
L-Cysteine Synthase Enhanced Sulfide Biotransformation in Subtropical Marine Mangrove Sediments as Revealed by Metagenomics Analysis


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
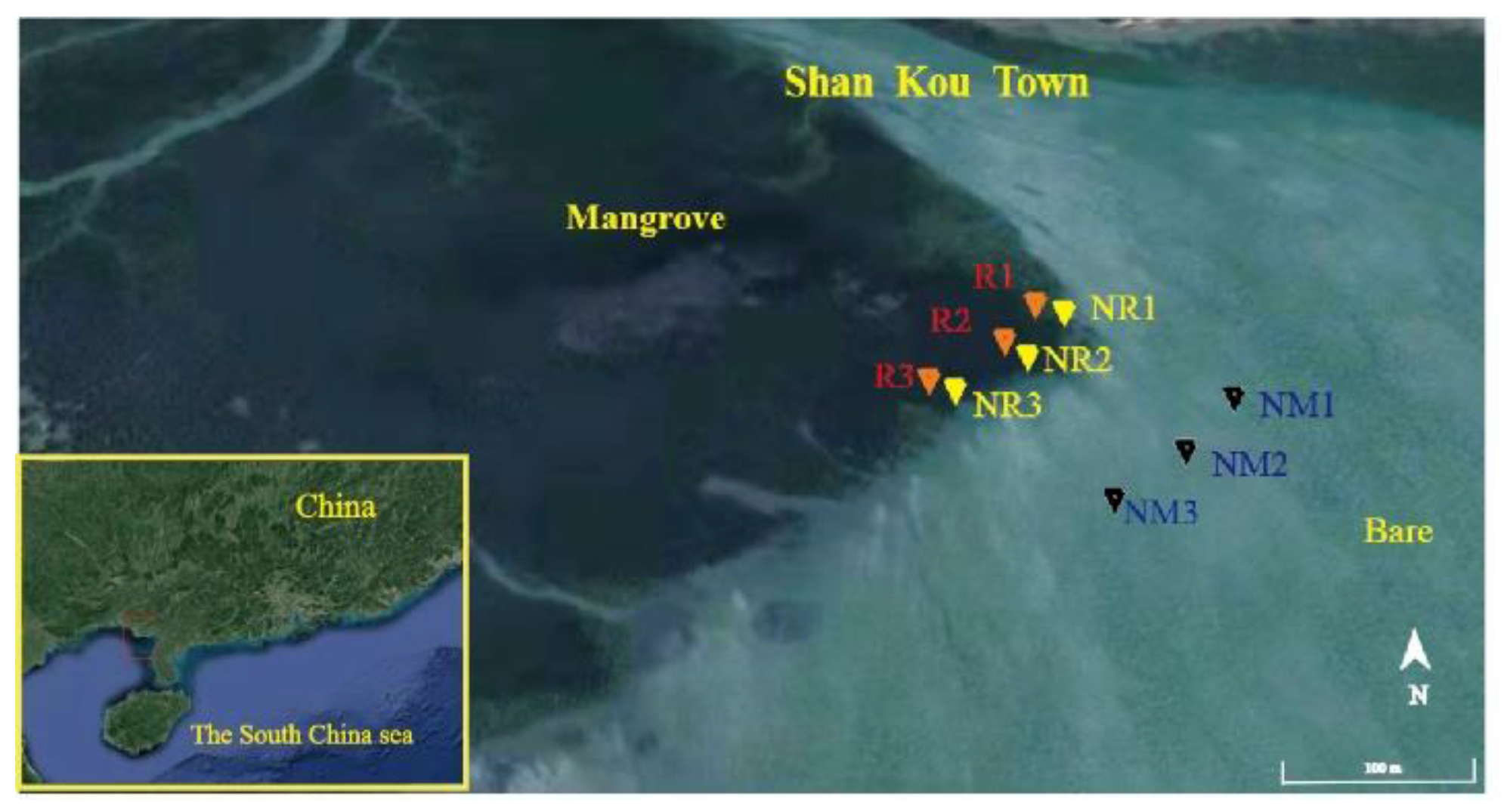
2.1. Sampling Sites and Sediment Collection
2.2. DNA Extraction and High-Throughput Sequencing
2.3. Shotgun Metagenomic Sequence Processing and Analysis
2.4. Quantification of Dissimilatory Sulfite Reductase (dsrB) and Adenylyl Sulfate Reductase (aprA) Gene Copy Numbers
3. Results
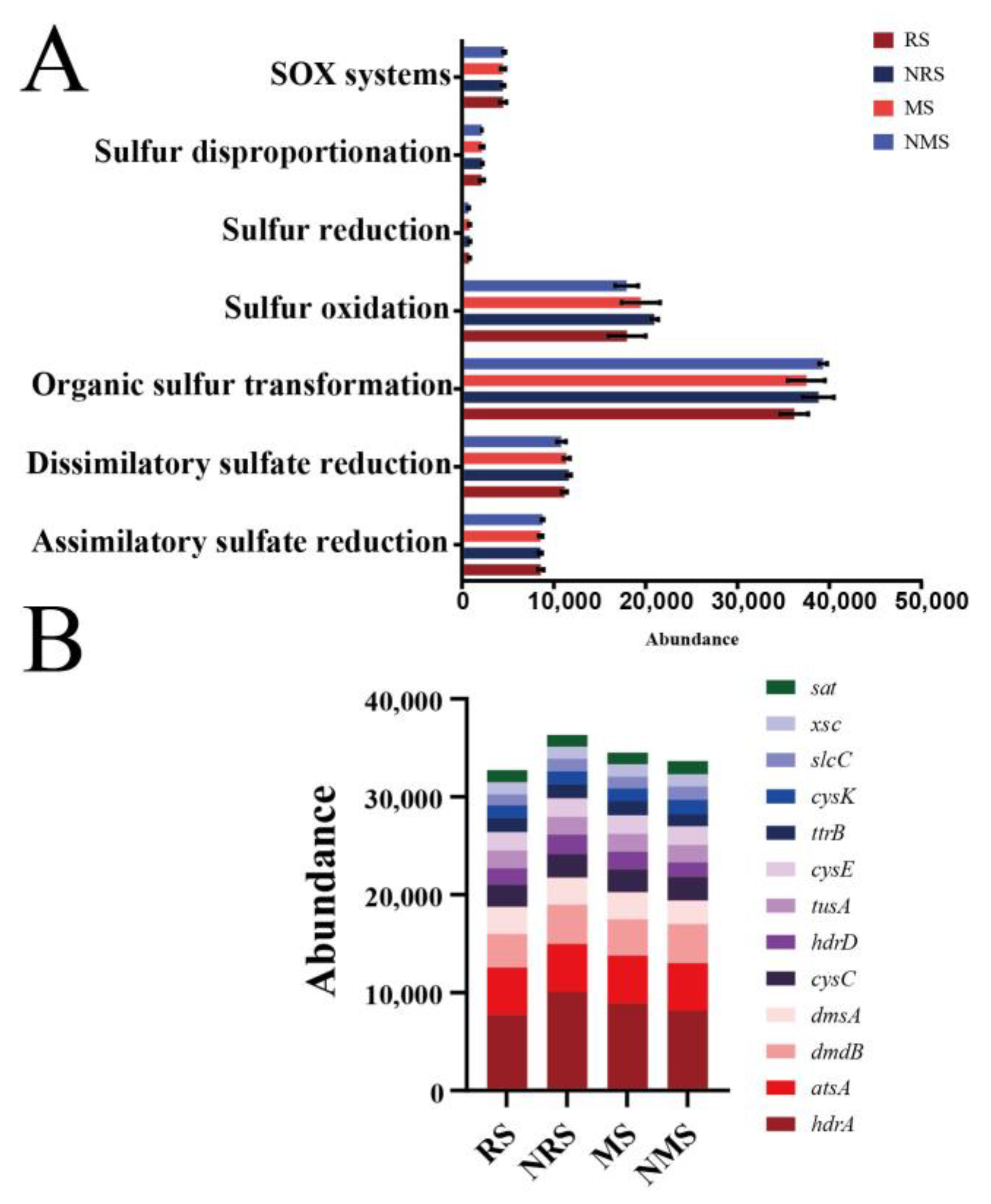
3.1. Abundance and Diversity of Sulfur (sub)Gene Families
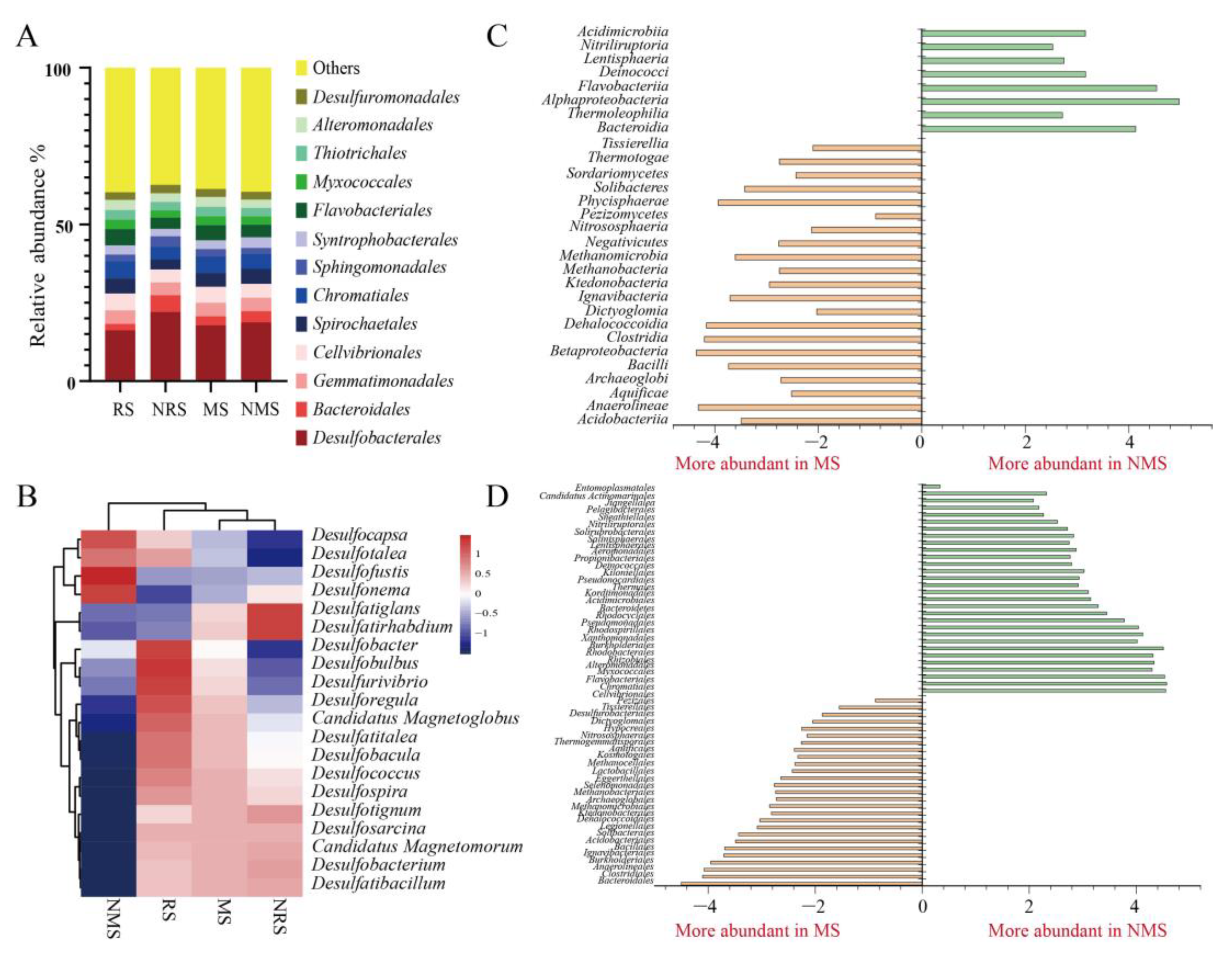
3.2. Microbial Diversity Based on Metagenomics
3.3. Genes for the Dissimilatory Sulfate Reduction
3.4. Taxonomic Assignment of Dissimilatory Sulfate-Reducing Genes
3.5. Quantification of dsrB and aprA Genes
3.6. Sediment Properties
4. Discussion
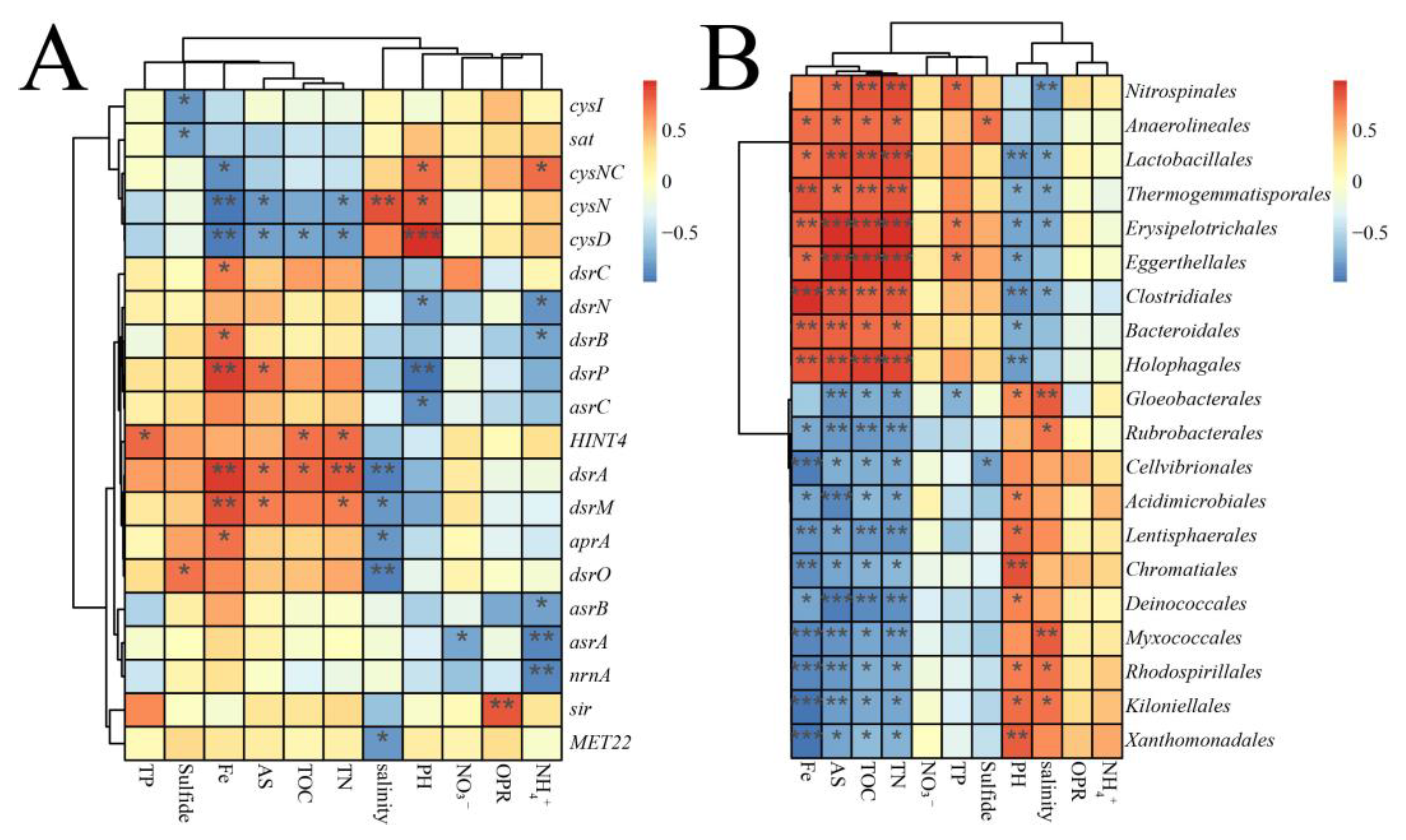
4.1. Sulfur-Cycling Genes in the Mangrove Ecosystem
4.2. Microorganisms Involved in the Sulfide Conversion
4.3. Major Environmental Factors Affecting the Sulfate Reduction
4.4. Mechanism of the Sulfide Conversion in the Mangrove Ecosystem
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rossi, G.S.; Tunnah, L.; Martin, K.E.; Turko, A.J.; Taylor, D.S.; Currie, S.; Wright, P.A. Mangrove Fishes Rely on Emersion Behavior and Physiological Tolerance to Persist in Sulfidic Environments. Physiol. Biochem. Zool. 2019, 92, 316–325. [Google Scholar] [CrossRef]
- Wiig, J.A.; Hu, Y.L.; Ribbe, M.W. Refining the pathway of carbide insertion into the nitrogenase M-cluster. Nat. Commun. 2015, 6, 8034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganigue, R.; Gutierrez, O.; Rootsey, R.; Yuan, Z.G. Chemical dosing for sulfide control in Australia: An industry survey. Water Res. 2011, 45, 6564–6574. [Google Scholar] [CrossRef] [PubMed]
- Gros, O.; Bisque, L.; Sadjan, M.; Azede, C.; Jean-Louis, P.; Guidi-Rontani, C. First description of a new uncultured purple sulfur bacterium colonizing marine mangrove sediment in the Caribbean: Halochromatium-like PSB from Guadeloupe. C. R. Biol. 2018, 341, 387–397. [Google Scholar] [CrossRef]
- Yang, W.; Vollertsen, J.; Hvitved-Jacobsen, T. Anoxic sulfide oxidation in wastewater of sewer networks. Water Sci. Technol. 2005, 52, 191–199. [Google Scholar] [CrossRef]
