
Gross Primary Production of Rainfed and Irrigated Potato (Solanum tuberosum L.) in the Colombian Andean Region Using Eddy Covariance Technique

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description and Crop Management
2.2. Microclimate and Eddy Covariance (EC) Measurements
2.3. Biometric Measurements
2.4. Data Processing and Quality Control
2.5. Gap-Filling Methods
2.6. NEE Partitioning
2.7. Uncertainty and Statistical Analysis
2.8. Energy Balance Closure
3. Results
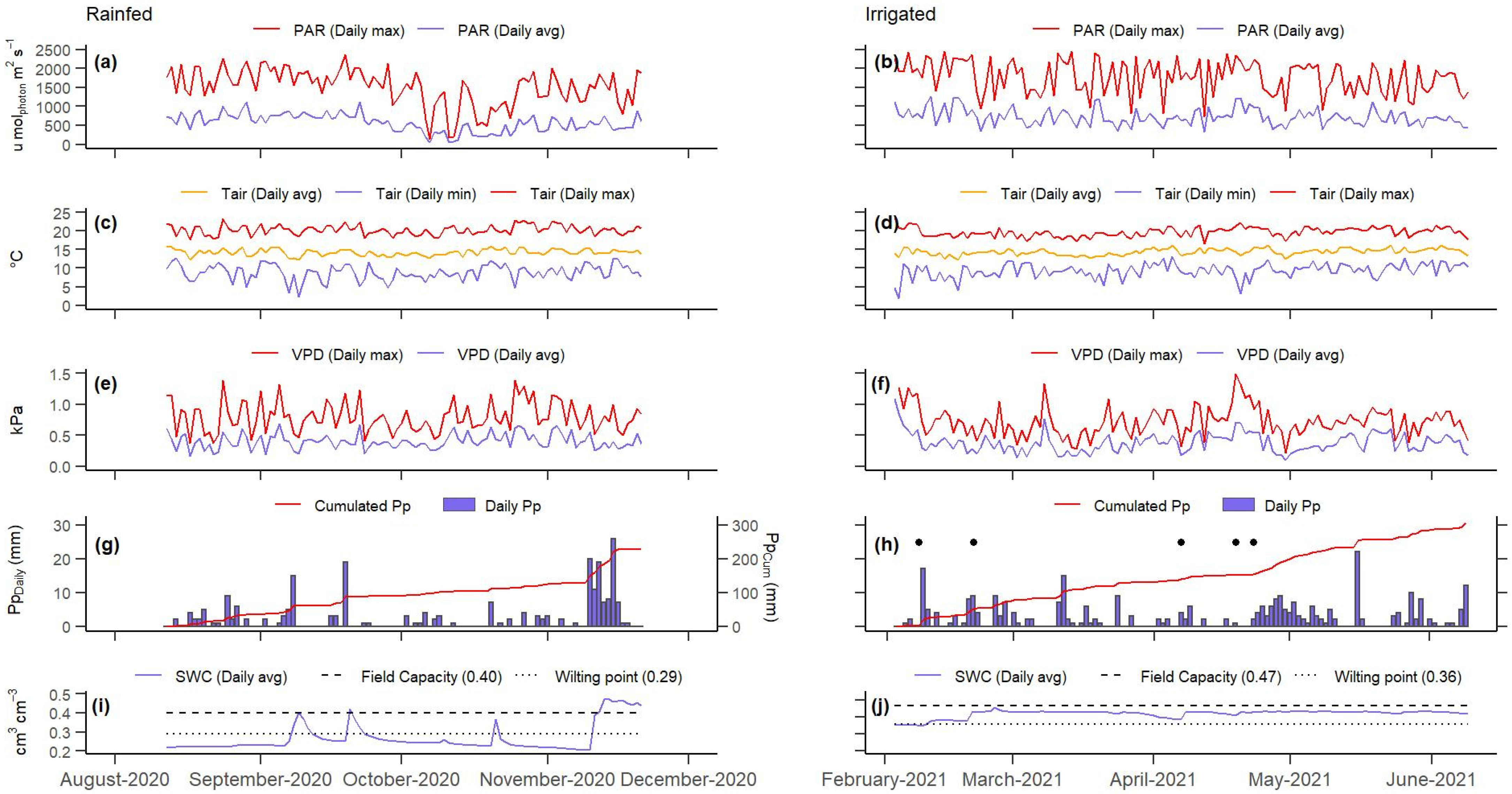
3.1. Meteorological Conditions
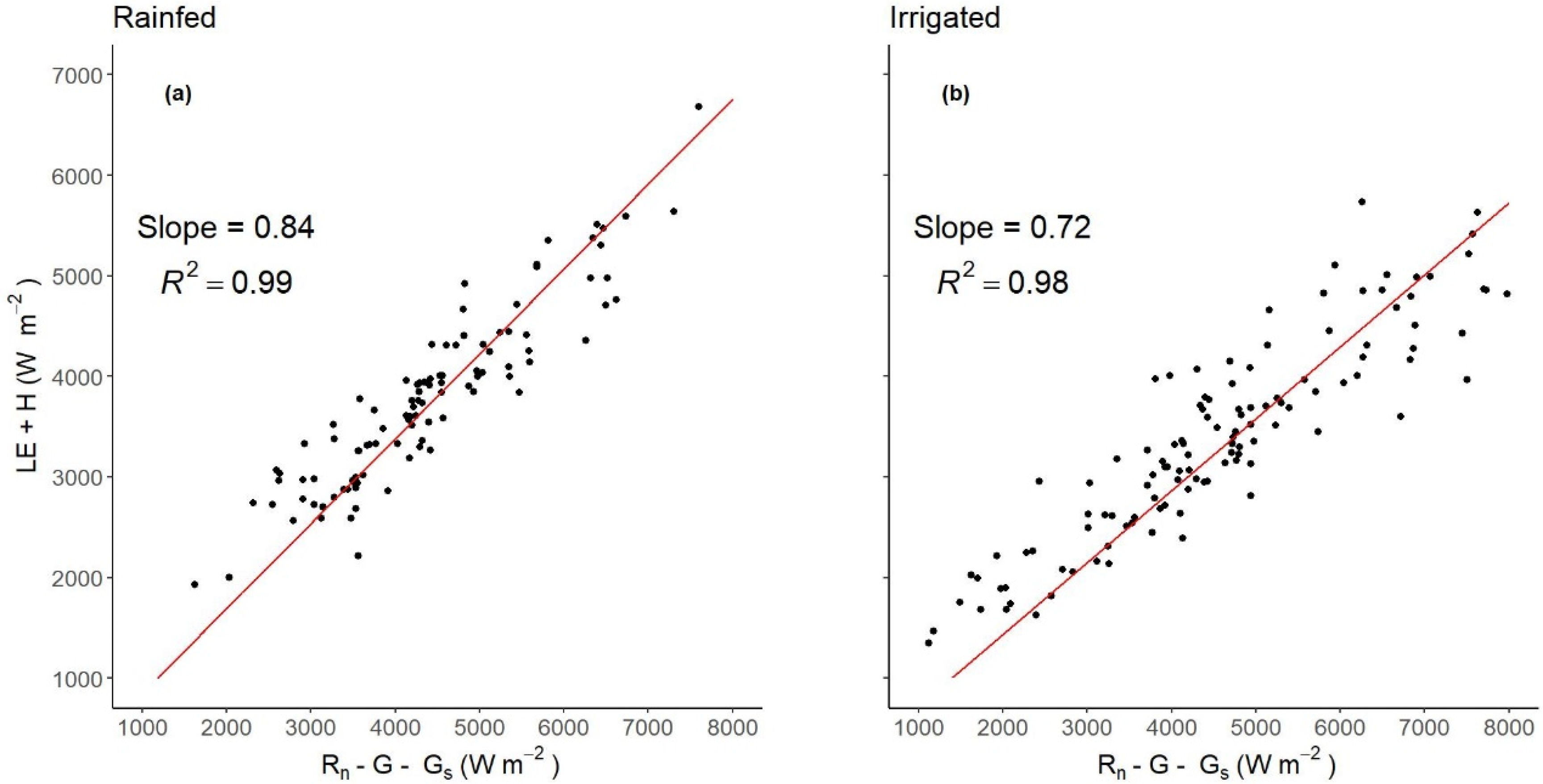
3.2. Energy Balance Closure and Uncertainty
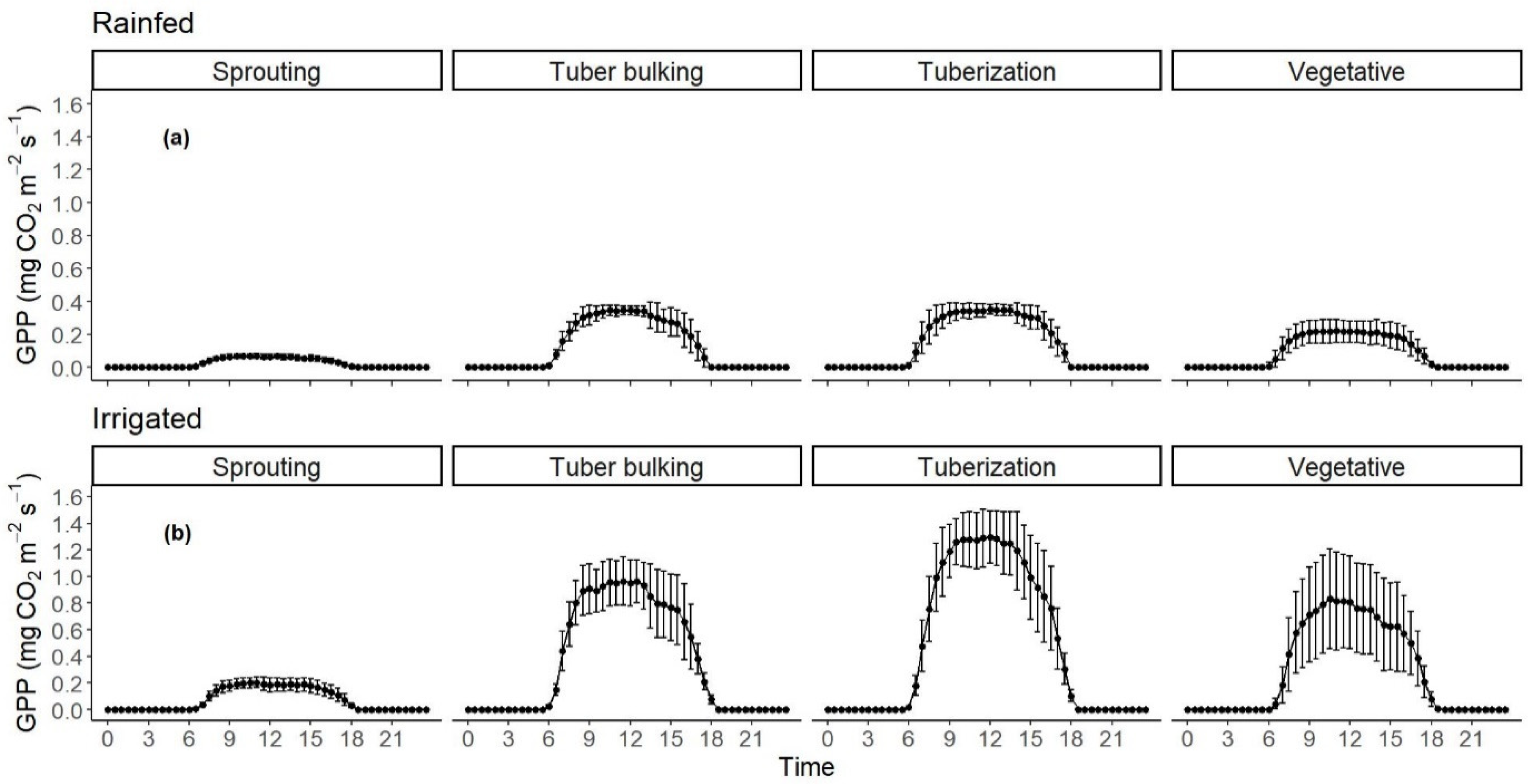
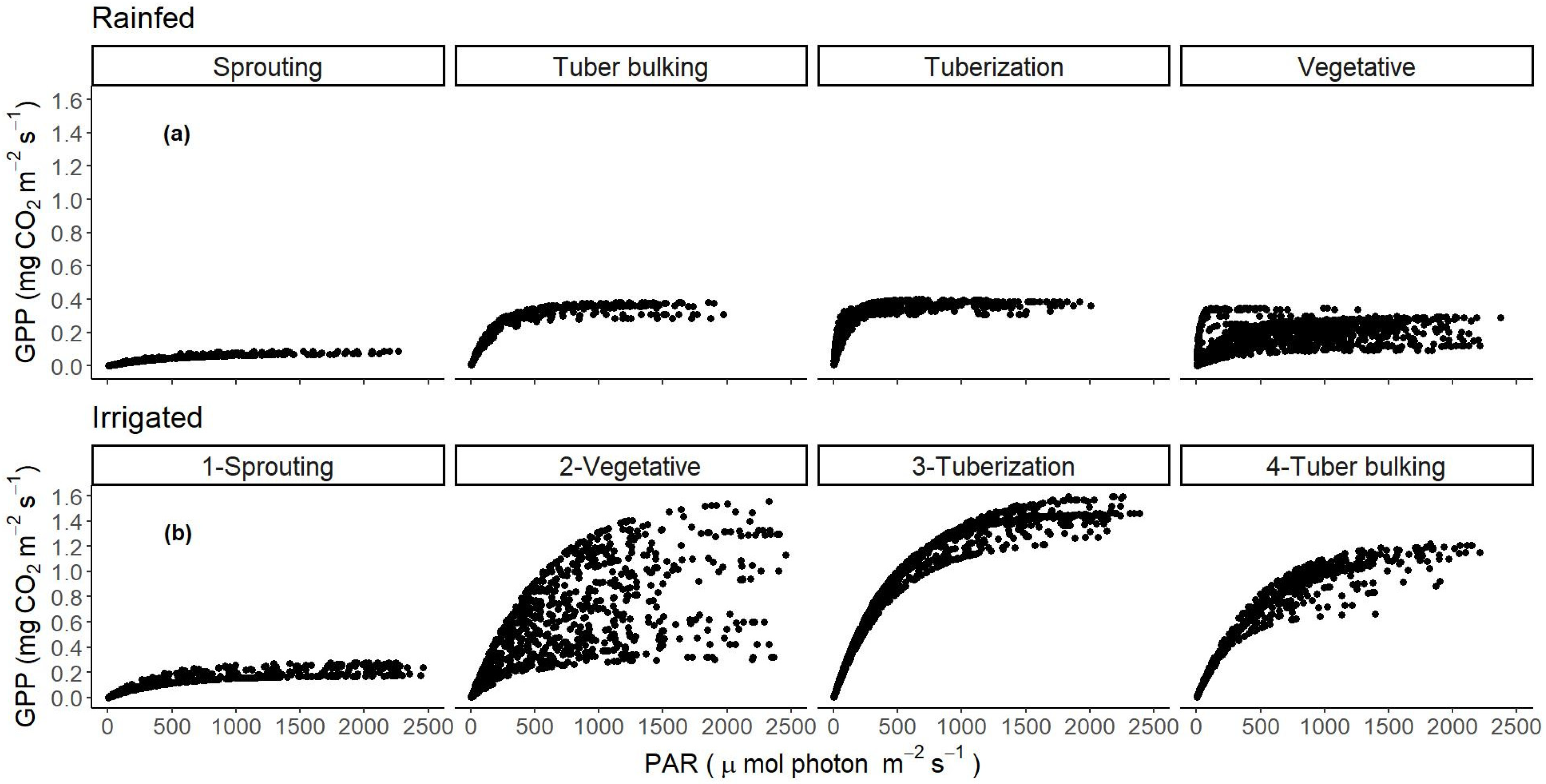
