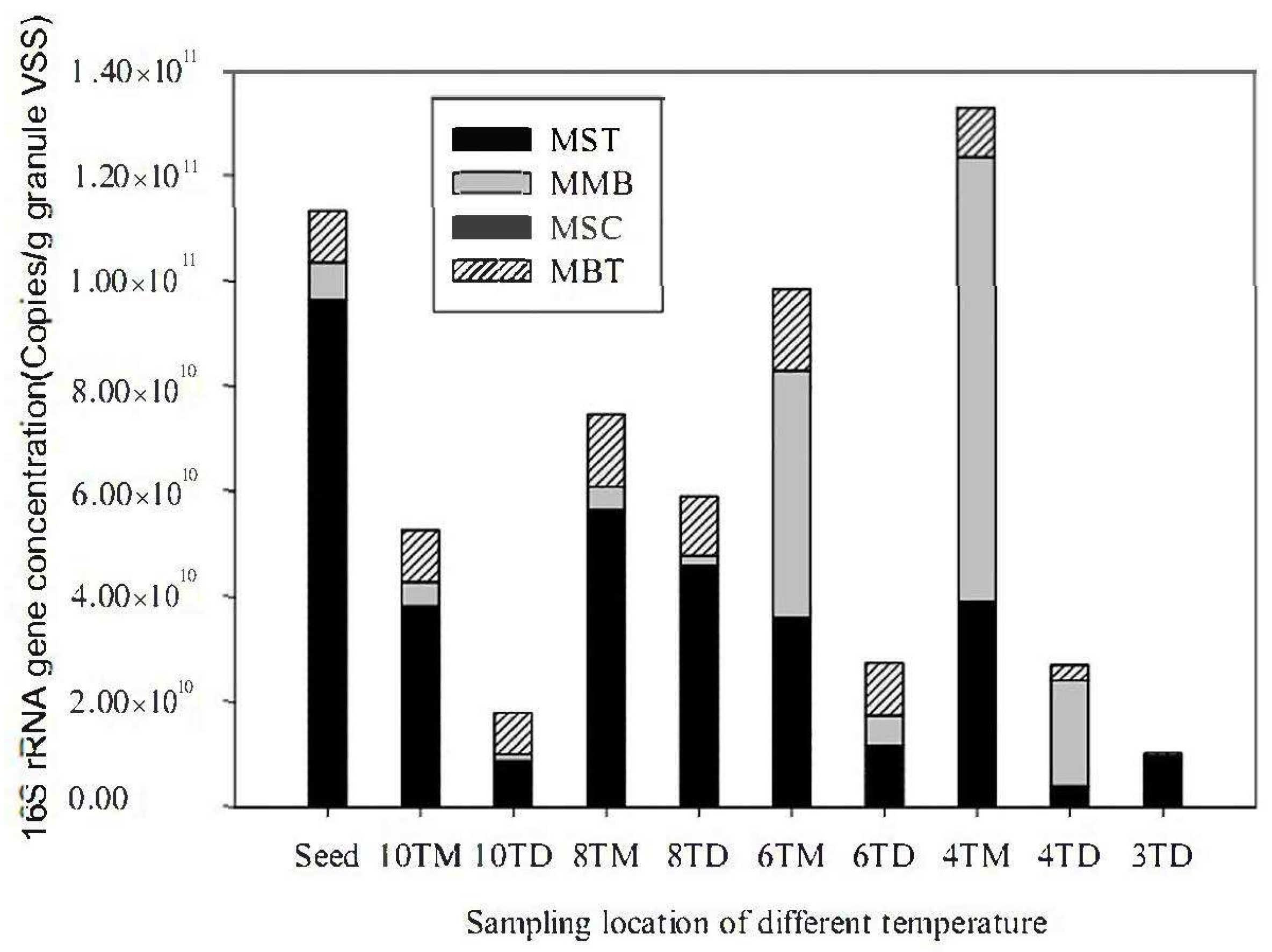
2.7. Quantitative PCR of Methanogens
3. Results and Discussion
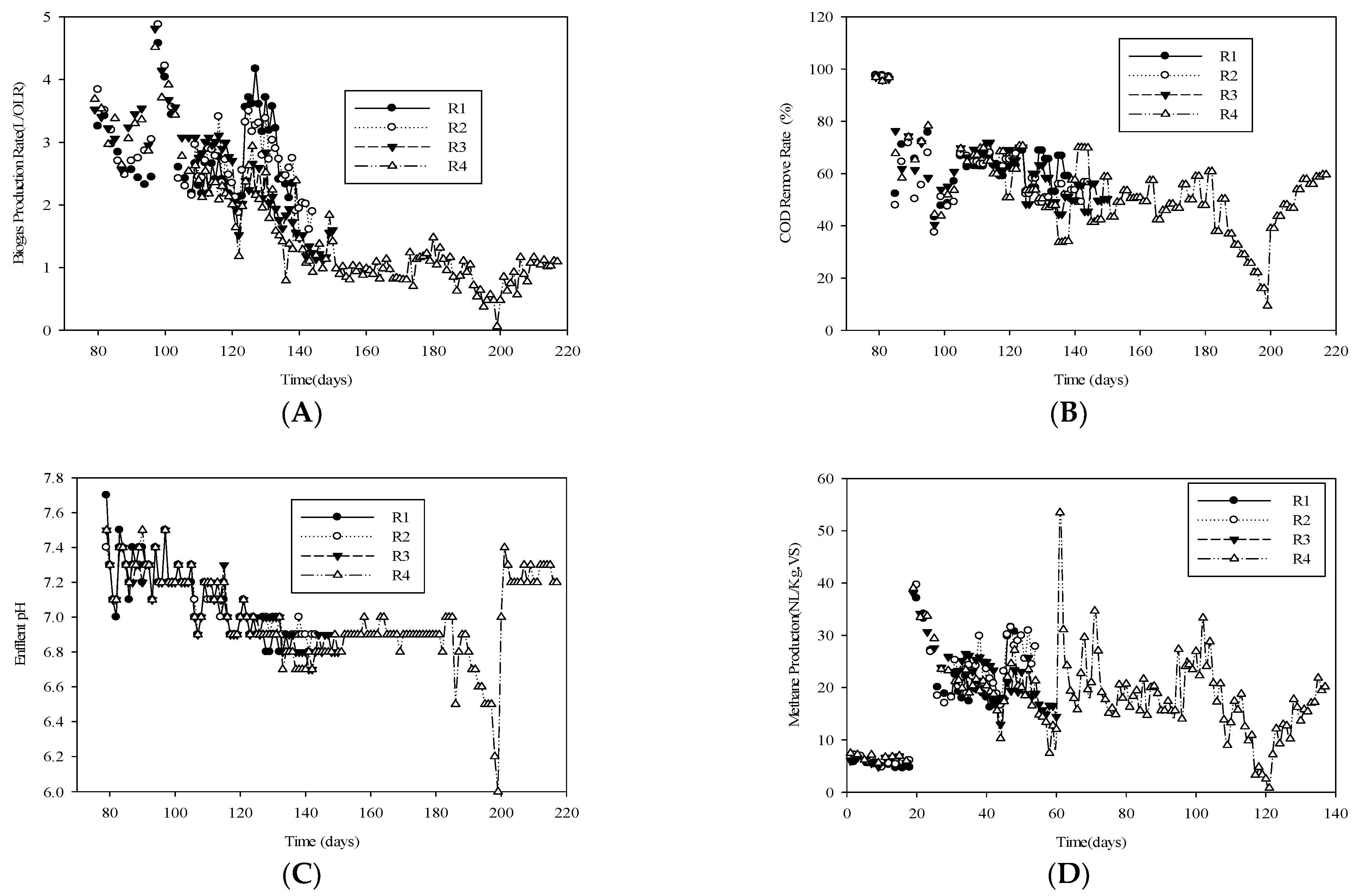
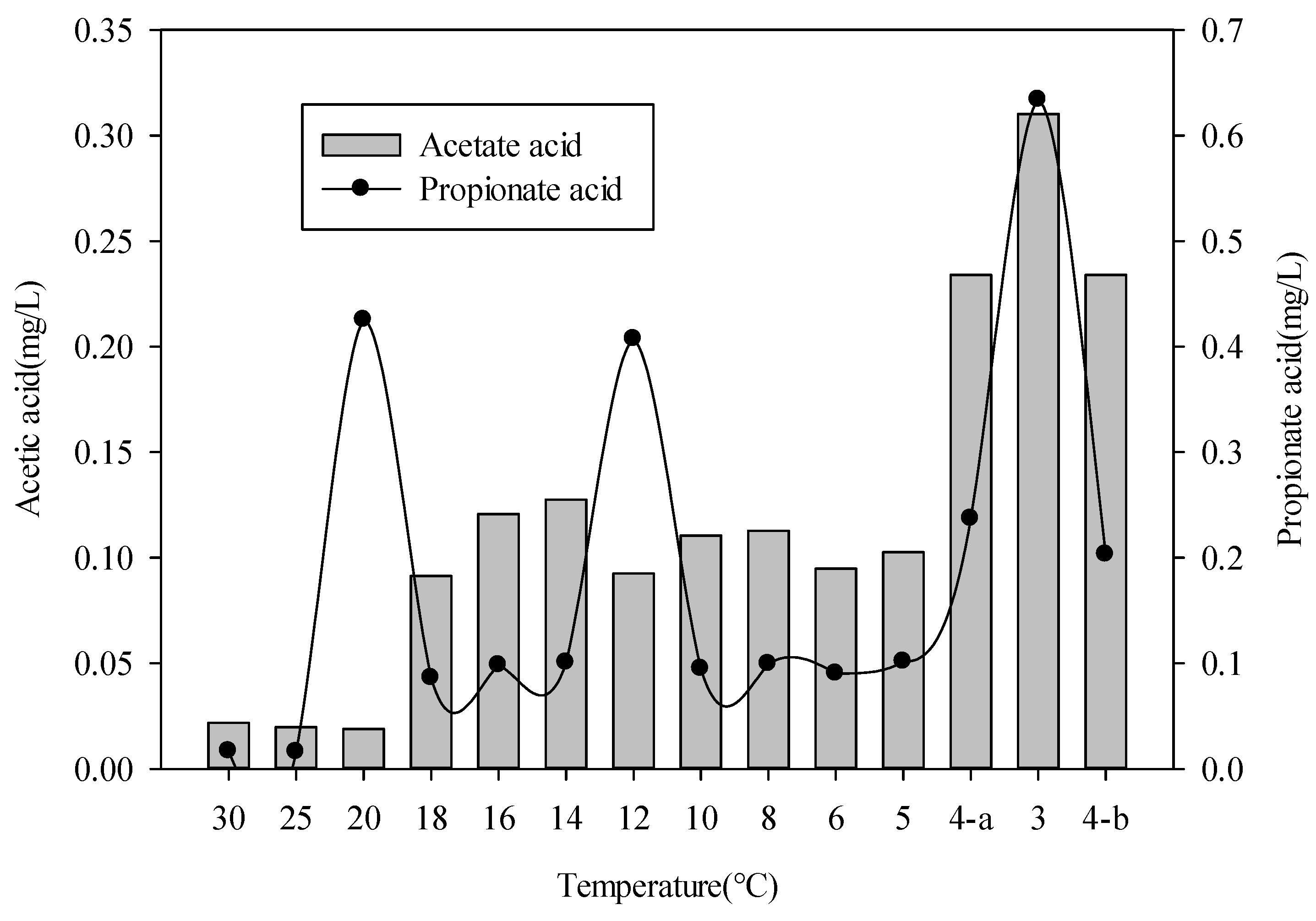
3.1. Bioreactor Performance
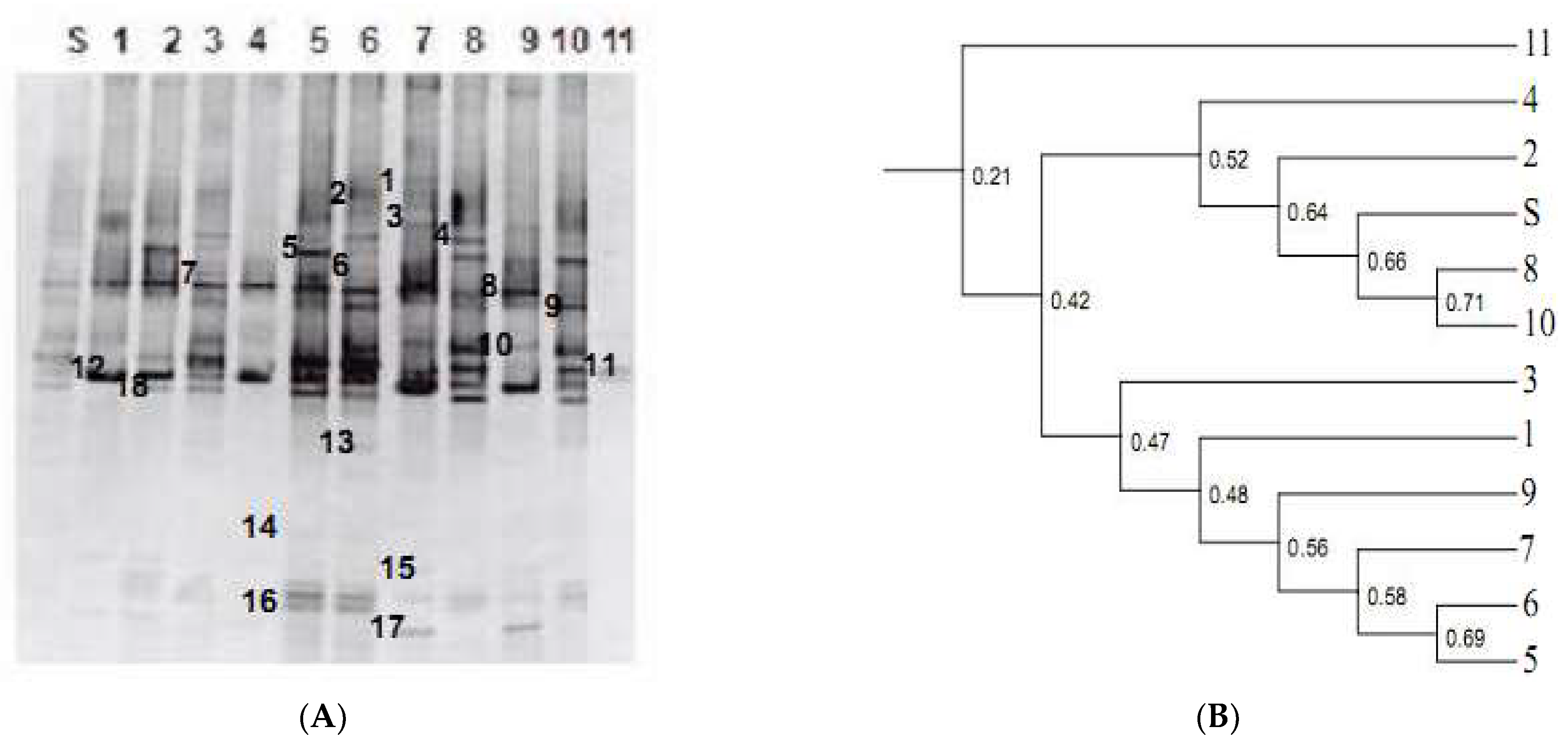
3.2. Archaea DGGE Profiles
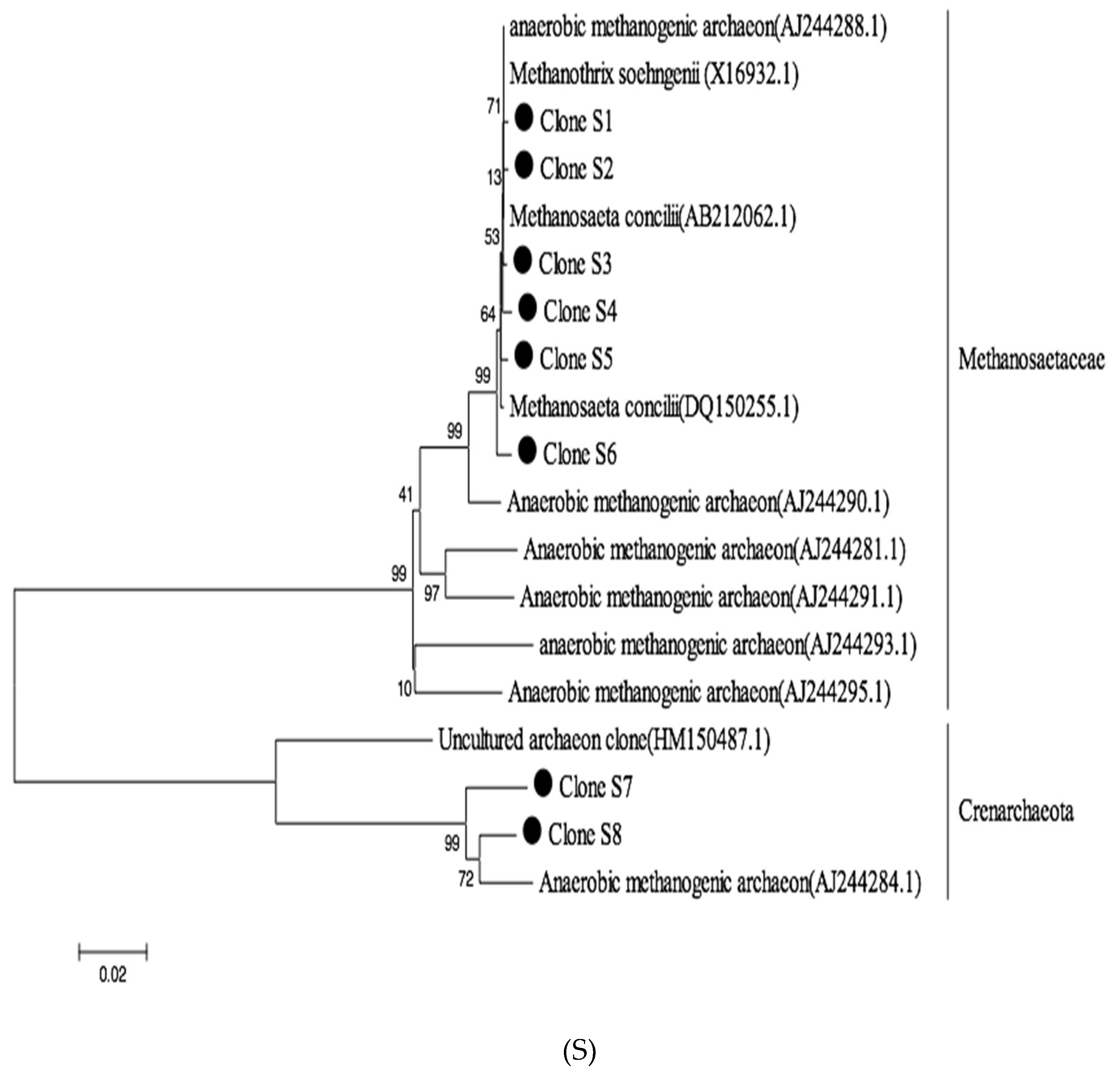
3.3. Methanogenic Community Clone Library Analysis
3.4. Quantitative Analysis of the Methanogenic Community
4. Advantages of Fixed-Bed Reactor Operation at Low Temperature
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Angelidaki, I.; Treu, L.; Tsapekos, P.; Luo, G.; Campanaro, S.; Wenzel, H.; Kougias, P.G. Biogas upgrading and utilization: Current status and perspectives. Biotechnol. Adv. 2018, 36, 452–466. [Google Scholar] [CrossRef] [Green Version]
- Westerholm, M.; Isaksson, S.; Karlsson Lindsjö, O.; Schnürer, A. Microbial community adaptability to altered temperature conditions determines the potential for process optimisation in biogas production. Appl. Energy 2018, 226, 838–848. [Google Scholar] [CrossRef]
- Wu, P.H.; Ng, K.K.; Hong, P.K.A.; Yang, P.Y.; Lin, C.F. Treatment of low-strength wastewater at mesophilic and psychrophilic conditions using immobilized anaerobic biomass. Chem. Eng. J. 2017, 311, 46–54. [Google Scholar] [CrossRef]
- Lin, R.; Cheng, J.; Ding, L.; Murphy, J.D. Improved efficiency of anaerobic digestion through direct interspecies electron transfer at mesophilic and thermophilic temperature ranges. Chem. Eng. J. 2018, 350, 681–691. [Google Scholar] [CrossRef]
