
The Holocene History of the Diatom Community in a Small Water Body on Shemya Island (Aleutian Arc, USA): The Influence of Global and Local Environmental Changes

 , , , , , and
, , , , , and 
Abstract
:1. Introduction
2. Materials and Methods
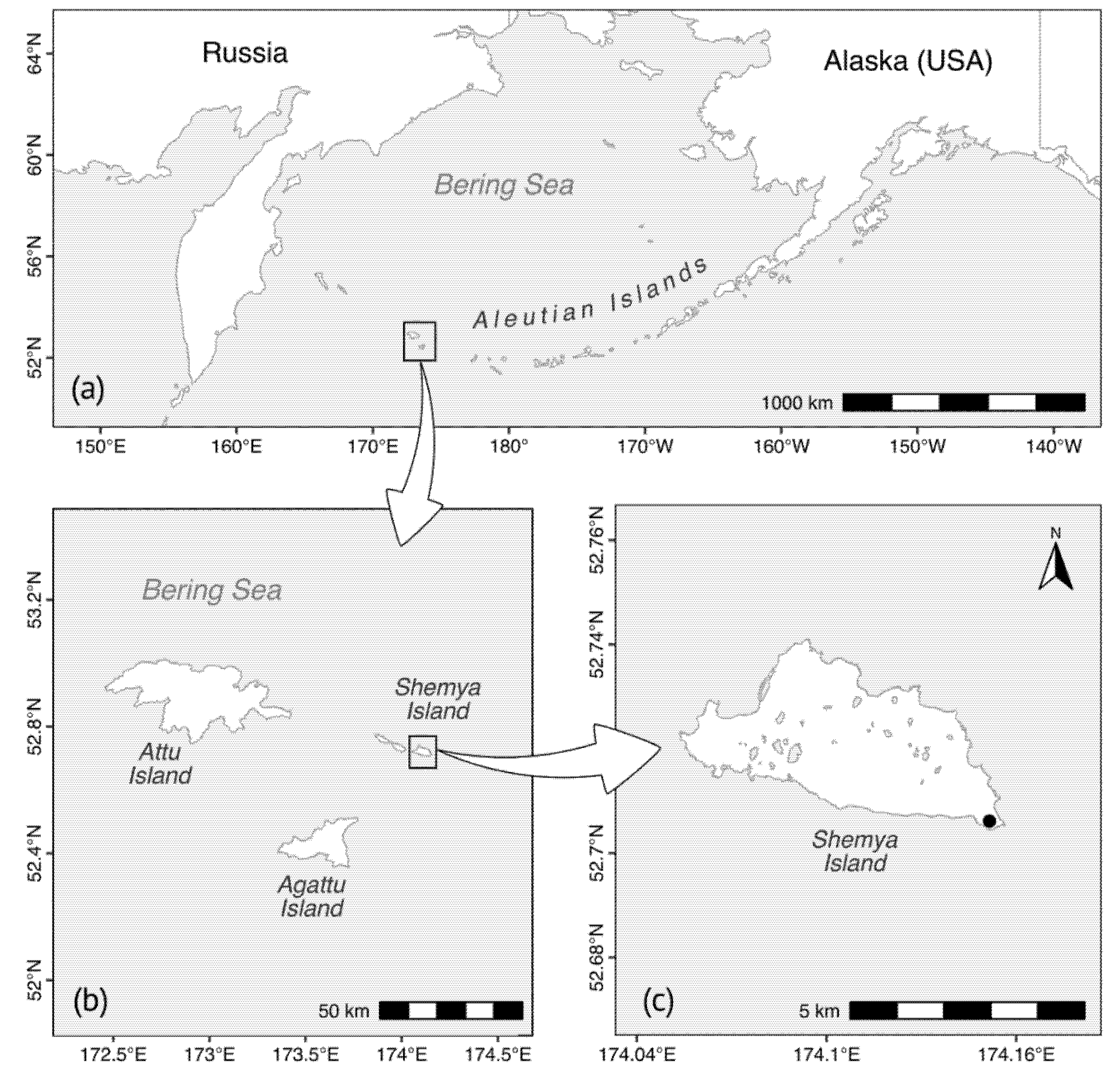
2.1. Study Area and Sampling
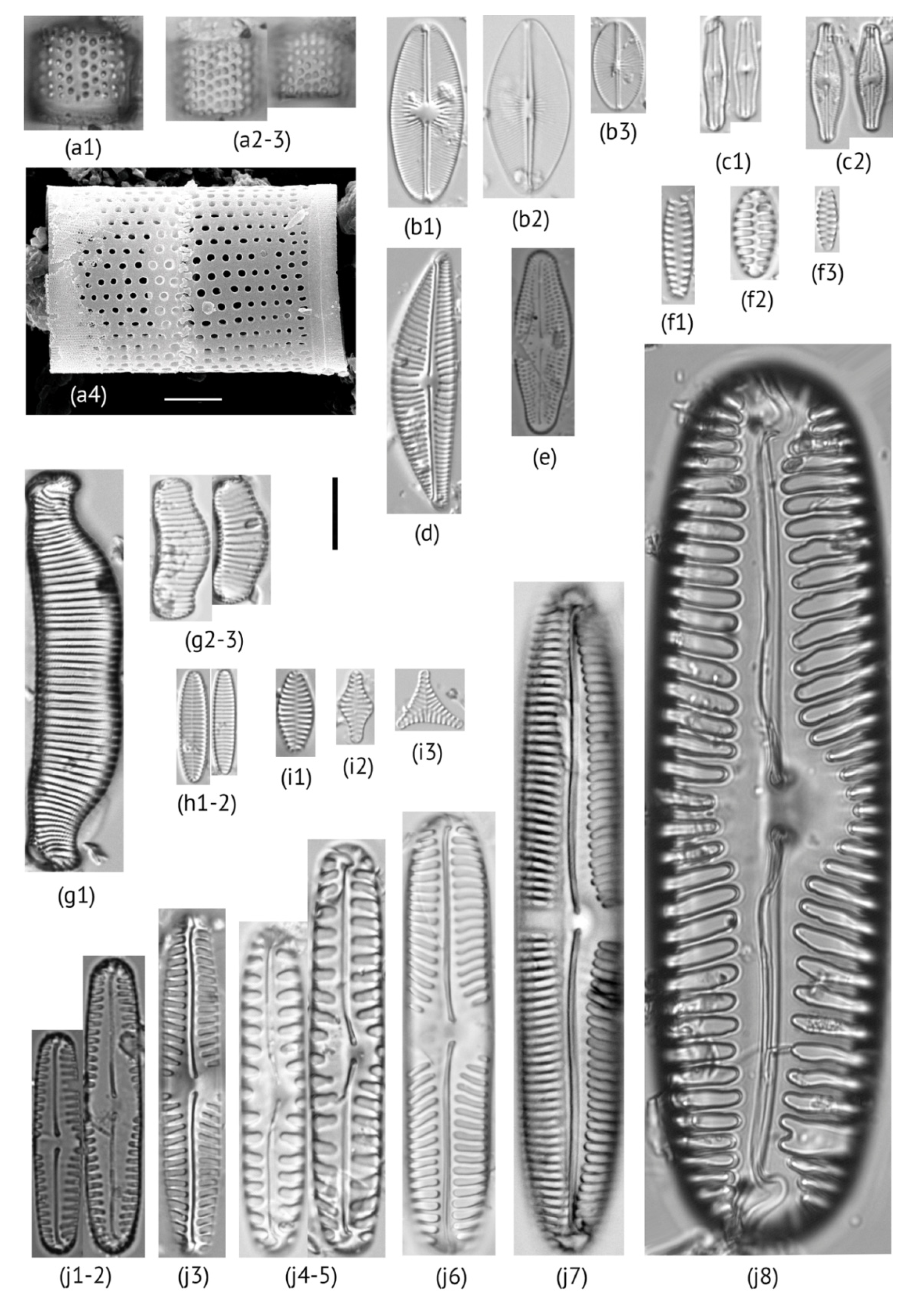
2.2. Diatom Analysis
3. Results
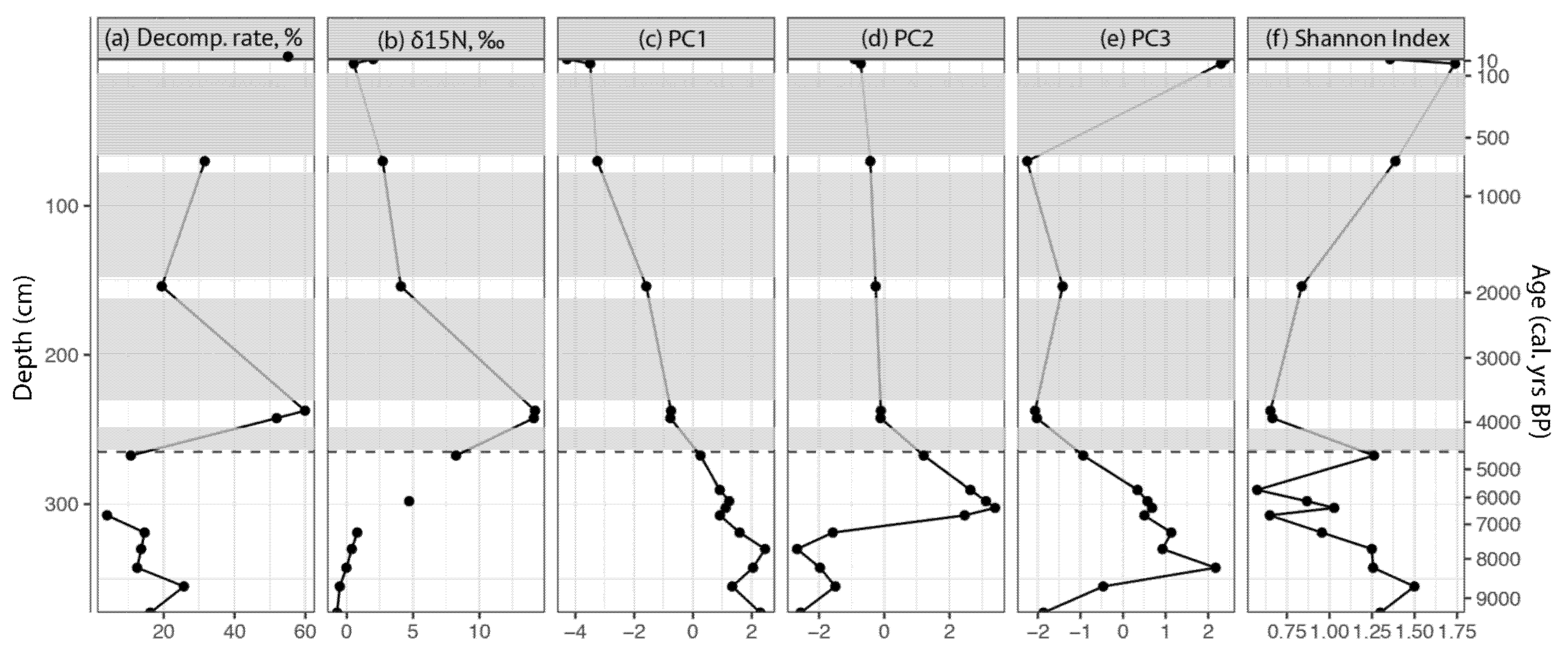
3.1. Environmental Factor Dynamics
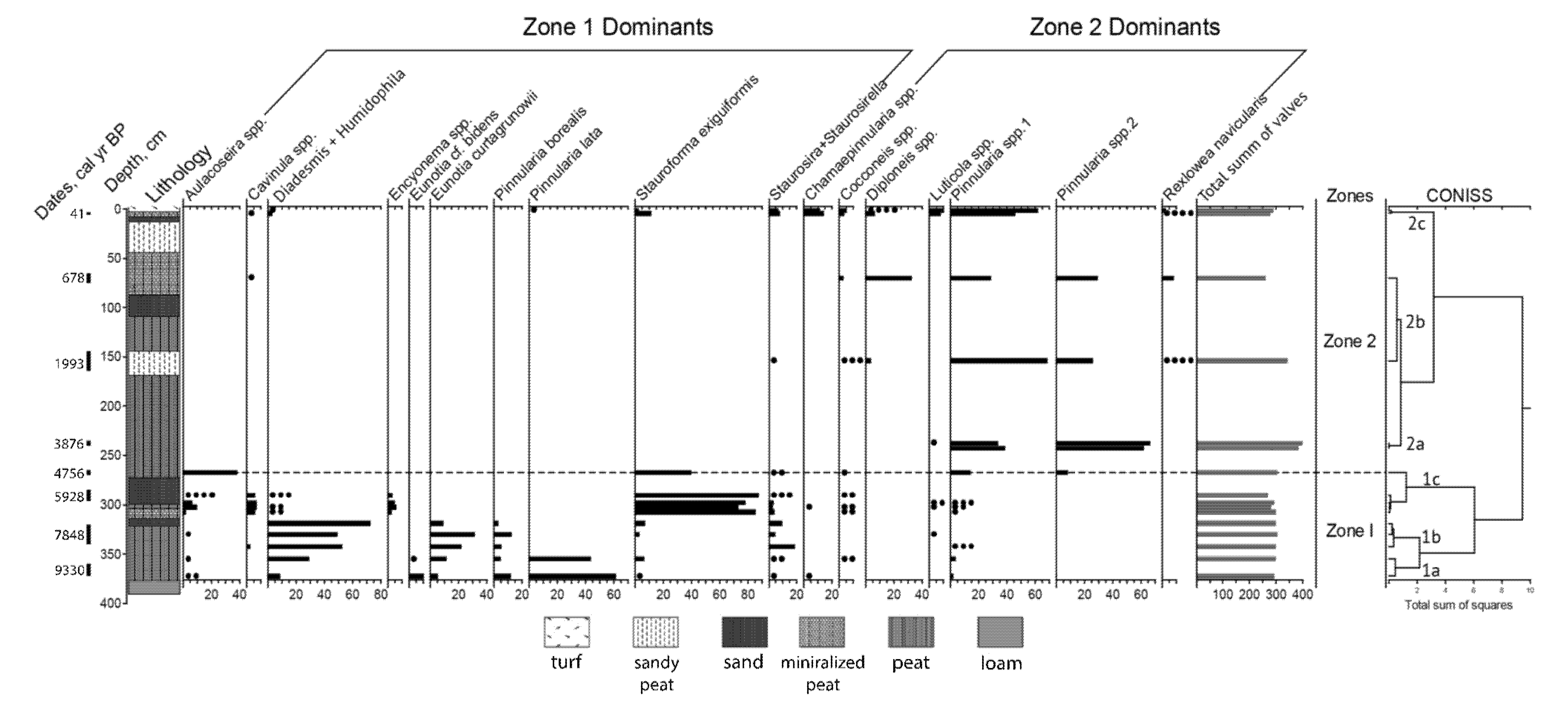
3.2. Diatom Community Dynamics
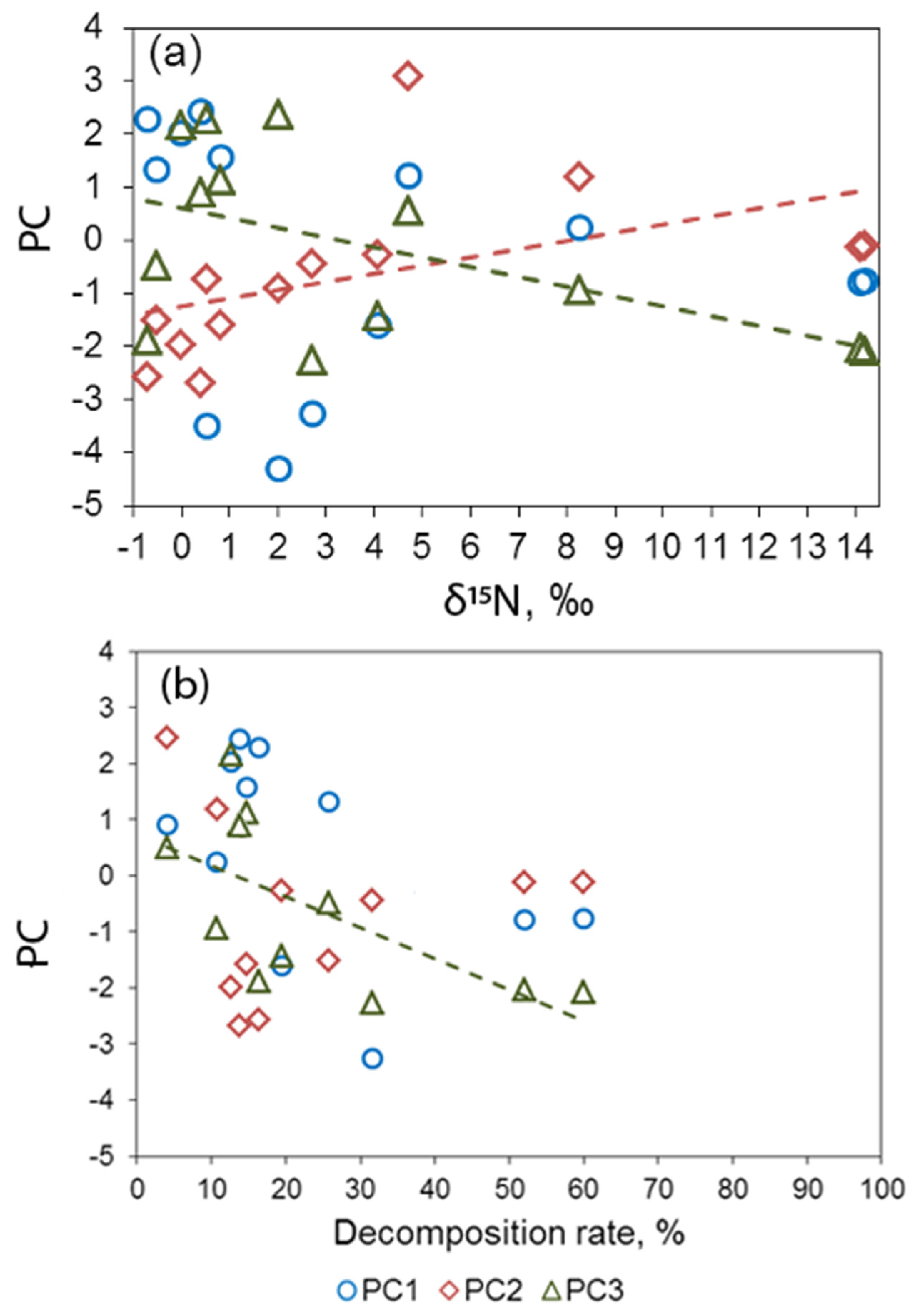
3.3. Principal Component Analysis
4. Discussion
4.1. Diatom Community Dynamics: Environmental Effects
4.2. Environmental Factors as Drivers of Gradual and Abrupt Changes in Diatom Community Y
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Paleoreconstruction Based on the Diatom Analysis
References
- Mackel, R.; Schneider, R.; Seidel, J. Anthropogenic impact on the landscape of southern Badenia (Germany) during the holocene-documented by colluvial and alluvial sediments. Archaeometry 2003, 45, 487–501. [Google Scholar] [CrossRef]
