
The Colonization of a Cold Spring Ecosystem by the Invasive Species Potamopyrgus antipodarum (Gray, 1843) (Gastropoda: Tateidae) (Southern Poland)


Abstract
:1. Introduction
2. Material and Methods
2.1. Study Area
2.2. Sampling Procedure
2.3. Data Analysis
3. Results
3.1. Water Chemistry
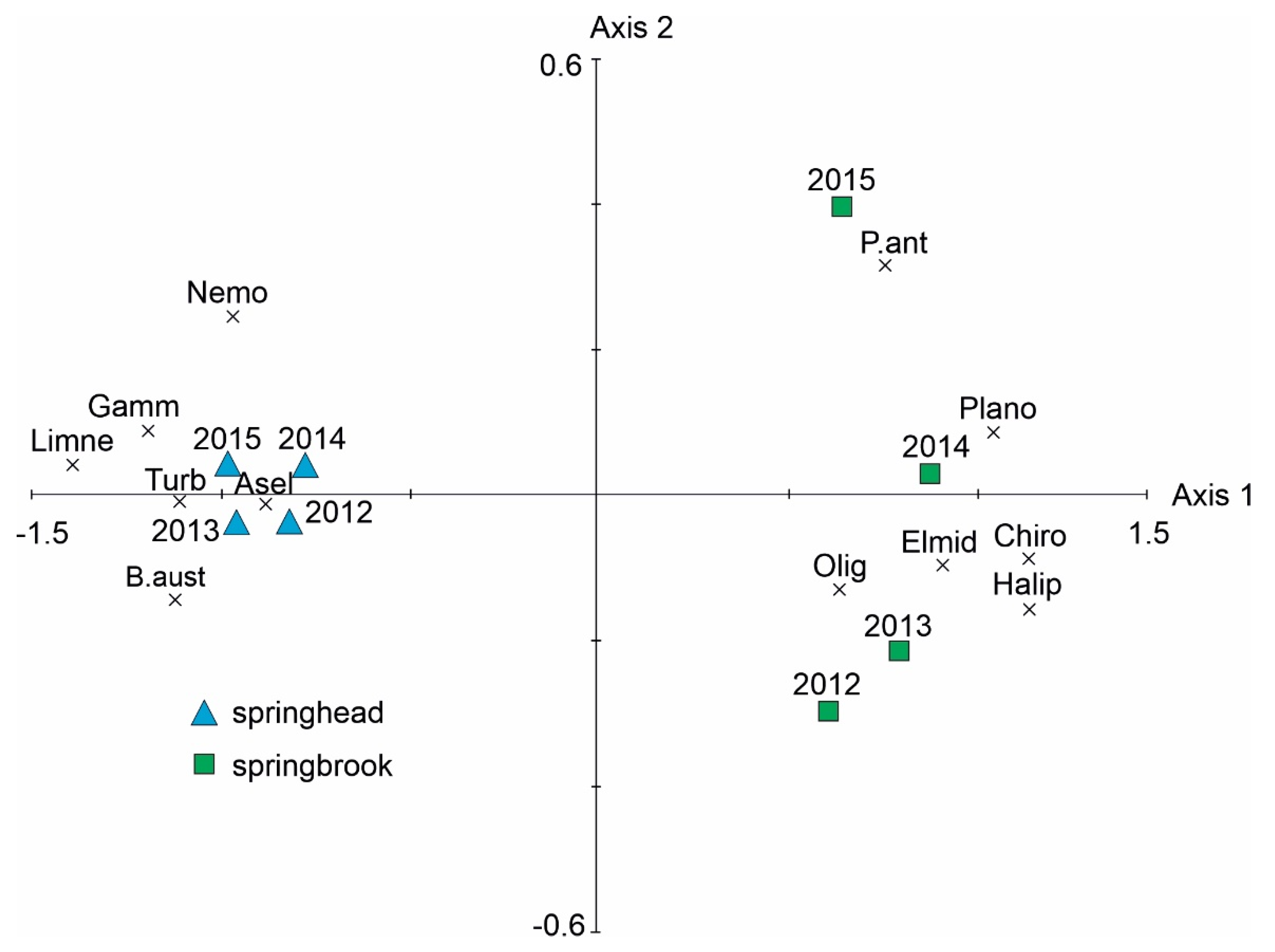
3.2. Macroinvertebrate Communities
3.3. Characteristic of the Potamopyrgus Antipodarum Populations
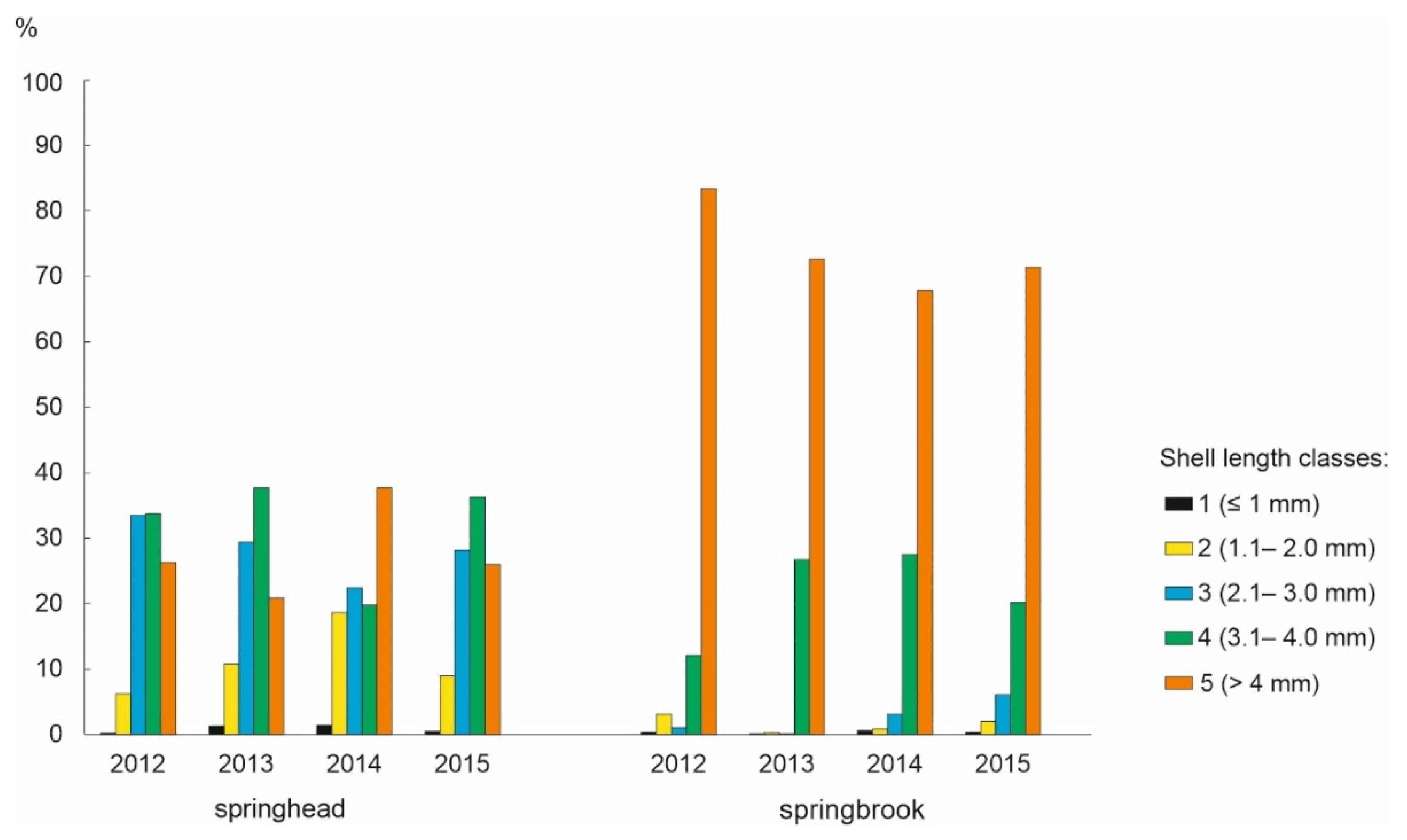
3.3.1. Size-Class Distribution
3.3.2. Fecundity of Potamopyrgus Antipodarum
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cantonati, M.; Gerecke, R.; Bertuzzi, E. Springs of the Alps—Sensitive ecosystems to environmental change: From biodiversity assessments to long-term studies. Hydrobiologia 2006, 562, 59–96. [Google Scholar] [CrossRef]
- Scarsbrook, M.; Barquĺn, J.; Gray, D. New Zealand Coldwater Springs and Their Biodiversity; Science for Conservation 278; Department of Conservation: Wellington, New Zealand, 2007. [Google Scholar]
- Glazier, D.S. Springs. In Encyclopedia of Inland Waters; Likens, G.E., Ed.; Academic Press Elsevier: Oxford, UK, 2009; Volume 1, pp. 734–755. [Google Scholar]
- McCabe, D.J. Biological communities in Springbrooks. In Studies in Crenobiology. The Biology of Springs and Springbrooks; Botosaneanu, L., Ed.; Backhuys Publishers: Leiden, The Netherlands, 1998. [Google Scholar]
- Ward, J.V.; Tockner, K. Biodiversity: Towards a unifying theme for river ecology. Freshw. Biol. 2001, 46, 807–819. [Google Scholar] [CrossRef]
- Gerecke, R.; Franz, H. Quellen im Nationalpark Berchtesgaden. Lebensgemeinschaften als Indikatoren des Klimawandels; Forschungsbericht 51; Nationalpark Berchtesgaden: Berchtesgaden, Germany, 2006. [Google Scholar]
- Cantonati, M.; Fuereder, L.; Gerecke, R.; Juettner, I.; Cox, E.J. Crenic habitats, hotspots for freshwater biodiversity conservation: Toward an understanding of their ecology. Freshw. Sci. 2012, 31, 463–480. [Google Scholar] [CrossRef]
- Cantonati, M.; Lange-Bertalot, H.; Scalfi, A.; Angeli, N. Cymbella tridentina sp. nov. (Bacillariophyta), a crenophilous diatom from carbonate springs of the Alps. J. N. Am. Benthol. Soc. 2010, 29, 775–788. [Google Scholar] [CrossRef]
- Ilmonen, J.; Mykrä, H.; Virtanen, R.; Paasivirta, L.; Muotka, T. Responses of spring macroinvertebrate and bryophyte communities to habitat modification: Community composition, species richness, and red-listed species. Freshw. Sci. 2012, 31, 657–667. [Google Scholar] [CrossRef]
- Martin, P.; Brunke, M. Faunal typology of lowland springs in Northern Germany. Freshw. Sci. 2012, 31, 542–562. [Google Scholar] [CrossRef]
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.; Knowler, D.; Lévêque, C.; Naiman, R.J.; Prieur-Richard, A.H.; Soto, D.; Stiassny, M.L.J.; et al. Freshwater biodiversity: Importance, threats, status, and conservation challenges. Biol. Rev. 2006, 81, 163–182. [Google Scholar] [CrossRef]
- Lindegaard, C.; Brodersen, K.P.; Wiberg-Larsen, P.; Skriver, J. Multivariate analyses of macrofaunal communities in Danish springs and Springbrooks. In Studies in Crenobiology: The Biology of Springs and Springbrooks; Botosaneanu, L., Ed.; Backhuys Publishers: Leiden, The Netherlands, 1998. [Google Scholar]
- Di Sabatino, A.; Cicolani, B.; Gerecke, R. Biodiversity and distribution of water mites (Acari, Hydrachnidia) in spring habitats. Freshw. Biol. 2003, 48, 2163–2173. [Google Scholar] [CrossRef]
- Minckley, W.L. Ecological review and management recommendations for recovery of the endangered Gila topminnow. Great Basin Nat. 1999, 59, 230–244. [Google Scholar]
- Ponder, W.F. Endemic aquatic macroinvertebrates of artesian springs of the Great Artesian Basin—Progress and future directions. Rec. South Aus. Mus. 2003, 7, 101–110. [Google Scholar]
- Minckley, W.L.; Unmack, P.J. Western springs: Their faunas, and threats to their existence. In Freshwater Ecoregions of North America; Abell, R.A., Olson, D.M., Dinerstein, E., Hurley, P.T., Diggs, J.T., Eichbaum, W., Walters, S., Wettengel, W., Allnutt, T., Loucks, C., et al., Eds.; Island Press: Washington, DC, USA, 2000. [Google Scholar]