- Simpson, A.G.; Tripp, L.; Shull, D.H.; Yang, S.J.; Science, S. Effects of Zostera marina roots and leaf detritus on the concentration and distribution of pore-water sulfide in marine sediments. Estuar. Coast. Shelf Sci. 2018, 209, 160–168. [Google Scholar] [CrossRef]
- Ferreira, T.O.; Otero, X.L.; Vidal-Torrado, P.; Macias, F. Effects of bioturbation by root and crab activity on iron and sulfur biogeochemistry in mangrove substrate. Geoderma 2007, 142, 36–46. [Google Scholar] [CrossRef]
- Bukhtiyarova, P.A.; Antsiferov, D.V.; Brasseur, G.; Avakyan, M.R.; Frank, Y.A.; Ikkert, O.P.; Pimenov, N.V.; Tuovinen, O.H.; Karnachuk, O.V. Isolation, characterization, and genome insights into an anaerobic sulfidogenic Tissierella bacterium from Cu-bearing coins. Anaerobe 2019, 56, 66–77. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Liu, H.; Wang, P.; Zhang, D.; Sun, Y.; Li, E. Oxygen Reduction Reaction Affected by Sulfate-Reducing Bacteria: Different Roles of Bacterial Cells and Metabolites. Indian J. Microbiol. 2017, 57, 344–350. [Google Scholar] [CrossRef] [PubMed]
- Dolla, A.; Fournier, M.; Dermoun, Z. Oxygen defense in sulfate-reducing bacteria. J. Biotechnol. 2006, 126, 87–100. [Google Scholar] [CrossRef]
- Ferreira, T.O.; Otero, X.L.; de Souza, V.S.; Vidal-Torrado, P.; Macias, F.; Firme, L.P. Spatial patterns of soil attributes and components in a mangrove system in Southeast Brazil (Sao Paulo). J. Soils Sediments 2010, 10, 995–1006. [Google Scholar] [CrossRef]
- Cochrane, P.V.; Rossi, G.S.; Tunnah, L.; Jonz, M.G.; Wright, P.A. Hydrogen sulphide toxicity and the importance of amphibious behaviour in a mangrove fish inhabiting sulphide-rich habitats. J. Comp. Physiol. B 2019, 189, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Kojima, H.; Takano, Y.; Fukui, M. Diversity of sulfur-cycle prokaryotes in freshwater lake sediments investigated using aprA as the functional marker gene. Syst. Appl. Microbiol. 2013, 36, 436–443. [Google Scholar] [CrossRef]
- Wu, S.; Li, R.; Xie, S.; Shi, C. Depth-related change of sulfate-reducing bacteria community in mangrove sediments: The influence of heavy metal contamination. Mar. Pollut. Bull. 2019, 140, 443–450. [Google Scholar] [CrossRef]