3.3. Carbon Fluxes, Daily Averages, Maximums, and Sums of NEE, GPP, and RECO in the Different Growth Stages for Non-Irrigated and Irrigated Crops
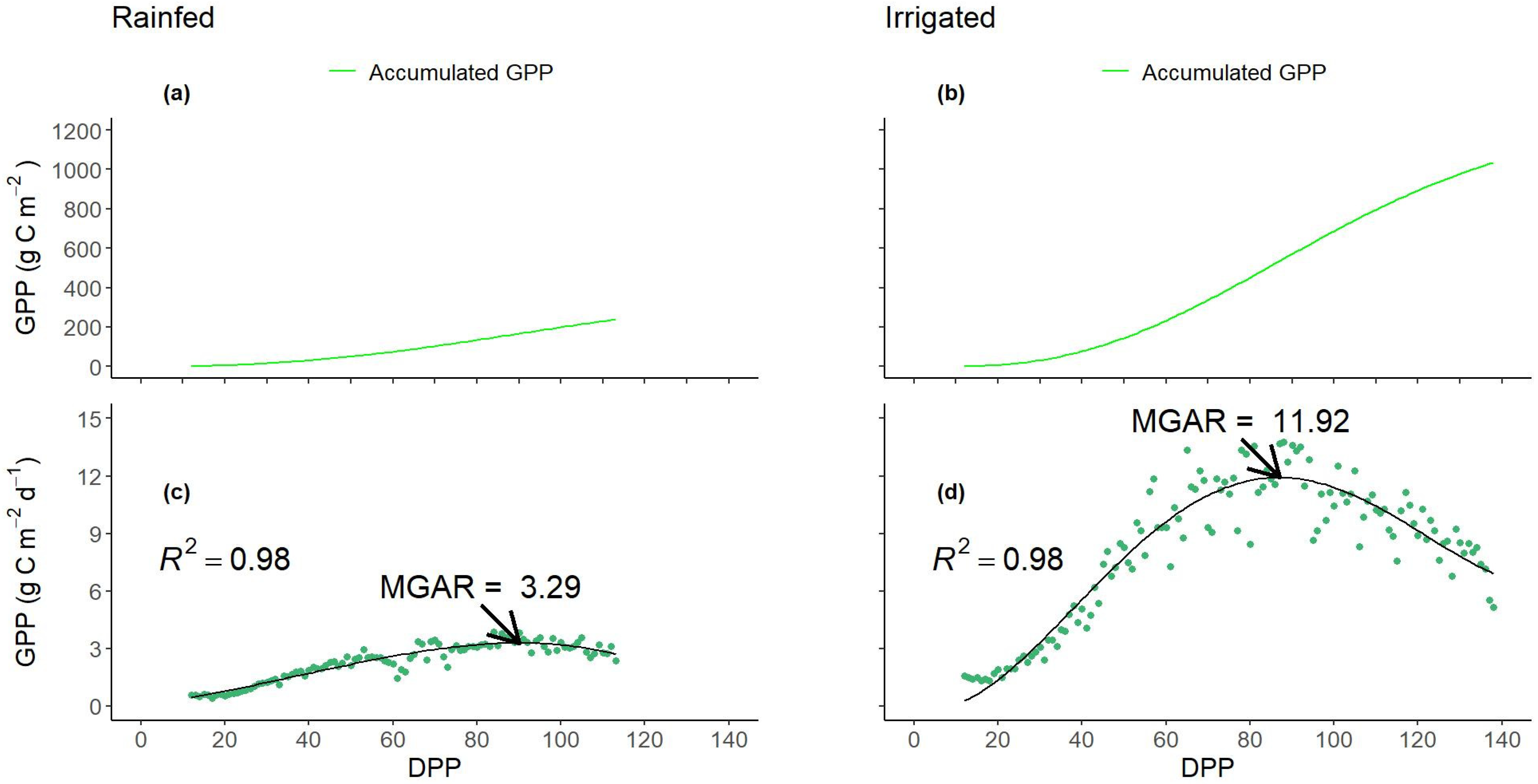
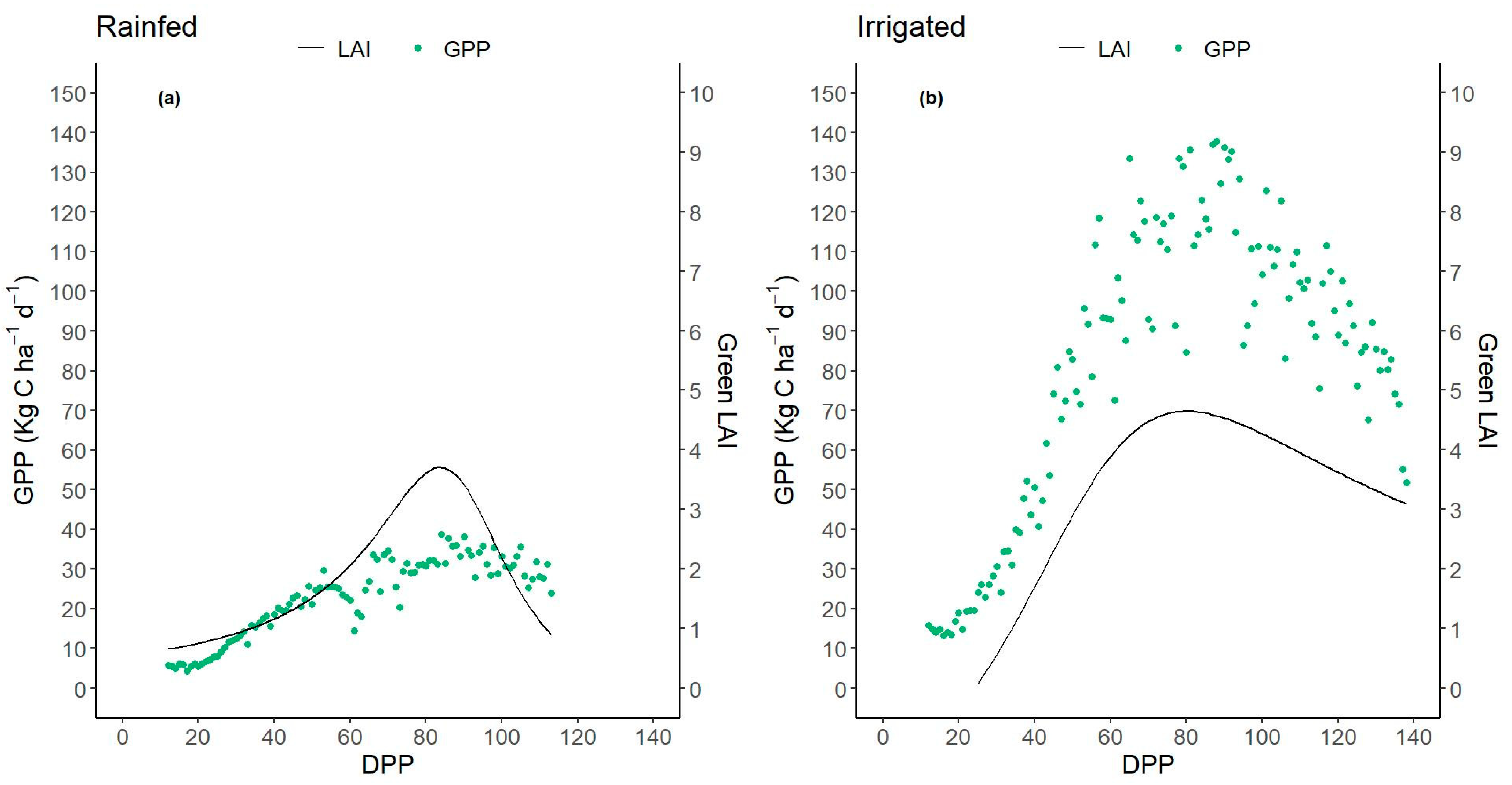
3.4. Dynamics of Daily and Accumulated Gross Primary Production—GPP
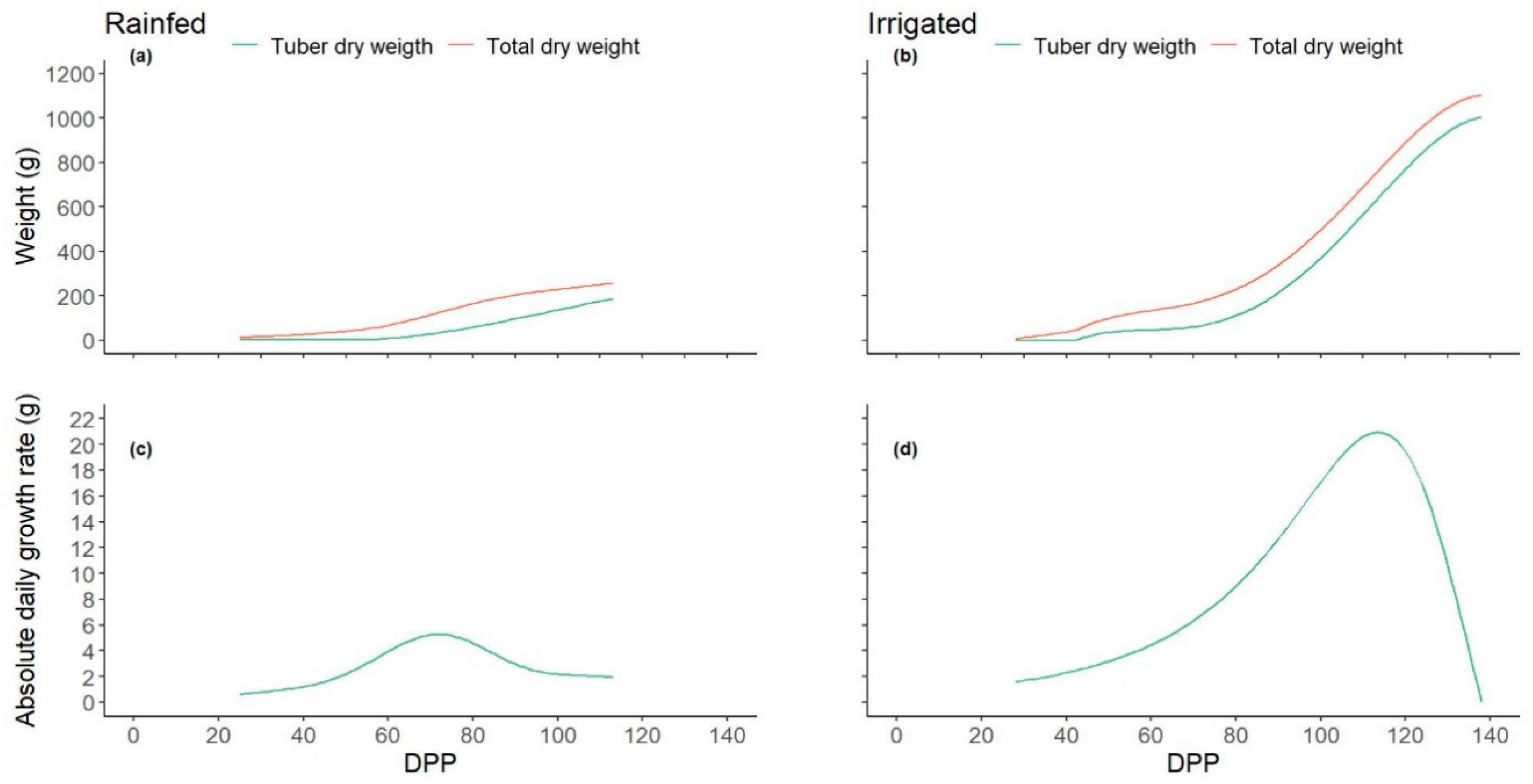
3.5. GPP and Growth Relationships
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Paillard, S.; Treyer, S.; Dorin, B. Agrimonde–Scenarios and Challenges for Feeding the World in 2050; Paillard, S., Treyer, S., Dorin, B., Eds.; Springer: Dordrecht, The Netherlands, 2014; ISBN 978-94-017-8744-4. [Google Scholar]
- FAO. The Future of Food and Agriculture Trends and Challenges; FAO—Food and Agriculture Organization of the United Nations: Roma, Italy, 2017; ISBN 978-92-5-109551-5. [Google Scholar]