- Gervasio, V.; Berg, D.J.; Lang, B.K.; Allan, N.L.; Guttman, S.I. Genetic diversity in the Gammarus pecos species complex: Implications for conservation and regional biogeography in the Chihuahuan Desert. Limnol. Oceanogr. 2004, 49, 520–531. [Google Scholar] [CrossRef] [Green Version]
- Meyer, J.L.; Strayer, D.L.; Wallace, J.B.; Eggert, S.L.; Helfman, G.S.; Leonard, N.E. The contribution of headwater streams to biodiversity in river networks. J. Am. Water Resour. Assoc. 2007, 43, 86–103. [Google Scholar] [CrossRef] [Green Version]
- Bogan, M.T.; Noriega-Felix, N.; Vidal-Aguilar, S.L.; Findley, L.T.; Lytle, D.A.; GutiérrezRuacho, O.G.; Alvarado-Castro, J.A.; Varela-Romero, A. Biogeography and conservation of aquatic fauna in spring-fed tropical canyons of the southern Sonoran Desert, Mexico. Biodivers. Conserv. 2014, 23, 2705–2748. [Google Scholar] [CrossRef]
- Cantonati, M. Diatom communities of springs in the Southern Alps. Diatom Res. 1998, 13, 201–220. [Google Scholar] [CrossRef]
- Cantonati, M.; Spitale, D. The role of environmental variables in structuring epiphytic and epilithic diatom assemblages in springs and streams of the Dolomiti Bellunesi National Park (south-eastern Alps). Fundam. Appl. Limnol.—Arch. Hydrobiol. 2009, 174, 117–133. [Google Scholar] [CrossRef]
- Cantonati, M.; Lange-Bertalot, H. Diatom biodiversity of springs in the Berchtesgaden National Park (northern Alps, Germany), with the ecological and morphological characterization of two species new to science. Diatom Res. 2010, 25, 251–280. [Google Scholar] [CrossRef]
- Barquin, J.; Scarsbrook, M. Management and conservation strategies for coldwater springs. Aquat. Conserv. 2008, 18, 580–591. [Google Scholar] [CrossRef]
- Unmack, P.J.; Minckley, W.L. The demise of desert springs. In Aridland Springs in North America: Ecology and Conservation; Stevens, L.E., Meretsky, V.J., Eds.; University of Arizona Press: Tucson, AZ, USA, 2008. [Google Scholar]
- Davis, J.A.; Kerezsy, A.; Nicol, S. Springs: Conserving perennial water is critical in arid landscapes. Biol. Conserv. 2017, 211, 30–35. [Google Scholar] [CrossRef]
- Boulton, A.J.; Brock, M.A.; Robson, B.J.; Chambers, J.M.; Ryder, D.S.; Davis, J.A. Australian Freshwater Ecology; Wiley-Blackwell: Hoboken, NJ, USA, 2014. [Google Scholar]
- Sada, D.W.; Williams, J.E.; Silvey, J.C.; Halford, A.; Ramakka, J.; Summers, P.; Lewis, L. A Guide to Managing, Restoring, and Conserving Springs in the Western United States; Desert Research Institute: Reno, NV, USA, 2001. [Google Scholar]
- Ponder, W.F. Desert springs of the Australian Great Artesian Basin. In Proceedings of the Meeting on Spring-Fed Wetlands: Important Scientific and Cultural Resources of the Intermountain Region, Las Vegas, NV, USA, 7–9 May 2002; Sada, D.W., Sharpe, S.E., Eds.; Desert Research Institute: Reno, NV, USA, 2002; pp. 1–13. [Google Scholar]
- Young, R.; Crowe, A.; Strickland, R. Ecological Assessments of Spring-Fed Streams on the Wairau Plain; prepared for Marlborough District Council; Cawthron Institute: Nelson, New Zealand, 2002. [Google Scholar]
- Fensham, R.J.; Ponder, W.F.; Fairfax, R.J. Recovery Plan for the Community of Native Species Dependent on Natural Discharge of Groundwater from the Great Artesian Basin; Report to Department of the Environment, Water, Heritage and the Arts; Department of Environment and Resource Management: Brisbane, QLD, Australia, 2007. [Google Scholar]
- Meffe, G.K.; Hendrickson, D.A.; Minckley, W.L.; Rinne, J.N. Factors resulting in decline of the endangered Sonoran topminnow Poeciliopsis occidentalis (Atheriniformes: Poeciliidae) in the United States. Biol. Conerv. 1983, 25, 135–159. [Google Scholar] [CrossRef]
- Rosen, P.C.; Schwalbe, C.R. Bullfrogs: Introduced predators in southwestern wetlands. In Our Living Resources: A Report to the Nation on the Distribution, Abundance, and Health of US; LaRoe, E.T., Farris, G.S., Puckett, C.E., Doran, P.D., Mac, M.J., Eds.; US Department of the Interior—National Biological Service: Washington, DC, USA, 1995. [Google Scholar]
- Fairfax, R.; Fensham, R.; Wager, R.; Brooks, S.; Webb, A.; Unmack, P. Recovery of the red-finned blue-eye: An endangered fish from springs in the great Artesian Basin. Wildl. Res. 2007, 34, 156–166. [Google Scholar] [CrossRef] [Green Version]
