
Characterization and Polyculture Analysis of Microalgae Strains Based on Biomass Production and Nutrient Consumption, and Bacterial Community in Municipal Wastewater


Abstract
:1. Introduction
2. Materials and Methods
2.1. Microalgae Strains and Proliferation Culture
2.2. Simulated Municipal Wastewater Culture Medium
2.3. Microalgal Culture Conditions
2.4. Analytical Methods
2.4.1. Biomass Analysis
2.4.2. Nitrogen and Phosphorus Analysis
2.5. Amplicon Sequencing and Data Analysis
3. Result and Discussion
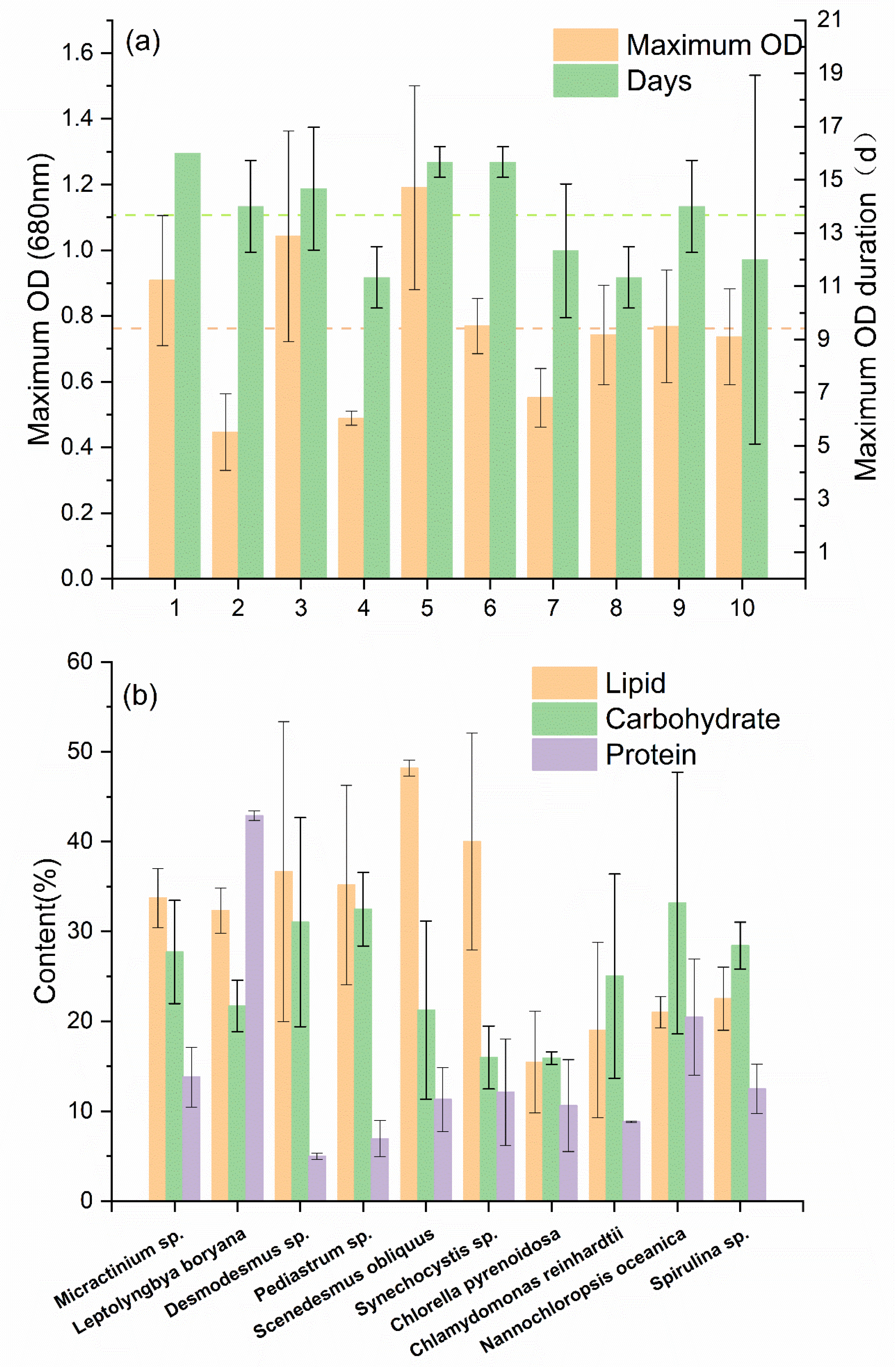
3.1. Biomass of Microalgae
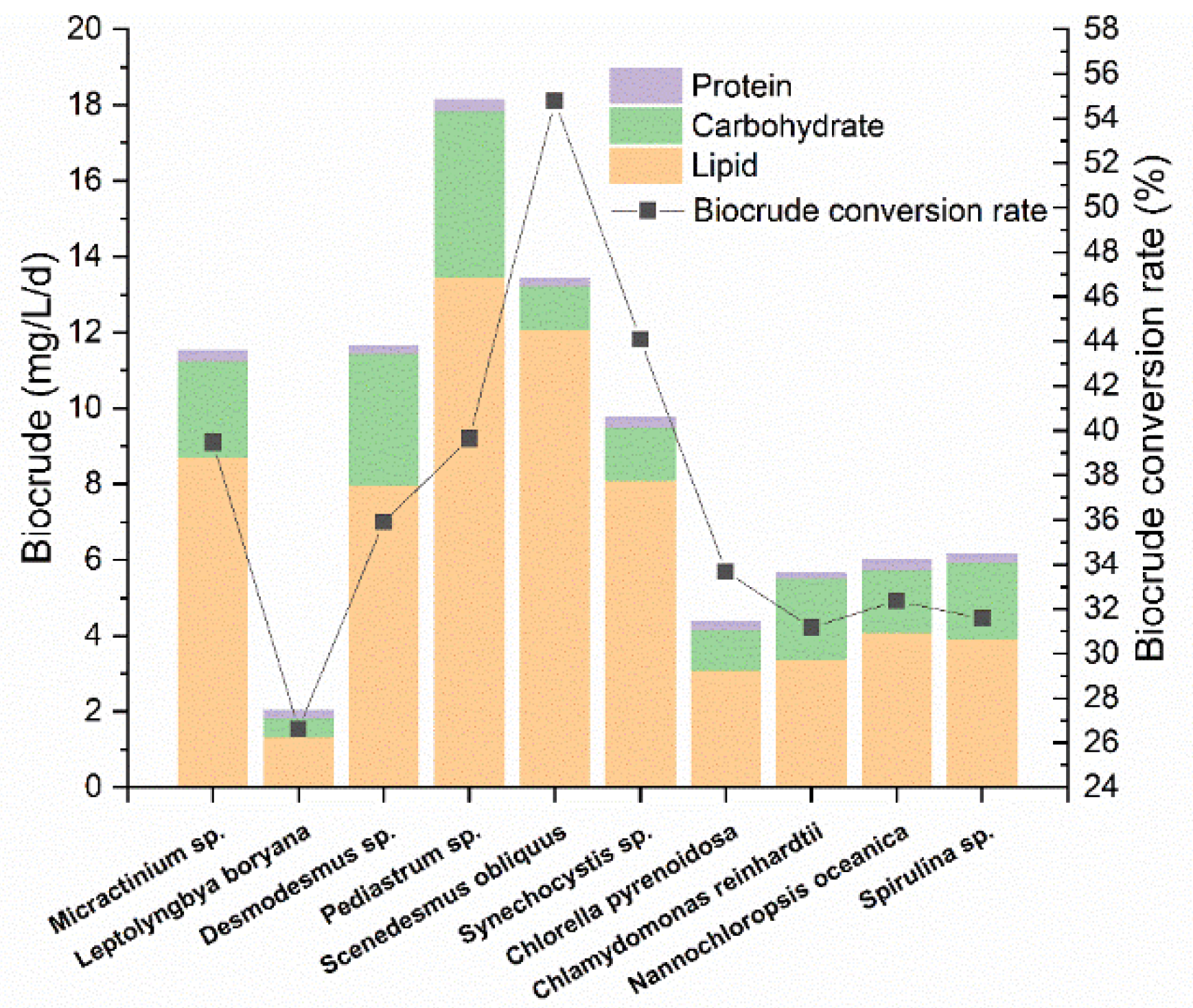
3.2. Potential as Biocrude
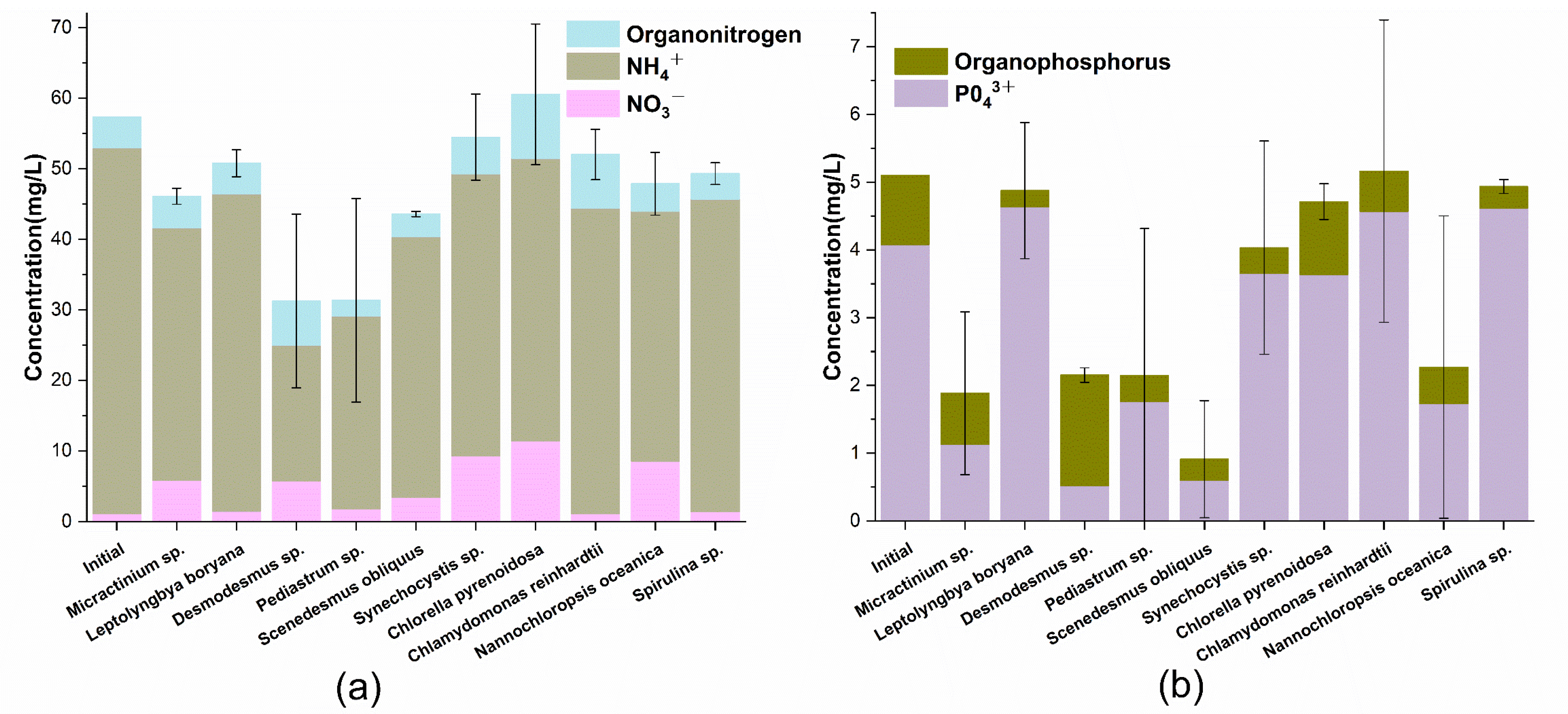
3.3. Nitrogen and Phosphorus Consumption
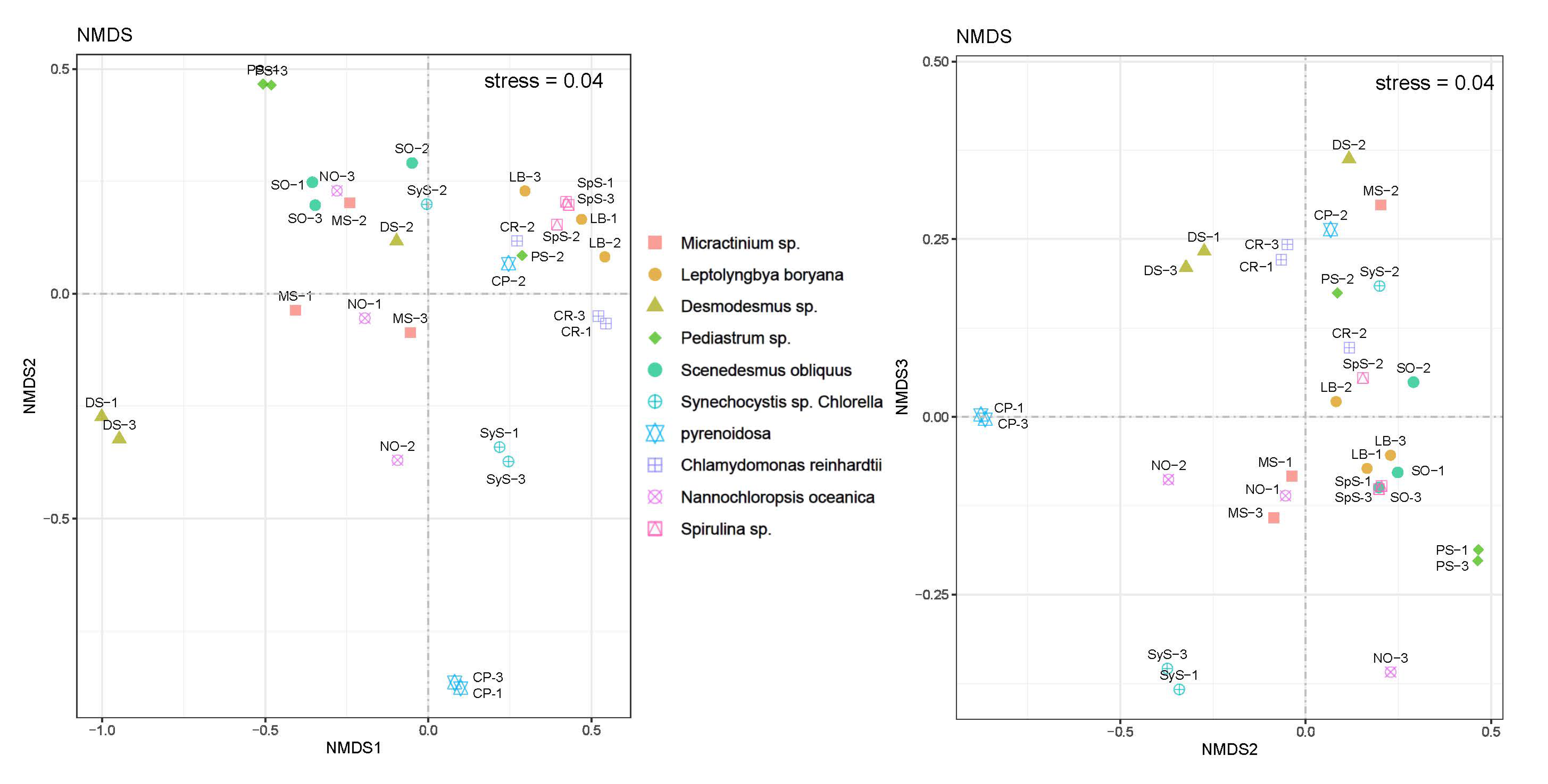
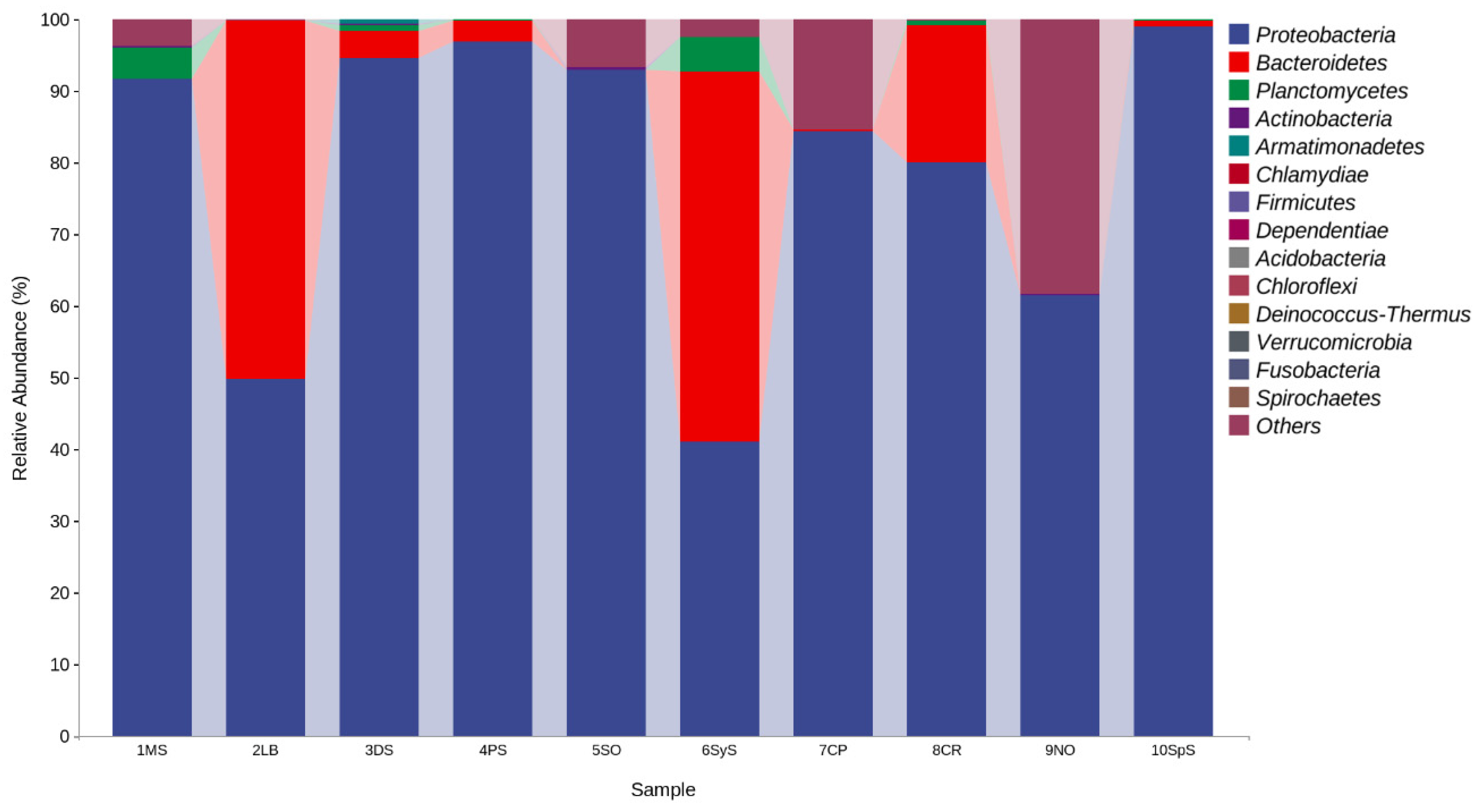
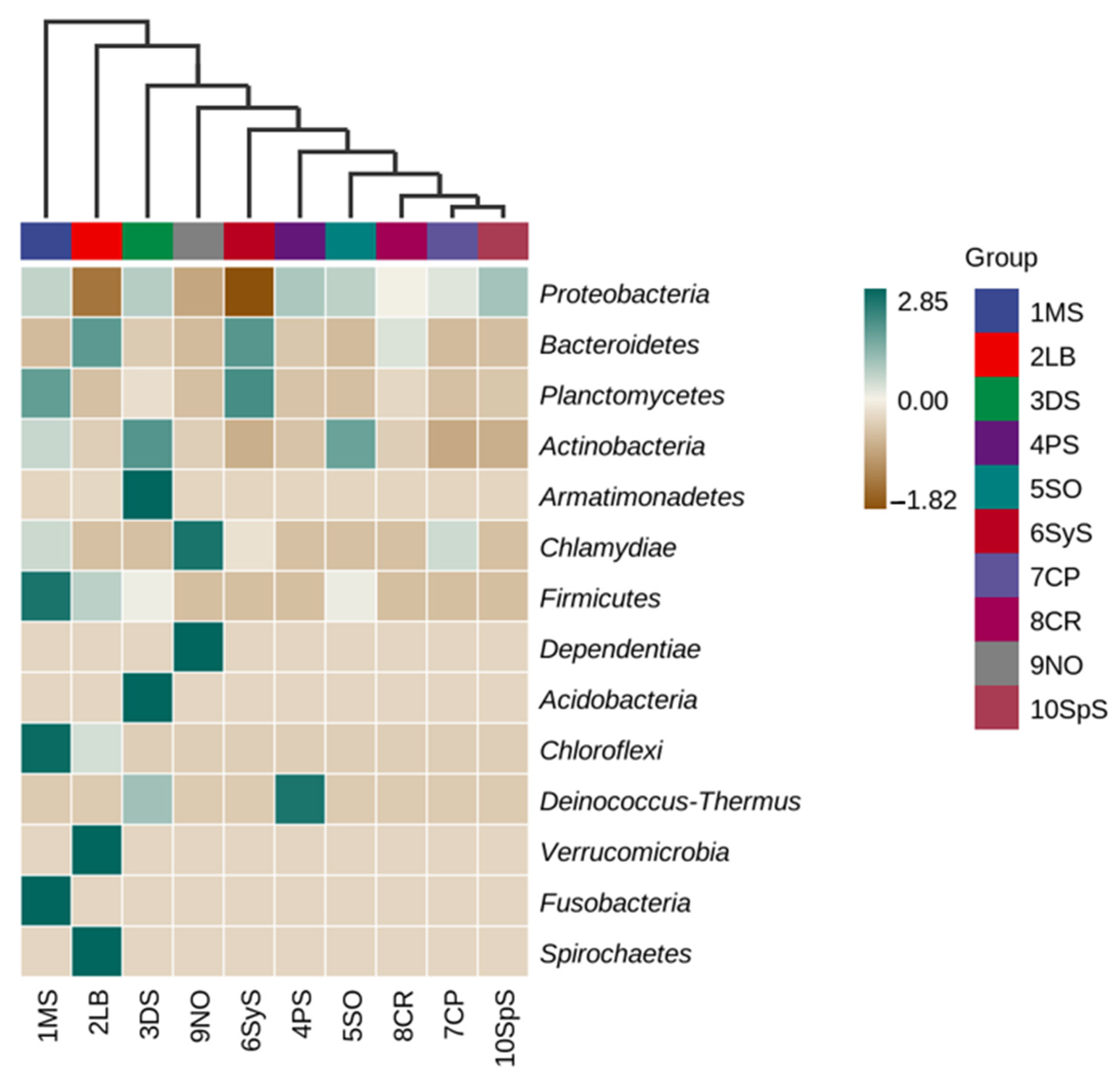
3.4. Bacterial Community Structure
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dudley, B. BP Statistical Review of World Energy; BP Statistical Review: London, UK, 2018; pp. 51–53. [Google Scholar]
- Brennan, L.; Owende, P. Biofuels from microalgae—A review of technologies for production, processing, and extractions of biofuels and co-products. Renew. Sustain. Energy Rev. 2010, 14, 557–577. [Google Scholar] [CrossRef]
- Chen, G.; Zhao, L.; Qi, Y. Enhancing the productivity of microalgae cultivated in wastewater toward biofuel production: A critical review. Appl. Energy 2015, 137, 282–291. [Google Scholar] [CrossRef]
- Rosenberg, J.N.; Mathias, A.; Korth, K.; Betenbaugh, M.J.; Oyler, G.A. Microalgal biomass production and carbon dioxide sequestration from an integrated ethanol biorefinery in Iowa: A technical appraisal and economic feasibility evaluation. Biomass Bioenergy 2011, 35, 3865–3876. [Google Scholar] [CrossRef]