- Wu, L.-J.; Higashimori, A.; Qin, Y.; Hojo, T.; Kubota, K.; Li, Y.-Y. Comparison of hyper-thermophilic-mesophilic two-stage with single-stage mesophilic anaerobic digestion of waste activated sludge: Process performance and microbial community analysis. Chem. Eng. J. 2016, 290, 290–301. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, J.; Feng, L.; Zhang, L. Effectiveness research of membrane bioreactor process treating low-temperature wastewater. Chin. J. Environ. Eng. 2011, 5, 1263–1267. [Google Scholar]
- Smith, A.L.; Skerlos, S.J.; Raskin, L. Anaerobic membrane bioreactor treatment of domestic wastewater at psychrophilic temperatures ranging from 15 °C to 3 °C. Environ. Sci. Water Res. Technol. 2015, 1, 56–64. [Google Scholar] [CrossRef]
- Lettinga, G.; Rebac, S.; Zeeman, G. Challenge of psychrophilic anaerobic wastewater treatment. Trends Biotechnol. 2001, 19, 363–370. [Google Scholar] [CrossRef]
- Petropoulos, E.; Dolfing, J.; Davenport, R.J.; Bowen, E.J.; Curtis, T.P. Developing cold-adapted biomass for the anaerobic treatment of domestic wastewater at low temperatures (4, 8 and 15 °C) with inocula from cold environments. Water Res. 2017, 112, 100–109. [Google Scholar] [CrossRef]
- Deng, L.; Yang, H.; Liu, G.; Zheng, D.; Chen, Z.; Liu, Y.; Pu, X.; Song, L.; Wang, Z.; Lei, Y. Kinetics of temperature effects and its significance to the heating strategy for anaerobic digestion of swine wastewater. Appl. Energy 2014, 134, 349–355. [Google Scholar] [CrossRef]
- McKeown, R.M.; Scully, C.; Mahony, T.; Collins, G.; O’Flaherty, V. Long-term (1243 days), low-temperature (4–15 °C), anaerobic biotreatment of acidified wastewaters: Bioprocess performance and physiological characteristics. Water Res. 2009, 43, 1611–1620. [Google Scholar] [CrossRef]