- Spring, S.; Sorokin, D.Y.; Verbarg, S.; Rohde, M.; Woyke, T.; Kyrpides, N.C. Sulfate-Reducing Bacteria That Produce Exopolymers Thrive in the Calcifying Zone of a Hypersaline Cyanobacterial Mat. Front. Microbiol. 2019, 10, 862. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.; Li, X.; Wang, C.; Cao, H.; Cui, Z. Metagenome complexity and template length are the main causes of bias in PCR-based bacteria community analysis. J. Basic Microbiol. 2018, 58, 987–997. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.L.; Hetharua, B.; Lin, L.; Xu, H.; Zheng, T.L.; He, Z.L.; Tian, Y. Mangrove Sediment Microbiome: Adaptive Microbial Assemblages and Their Routed Biogeochemical Processes in Yunxiao Mangrove National Nature Reserve, China. Microb. Ecol. 2019, 78, 57–69. [Google Scholar] [CrossRef]
- Wang, W.L.; Xu, S.Y.; Ren, Z.G.; Tao, L.; Jiang, J.W.; Zheng, S.S. Application of metagenomics in the human gut microbiome. World J. Gastroenterol. 2015, 21, 803–814. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.L.; Zhou, J.Y.; Song, W.; Xu, M.Z.; He, Q.; Peng, Y.S.; Tian, Y.; Wang, C.; Shu, L.F.; Wang, S.Q.; et al. SCycDB: A curated functional gene database for metagenomic profiling of sulphur cycling pathways. Mol. Ecol. Resour. 2020, 21, 924–940. [Google Scholar] [CrossRef]
- Fu, X.; Li, C.; Zhou, X.; Liu, S.; Wu, F. Physiological response and sulfur metabolism of the V. dahliae-infected tomato plants in tomato/potato onion companion cropping. Sci. Rep. 2016, 6, 36445. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Chen, Q.Q.; Zhang, G.R. Determination of Sulfate in Coastal Salt Marsh Sediments with High-Chloride Concentration by Ion Chromatography: A Revised Method. Instrum. Sci. Technol. 2013, 41, 37–47. [Google Scholar] [CrossRef]
- Guenther, E.A.; Johnson, K.S.; Coale, K.H. Direct ultraviolet spectrophotometric determination of total sulfide and iodide in natural waters. Anal. Chem. 2001, 73, 3481–3487. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.T.; Liu, J.; Chen, W.C.; Chen, X.; Shu, H.Y.; Jia, P.; Liao, B.; Shu, W.S.; Li, J.T. Changes in microbial community composition following phytostabilization of an extremely acidic Cu mine tailings. Soil Biol. Biochem. 2017, 114, 52–58. [Google Scholar] [CrossRef]