- Liang, M.-H.; Jiang, J.-G. Advancing oleaginous microorganisms to produce lipid via metabolic engineering technology. Prog. Lipid Res. 2013, 52, 395–408. [Google Scholar] [CrossRef] [PubMed]
- Chisti, Y. Constraints to commercialization of algal fuels. J. Biotechnol. 2013, 167, 201–214. [Google Scholar] [CrossRef]
- Lam, M.K.; Lee, K.T. Microalgae biofuels: A critical review of issues, problems and the way forward. Biotechnol. Adv. 2012, 30, 673–690. [Google Scholar] [CrossRef]
- Godwin, C.M.; Lashaway, A.R.; Hietala, D.C.; Savage, P.E.; Cardinale, B.J. Biodiversity improves the ecological design of sustainable biofuel systems. GCB Bioenergy 2018, 10, 752–765. [Google Scholar] [CrossRef]
- Tumilar, A.S.; Milani, D.; Cohn, Z.; Florin, N.; Abbas, A. A Modelling Framework for the Conceptual Design of Low-Emission Eco-Industrial Parks in the Circular Economy: A Case for Algae-Centered Business Consortia. Water 2021, 13, 69. [Google Scholar] [CrossRef]
- Choi, W.J.; Chae, A.N.; Song, K.G.; Park, J.; Lee, B.C. Effect of trophic conditions on microalga growth, nutrient removal, algal organic matter, and energy storage products in Scenedesmus (Acutodesmus) obliquus KGE-17 cultivation. Bioprocess Biosyst. Eng. 2019, 42, 1225–1234. [Google Scholar] [CrossRef]
- Elser, J.J.; Fagan, W.F.; Denno, R.F.; Dobberfuhl, D.R.; Folarin, A.; Huberty, A.; Interlandi, S.; Kilham, S.S.; McCauley, E.; Schulz, K.L.; et al. Nutritional constraints in terrestrial and freshwater food webs. Nature 2000, 408, 578–580. [Google Scholar] [CrossRef]