- Siggins, A.; Enright, A.M.; O’Flaherty, V. Low-temperature (7 degrees C) anaerobic treatment of a trichloroethylene-contaminated wastewater: Microbial community development. Water Res. 2011, 45, 4035–4046. [Google Scholar] [CrossRef]
- Smith, K.; Liu, S.; Liu, Y.; Guo, S. Can China reduce energy for water? A review of energy for urban water supply and wastewater treatment and suggestions for change. Renew. Sustain. Energy Rev. 2018, 91, 41–58. [Google Scholar] [CrossRef]
- Zhao, H.; Yan, F.; Li, X.; Piao, R.; Wang, W.; Cui, Z. Impact of Organic Loading Rate on Performance and Methanogenic Microbial Communities of a Fixed-Bed Anaerobic Reactor at 4 °C. Water 2020, 12, 2586. [Google Scholar] [CrossRef]
- Connaughton, S.; Collins, G.; O’Flaherty, V. Development of microbial community structure and actvity in a high-rate anaerobic bioreactor at 18 °C. Water Res. 2006, 40, 1009–1017. [Google Scholar] [CrossRef]
- Raheem, A.; Sikarwar, V.S.; He, J.; Dastyar, W.; Dionysiou, D.D.; Wang, W.; Zhao, M. Opportunities and challenges in sustainable treatment and resource reuse of sewage sludge: A review. Chem. Eng. J. 2018, 337, 616–641. [Google Scholar] [CrossRef]
- Yurtsever, A.; Calimlioglu, B.; Sahinkaya, E. Impact of SRT on the efficiency and microbial community of sequential anaerobic and aerobic membrane bioreactors for the treatment of textile industry wastewater. Chem. Eng. J. 2017, 314, 378–387. [Google Scholar] [CrossRef]
- Zhang, D.; Zhu, W.; Tang, C.; Suo, Y.; Gao, L.; Yuan, X.; Wang, X.; Cui, Z. Bioreactor performance and methanogenic population dynamics in a low-temperature (5–18 °C) anaerobic fixed-bed reactor. Bioresour. Technol. 2012, 104, 136–143. [Google Scholar] [CrossRef]