- Kerezsy, A.; Fensham, R. Conservation of the endangered red-finned blue-eye, Scaturiginichthys vermeilipinnis, and control of alien eastern gambusia, Gambusia holbrooki, in a spring wetland complex. Mar. Freshw. Res. 2013, 64, 851–863. [Google Scholar] [CrossRef]
- Baillie, J.; Butcher, E. Priceless or Worthless? The World’s Most Threatened Species; Zoological Society of London: London, UK, 2012. [Google Scholar]
- Minckley, W.L.; Meffe, G.K.; Soltz, D.L. Conservation and management of short-lived fishes: The cyprinodonts. In Battle against Extinction: Native Fish Management in the American West; Minckley, W.L., Deacon, J.E., Eds.; University of Arizona Press: Tucson, AZ, USA, 1991. [Google Scholar]
- Pister, E.P. The Desert Fishes Council: Catalyst for change. In Battle against Extinction: Native Fish Management in the American West; Minckley, W.L., Deacon, J.E., Eds.; University of Arizona Press: Tucson, AZ, USA, 1991. [Google Scholar]
- Williams, J.E.; Bowman, D.B.; Brooks, J.E.; Echelle, A.A.; Edwards, R.J.; Hendrickson, D.A.; Landye, J.J. Endangered aquatic ecosytems in North American deserts with a list of vanishing fishes of the region. J. AZ-NV Acad. Sci. 1985, 20, 1–62. [Google Scholar]
- Contreras-Arquieta, A.; Guajardo-Martinez, G.; Contreras-Balderas, S. Thiara (Melanoides) Tuberculata (Müller, 1774) (Gastropoda: Thiaridae), su Probable Impacto Ecológico en México. Publ. Biol. FCB/UANL 1995, 8, 17–24. [Google Scholar]
- Pyšek, P.; Jarošík, V.; Hulme, P.E.; Kühn, I.; Wild, J.; Arianoutsou, M.; Bacher, S.; Chiron, F.; Didžiulis, V.; Essl, F.; et al. Disentangling the role of environmental and human pressures on biological invasions across Europe. Proc. Natl. Acad. Sci. USA 2010, 107, 12157–12162. [Google Scholar] [CrossRef] [Green Version]
- Nielson, K.G.; Gill, K.M.; Springer, A.E.; Ledbetter, J.D.; Stevens, L.E.; Rood, S.B. Springs ecosystems: Vulnerable ecological islands where environmental conditions, life history traits, and human disturbance facilitate non-native plant invasions. Biol. Invasions 2019, 21, 2963–2981. [Google Scholar] [CrossRef]
- Mack, R.N.; Lonsdale, W.M. Humans as global plant dispersers: Getting more than we bargained for. Bioscience 2001, 51, 95–102. [Google Scholar] [CrossRef] [Green Version]
- Alonso, A.; Castro-Díez, P. The exotic aquatic mud snail Potamopyrgus antipodarum (Hydrobiidae, Mollusca): State of the art of a worldwide invasion. Aquat. Sci. 2012, 74, 375–383. [Google Scholar] [CrossRef]
- Collado, G.A. Out of New Zealand: Molecular identification of the highly invasive freshwater mollusc Potamopyrgus antipodarum (Gray, 1843) in South America. Zool. Stud. 2014, 53, 70. [Google Scholar] [CrossRef] [Green Version]
- Taybi, A.F.; Mabrouki, Y.; Glöer, P. First record of the New Zealand mudsnail Potamopyrgus antipodarum (J.E. Gray, 1843) (Tateidae, Mollusca) in Africa. Graellsia 2021, 77, e140. [Google Scholar] [CrossRef]
- Nentwig, W.; Bacher, S.; Kumschick, S.; Pysěk, P.; Vilà, M. More than ‘‘100 worst’’ alien species in Europe. Biol. Invasions 2018, 20, 1611–1621. [Google Scholar] [CrossRef] [Green Version]
- Alonso, A.; Castro-Diez, P. What explains the invading success of the aquatic mud snail Potamopyrgus antipodarum (Hydrobiidae: Mollisca)? Hydrobiologia 2008, 614, 107–117. [Google Scholar] [CrossRef]
- Alonso, A.; Castro-Diez, P. Tolerance to air exposure of the new Zealand mudsnail Potamopyrgus antipodarum (Hydrobiidae, Mollusca) as a prerequisite to survival in overland translocation. NeoBiota 2012, 14, 67–74. [Google Scholar] [CrossRef]
- Alonso, A.; Valle-Torres, G.; Castro-Díez, P. Survival of an invasive aquatic snail to overland translocation in non-aquatic media: Implications for spreading. Limnologica 2016, 57, 60–65. [Google Scholar] [CrossRef]
- Gérard, C.; Blanc, A.; Costil, K. Potamopyrgus antipodarum (Mollusca:Hydrobiidae) in continental aquatic gastropod communities: Impact of salinity and trematode parasitism. Hydrobiologia 2003, 493, 167–172. [Google Scholar] [CrossRef]
- Kopp, K.; Jokela, J. Resistant invaders can convey benefits to native species. Oikos 2007, 116, 295–301. [Google Scholar] [CrossRef]