- Van Bellen, S.; Garneau, M.; Booth, R.K. Holocene carbon accumulation rates from three ombrotrophic peatlands in boreal Quebec, Canada: Impact of climate-driven ecohydrological change. Holocene 2011, 21, 1217–1231. [Google Scholar] [CrossRef]
- Anderson, T.W.; Lewis, C.F.M. A new water-level history for Lake Ontario basin: Evidence for a climate-driven early Holocene lowstand. J. Paleolimnol. 2012, 47, 513–530. [Google Scholar] [CrossRef]
- García-Alix, A.; Jimenez-Espejo, F.J.; Lozano, J.A.; Jiménez-Moreno, G.; Martinez-Ruiz, F.; García Sanjuán, L.; Aranda Jiménez, G.; García Alfonso, E.; Ruiz-Puertas, G.; Anderson, R.S. Anthropogenic impact and lead pollution throughout the Holocene in Southern Iberia. Sci. Total Environ. 2013, 449, 451–460. [Google Scholar] [CrossRef] [PubMed]
- Barrow, J.L.; Jeziorski, A.; Rühland, K.M.; Hadley, K.R.; Smol, J.P. Diatoms indicate that calcium decline, not acidification, explains recent cladoceran assemblage changes in south-central Ontario softwater lakes. J. Paleolimnol. 2014, 52, 61–75. [Google Scholar] [CrossRef]