- Kuroda, K.C.; Nakahara, T.; Hatamoto, N.; Wakabayashi, M.; Kawai, T.; Araki, T.; Syutsubo, N.; Yamaguchi, K.T. High organic loading treatment for industrial molasses wastewater and microbial community shifts corresponding to system development. Bioresour. Technol. 2015, 196, 225–234. [Google Scholar] [CrossRef]
- Li, L.; Yuan, Z.; Sun, Y.; Kong, X.; Dong, P.; Zhang, J. A reused method for molasses-processed wastewater: Effect on silage quality and anaerobic digestion performance of Pennisetum purpereum. Bioresour. Technol. 2017, 241, 1003–1011. [Google Scholar] [CrossRef]
- Meng, X.Y.; Yuan, X.; Ren, J.; Wang, X.; Zhu, W.; Cui, Z. Methane production and characteristics of the microbial community in a two-stage fixed-bed anaerobic reactor using molasses. Bioresour. Technol. 2017, 241, 1050–1059. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Li, J.; Guo, P.; Li, P.; Suo, Y.; Wang, X.; Cui, Z. Dynamic transition of microbial communities in response to acidification in fixed-bed anaerobic baffled reactors (FABR) of two different flow directions. Bioresour. Technol. 2011, 102, 4703–4711. [Google Scholar] [CrossRef] [PubMed]
- Lew, B.; Tarre, S.; Beliavski, M.; Green, M. Anaerobic degradation pathway and kinetics of domestic wastewater at low temperatures. Bioresour. Technol. 2009, 100, 6155–6162. [Google Scholar] [CrossRef]
- Sasaki, K.; Morita, M.; Hirano, S.; Ohmura, N.; Igarashi, Y. Effect of adding carbon fiber textiles to methanogenic bioreactors used to treat an artificial garbage slurry. J. Biosci. Bioeng. 2009, 108, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Salvador, M.; Haruta, S.; Hazaka, M.; Shimada, R.; Yoshida, C.; Hiura, K.; Ishii, M.; Igarashi, Y. Denaturing gradient gel electrophoresis analyses of microbial community from field-scale composter. J. Biosci. Bioeng. 2001, 91, 159–165. [Google Scholar] [CrossRef]