- Li, D.; Luo, R.; Liu, C.M.; Leung, C.M.; Ting, H.F.; Sadakane, K.; Yamashita, H.; Lam, T.W. MEGAHIT v1.0: A fast and scalable metagenome assembler driven by advanced methodologies and community practices. Methods 2016, 102, 3–11. [Google Scholar] [CrossRef]
- Li, R.Q.; Yu, C.; Li, Y.R.; Lam, T.W.; Yiu, S.M.; Kristiansen, K.; Wang, J. SOAP2: An improved ultrafast tool for short read alignment. Bioinformatics 2009, 25, 1966–1967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villar, E.; Farrant, G.K.; Follows, M.; Garczarek, L.; Speich, S.; Audic, S.; Bittner, L.; Blanke, B.; Brum, J.R.; Brunet, C.; et al. Environmental characteristics of Agulhas rings affect interocean plankton transport. Science 2015, 348, 1261447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef]
- George, D.; Mallery, P. IBM SPSS Statistics 21 Step by Step: A Simple Guide and Reference; Pearson: New York, NY, USA, 2013. [Google Scholar]
- Huson, D.H.; Beier, S.; Flade, I.; Gorska, A.; El-Hadidi, M.; Mitra, S.; Ruscheweyh, H.J.; Tappu, R. MEGAN Community Edition-Interactive Exploration and Analysis of Large-Scale Microbiome Sequencing Data. PLoS Comput. Biol. 2016, 12, e1004957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, J.; Sievert, S.M.; Wang, Y.; Seewald, J.S.; Natarajan, V.P.; Wang, F.; Xiao, X. Microbial succession during the transition from active to inactive stages of deep-sea hydrothermal vent sulfide chimneys. Microbiome 2020, 8, 102. [Google Scholar] [CrossRef]
- Nadkarni, M.A.; Martin, F.E.; Jacques, N.A.; Hunter, N. Determination of bacterial load by real-time PCR using a broad-range (universal) probe and primers set. Microbiology 2002, 148, 257–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geets, J.; Borremans, B.; Diels, L.; Springael, D.; Vangronsveld, J.; van der Lelie, D.; Vanbroekhoven, K. DsrB gene-based DGGE for community and diversity surveys of sulfate-reducing bacteria. J. Microbiol. Methods 2006, 66, 194–205. [Google Scholar] [CrossRef]
- Meyer, B.; Kuever, J. Molecular analysis of the diversity of sulfate-reducing and sulfur-oxidizing prokaryotes in the environment, using aprA as functional marker gene. Appl. Environ. Microbiol. 2007, 73, 7664–7679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, J.T.; Li, X.M.; Li, F.B.; Liu, T.X.; Young, L.Y.; Huang, W.L.; Sun, K.; Tong, H.; Hu, M. Humic Substances Facilitate Arsenic Reduction and Release in Flooded Paddy Soil. Environ. Sci. Technol. 2019, 53, 5034–5042. [Google Scholar] [CrossRef]