- Mollamohammada, S.; Aly Hassan, A.; Dahab, M.; Kumar, S. A Hybrid Biological-Adsorption Approach for the Treatment of Contaminated Groundwater Using Immobilized Nanoclay-Algae Mixtures. Water 2021, 13, 633. [Google Scholar] [CrossRef]
- Georgianna, D.R.; Mayfield, S.P. Exploiting diversity and synthetic biology for the production of algal biofuels. Nature 2012, 488, 329–335. [Google Scholar] [CrossRef]
- Newby, D.T.; Mathews, T.J.; Pate, R.C.; Huesemann, M.H.; Lane, T.W.; Wahlen, B.D.; Mandal, S.; Engler, R.K.; Feris, K.P.; Shurin, J.B. Assessing the potential of polyculture to accelerate algal biofuel production. Algal Res. 2016, 19, 264–277. [Google Scholar] [CrossRef] [Green Version]
- Tian, Y.-T.; Wang, X.; Cui, Y.-H.; Wang, S.-K. A symbiotic yeast to enhance heterotrophic and mixotrophic cultivation of Chlorella pyrenoidosausing sucrose as the carbon source. Bioprocess Biosyst. Eng. 2020, 43, 2243–2252. [Google Scholar] [CrossRef]
- Shurin, J.B.; Abbott, R.L.; Deal, M.S.; Kwan, G.T.; Litchman, E.; McBride, R.C.; Mandal, S.; Smith, V.H. Industrial-strength ecology: Trade-offs and opportunities in algal biofuel production. Ecol. Lett. 2013, 16, 1393–1404. [Google Scholar] [CrossRef] [Green Version]
- Godwin, C.M.; Hietala, D.C.; Lashaway, A.R.; Narwani, A.; Savage, P.E.; Cardinale, B.J. Ecological Stoichiometry Meets Ecological Engineering: Using Polycultures to Enhance the Multifunctionality of Algal Biocrude Systems. Environ. Sci. Technol. 2017, 51, 11450–11458. [Google Scholar] [CrossRef]
- Bell, T.A.S.; Doig, L.; Peyton, B.M.; Gerlach, R.; Fields, M.W. Contributions of the microbial community to algal biomass and biofuel productivity in a wastewater treatment lagoon system. Algal Res. 2019, 39, 101461. [Google Scholar] [CrossRef]