- Lueders, T.; Friedrich, M.W. Effects of Amendment with Ferrihydrite and Gypsum on the Structure and Activity of Methanogenic Populations in Rice Field Soil. Appl. Environ. Microbiol. 2002, 68, 2484–2494. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.; Yang, H.; Wang, X.; Huang, Z.; Ishii, M.; Igarashi, Y.; Cui, Z. Rice straw fermentation using lactic acid bacteria. Bioresour. Technol. 2008, 99, 2742–2748. [Google Scholar] [CrossRef]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef]
- Yu, Y.; Lee, C.; Kim, J.; Hwang, S. Group-specific primer and probe sets to detect methanogenic communities using quantitative real-time polymerase chain reaction. Biotechnol. Bioeng. 2005, 89, 670–679. [Google Scholar] [CrossRef]
- Zhao, H.; Li, J.; Li, J.; Yuan, X.; Piao, R.; Zhu, W.; Li, H.; Wang, X.; Cui, Z. Organic loading rate shock impact on operation and microbial communities in different anaerobic fixed-bed reactors. Bioresour. Technol. 2013, 140, 211–219. [Google Scholar] [CrossRef]
- Bandara, W.M.K.R.T.W.; Kindaichi, T.; Satoh, H.; Sasakawa, M.; Nakahara, Y.; Takahashi, M.; Okabe, S. Anaerobic treatment of municipal wastewater at ambient temperature: Analysis of archaeal community structure and recovery of dissolved methane. Water Res. 2012, 46, 5756–5764. [Google Scholar] [CrossRef] [Green Version]
- Collins, G.; Foy, C.; McHugh, S.; Mahony, T.; O’Flaherty, V. Anaerobic biological treatment of phenolic wastewater at 15–18 °C. Water Res. 2005, 39, 1614–1620. [Google Scholar] [CrossRef]