- Corbett, D.G.; West, D.L.; Lefèvre, C. (Eds.) The People at the End of the World: The Western Aleutians Project and the Archaeology of Shemya Island; Alaska Anthropological Association: Anchorage, AK, USA, 2010; ISBN 1890396087. [Google Scholar]
- Savinetsky, A.B.; Kiseleva, N.K.; Khassanov, B.F. Dynamics of sea mammal and bird populations of the Bering Sea region over the last several millennia. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2004, 209, 335–352. [Google Scholar] [CrossRef]
- Savinetsky, A.B.; Khasanov, B.F.; West, D.L.; Kiseleva, N.K.; Krylovich, O.A. Nitrogen Isotope Composition of Peat Samples as a Proxy for Determining Human Colonization of Islands. Arct. Anthropol. 2014, 51, 78–85. [Google Scholar] [CrossRef]
- Hatfield, V.L.; Nicolaysen, K.; WEST, D.L.; Krylovich, O.A.; Bruner, K.M.; Savinetsky, A.B.; Vasyukov, D.D.; MacInnes, B.T.; Khasanov, B.F.; Persico, L.; et al. Human resilience and resettlement among the Islands of Four Mountains, Aleutians, Alaska. Quat. Res. 2019, 91, 917–933. [Google Scholar] [CrossRef]
- Krylovich, O.A.; Vasyukov, D.D.; Khasanov, B.F.; Hatfield, V.; West, D.; Savinetsky, A.A. Hunter-gatherers subsistence and impact on fauna in the Islands of Four Mountains, Eastern Aleutians, Alaska, over 3000 yr. Quat. Res. 2019, 91, 983–1002. [Google Scholar] [CrossRef]
- Coats, R.R. Volcanic Activity in the Aleutian Arc: Contributions to General Geology; United States Government Printing Office: Washington, DC, USA, 1950. [Google Scholar]
- Black, R.F. Isostatic, tectonic, and eustatic movements of sea level in the Aleutian Islands, Alaska. In Earth Rheology, Isostasy and Eustacy; Moerner, N.-A., Ed.; John Wiley & Sons: Chichester, UK, 1980; pp. 231–248. [Google Scholar]
- Murie, O.J. Fauna of the Aleutian Islands and Alaska Peninsula; North American Fauna 61–63, 1959. Available online: http://pubs.er.usgs.gov/publication/naf61A (accessed on 30 September 2021).