- Mhedhbi, E.; Khelifi, N.; Foladori, P.; Smaali, I. Real-Time Behavior of a Microalgae–Bacteria Consortium Treating Wastewater in a Sequencing Batch Reactor in Response to Feeding Time and Agitation Mode. Water 2020, 12, 1893. [Google Scholar] [CrossRef]
- Aubakirova, M.; Krupa, E.; Mazhibayeva, Z.; Isbekov, K.; Assylbekova, S. The Role of External Factors in the Variability of the Structure of the Zooplankton Community of Small Lakes (South-East Kazakhstan). Water 2021, 13, 962. [Google Scholar] [CrossRef]
- Shetty, P.; Boboescu, I.Z.; Pap, B.; Wirth, R.; Kovács, K.L.; Bíró, T.; Futó, Z.; White, R.A.; Maróti, G. Exploitation of Algal-Bacterial Consortia in Combined Biohydrogen Generation and Wastewater Treatment. Front. Energy Res. 2019, 7, 230. [Google Scholar] [CrossRef]
- Stanier, R.Y.; Kunisawa, R.; Mandel, M.; Cohenbaz, G. Purification and Properties of Unicellular Blue-green Algae. Bacteriol. Rev. 1971, 35, 171–205. [Google Scholar] [CrossRef]
- Mehrabadi, A.; Farid, M.M.; Craggs, R. Potential of five different isolated colonial algal species for wastewater treatment and biomass energy production. Algal Res. 2017, 21, 1–8. [Google Scholar] [CrossRef]
- Cho, D.-H.; Choi, J.-W.; Kang, Z.; Kim, B.-H.; Oh, H.-M.; Kim, H.-S.; Ramanan, R. Microalgal diversity fosters stable biomass productivity in open ponds treating wastewater. Sci. Rep. 2017, 7, 1979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerber, L.N.; Tester, J.W.; Beal, C.M.; Huntley, M.E.; Sills, D.L. Target Cultivation and Financing Parameters for Sustainable Production of Fuel and Feed from Microalgae. Environ. Sci. Technol. 2016, 50, 3333–3341. [Google Scholar] [CrossRef]