- Kashyap, D.R.; Dadhich, K.S.; Sharma, S.K. Biomethanation under psychrophilic conditions: A review. Bioresour. Technol. 2003, 87, 147–153. [Google Scholar] [CrossRef]
- Dhaked, R.K.; Singh, P.; Singh, L. Biomethanation under psychrophilic conditions. Waste Manag. 2010, 30, 2490–2496. [Google Scholar] [CrossRef]
- O’Reilly, J.; Lee, C.; Collins, G.; Chinalia, F.; Mahony, T.; O’Flaherty, V. Quantitative and qualitative analysis of methanogenic communities in mesophilically and psychrophilically cultivated anaerobic granular biofilims. Water Res. 2009, 43, 3365–3374. [Google Scholar] [CrossRef] [PubMed]
- Kalyuzhnyl, S.V.; Gladchenko, M.A.; Skylar, V.I.; Kizimenko, Y.S.; Shcherbakov, S.S. One- and Two-Stage Upflow Anaerobic Sludge-Bed Reactor Pretreatment of Winery Wastewater at 4–10 °C. Appl. Biochem. Biotechnol. 2001, 90, 107–124. [Google Scholar] [CrossRef]
- Lettinga, G.; Rebic, S.; Parshina, S.; Nozhevnikoa, A.; Vanlier, J.; Stams, A. High-Rate Anaerobic Treatment of Wastewater at Low Temperatures. Appl. Environ. Microbiol. 1999, 65, 1696–1702. [Google Scholar] [CrossRef] [Green Version]
- McKeown, R.M.; Hughes, D.; Collins, G.; Mahony, T.; O’Flaherty, V. Low-temperature anaerobic digestion for wastewater treatment. Curr. Opin. Biotechnol. 2012, 23, 444–451. [Google Scholar] [CrossRef] [PubMed]
- Parshina, S.N.; Ermakova, A.V.; Bomberg, M.; Detkova, E.N. Methanospirillum stamsii sp. nov., a psychrotolerant, hydrogenotrophic, methanogenic archaeon isolated from an anaerobic expanded granular sludge bed bioreactor operated at low temperature. Int. J. Syst. Evol. 2014, 64, 180–186. [Google Scholar] [CrossRef] [Green Version]