- Vinson, M.R.; Baker, M.A. Poor growth of rainbow trout fed New Zealand mud snails Potamopyrgus antipodarum. N. Am. J. Fish. Manag. 2008, 28, 701–709. [Google Scholar] [CrossRef]
- Verhaegen, G.; McElroy, K.E.; Bankers, L.; Neiman, M.; Haase, M. Adaptive phenotypic plasticity in a clonal invader. Ecol. Evol. 2018, 8, 4465–4483. [Google Scholar] [CrossRef]
- Verhaegen, G.; von Jungmeister, K.; Haase, M. Life history variation in space and time: Environmental and seasonal responses of a parthenogenetic invasive freshwater snail in northern Germany. Hydrobiologia 2021, 848, 2153–2168. [Google Scholar] [CrossRef]
- Zbikowski, J.; Zbikowska, E. Invaders of an invader: Trematodes in Potamopyrgus antipodarum in Poland. J. Invertebr. Pathol. 2009, 101, 67–70. [Google Scholar] [CrossRef]
- Cichy, A.; Marszewska, A.; Parzonko, J.; Zbikowski, J.; Zbikowska, E. Infection of Potamopyrgus antipodarum (Gray, 1843) (Gastropoda: Tateidae) by trematodes in Poland, including the first record of aspidogastrid acquisition. J. Invertebr. Pathol. 2017, 150, 32–34. [Google Scholar] [CrossRef]
- Adema, C.M.; Lun, C.-M.; Hanelt, B.; Seville, R.S. Digenean trematode infections of native freshwater snails and invasive Potamopyrgus antipodarum in the Grand Teton National Park/John D. Rockefeller Memorial Parkway Area. J. Parasitol. 2009, 95, 224–227. [Google Scholar] [CrossRef]
- McKenzie, V.J.; Hall, W.E.; Guralnick, R.P. New Zealand mudsnails (Potamopyrgus antipodarum) in Boulder Creek, Colorado: Environmental factors associated with fecundity of a parthenogenic invader. Can. J. Zool. 2013, 91, 30–36. [Google Scholar] [CrossRef]
- Gérard, C.; Dussart, G.B.J. Invader and invaded colonisation by, and of, Potamopyrgus antipodarum Gray (Mollusca, Hydrobiidae). In BCPC Symposium Proceedings No 80, Slugs & Snails: Agricultural, Veterinary and Environmental Perspectives; British Crop Protection Council: Alton, UK, 2003; pp. 281–286. [Google Scholar]
- Gérard, C.; Le Lannic, J. Establishment of a new host-parasite association between the introduced invasive species Potamopyrgus antipodarum (Smith) (Gastropoda) an Sanguinicola sp. Plehn (Trematoda) in Europe. J. Zool. 2003, 261, 213–216. [Google Scholar] [CrossRef]
- Evans, N.A.; Whitfield, P.J.; Dobson, A.P. Parasite utilization of a host community: The distribution and occurrence of metacercarial cysts of Echinoparyphium recurvatum (Digenea: Echinostomatidae) in seven species of mollusc at Harting Pond, Sussex. Parasitology 1981, 83, 1–12. [Google Scholar] [CrossRef]
- Schreiber, E.S.G.; Glaister, A.; Quinn, G.P.; Lake, P.S. Life history and population dynamics of the exotic snail Potamopyrgus antipodarum (Prosobranchia:Hydrobiidae) in Lake Purrumbete, Victoria, Australia. Mar. Freshw. Res. 1998, 49, 73–78. [Google Scholar] [CrossRef]
- Schreiber, E.S.G.; Lake, P.S.; Quinn, G.P. Facilitation of native stream fauna by an invading species? Experimental investigations of the interaction of the snail, Potamopyrgus antipodarum (Hydrobiidae) with native benthic fauna. Biol. Invasions 2002, 4, 317–325. [Google Scholar] [CrossRef]
- Kerans, B.L.; Dybdahl, M.F.; Gangloff, M.M.; Jannot, J.E. Potamopyrgus antipodarum: Distribution, density, and effects on native macroinvertebrate assemblages in the Greater Yellowstone ecosystem. J. North Am. Benthol. Soc. 2005, 24, 123–138. [Google Scholar] [CrossRef]
- Richards, D.C. Competition between the Threatened Bliss Rapids snail, Taylorconcha serpenticola (Hershler et al.) and the Invasive Aquatic Snail, Potamopyrgus antipodarum (Gray). Ph.D. Thesis, Montana State University, Bozeman, MT, USA, 2004. [Google Scholar]
- Riley, L.A.; Dybdahl, M.F.; Hall, R.O., Jr. Invasive species impact: Asymmetric interactions between invasive and endemic freshwater snails. J. North. Am. Benthol. Soc. 2008, 27, 509–520. [Google Scholar] [CrossRef]
- Collado, G.A.; Aguayo, K.P.; Cazzaniga, N.J.; Gutiérrez Gregoric, D.E.; De Lucıá, M.; Haase, M. Systematic evaluation of cryptic freshwater snails from central Chile, including the enigmatic Littoridina santiagensis (Gastropoda, Truncatelloidea). Eur. J. Taxon. 2019, 524, 1–15. [Google Scholar] [CrossRef]