- Kole, C. Genomic Designing of Climate-Smart Cereal Crops; Springer Nature Switzerland AG 2020: New Delhi, India, 2020; ISBN 978-3-319-97414-9. [Google Scholar]
- Ortiz, O.; Mares, V. The Historical, Social, and Economic Importance of the Potato Crop. Compend. Plant Genomes 2017, 1–10. [Google Scholar] [CrossRef]
- Smith, L.G.; Kirk, G.J.D.; Jones, P.J.; Williams, A.G. The greenhouse gas impacts of converting food production in England and Wales to organic methods. Nat. Commun. 2019, 10, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Martin-Gorriz, B.; Martínez-Alvarez, V.; Maestre-Valero, J.F.; Gallego-Elvira, B. Influence of the Water Source on the Carbon Footprint of Irrigated Agriculture: A Regional Study in South-Eastern Spain. Agronomy 2021, 11, 351. [Google Scholar] [CrossRef]
- Verma, S.B.; Dobermann, A.; Cassman, K.G.; Walters, D.T.; Knops, J.M.; Arkebauer, T.J.; Suyker, A.E.; Burba, G.G.; Amos, B.; Yang, H.; et al. Annual carbon dioxide exchange in irrigated and rainfed maize-based agroecosystems. Agric. For. Meteorol. 2005, 131, 77–96. [Google Scholar] [CrossRef] [Green Version]
- Litton, C.M.; Giardina, C.P. Below-ground carbon flux and partitioning: Global patterns and response to temperature. Funct. Ecol. 2008, 22, 941–954. [Google Scholar] [CrossRef]
- Rambal, S.; Lempereur, M.; Limousin, J.M. How drought severity constrains GPP and its partitioning among carbon pools in a Quercus ilex coppice? Biogeosciences Discuss. 2014, 11, 8673–8711. [Google Scholar] [CrossRef]