- Garroutte, M.; Huettmann, F.; Webb, C.O.; Ickert-Bond, S.M. Biogeographic and anthropogenic correlates of Aleutian Islands plant diversity: A machine-learning approach. J. Syst. Evol. 2018, 56, 476–497. [Google Scholar] [CrossRef] [Green Version]
- MacArthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; Princeton University Press: Princeton, NJ, USA, 2001; ISBN 9780691088365. [Google Scholar]
- Edwards, K.J.; Dugmore, A.J.; Blackford, J.J. Vegetational response to tephra deposition and land-use change in Iceland: A modern analogue and multiple working hypothesis approach to tephropalynology. Polar Rec. 2004, 40, 113–120. [Google Scholar] [CrossRef] [Green Version]
- Hotes, S.; Grootjans, A.P.; Takahashi, H.; Ekschmitt, K.; Poschlod, P. Resilience and alternative equilibria in a mire plant community after experimental disturbance by volcanic ash. Oikos 2010, 119, 952–963. [Google Scholar] [CrossRef]
- McConnell, J.R.; Sigl, M.; Plunkett, G.; Burke, A.; Kim, W.M.; Raible, C.C.; Wilson, A.I.; Manning, J.G.; Ludlow, F.; Chellman, N.J.; et al. Extreme climate after massive eruption of Alaska’s Okmok volcano in 43 BCE and effects on the late Roman Republic and Ptolemaic Kingdom. Proc. Natl. Acad. Sci. USA 2020, 117, 15443–15449. [Google Scholar] [CrossRef] [PubMed]
- Hulten, E. Flora of Alaska and Neighboring Territories: A Manual of the Vascular Plants; Stanford University Press: Stanford, UK, 1968; ISBN 9780804706438. [Google Scholar]
- Heusser, C.J. Late Quaternary vegetation of the Aleutian Islands, southwestern Alaska. Can. J. Bot. 1990, 68, 1320–1326. [Google Scholar] [CrossRef]
- West, D.; Crawford, M.; Savinetsky, A.B. Genetics, prehistory and the colonisation of the Aleutian Islands. Earth Environ. Sci. Trans. R. Soc. Edinb. 2007, 98, 47–57. [Google Scholar] [CrossRef]
- Crockford, S.J. Archaeozoology of Adak Island: 6000 Years of Subsistence History in the Central Aleutians. In The People Before: The Geology, Paleoecology and Archaeology of Adak Island, Alaska; West, D., Hatfield, V., Wilmerding, E., Lefèvre, C., Gualtieri, L., Eds.; Archaeopress: Oxford, UK, 2012; pp. 107–143. ISBN 9781407309057. [Google Scholar]
- Vasyukov, D.D.; Krylovich, O.A.; West, D.L.; Hatfield, V.; Savinetsky, A.B. Ancient canids of the Aleutian Islands (new archaeological discoveries from the Islands of Four Mountains). Quat. Res. 2019, 91, 1028–1044. [Google Scholar] [CrossRef]
- Byrd, V.G.; Renner, H.M.; Renner, M. Distribution patterns and population trends of breeding seabirds in the Aleutian Islands. Fish. Oceanogr. 2005, 14, 139–159. [Google Scholar] [CrossRef]
- Maron, J.L.; Estes, J.A.; Croll, D.A.; Danner, E.M.; Elmendorf, S.C.; Buckelew, S.L. An introduced predator alters Aleutian Island plant communities by thwarting nutrient subsidies. Ecol. Monogr. 2006, 76, 3–24. [Google Scholar] [CrossRef] [Green Version]
- Kuzmicheva, E.A.; Smyshlyaeva, O.I.; Vasyukov, D.D.; Khasanov, B.F.; Krylovich, O.A.; Okuno, M.; West, D.L.; Hatfield, V.L.; Savinetsky, A.B. A 7300-yr-old environmental history of seabird, human, and volcano impacts on Carlisle Island (the Islands of Four Mountains, eastern Aleutians, Alaska). Quat. Res. 2019, 91, 934–952. [Google Scholar] [CrossRef]
- Smyshlyaeva, O.I.; Severova, E.E.; Krylovich, O.A.; Kuzmicheva, E.A.; Savinetsky, A.B.; Dixie, W.; Hatfield, V. Ornithogenic vegetation: How significant has the seabird influence been on the Aleutian Island vegetation during the Holocene? Ecol. Evol. 2021, 11, 14088–14100. [Google Scholar] [CrossRef]
- Croll, D.A.; Maron, J.L.; Estes, J.A.; Danner, E.M.; Byrd, G.V. Introduced predators transform subarctic islands from grassland to tundra. Science 2005, 307, 1959–1961. [Google Scholar] [CrossRef] [Green Version]
- Davis, R.S.; Knecht, R.A. Continuity and change in the eastern Aleutian archaeological sequence. Hum. Biol. 2010, 82, 507–524. [Google Scholar] [CrossRef] [Green Version]
- Causey, D.; Corbett, D.G.; Lefevre, C.; West, D.L.; Savinetsky, A.B.; Kiseleva, N.K.; Khassanov, B.F. The palaeoenvironment of humans and marine birds of the Aleutian Islands: Three millennia of change. Fish. Oceanogr. 2005, 14, 259–276. [Google Scholar] [CrossRef]
- Lefèvre, C.; Corbett, D.G. Appendix G: WAAPP Radiocarbon Dates. In The People at the End of the World: The Western Aleutians Project and the Archaeology of Shemya Island; Corbett, D.G., West, D.L., Lefèvre, C., Eds.; Alaska Anthropological Association: Anchorage, AK, USA, 2010; pp. 243–257. ISBN 1890396087. [Google Scholar]
- Bailey, H.L.; Kaufman, D.S.; Sloane, H.J.; Hubbard, A.L.; Henderson, A.C.; Leng, M.J.; Meyer, H.; Welker, J.M. Holocene atmospheric circulation in the central North Pacific: A new terrestrial diatom and δ18O dataset from the Aleutian Islands. Quat. Sci. Rev. 2018, 194, 27–38. [Google Scholar] [CrossRef] [Green Version]
- Smol, J.P. Diatoms and Lake Acidity: Reconstructing pH from Siliceous Algal Remains in Lake Sediments; Springer: Dordrecht, The Netherlands, 1986; ISBN 978-90-6193-536-0. [Google Scholar]
- Battarbee, R.W. Diatom analysis. In Handbook of Holocene Palaeoecology and Palaeohydrology; Berglund, B.E., Ralska-Jasiewiczowa, M., Eds.; Wiley: Chichester, UK, 1986; ISBN 9780471906919. [Google Scholar]
- Vos, P.C.; de Wolf, H. Diatoms as a tool for reconstructing sedimentary environments in coastal wetlands; methodological aspects. Hydrobiologia 1993, 269, 285–296. [Google Scholar] [CrossRef]
- Reid, M.A.; Tibby, J.C.; Penny, D.; Gell, P.A. The use of diatoms to assess past and present water quality. Aust. J. Ecol. 1995, 20, 57–64. [Google Scholar] [CrossRef]
- Pandey, L.K.; Bergey, E.A.; Lyu, J.; Park, J.; Choi, S.; Lee, H.; Depuydt, S.; Oh, Y.-T.; Lee, S.-M.; Han, T. The use of diatoms in ecotoxicology and bioassessment: Insights, advances and challenges. Water Res. 2017, 118, 39–58. [Google Scholar] [CrossRef] [PubMed]
- Hein, M. Flora of Adak Island, Alaska: Bacillariophyceae (Diatoms); J.Cramer: Berlin, Stuttgart, 1990; ISBN 978-3443570125. [Google Scholar]
- Neplyukhina, A.A.; Chudaev, D.A.; Gololobova, M.A. Diatomovie vodorosly torfyanogo otlojenia ostrova Karlayl (Aleutskye ostrova, S.Sh.A) (in russian). In Materialy V Vserossiyskoy Naychnoy Konferencii s Mezjdunarodnym Uchastiem «Dinamika Ecosistem v Golocene» (k 100-Letiy L.G. Dinesmana); Savinetsky, A.B., Ed.; Media-PRESS: Moskva, Russia, 2019; pp. 235–236. ISBN 978-5-901003-57-2. [Google Scholar]
- Potapova, M. Diatoms of Bering Island, Kamchatka, Russia. Nova Hedwig. 2014, 143, 63–102. [Google Scholar]
- Neplyukhina, A.A.; Chudaev, D.A.; Krylovich, O.A.; Gololobova, M.A. Diatoms of the Peatbog Sediments from Shemya Island (Aleutian Islands, United States). Mosc. Univ. Biol. Sci. Bull. 2018, 73, 76–81. [Google Scholar] [CrossRef]
- Neplyukhina, A.A.; Chudaev, D.A.; Gololobova, M.A. Pinnularia arkadii sp. nov., a new diatom (Naviculales, Bacillariophyceae) from Shemya Island, Alaska, USA. Nov. Sist. Nizshikh Rastenii 2018, 52, 83–89. [Google Scholar] [CrossRef]
- Savinetsky, A.B.; Kiseleva, N.K.; Khassanov, B.F. Paleoenvironment—Holocene deposits from Shemya Island. In The People at the End of the World: The Western Aleutians Project and the Archaeology of Shemya Island; Corbett, D.G., West, D.L., Lefèvre, C., Eds.; Alaska Anthropological Association: Anchorage, AK, USA, 2010; ISBN 1890396087. [Google Scholar]
- Parnell, A. Bchron: Radiocarbon Dating, Age-Depth Modelling, Relative Sea Level Rate Estimation, and Non-Parametric Phase Modelling: R package version 4.1.1. Available online: https://CRAN.R-project.org/package=Bchron (accessed on 30 September 2021).