- Collado, G.A.; Vidal, M.A.; Aguayo, K.P.; Méndez, M.A.; Valladares, M.A.; Cabrera, F.J.; Pastenes, L.; Gutiérrez Gregoric, D.E.; Puillandre, N. Morphological and molecular analysis of cryptic native and invasive freshwater snails in Chile. Sci. Rep. 2019, 9, 7846. [Google Scholar] [CrossRef]
- Múrria, C.; Bonada, N.; Prat, N. Effects of the invasive species Potamopyrgus antipodarum (Hydrobiidae, Mollusca) on community structure in a small Mediterranean stream. Fundam. Appl. Limnol. 2008, 71, 131–143. [Google Scholar] [CrossRef]
- Schmidlin, S.; Schmera, D.; Baur, B. Alien molluscs affect the composition and diversity of native macroinvertebrates in a sandy flat of Lake Neuchâtel, Switzerland. Hydrobiologia 2012, 679, 233–249. [Google Scholar] [CrossRef]
- Hall, R.O., Jr.; Tank, J.L.; Dybdahl, M.F. Exotic snails dominate nitrogen and carbon cycling in a highly productive stream. Front. Ecol. Environ. 2003, 1, 407–411. [Google Scholar] [CrossRef]
- Arango, C.P.; Riley, L.; Tank, J.L.; Hall, R.O., Jr. Herbivory by an invasive snail increases nitrogen fixation in a nitrogen-limited stream. Can. J. Fish. Aquat. Sci. 2009, 66, 1309–1317. [Google Scholar] [CrossRef]
- Bennett, D.M.; Dudley, T.L.; Cooper, S.D.; Sweet, S.S. Ecology of the invasive New Zealand mud snail, Potamopyrgus antipodarum (Hydrobiidae), in mediterranean-climate stream system. Hydrobiologia 2015, 746, 375–399. [Google Scholar] [CrossRef]
- Bruce, R.L.; Moffitt, C.M. Survival and passage of ingested New Zealand Mudsnails through the intestinal tract of rainbow trout. N. Am. J. Aquac. 2009, 71, 287–301. [Google Scholar] [CrossRef]
- Brenneis, V.E.F.; Sih, A.; de Rivera, C.E. Integration of an invasive consumer into an estuarine food web: Direct and indirect effects of the New Zealand mud snail. Oecologia 2011, 167, 169–179. [Google Scholar] [CrossRef] [Green Version]
- Rakauskas, V.; Butkus, R.; Merkytė, E. Consumption of the invasive New Zealand mud snail (Potamopyrgus antipodarum) by benthivorous predators in temperate lakes: A case study from Lithuania. Hydrobiologia 2016, 775, 213–230. [Google Scholar] [CrossRef]
- Schreiber, E.S.G.; Quinn, G.P.; Lake, P.S. Distribution of an alien aquatic snail in relation to flow variability, human activity and water quality. Freshw. Biol. 2003, 48, 951–961. [Google Scholar] [CrossRef]
- Bodon, M.; Cianfanelli, S.; Montanari, A. Mollusks of the Frasassi karstic complex and adjacent sulfidic spring. In Proceedings of the Frasassi Stygobionts and Their Sulfidic Environment. A Scientific Conference and Workshop: Abstracts, Genga, Italy, 10–13 September 2009; pp. 9–11. [Google Scholar]
- Moseley, M. Early British reports of aquatic mollusca living in underground water pipes. Cave Karst. Sci. 2002, 29, 123–124. [Google Scholar]
- Ross, R.M.; Lellis, W.A.; Benett, R.M.; Johnson, C.S. Landscape determinants of nonindigenous fish invasions. Biol. Invasions 2001, 3, 347–361. [Google Scholar] [CrossRef]
- Cada, C.A. Interactions between the Invasive New Zealand mudsnail, Potamopyrgus antipodarum, Baetid Mayflies, and Fish Predators. Ph.D. Thesis, University of Montana State, Bozeman, MT, USA, 2004. [Google Scholar]
- Richards, D.C.; O’Connell, P.; Shinn, D.C. Simple control method to limit spread of New Zealand mudsnail, Potamopyrgus antipodarum. N. Am. J. Fish. Manag. 2004, 24, 114–117. [Google Scholar] [CrossRef]
- Alonso, A. Valoración del efecto de la degradación ambiental sobre los macroinvertebrados bentónicos en la cabecera del río Henares. Ecosistemas, 2006; pp. 101–105. [Google Scholar]
- Hinz, W.; Boeters, H.D.; Guenther, A. On the freshwater mollusc fauna in the Spanish Province of Cuenca. Malakol. Abh. 1994, 17, 65–81. [Google Scholar]
- Kerney, M. Atlas of the Land and Freshwater Molluscs of Britain and Ireland; Harley Books Martins, Great Horskesley: Colchester, Essex, UK, 1999. [Google Scholar]
- Richards, D.C.; Cazier, L.D.; Lester, G.T. Spatial distribution of three snail species including the invader Potamopyrgus antipodarum in a freshwater spring. West. N. Am. Naturalist 2001, 6, 375–380. Available online: https://www.jstor.org/stable/41717182 (accessed on 5 November 2021).