- Pott, A.S.; Dahl, C. Sirohaem sulfite reductase and other proteins encoded by genes at the dsr locus of Chromatium vinosum are involved in the oxidation of intracellular sulfur. Microbiology 1998, 144, 1881–1894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canfield, D.E.; Boudreau, B.P.; Mucci, A.; Gundersen, J.K. The early diagenetic formation of organic sulfur in the sediments of Mangrove Lake, Bermuda. Geochim. Cosmochim. Acta 1998, 62, 767–781. [Google Scholar] [CrossRef]
- Quatrini, R.; Appia-Ayme, C.; Denis, Y.; Jedlicki, E.; Holmes, D.S.; Bonnefoy, V. Extending the models for iron and sulfur oxidation in the extreme Acidophile Acidithiobacillus ferrooxidans. BMC Genom. 2009, 10, 394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takagi, H.; Ohtsu, I. L-Cysteine Metabolism and Fermentation in Microorganisms. Adv. Biochem. Eng. Biot. 2017, 159, 129–151. [Google Scholar] [CrossRef]
- Varon-Lopez, M.; Dias, A.C.; Fasanella, C.C.; Durrer, A.; Melo, I.S.; Kuramae, E.E.; Andreote, F.D. Sulphur-oxidizing and sulphate-reducing communities in Brazilian mangrove sediments. Environ. Microbiol. 2014, 16, 845–855. [Google Scholar] [CrossRef]
- Nie, S.; Zhang, Z.; Mo, S.; Li, J.; He, S.; Kashif, M.; Liang, Z.; Shen, P.; Yan, B.; Jiang, C. Desulfobacterales stimulates nitrate reduction in the mangrove ecosystem of a subtropical gulf. Sci. Total Environ. 2021, 769, 144562. [Google Scholar] [CrossRef]
- Ettinger, C.L.; Williams, S.L.; Abbott, J.M.; Stachowicz, J.J.; Eisen, J.A. Microbiome succession during ammonification in eelgrass bed sediments. PeerJ 2017, 5, e3674. [Google Scholar] [CrossRef] [Green Version]
- Fan, L.F.; Tang, S.L.; Chen, C.P.; Hsieh, H.L. Diversity and Composition of Sulfate- and Sulfite-Reducing Prokaryotes as Affected by Marine-Freshwater Gradient and Sulfate Availability. Microb. Ecol. 2012, 63, 224–237. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Dey, N.; Bera, A.; Tiwari, A.; Sathyaniranjan, K.B.; Chakrabarti, K.; Chattopadhyay, D. Culture independent molecular analysis of bacterial communities in the mangrove sediment of Sundarban, India. Saline Syst. 2010, 6, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wasmund, K.; Mussmann, M.; Loy, A. The life sulfuric: Microbial ecology of sulfur cycling in marine sediments. Environ. Microbiol. Rep. 2017, 9, 323–344. [Google Scholar] [CrossRef]