- Msanne, J.; Polle, J.; Starkenburg, S. An assessment of heterotrophy and mixotrophy in Scenedesmus and its utilization in wastewater treatment. Algal Res. 2020, 48, 101911. [Google Scholar] [CrossRef]
- Branco Dos Santos, F.; Du, W.; Hellingwerf, K.J. Synechocystis: Not Just a Plug-Bug for CO2, but a Green E. coli. Front. Bioeng. Biotechnol. 2014, 2, 36. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Han, D.; Yoon, K.; Hu, Q.; Li, Y. Characterization of type 2 diacylglycerol acyltransferases in Chlamydomonas reinhardtii reveals their distinct substrate specificities and functions in triacylglycerol biosynthesis. Plant J. 2016, 86, 3–19. [Google Scholar] [CrossRef] [Green Version]
- Biondi, N.; Bassi, N.; Chini Zittelli, G.; de Faveri, D.; Giovannini, A.; Rodolfi, L.; Allevi, C.; Macrì, C.; Tredici, M.R. Nannochloropsis sp. F&M-M24: Oil production, effect of mixing on productivity and growth in an industrial wastewater. Environ. Prog. Sustain. Energy 2013, 32, 846–853. [Google Scholar] [CrossRef]
- Hupfauf, B.; Süß, M.; Dumfort, A.; Fuessl-Le, H. Cultivation of Microalgae in Municipal Wastewater and Conversion by Hydrothermal Carbonization: A Review. ChemBioEng Rev. 2016, 3, 186–200. [Google Scholar] [CrossRef]
- Mark Ibekwe, A.; Murinda, S.E.; Murry, M.A.; Schwartz, G.; Lundquist, T. Microbial community structures in high rate algae ponds for bioconversion of agricultural wastes from livestock industry for feed production. Sci. Total Environ. 2017, 580, 1185–1196. [Google Scholar] [CrossRef] [Green Version]
- Beigbeder, J.-B.; Boboescu, I.Z.; Damay, J.; Duret, X.; Bhatti, S.; Lavoie, J.-M. Phytoremediation of bark-hydrolysate fermentation effluents and bioaccumulation of added-value molecules by designed microalgal consortia. Algal Res. 2019, 42, 101585. [Google Scholar] [CrossRef]