- Dumnicka, E.; Galas, J.; Koperski, P. Benthic invertebrates in karst springs: Does substratum or location define communities? Int. Rev. Hydrobiol. 2007, 92, 452–464. [Google Scholar] [CrossRef]
- Koperski, P.; Dumnicka, E.; Galas, J. Abiotic parameters determining fauna composition in karstic springs. Pol. J. Ecol. 2011, 59, 153–163. [Google Scholar]
- Wilke, T.; Haase, M.; Hershler, R.; Liu, H.-P.; Misof, B.; Ponder, W. Pushing short DNA fragments to the limit: Phylogenetic relationships of ‘hydrobioid’ gastropods (Caenogastropoda: Rissooidea). Mol. Phylogenet. Evol. 2013, 66, 715–736. [Google Scholar] [CrossRef]
- Thienemann, A. Hydrobiologische Untersuchungen an Quellen. Arch. Hydrobiol. 1924, 14, 41. [Google Scholar]
- Springer, A.E.; Stevens, L.E. Spheres of discharge of springs. Hydrogeol. J. 2009, 17, 83–93. [Google Scholar] [CrossRef]
- Galas, J. Human impact on physical and chemical properties of springs from Cracow-Częstochowa Upland (Southern Poland). Pol. J. Ecol. 2005, 53, 329–341. [Google Scholar]
- Erman, N.A.; Erman, D.C. Spring permanence, Trichoptera species richness and the role of drought. J. Kans. Entomol. Soc. 1995, 68, 50–64. [Google Scholar]
- Von Fumetti, S.; Nagel, P.; Baltes, B. Where a springhead becomes a springbrook—A regional zonation of springs. Arch. Hydrobiol. 2007, 169, 37–48. [Google Scholar] [CrossRef]
- Glöer, P. Mollusca, I. Süsswassergastropoden. Nord- und Mitteleuropas Bestimmungsschlüssel, Lebensweise, Verbreitung; ConchBooks: Hackenheim, Germany, 2002. [Google Scholar]
- Eggers, T.O.; Martens, A. A key to the freshwater Amphipoda (Crustacea) of Germany. Lauterbornia 2001, 42, 1–68. [Google Scholar]
- Hermanowicz, W.; Dojlido, J.; Dożańska, W.; Koziorowski, B.; Zerbe, J. Fizyczno-Chemiczne Badania Wody i Ścieków; Arkady: Warszawa, Poland, 1999. [Google Scholar]
- Møller, V.; Forbes, V.E.; Depledge, M.H. Influence of acclimation and exposure temperature on the acute toxicity of cadmium to freshwater snail Potamopyrgus antipodarum (Hydrobiidae). Environ. Toxicol. Chem. 1994, 3, 1519–1524. [Google Scholar] [CrossRef]
- Richards, D.C. The New Zealand mud snail invades the Western United States. Aquat. Nuis. Species Dig. 2002, 4, 42–44. [Google Scholar]
- Gauch, H.G. Noise reduction by eigenvector ordinations. Ecology 1982, 63, 1643–1649. [Google Scholar] [CrossRef]
- Ter Braak, C.J.F.; Šmilauer, P. CANOCO Reference Manual and User’s Guide: Software for Ordination (Version 5.0); Microcomputer Power: Ithaca, NY, USA, 2012. [Google Scholar]
- Moffitt, C.M.; James, C.A. Response of New Zealand mudsnails Potamopyrgus antipodarum to freezing and near-freezing fluctuating water temperatures. Freshw. Sci. 2012, 31, 1035–1041. [Google Scholar] [CrossRef]
- Moffitt, C.M.; James, C.A. Dynamics of Potamopyrgus antipodarum infestations and seasonal water temperatures in a heavily used recreational watershed in intermountain North America. Aquat. Invasions 2012, 7, 193–202. [Google Scholar] [CrossRef]
- Hamada, K.; Takeda, N.; Tatara, Y.; Ogata, D.; Nakajima, M.; Sonohara, T.; Urabe, M. Habitat Description of Potamopyrgus antipodarum (Caenogastropoda: Hydrobiidae) in Some Areas of Japan: How Far Will It Spread? Venus 2013, 71, 61–79. [Google Scholar]
- Hall, R.O., Jr.; Dybdahl, M.F.; VanderLoop, M.C. Extremely high secondary production of introduced snails in rivers. Ecol. Appl. 2006, 16, 1121–1131. [Google Scholar] [CrossRef]
- Verhaegen, G.; Neiman, M.; Haase, M. Ecomorphology of a generalist freshwater gastropod: Complex relations of shell morphology, habitat, and fecundity. Org. Divers. Evol. 2018, 18, 425–441. [Google Scholar] [CrossRef]
- Dybdahl, M.F.; Kane, S.L. Adaptation vs. phenotypic plasticity in the success of a clonal invader. Ecology 2005, 86, 1592–1601. [Google Scholar] [CrossRef] [Green Version]
- Gust, M.; Buronfosse, T.; André, C.; Mons, R.; Gagné, F.; Garric, J. Is exposure temperature a confounding factor for the assessment of reproductive parameters of New Zealand mudsnails Potamopyrgus antipodarum (Gray)? Aquat. Toxicol. 2011, 101, 396–404. [Google Scholar] [CrossRef] [PubMed]
- Macken, A.; Le Page, G.; Hayfield, A.; Williams, T.D.; Brown, R.J. Effects of test design and temperature in a partial life-cycle study with the freshwater gastropod Potamopyrgus antipodarum. Environ. Toxicol. Chem. 2012, 31, 1989–1994. [Google Scholar] [CrossRef] [PubMed]
- Winterbourn, M.J. Water temperature as a factor limiting the distribution of Potamopyrgus antipodarum (Gastropoda, Prosobranchia) in the New Zealand thermal region. New Zealand J. Mar. Freshw. Res. 1969, 3, 453–458. [Google Scholar] [CrossRef]
- Hylleberg, J.; Siegismund, H.R. Niche overlap in mud snails (Hydrobiidae): Freezing tolerance. Mar. Biol. 1987, 94, 403–407. [Google Scholar] [CrossRef]
- Lewin, I. Occurrence of the Invasive Species Potamopyrgus antipodarum (Prosobranchia: Hydrobiidae) in Mining Subsidence Reservoirs in Poland in Relation to Environmental Factors. Malacologia 2012, 55, 15–31. [Google Scholar] [CrossRef]