- Griffiths, R.I.; Thomson, B.C.; James, P.; Bell, T.; Bailey, M.; Whiteley, A.S. The bacterial biogeography of British soils. Environ. Microbiol. 2011, 13, 1642–1654. [Google Scholar] [CrossRef]
- Christian, J.; Letters, S.J.G.R. Elevated rates of organic carbon, nitrogen, and phosphorus accumulation in a highly impacted mangrove wetland. Geophys. Res. Lett. 2014, 41, 2475–2480. [Google Scholar] [CrossRef]
- Taketani, R.G.; Yoshiura, C.A.; Dias, A.C.; Andreote, F.D.; Tsai, S.M. Diversity and identification of methanogenic archaea and sulphate-reducing bacteria in sediments from a pristine tropical mangrove. Antonie Leeuwenhoek 2010, 97, 401–411. [Google Scholar] [CrossRef] [PubMed]
- Sherman, R.E.; Fahey, T.J.; Howarth, R.W. Soil-plant interactions in a neotropical mangrove forest: Iron, phosphorus and sulfur dynamics. Oecologia 1998, 115, 553–563. [Google Scholar] [CrossRef]
- Dizicheh, Z.B.; Halloran, N.; Asma, W.; Ghirlanda, G. De Novo Design of Iron-Sulfur Proteins. Methods Enzymol. 2017, 595, 33–53. [Google Scholar] [CrossRef]
- Guo, J.; Kang, Y.; Feng, Y. Bioassessment of heavy metal toxicity and enhancement of heavy metal removal by sulfate-reducing bacteria in the presence of zero valent iron. J. Environ. Manag. 2017, 203 Pt 1, 278–285. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, Y.; Ni, B.J. Zero valent iron simultaneously enhances methane production and sulfate reduction in anaerobic granular sludge reactors. Water Res. 2015, 75, 292–300. [Google Scholar] [CrossRef]
- Haaning Nielsen, A.; Lens, P.; Vollertsen, J.; Hvitved-Jacobsen, T. Sulfide-iron interactions in domestic wastewater from a gravity sewer. Water Res. 2005, 39, 2747–2755. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Li, C.; Feng, Q.; Wei, Y.; Zheng, H.; Zhao, Y.; Feng, Y.; Li, H. Shifts in soil microbial metabolic activities and community structures along a salinity gradient of irrigation water in a typical arid region of China. Sci. Total Environ. 2017, 598, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.B.; Zhang, C.J.; Wei, Q.Y.; Zhang, S.Y.; Quan, Z.X.; Li, M. Temperature and salinity drive comammox community composition in mangrove ecosystems across southeastern China. Sci. Total Environ. 2020, 742, 140456. [Google Scholar] [CrossRef] [PubMed]