- Matyash, V.; Liebisch, G.; Kurzchalia, T.V.; Shevchenko, A.; Schwudke, D. Lipid extraction by methyl-tert-butyl ether for high-throughput lipidomics. J. Lipid Res. 2008, 49, 1137–1146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Rausch, T. The estimation of micro-algal protein content and its meaning to the evaluation of algal biomass I. Comparison of methods for extracting protein. Hydrobiologia 1981, 78, 237–251. [Google Scholar] [CrossRef]
- Bradford, M.M. Bacteriological Reviewsrapid and Sensitive Method for Quantitation of Microgram Quantities of Protein Utilizing Principle of Protein-dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Biller, P.; Ross, A.B. Potential yields and properties of oil from the hydrothermal liquefaction of microalgae with different biochemical content. Bioresour. Technol. 2011, 102, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Fusheng, W. Water and Wastewater Monitoring and Analysis Methods, 4th ed.; China Environmental Publishing Group: Beijing, China, 2002; pp. 88–697. [Google Scholar]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat Biotechnol 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Gregory Caporaso, J. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef] [PubMed]
- Ye, S.; Gao, L.; Zhao, J.; An, M.; Wu, H.; Li, M. Simultaneous wastewater treatment and lipid production by Scenedesmus sp. HXY2. Bioresour. Technol. 2020, 302, 122903. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Min, M.; Li, Y.; Hu, B.; Ma, X.; Cheng, Y.; Liu, Y.; Chen, P.; Ruan, R. A hetero-photoautotrophic two-stage cultivation process to improve wastewater nutrient removal and enhance algal lipid accumulation. Bioresour. Technol. 2012, 110, 448–455. [Google Scholar] [CrossRef]