- Haslett, J.; Parnell, A. A simple monotone process with application to radiocarbon-dated depth chronologies. J. R. Stat. Soc. Ser. C Appl. Stat. 2008, 57, 399–418. [Google Scholar] [CrossRef]
- Kiseleva, N.K.; Savinetsky, A.B.; Khassanov, B.F. Development of Natural Processes over the Holocene in Aleutian Islands. Izv. AN Seriya Geogr. 2002, 1, 97–103. [Google Scholar]
- Gorham, E. Northern Peatlands: Role in the Carbon Cycle and Probable Responses to Climatic Warming. Ecol. Appl. 1991, 1, 182–195. [Google Scholar] [CrossRef] [PubMed]
- Ise, T.; Dunn, A.L.; Wofsy, S.C.; Moorcroft, P.R. High sensitivity of peat decomposition to climate change through water-table feedback. Nat. Geosci. 2008, 1, 763–766. [Google Scholar] [CrossRef]
- Biester, H.; Knorr, K.-H.; Schellekens, J.; Basler, A.; Hermanns, Y.-M. Comparison of different methods to determine the degree of peat decomposition in peat bogs. Biogeosciences 2014, 11, 2691–2707. [Google Scholar] [CrossRef] [Green Version]
- Kelly, M.G.; Adams, C.; Graves, A.C. The Trophic Diatom Index: A User’s Manual; Revised Edition; WRC: Swindon, UK, 2001; ISBN 1-857-05597-7. [Google Scholar]
- Dam, H.; Mertens, A.; Sinkeldam, J. A coded checklist and ecological indicator values of freshwater diatoms from The Netherlands. Neth. J. Aquat. Ecol. 1994, 28, 117–133. [Google Scholar] [CrossRef]
- Lange-Bertalot, H. (Ed.) Diatoms of Europe; A.R.G. Gantner Verlag (Koeltz Scientific Books): Königstein, Germany, 2000. [Google Scholar]
- Spaulding, S.A.; Bishop, I.W.; Edlund, M.B.; Lee, S.; Fure, P.; Jovanovska, E.; Potapova, M. Diatoms of North America. Available online: https://diatoms.org (accessed on 27 April 2021).
- Guiry, M.D.; Guiry, G. AlgaeBase: World-Wide Electronic Publication. Available online: http://www.algaebase.org (accessed on 27 April 2021).
- Lange-Bertalot, H.; Bąk, M.; Witkowski, A. Eunotia and Some Related Genera. In Süßwasserflora von Mitteleuropa; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; Spektrum Akademischer Verlag: Stuttgart, Germany, 2007. [Google Scholar]
- Lange-Bertalot, H. Navicula Sensu Stricto, 10 Genera Separated from Navicula Sensu Lato, Frustulia. In Diatoms of Europe; Lange-Bertalot, H., Ed.; A.R.G. Gantner Verlag (Koeltz Scientific Books): Königstein, Germany, 2000. [Google Scholar]
- Grimm, E.C. Tilia (software): Version 2.6.1. Available online: https://www.tiliait.com/download/ (accessed on 12 June 2021).
- Grimm, E.C. CONISS: A FORTRAN 77 program for stratigraphically constrained cluster analysis by the method of incremental sum of squares. Comput. Geosci. 1987, 13, 13–35. [Google Scholar] [CrossRef]
- Schaumburg, J.; Schranz, C.; Stelzer, D.; Hofmann, G.; Gutowski, A.; Foerster, J. Instruction Protocol for the ecological Assessment of Running Waters for Implementation of the EC Water Framework Directive: Macrophytes and Phytobenthos. 2006. Available online: http://www.planktonforum.eu/fileadmin/_migrated/content_uploads/PHYLIB_instruction_protocol_running_waters_01_2006__Schaumburg_etal_2006_.pdf (accessed on 12 June 2021).
- Spaulding, S.; Edlund, M. Staurosira. In Diatoms of North America. Available online: https://diatoms.org/genera/staurosira (accessed on 13 May 2021).
- Spaulding, S.; Edlund, M. Chamaepinnularia. In Diatoms of North America. Available online: https://diatoms.org/genera/chamaepinnularia (accessed on 13 May 2021).
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- TIBCO Software Inc. TIBCO Statistica, v. 12.5.192.7. Available online: https://www.tibco.com/products/tibco-statistica (accessed on 30 September 2021).
- Juggins, S. Rioja: Analysis of Quaternary Science Data.: R Package Version 0.9-26. Available online: https://cran.r-project.org/package=rioja (accessed on 12 June 2021).