- Ceccon, D.M.; Faoro, H.; Lana, P.D.; Souza, E.M.; Pedrosa, F.O. Metataxonomic and metagenomic analysis of mangrove microbiomes reveals community patterns driven by salinity and pH gradients in Paranagua Bay, Brazil. Sci. Total Environ. 2019, 694, 133609. [Google Scholar] [CrossRef]
- He, H.; Zhen, Y.; Mi, T.Z.; Xu, B.C.; Wang, G.S.; Zhang, Y.; Yu, Z.G. Community composition and distribution of sulfate- and sulfite-reducing prokaryotes in sediments from the Changjiang estuary and adjacent East China Sea. Estuar. Coast. Shelf Sci. 2015, 165, 75–85. [Google Scholar] [CrossRef]
- Kopriva, S.; Mugford, S.G.; Matthewman, C.; Koprivova, A. Plant sulfate assimilation genes: Redundancy versus specialization. Plant. Cell Rep. 2009, 28, 1769–1780. [Google Scholar] [CrossRef] [PubMed]
- Bogicevic, B.; Berthoud, H.; Portmann, R.; Bavan, T.; Meile, L.; Irmler, S. Cysteine biosynthesis in Lactobacillus casei: Identification and characterization of a serine acetyltransferase. FEMS Microbiol. Lett. 2016, 363, fnw012. [Google Scholar] [CrossRef] [Green Version]
- Tanous, C.; Soutourina, O.; Raynal, B.; Hullo, M.F.; Mervelet, P.; Gilles, A.M.; Noirot, P.; Danchin, A.; England, P.; Martin-Verstraete, I. The CymR Regulator in Complex with the Enzyme CysK Controls Cysteine Metabolism in Bacillus subtilis. J. Biol. Chem. 2008, 283, 35551–35560. [Google Scholar] [CrossRef] [Green Version]
- Friedrich, C.G.; Rother, D.; Bardischewsky, F.; Quentmeier, A.; Fischer, J. Oxidation of reduced inorganic sulfur compounds by bacteria: Emergence of a common mechanism? Appl. Environ. Microb. 2001, 67, 2873–2882. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxonomy | RS | NRS | NMS |
|---|---|---|---|
| Chromatiales | cysJ; dsrK; aprA; cysN; dsrA; sir; sat; aprM; cysQ; cysNC; cysD; cysH; dsrO; asrB; dsrN; dsrP; asrA; nrnA; MET3; dsrB; asrC; HINT4; cysI | cysN; aprA; dsrB; dsrO; sir; sat; cysQ; dsrK; cysNC; cysD; MET3; dsrM; cysH; cysJ; asrB; dsrP; dsrN; cysI; dsrA; aprB; nrnA; dsrC; HINT4 | cysJ; cysN; asrB; cysD; cysNC; dsrK; asrA; sat; cysI; cysH; cysQ; aprA; dsrC; sir; aprM; nrnA; dsrP; MET3; dsrM; aprB; dsrB; dsrA |