- Sekiguchi, Y.; Kamagata, Y. Microbial community structure and functions in methane fermentation technology for wastewater treatment. Strict and facultative anaerobes: Medical and environmental aspects. BioScience 2004, 361–384. [Google Scholar]
- McKeown, R.M.; Scully, C.; Enright, A.M.; Chinalia, F.A.; Lee, C.; Mahony, T.; Collins, G.; O’Flaherty, V. Psychrophilic methanogenic community development during long-term cultivation of anaerobic granular biofilms. ISME J. 2009, 3, 1231–1242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshinaga, M.Y.; Holler, T.; Goldhammer, T.; Wegener, G.; Pohlman, J.W.; Brunner, B.; Kuypers, M.; Hinrichs, K.; Elvert, M. Carbon isotope equilibration during sulphate-limited anaerobic oxidation of methane. Nat. Geosci. 2014, 7, 190–194. [Google Scholar] [CrossRef]
- Liu, Y.F.; Chen, Y.W.; Li, T.; Wang, D.G.; Wang, D.K. Investigation on the heat loss characteristic of underground household biogas digester using dynamic simulations and experiments. Biosyst. Eng. 2017, 163, 116–133. [Google Scholar] [CrossRef]
- Smith, A.L.; Skerlos, S.J.; Raskin, L. Psychrophilic anaerobic membrane bioreactor treatment of domestic wastewater. Water Res. 2013, 47, 1655–1665. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Operation Stage | T (°C) | HRT | Operation Time (Day) | Molasses Add (g/d) | Influent COD (mg/L) | OLR (kg COD/m3·d) |
|---|---|---|---|---|---|---|
| Start-up | 35 ± 0.2 | 13 | 1–78 | 102.56 ± 0.1 | 20,000 ± 1 | 6.67 ± 0.1 |
| Decrease state | 30 ± 0.2 | 2 | 79–85 | 102.56 ± 0.1 | 20,000 ± 1 | 6.67 ± 0.1 |
| 25 ± 0.2 | 2 | 86–91 | 76.92 ± 0.1 | 15,000 ± 1 | 5.00 ± 0.1 | |
| 20 ± 0.2 | 2 | 92–97 | 51.28 ± 0.1 | 10,000 ± 1 | 3.33 ± 0.1 | |