- Jari, O.; Guillaume, B.F.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package: R Package Version 2.5-7. Available online: https://CRAN.R-project.org/package=vegan (accessed on 12 June 2021).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer International Publishing: Cham, Switzerland, 2016; ISBN 978-3-319-24275-0. [Google Scholar]
- Mann, D.H.; Crowell, A.L.; Hamilton, T.D.; Finney, B.P. Holocene Geologic and Climatic History around the Gulf of Alaska. Arct. Anthropol. 1998, 35, 112–131. [Google Scholar]
- Nelson, A.R.; Briggs, R.W.; Dura, T.; Engelhart, S.E.; Gelfenbaum, G.; Bradley, L.-A.; Forman, S.L.; Vane, C.H.; Kelley, K.A. Tsunami recurrence in the eastern Alaska-Aleutian arc: A Holocene stratigraphic record from Chirikof Island, Alaska. Geosphere 2015, 11, 1172–1203. [Google Scholar] [CrossRef] [Green Version]
- Kaufman, D.S.; Axford, Y.L.; Henderson, A.C.; McKay, N.P.; Oswald, W.W.; Saenger, C.; Anderson, R.S.; Bailey, H.L.; Clegg, B.; Gajewski, K.; et al. Holocene climate changes in eastern Beringia (NW North America)—A systematic review of multi-proxy evidence. Quat. Sci. Rev. 2016, 147, 312–339. [Google Scholar] [CrossRef] [Green Version]
- Kaplin, P.A. Osnovnye Etapy Razvitiya Beregovoi Zony v Golotsene[Main Stages of Development of Coastal Area in the Holocene]. Geogr. Investig. Quat. Period 1982, 87–96. [Google Scholar]
- Kaplin, P.A.; Leont’ev’, O.K.; Luk’yanova, S.A.; Nikiforov, L.G. Berega [Shores]; Mysl’: Moscow, Russia, 1991. [Google Scholar]
- Caut, S.; Angulo, E.; Pisanu, B.; Ruffino, L.; Faulquier, L.; Lorvelec, O.; Chapuis, J.-L.; Pascal, M.; Vidal, E.; Courchamp, F. Seabird modulations of isotopic nitrogen on islands. PLoS ONE 2012, 7, e39125. [Google Scholar] [CrossRef] [Green Version]
- González-Bergonzoni, I.; Johansen, K.L.; Mosbech, A.; Landkildehus, F.; Jeppesen, E.; Davidson, T.A. Small birds, big effects: The little auk (Alle alle) transforms high Arctic ecosystems. Proc. Biol. Sci. 2017, 284. [Google Scholar] [CrossRef] [Green Version]
- Manny, B.A.; Johnson, W.C.; Wetzel, R.G. Nutrient additions by waterfowl to lakes and reservoirs: Predicting their effects on productivity and water quality. Hydrobiologia 1994, 279-280, 121–132. [Google Scholar] [CrossRef]
- Otero, X.L.; de La Peña-Lastra, S.; Pérez-Alberti, A.; Ferreira, T.O.; Huerta-Diaz, M.A. Seabird colonies as important global drivers in the nitrogen and phosphorus cycles. Nat. Commun. 2018, 9, 246. [Google Scholar] [CrossRef] [PubMed]
- Ganning, B.; Wulff, F.; Ganning, B. The Effects of Bird Droppings on Chemical and Biological Dynamics in Brackish Water Rockpools. Oikos 1969, 20, 274. [Google Scholar] [CrossRef]
- Mulder, C.P.H.; Anderson, W.B.; Towns, D.R.; Bellingham, P.J. Seabird Islands; Oxford University Press: Oxford, UK, 2011; ISBN 9780199735693. [Google Scholar]
- Manley, W.F. Postglacial Flooding of the Bering land Bridge: A Geospatial Animation: INSTAAR, Vol. 1. Available online: http://instaar.colorado.edu/QGISL/bering_land_bridge (accessed on 12 June 2021).
- Smith, D.E.; Harrison, S.; Firth, C.R.; Jordan, J.T. The early Holocene sea level rise. Quat. Sci. Rev. 2011, 30, 1846–1860. [Google Scholar] [CrossRef]
- Gladwell, M. The Tipping Point: How Little Things Can Make a Big Difference; Little: Boston, MA, USA, 2000; ISBN 9780316316965. [Google Scholar]
- Dakos, V.; Matthews, B.; Hendry, A.P.; Levine, J.; Loeuille, N.; Norberg, J.; Nosil, P.; Scheffer, M.; De Meester, L. Ecosystem tipping points in an evolving world. Nat. Ecol. Evol. 2019, 3, 355–362. [Google Scholar] [CrossRef] [Green Version]
- Dudney, J.; Suding, K.N. The elusive search for tipping points. Nat. Ecol. Evol. 2020, 4, 1449–1450. [Google Scholar] [CrossRef]
- Souffreau, C.; Vanormelingen, P.; Verleyen, E.; Sabbe, K.; Vyverman, W. Tolerance of benthic diatoms from temperate aquatic and terrestrial habitats to experimental desiccation and temperature stress. Phycologia 2010, 49, 309–324. [Google Scholar] [CrossRef] [Green Version]
- Kulikovskiy, M.S. The species composition and distribution of diatom algae in sphagnum bogs in European Russia: The Polistovo-Lovatskii land tract. Inland Water Biol. 2009, 2, 135–143. [Google Scholar] [CrossRef]
- Cantonati, M.; Lange-Bertalot, H.; Decet, F.; Gabrieli, J. Diatoms in very-shallow pools of the site of community importance Danta di Cadore Mires (south-eastern Alps), and the potential contribution of these habitats to diatom biodiversity conservation. Nova Hedwig. 2011, 93, 475–507. [Google Scholar] [CrossRef]
- Gottschalk, S.; Kahlert, M. Shifts in taxonomical and guild composition of littoral diatom assemblages along environmental gradients. Hydrobiologia 2012, 694, 41–56. [Google Scholar] [CrossRef]
- Cvetkoska, A.; Levkov, Z.; Hamilton, P.B.; Potapova, M. The biogeographic distribution of Cavinula (Bacillariophyceae) in North America with the descriptions of two new species. Phytotaxa 2014, 184, 181. [Google Scholar] [CrossRef]