- Li, Z.; Savage, P.E. Feedstocks for fuels and chemicals from algae: Treatment of crude bio-oil over HZSM-5. Algal Res. 2013, 2, 154–163. [Google Scholar] [CrossRef]
- Ross, A.B.; Jones, J.M.; Kubacki, M.L.; Bridgeman, T. Classification of macroalgae as fuel and its thermochemical behaviour. Bioresour. Technol. 2008, 99, 6494–6504. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Ning, K.; Zeng, X.; Luo, Y.; Wang, D.; Hu, J.; Li, J.; Xu, H.; Huang, J.; Wan, M.; et al. Genomic Foundation of Starch-to-Lipid Switch in Oleaginous Chlorella spp. Plant Physiol. 2015, 169, 2444–2461. [Google Scholar] [CrossRef] [Green Version]
- Perez-Garcia, O.; Escalante, F.M.E.; de-Bashan, L.E.; Bashan, Y. Heterotrophic cultures of microalgae: Metabolism and potential products. Water Res. 2011, 45, 11–36. [Google Scholar] [CrossRef] [PubMed]
- Tian, B.; Hua, Y. Carotenoid biosynthesis in extremophilic Deinococcus-Thermus bacteria. Trends Microbiol. 2010, 18, 512–520. [Google Scholar] [CrossRef]
- Zhang, T.; Shao, M.-F.; Ye, L. 454 pyrosequencing reveals bacterial diversity of activated sludge from 14 sewage treatment plants. ISME J. 2012, 6, 1137–1147. [Google Scholar] [CrossRef] [PubMed]
- Vik, U.; Logares, R.; Blaalid, R.; Halvorsen, R.; Carlsen, T.; Bakke, I.; Kolstø, A.-B.; Økstad, O.A.; Kauserud, H. Different bacterial communities in ectomycorrhizae and surrounding soil. Sci. Rep. 2013, 3, 3471. [Google Scholar] [CrossRef] [Green Version]