| 18 ± 0.2 | 2 | 98–103 | 25.64 ± 0.1 | 5000 ± 1 | 1.67 ± 0.1 | |
| 16 ± 0.2 | 2 | 104–109 | 25.64 ± 0.1 | 5000 ± 1 | 1.67 ± 0.1 | |
| 14 ± 0.2 | 2 | 110–115 | 25.64 ± 0.1 | 5000 ± 1 | 1.67 ± 0.1 | |
| 12 ± 0.2 | 2 | 116–121 | 25.64 ± 0.1 | 5000 ± 1 | 1.67 ± 0.1 | |
| 10 ± 0.2 | 2 | 122–128 | 25.64 ± 0.1 | 5000 ± 1 | 1.67 ± 0.1 | |
| 8 ± 0.2 | 2 | 129–134 | 25.64 ± 0.1 | 5000 ± 1 | 1.67 ± 0.1 | |
| 6 ± 0.2 | 2 | 135–140 | 25.64 ± 0.1 | 5000 ± 1 | 1.67 ± 0.1 | |
| 5 ± 0.2 | 4 | 141–152 | 10.26 ± 0.1 | 2000 ± 1 | 0.67 ± 0.1 | |
| 4 ± 0.2 | 10 | 153–182 | 10.26 ± 0.1 | 2000 ± 1 | 0.67 ± 0.1 | |
| Source state | 3 ± 0.2 | 6 | 183–200 | 10.26 ± 0.1 | 2000 ± 1 | 0.67 ± 0.1 |
| Recovery state | 4 ± 0.2 | 6 | 201–217 | 10.26 ± 0.1 | 2000 ± 1 | 0.67 ± 0.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, H.; Liu, H.; Sun, Y.; Zhang, L.; Fazl, U.; Piao, R.; Wang, W.; Cui, Z. Impact of Temperature on the Performance and Character of the Methanogenic Community of a Fixed-Bed Anaerobic Reactor at Psychrophilic Temperature. Water 2021, 13, 3051. https://doi.org/10.3390/w13213051
Zhao H, Liu H, Sun Y, Zhang L, Fazl U, Piao R, Wang W, Cui Z. Impact of Temperature on the Performance and Character of the Methanogenic Community of a Fixed-Bed Anaerobic Reactor at Psychrophilic Temperature. Water. 2021; 13(21):3051. https://doi.org/10.3390/w13213051
Chicago/Turabian StyleZhao, Hongyan, Hongdou Liu, Yu Sun, Liqiang Zhang, Ullah Fazl, Renzhe Piao, Weidong Wang, and Zongjun Cui. 2021. "Impact of Temperature on the Performance and Character of the Methanogenic Community of a Fixed-Bed Anaerobic Reactor at Psychrophilic Temperature" Water 13, no. 21: 3051. https://doi.org/10.3390/w13213051