- Coulaud, R.; Mouthon, J.; Quéau, H.; Charles, S.; Chaumot, A. Life-history phenology strongly influences population vulnerability to toxicants: A case study with the mudsnail Potamopyrgus antipodarum. Environ. Toxicol. Chem. 2013, 32, 721–1736. [Google Scholar] [CrossRef]
- Wallace, C. On the distribution of the sexes of Potamopyrgus jenkinsi (Smith). J. Mollusc. Stud. 1985, 51, 290–296. [Google Scholar] [CrossRef]
- Økland, J. Lakes and Snails; Universal Book Services/Dr. W. Backhuys: Oegstgeest, The Netherlands, 1990. [Google Scholar]
- Falniowski, A. A male of Potamopyrgus jenkinsi (Smith, 1889) in Poland. Basteria 1979, 43, 109–111. [Google Scholar]
- Tobin, P.C.; Berec, L.; Liebhold, A.M. Exploiting Allee effects for managing biological invasions. Ecol. Lett. 2011, 14, 615–624. [Google Scholar] [CrossRef]
- Havel, J.E.; Kovalenko, K.E.; Thomaz, S.M.; Amalfitano, S.; Kats, L.B. Aquatic invasive species: Challenges for the future. Hydrobiologia 2015, 750, 147–170. [Google Scholar] [CrossRef]
- Cuttelod, A.; Seddon, M.; Neubert, E. European Red List of Non-Marine Molluscs; Publications Office of the European Union: Luxembourg, 2011. [Google Scholar] [CrossRef]
- Sturm, R. Modelling ecological specifities of freshwater molluscs: The exemplary case of Bythinella austriaca (v. Frauenfeld, 1857) (Gastropoda, Prosobranchia). J. Limnol. 2016, 75, 626–633. [Google Scholar] [CrossRef] [Green Version]
- Sturm, R. Malacological mapping in Austria: Distribution of the Austrian spring snail Bythinella austriaca (v. FRAUENFELD, 1857) in the federal state of Salzburg. Linzer. Biol. Beitr. 2018, 50/1, 837–843. [Google Scholar]
- Sturm, R. Modelling optimum ranges of selected environmental variables for habitats colonized by the spring snail Bythinella austriaca (v. Frauenfeld, 1857) (Gastropoda, Prosobranchia). Malakol. Abh. 2005, 23, 67–76. [Google Scholar]
- Klockmann, M.; Scharre, M.; Haase, M.; Fischer, K. Does narrow niche space in a ‘cold-stenothermic’spring snail indicate high vulnerability to environmental change? Hydrobiologia 2016, 765, 71–83. [Google Scholar] [CrossRef]
- Nicol, S.; Haynes, T.B.; Fensham, R.; Kerezsy, A. Quantifying the impact of Gambusia holbrooki on the extinction risk of the critically endangered red-finned blue-eye. Ecosphere 2015, 6, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Box, J.B.; Duguid, A.; Read, R.; Kimber, R.; Knapton, A.; Davis, J.; Bowland, A. Central Australian waterbodies: The importance of permanence in a desert landscape. J. Arid Environ. 2008, 72, 1395–1413. [Google Scholar] [CrossRef]
- Fensham, R.J.; Silcock, J.; Kerezsy, A.; Ponder, W. Four desert waters: Setting arid zone wetland conservation priorities through understanding patterns of endemism. Biol. Conserv. 2011, 144, 2459–2467. [Google Scholar] [CrossRef]
- Rossini, R.A.; Fensham, R.J.; Walter, G.H. Determining optimal sampling strategies for monitoring threatened endemic macroinvertebrates in Australia’s artesian springs. Mar. Freshw. Res. 2015, 67, 653–665. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Description | |
|---|---|---|
| Type of a spring | valley, rheocrene, in the bottom of the pond | |
| Discharge (1 s−1) | 50 | |
| Drainage basin of a river | Przemsza | |
| Altitude | 290 m a.s.l. | |
| Geographical location | Katowice Upland | |
| Geographical coordinates | N 50°18.399′ E 19°19.422′ | |
| Management | natural monument named “Wywierzyska in Strzemieszyce Wielkie” | |
| Range of water temperature (°C) | 9.0–11.0 | |
| springhead | springbrook | |
| Location | within 1 m of the spring’s source | within 40 m of the source |
| Dominant type of substrate | loamy sand | loamy sand |
| Aquatic macrophytes | none | Sparganium erectum L., 1753 |
| Parameter | Site | Year | |||
|---|---|---|---|---|---|
| 2012 | 2013 | 2014 | 2015 | ||
| Temperature (°C) | SH | 10.1 | 9.9 | 10.0 | 10.1 |
| SB | 14.1 | 13.9 | 14.0 | 14.2 | |
| Dissolved oxygen (mg L −1) | SH | 7.7 | 7.8 | 7.9 | 7.7 |
| SB | 7.6 | 7.6 | 7.8 | 7.3 | |
| pH | SH | 7.2 | 7.3 | 7.6 | 7.3 |
| SB | 7.3 | 7.7 | 7.7 | 7.4 | |
| Conductivity (µS cm−1) | SH | 720 | 799 | 760 | 830 |
| SB | 770 | 798 | 740 | 810 | |
| Total dissolved solids (mg L −1) | SH | 460 | 520 | 370 | 410 |
| SB | 380 | 518 | 360 | 400 | |
| Chlorides (mg L −1) | SH | 60 | 40 | 42 | 46 |
| SB | 60 | 46 | 44 | 40 | |
| Nitrate nitrogen (mg L −1) | SH | 2.1 | 4.6 | 7.2 | 5 |
| SB | 4.2 | 4.4 | 5.2 | 8.4 | |
| Ammonium nitrogen (mg L −1) | SH | 0.16 | 0.09 | 0.5 | 0.1 |
| SB | 0.17 | 0.1 | 0.05 | 0.21 | |
| Phosphates (mg L −1) | SH | 0.14 | 0.11 | 0.08 | 0.08 |
| SB | 0.14 | 0.13 | 0.07 | 0.1 | |
| Total hardness (mgCaCO3 L −1) | SH | 435 | 501.9 | 405 | 355 |
| SB | 435 | 494.7 | 325 | 355 | |
| Calcium (mg L −1) | SH | 86.0 | 104.0 | 102.0 | 100.0 |