- Kalam, S.; Basu, A.; Ahmad, I.; Sayyed, R.Z.; El-Enshasy, H.A.; Dailin, D.J.; Suriani, N.L. Recent Understanding of Soil Acidobacteria and Their Ecological Significance: A Critical Review. Front. Microbiol. 2020, 11, 580024. [Google Scholar] [CrossRef]
- Shu, Q. Molecular progresses of marine Planctomycetes: A review. Afr. J. Microbiol. Res. 2011, 5, 6018–6023. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Latin Species Name | FACHB-Number * | Reference |
|---|---|---|---|
| 1 | Micractinium sp. | 1267 | [23] |
| 2 | Leptolyngbya boryana | 2210 | [24] |
| 3 | Desmodesmus sp. | 2042 | [25] |
| 4 | Pediastrum sp. | 931 | [23] |
| 5 | Scenedesmus obliquus | 13 | [26] |
| 6 | Synechocystis sp. | 898 | [27] |
| 7 | Chlorella pyrenoidosa | 5 | [24] |
| 8 | Chlamydomonas reinhardtii | 359 | [28] |
| 9 | Nannochloropsis oceanica | 926 | [29] |
| 10 | Spirulina sp. | 810 | [30] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, W.; Fu, D.; Pan, T.; Singh, R.P. Characterization and Polyculture Analysis of Microalgae Strains Based on Biomass Production and Nutrient Consumption, and Bacterial Community in Municipal Wastewater. Water 2021, 13, 3190. https://doi.org/10.3390/w13223190
Liu W, Fu D, Pan T, Singh RP. Characterization and Polyculture Analysis of Microalgae Strains Based on Biomass Production and Nutrient Consumption, and Bacterial Community in Municipal Wastewater. Water. 2021; 13(22):3190. https://doi.org/10.3390/w13223190
Chicago/Turabian StyleLiu, Weixing, Dafang Fu, Ting Pan, and Rajendra Prasad Singh. 2021. "Characterization and Polyculture Analysis of Microalgae Strains Based on Biomass Production and Nutrient Consumption, and Bacterial Community in Municipal Wastewater" Water 13, no. 22: 3190. https://doi.org/10.3390/w13223190