| Desulfobacterales | cysJ;cysN; aprA; asrB; cysQ; asrC; dsrK; nrnA; sat; dsrM; asrA; dsrN; cysH; dsrB; sir; dsrP; cysD; SAL; MET3; cysNC; dsrA; aprB; cysI; dsrO; MET22; dsrC;dsrJ | dsrK; cysJ; sat; aprA; cysN; cysQ; nrnA; dsrJ; dsrM; dsrC; asrB; cysNC; dsrN; cysH; dsrB; asrC; dsrA; sir; dsrP; aprB; SAL; asrA; MET3; MET22; dsrO; cysI; cysD | dsrK; cysJ; sat; asrB; cysN; asrA; cysQ; aprA; dsrB; MET22; nrnA; sir; dsrP; aprB; dsrJ; dsrN; dsrM; cysD; cysNC; dsrA; asrC;cysH; dsrO; cysI; MET3; dsrC |
| Rhizobiales | sat;dsrO; cysI; cysJ; cysH; asrA; cysQ; cysN; sir; nrnA; asrB | sat; aprA; cysJ; cysH; cysQ; asrB; HINT4; cysN | aprM; sat; dsrA; aprA; cysN; cysI; cysQ; asrB; cysJ; dsrO; sir; dsrK; nrnA |
| Desulfovibrionales | sat; asrC; dsrK; sir; cysN; cysJ | aprA; sat; dsrJ; cysN; asrC; dsrK; cysJ | sat; dsrK; cysN |
| Desulfuromonadales | cysI; cysN; asrB; nrnA; cysQ; sir; asrC | cysN; sir; asrC | cysI; cysN; asrB; sat; dsrA; sir; cysQ; dsrP; nrnA |
| Sample | RS | NRS | MS | NMS |
|---|---|---|---|---|
| 16S rDNA (108 copies/g soil) | 24.56 ± 8.59 | 36.50 ± 4.95 | 30.53 ± 9.21 | 11.02 ± 3.34 |
| dsrB (107 copies/g soil) | 54.45 ± 10.68 | 53.80 ± 31.69 | 48.54 ± 24.22 | 13.08 ± 8.91 |
| aprA (107 copies/g soil) | 142.54 ± 33.07 | 160.48 ± 88.53 | 151.51 ± 67.42 | 48.87 ± 31.60 |
| dsrB (10−1 copies/16r rDNA) | 1.82 ± 0.19 | 1.43 ± 0.78 | 1.63 ± 0.60 | 1.09 ± 0.46 |
| aprA (10−1 copies/16r rDNA) | 6.04 ± 0.68 | 4.31 ± 2.21 | 5.17 ± 1.85 | 4.13 ± 1.62 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mo, S.; Li, J.; Li, B.; Kashif, M.; Nie, S.; Liao, J.; Su, G.; Jiang, Q.; Yan, B.; Jiang, C. L-Cysteine Synthase Enhanced Sulfide Biotransformation in Subtropical Marine Mangrove Sediments as Revealed by Metagenomics Analysis. Water 2021, 13, 3053. https://doi.org/10.3390/w13213053
Mo S, Li J, Li B, Kashif M, Nie S, Liao J, Su G, Jiang Q, Yan B, Jiang C. L-Cysteine Synthase Enhanced Sulfide Biotransformation in Subtropical Marine Mangrove Sediments as Revealed by Metagenomics Analysis. Water. 2021; 13(21):3053. https://doi.org/10.3390/w13213053
Chicago/Turabian StyleMo, Shuming, Jinhui Li, Bin Li, Muhammad Kashif, Shiqing Nie, Jianping Liao, Guijiao Su, Qiong Jiang, Bing Yan, and Chengjian Jiang. 2021. "L-Cysteine Synthase Enhanced Sulfide Biotransformation in Subtropical Marine Mangrove Sediments as Revealed by Metagenomics Analysis" Water 13, no. 21: 3053. https://doi.org/10.3390/w13213053