| SB | 86.0 | 108.0 | 92.0 | 120.0 | |
| Magnesium (mg L −1) | SH | 53.5 | 58.8 | 46.5 | 45.6 |
| SH | 53.5 | 54.7 | 53.1 | 53.4 | |
| Alkalinity (mgCaCO3 L −1) | SB | 285 | 250 | 235 | 290 |
| SH | 240 | 240 | 215 | 285 | |
| Iron (mg L −1) | SB | 0.02 | 0.05 | 0.02 | 0.01 |
| SB | 0.02 | 0.05 | 0.18 | 0.03 | |
| Higher Taxa | Family | Species | Springhead | Springbrook | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2012 | 2013 | 2014 | 2015 | 2012 | 2013 | 2014 | 2015 | |||||||||||
| n | % | n | % | n | % | n | % | n | % | n | % | n | % | n | % | |||
| Gastropoda | Tateidae | Potamopyrgus antipodarum (Gray, 1843) | 468 | 20.0 | 398 | 19.1 | 419 | 18.8 | 589 | 19.9 | 579 | 64.0 | 706 | 69.5 | 828 | 69.0 | 2303 | 86.1 |
| Bythinellidae | Bythinella cf. austriaca (v. Frauenfeld 1857) | 478 | 20.5 | 410 | 19.7 | 435 | 19.6 | 625 | 21.1 | 19 | 2.1 | 14 | 1.4 | 9 | 0.8 | 4 | 0.1 | |
| Lymnaeidae | Radix balthica (Linnaeus, 1758) | 5 | 0.4 | |||||||||||||||
| Stagnicola palustris (O. F. Müller, 1774) | 9 | 0.8 | 6 | 0.2 | ||||||||||||||
| Planorbidae | Anisus spirorbis (Linnaeus, 1758) | 2 | 0.2 | 4 | 0.4 | 2 | 0.2 | 13 | 0.5 | |||||||||
| Bivalvia | Sphaeriidae | 24 | 1.0 | 28 | 1.3 | 33 | 1.5 | 37 | 1.2 | 35 | 3.9 | 29 | 2.9 | 63 | 5.3 | 52 | 1.9 | |
| Crustacea | Gammaridae | Gammarus fossarum Koch, 1836 | 1123 | 48.1 | 943 | 45.3 | 1073 | 48.3 | 1358 | 45.8 | 51 | 5.6 | 58 | 5.7 | 67 | 5.6 | 65 | 2.4 |
| Asellidae | Asellus aquaticus (Linnaeus, 1758) | 5 | 0.2 | 6 | 0.3 | 4 | 0,2 | 7 | 0.2 | 3 | 0.3 | 4 | 0.4 | 2 | 0.2 | 3 | 0.1 | |
| Oligochaeta | 7 | 0.3 | 8 | 0.4 | 10 | 0.4 | 12 | 0.4 | 16 | 1.8 | 18 | 1.8 | 13 | 1.1 | 15 | 0.6 | ||
| Hirudinea | Erpobdellidae | 1 | 0.1 | 1 | 0.04 | |||||||||||||
| Plecoptera | Nemouridae | 3 | 0.1 | 2 | 0.1 | 2 | 0.1 | 2 | 0.1 | 1 | 0.04 | |||||||
| Trichoptera | Limnephilidae | 186 | 8.0 | 240 | 11.5 | 203 | 9.1 | 284 | 9.6 | 23 | 2.5 | 24 | 2.4 | 23 | 1.9 | 25 | 0.9 | |
| Hydroptilidae | 12 | 0.4 | ||||||||||||||||
| Coleoptera | Haliplidae | 3 | 0.3 | 4 | 0.4 | 4 | 0.3 | 2 | 0.1 | |||||||||
| Elmidae | 3 | 0.1 | 2 | 0.1 | 3 | 0.1 | 2 | 0.1 | 5 | 0.6 | 3 | 0.3 | 4 | 0.3 | 4 | 0.1 | ||
| Odonata | Coenagrionidae | 1 | 0.04 | |||||||||||||||
| Megaloptera | Sialidae | 1 | 0.04 | |||||||||||||||
| Diptera | Chironomidae | 25 | 1.1 | 27 | 1.3 | 29 | 1.3 | 28 | 0.9 | 169 | 18.7 | 151 | 14.9 | 171 | 14.3 | 164 | 6.1 | |
| Ceratopogonidae | 1 | 0.04 | ||||||||||||||||
| Stratiomyidae | 1 | 0.04 | ||||||||||||||||
| Turbellaria | 15 | 0.6 | 16 | 0.8 | 12 | 0.5 | 18 | 0.6 | ||||||||||
| Number of macroinvertebrates | 2337 | 2080 | 2223 | 2962 | 905 | 1016 | 1200 | 2674 | ||||||||||
| Density of macroinvertebrates (ind m−2) | 18,696 | 16,640 | 17,784 | 23,696 | 7240 | 8128 | 9600 | 21,392 | ||||||||||
| Taxa | Axis 1 |
|---|---|
| Potamopyrgus antipodarum | 0.8491 ** |
| Bythinella cf. austriaca | −0.9898 *** |
| Planorbidae | 0.9643 *** |
| Gammaridae | −0.9853 *** |
| Asellidae | −0.8287 * |
| Oligochaeta | 0.8230 * |
| Nemouridae | −0.8361 ** |
| Limnephilidae | −0.9882 *** |
| Haliplidae | 0.9851 *** |
| Elmidae | 0.7649 * |
| Chironomidae | 0.9938 *** |
| Turbellaria | −0.9937 *** |
| Water temperature | 0.9460 *** |
| Variable | Site | Year | One-Way Anova | ||||
|---|---|---|---|---|---|---|---|
| 2012 | 2013 | 2014 | 2015 | F Ratio | P | ||
| Shell length (mm) | springhead | 0.8–4.7 | 0.6–4.7 | 0.7–4.7 | 0.6–4.7 | ||
| 3.3 ± 0.8 | 3.2 ± 0.9 | 3.2 ± 0.9 | 3.3 ± 0.9 | 1.741 | 0.157 | ||
| springbrook | 0.7–4.9 | 0.6–4.9 | 0.6–5.1 | 0.6–5.1 | |||
| 4.2 ± 0.6 | 4.2 ± 0.4 | 4.1 ± 0.6 | 4.1 ± 0.7 | 2.315 | 0.074 | ||
| t-test | 19.757 | 25.539 | 18.848 | 24.682 | |||
| p | <0.001 | <0.001 | <0.001 | <0.001 | |||
| Fecundity (number of embryos per female) | springhead | 0–36 | 0–34 | 0–37 | 0–37 | ||
| 15.4 ± 12.5 | 14.5 ± 11.9 | 14.9 ± 12.2 | 15.2 ± 12.5 | 2.087 | 0.106 | ||
| springbrook | 0–41 | 0–42 | 0–39 | 0–42 | |||
| 19.4 ± 12.7 | 21.4 ± 12.3 | 20.3 ± 10.3 | 19.8 ± 12.1 | 1.775 | 0.156 | ||
| t-test | 2.377 | 2.585 | 2.159 | 2.025 | |||
| p | <0.01 | <0.01 | <0.01 | <0.01 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krodkiewska, M.; Cieplok, A.; Spyra, A. The Colonization of a Cold Spring Ecosystem by the Invasive Species Potamopyrgus antipodarum (Gray, 1843) (Gastropoda: Tateidae) (Southern Poland). Water 2021, 13, 3209. https://doi.org/10.3390/w13223209
Krodkiewska M, Cieplok A, Spyra A. The Colonization of a Cold Spring Ecosystem by the Invasive Species Potamopyrgus antipodarum (Gray, 1843) (Gastropoda: Tateidae) (Southern Poland). Water. 2021; 13(22):3209. https://doi.org/10.3390/w13223209
Chicago/Turabian StyleKrodkiewska, Mariola, Anna Cieplok, and Aneta Spyra. 2021. "The Colonization of a Cold Spring Ecosystem by the Invasive Species Potamopyrgus antipodarum (Gray, 1843) (Gastropoda: Tateidae) (Southern Poland)" Water 13, no. 22: 3209. https://doi.org/10.3390/w13223209