- Li, X.; Ramírez, D.A.; Qin, J.; Dormatey, R.; Bi, Z.; Sun, C.; Wang, H.; Bai, J. Water restriction scenarios and their effects on traits in potato with different degrees of drought tolerance. Sci. Hortic. 2019, 256. [Google Scholar] [CrossRef]
- Blom-Zandstra, G.; Verhagen, J. Potato production systems in different agro ecological regions and their relation with climate change. Wagening. Res. Rep. 2015, 614, 32. [Google Scholar]
- Cassman, K.G.; Wood, S. Chapter 26 Cultivated Systems. In Millennium Ecosystem Assessment: Global Ecosystem Assessment Report on Conditions and Trends; World Research Institute: Washington, DC, USA, 2005; ISBN 1-59726-040-1. [Google Scholar]
- Malhi, Y.; Aragão, L.E.O.C.; Metcalfe, D.B.; Paiva, R.; Quesada, C.A.; Almeida, S.; Anderson, L.; Brando, P.; Chambers, J.Q.; da Costa, A.C.L.; et al. Comprehensive assessment of carbon productivity, allocation and storage in three Amazonian forests. Glob. Chang. Biol. 2009, 15, 1255–1274. [Google Scholar] [CrossRef]
- Wagle, P.; Xiao, X.; Suyker, A.E. Estimation and analysis of gross primary production of soybean under various management practices and drought conditions. ISPRS J. Photogramm. Remote Sens. 2015, 99, 70–83. [Google Scholar] [CrossRef] [Green Version]
- Baldocchi, D.D. Assessing the eddy covariance technique for evaluating carbon dioxide exchange rates of ecosystems: Past, present and future. Glob. Chang. Biol. 2003, 9, 479–492. [Google Scholar] [CrossRef] [Green Version]