- Barinova, S.S.; Niyatbekov, T. Bioindication of aquatic habitats with diatom algae in the Pamir Mountains, Tajikistan. MOJES 2018, 3, 117–120. [Google Scholar] [CrossRef] [Green Version]
- Antoniades, D.; Douglas, M.S.V.; Smol, J.P. benthic diatom autecology and inference model development from the Canadian High Arctic archipelago. J. Phycol. 2005, 41, 30–45. [Google Scholar] [CrossRef]
- Foets, J.; Stanek-Tarkowska, J.; Teuling, A.J.; van de Vijver, B.; Wetzel, C.E.; Pfister, L. Autecology of terrestrial diatoms under anthropic disturbance and across climate zones. Ecol. Indic. 2021, 122, 107248. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxonomic Groups | Ecological Characteristics | |||
|---|---|---|---|---|
| Combined Groups | Taxa Included in the Combined Groups | Ecological Group | Habitat | Water Patterns |
| Aulacoseira spp. | Aulacoseira canadensis, Aulacoseira crassipunctata, Aulacoseira italica | Freshwater | Benthic, planktonic | Slightly acidic waters |
| Cavinula spp. | Cavinula cocconeiformis, Cavinula cocconeiformis f. elliptica, Cavinula davisiae | Freshwater, terrestrial | Benthic | Oligotrophic, slightly acidic waters with a low conductivity |
| Chamaepinnularia spp. | Chamaepinnularia krookii, Chamaepinnularia krookiiformis, | Freshwater, brackish, terrestrial | Benthic | |
| Cocconeis spp. | Cocconeis cf. californica, Cocconeis cf. islandica, Cocconeis cf. scutellum, Cocconeis costata, Cocconeis placentula s.l. | Marine/freshwater | Epiphytic | |
| Diadesmis + Humidophila | Diadesmis mochalovae, Humidophila laevissima, | Freshwater, terrestrial | Benthic | |
| Diploneis spp. | Diploneis krammeri, Diploneis ovalis, Diploneis pseudovalis | Freshwater, terrestrial | Benthic | Low alkalinity, slightly acid waters |
| Encyonema spp. | Encyonema silesiacum, Encyonema minutum | Freshwater, marine, terrestrial | Benthic | Circumneutral waters |
| Luticola spp. | Luticola arctica, Luticola cf. mutica | Freshwater, terrestrial | Benthic | |
| Pinnularia spp.1 | Pinnularia arkadii, Pinnularia intermedia, Pinnularia lagerstedtii, Pinnularia pseudoparva | Freshwater, terrestrial | Benthic | Slightly acidic, oligotrophyc, low conductivity waters |
| Pinnularia spp.2 | Pinnularia inconstans, Pinnularia cf. islandica | Freshwater | Benthic | |
| Staurosira spp. + Staurosirella spp. | Staurosira construens, Staurosira construens var. exigua, Staurosira construens var. venter, Staurosirella lapponica, Staurosirella martyi, Staurosirella minuta | Freshwater | Benthic, planktonic, epiphytic | |
| Individual species | ||||
| Eunotia cf. bidens | Freshwater | Benthic, epiphytic | Acid waters with a low conductivity | |
| Eunotia curtagrunowii | Freshwater, terrestrial | Benthic | Cold, acidic waters with low conductivity | |
| Pinnularia borealis | Freshwater, terrestrial | Benthic | Cold, oligotrophic waters | |
| Pinnularia lata | Freshwater, terrestrial | Benthic | Cold, oligotrophic waters | |
| Rexlowea navicularis | Freshwater | Benthic | ||
| Stauroforma exiguiformis | Freshwater | Epiphytic | Circumneutral waters | |
| Taxa | PC1 | PC2 | PC3 |
|---|---|---|---|
| Aulacoseira spp. | 0.11 | 0.36 | −0.11 |
| Cavinula spp. | 0.32 | 0.83 | 0.27 |
| Chamaepinnularia spp. | −0.69 | −0.15 | 0.56 |
| Cocconeis spp. | −0.85 | −0.05 | 0.37 |
| Diadesmis + Humidophila | 0.51 | −0.58 | 0.39 |
| Diploneis spp. | −0.53 | −0.08 | −0.31 |
| Encyonema spp. | 0.28 | 0.84 | 0.19 |
| Eunotia cf. bidens | 0.29 | −0.35 | −0.31 |
| Eunotia curtagrunowii | 0.54 | −0.61 | 0.32 |
| Luticola spp. | −0.70 | −0.13 | 0.58 |
| Pinnularia borealis | 0.58 | −0.68 | 0.02 |
| Pinnularia lata | 0.34 | −0.41 | −0.30 |
| Pinnularia spp.1 | −0.86 | −0.16 | −0.12 |
| Pinnularia spp.2 | −0.32 | −0.04 | −0.68 |
| Rexlowea navicularis | −0.65 | −0.11 | −0.23 |
| Stauroforma exiguiformis | 0.29 | 0.90 | 0.20 |
| Staurosira + Staurosirella | 0.13 | −0.31 | 0.77 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neplyukhina, A.A.; Tchabovsky, A.V.; Gololobova, M.A.; Smyshlyaeva, O.I.; Krylovich, O.A.; Savinetsky, A.B.; Kotov, A.A. The Holocene History of the Diatom Community in a Small Water Body on Shemya Island (Aleutian Arc, USA): The Influence of Global and Local Environmental Changes. Water 2021, 13, 3134. https://doi.org/10.3390/w13213134
Neplyukhina AA, Tchabovsky AV, Gololobova MA, Smyshlyaeva OI, Krylovich OA, Savinetsky AB, Kotov AA. The Holocene History of the Diatom Community in a Small Water Body on Shemya Island (Aleutian Arc, USA): The Influence of Global and Local Environmental Changes. Water. 2021; 13(21):3134. https://doi.org/10.3390/w13213134
Chicago/Turabian StyleNeplyukhina, Alisa A., Andrey V. Tchabovsky, Maria A. Gololobova, Olesya I. Smyshlyaeva, Olga A. Krylovich, Arkady B. Savinetsky, and Alexey A. Kotov. 2021. "The Holocene History of the Diatom Community in a Small Water Body on Shemya Island (Aleutian Arc, USA): The Influence of Global and Local Environmental Changes" Water 13, no. 21: 3134. https://doi.org/10.3390/w13213134