- Anthoni, P.M.; Knohl, A.; Rebmann, C.; Freibauer, A.; Mund, M.; Ziegler, W.; Kolle, O.; Schulze, E.D. Forest and agricultural land-use-dependent CO2 exchange in Thuringia, Germany. Glob. Chang. Biol. 2004, 10, 2005–2019. [Google Scholar] [CrossRef]
- Centro Internacional de la Papa (CIP) Hechos y Cifras Sobre la Papa 2017, 2. Available online: https://cgspace.cgiar.org/bitstream/handle/10568/87957/CIP-Hechos-y-cifras-sobre-la-papa-Espanol-2017.pdf?sequence=1&isAllowed=y (accessed on 29 September 2021).
- Mosquera Vásquez, T.; Del Castillo, S.; Gálvez, D.C.; Rodríguez, L.E. Breeding Differently: Participatory Selection and Scaling Up Innovations in Colombia. Potato Res. 2017, 60, 361–381. [Google Scholar] [CrossRef] [Green Version]
- U.S. Department of Agriculture-USDA. Natural Resources Conservation Services—NRCS Claves para la Taxonomía de Suelos; XII; Estado de México. 2014; ISBN 0926487221. Available online: https://www.nrcs.usda.gov/Internet/FSE_DOCUMENTS/nrcs142p2_051546.pdf (accessed on 29 September 2021).
- Foken, T.; Leuning, R.; Oncley, S.R.; Mauder, M.; Aubinet, M. Corrections and Data Quality Control. In A Practical Guide to Measurement and Data Analysis; Springer Science + Business Media B.V.: Bayreuth, Germany, 2012; pp. 85–131. ISBN 9789400723511. [Google Scholar]
- Hunt, R. BASIC GROWTH ANALYSIS; Academic Division of Unwin Hyman Ltd.: London, UK, 1990; Volume 148, ISBN 9780044453734. [Google Scholar]
- Wilczak, J.M.; Oncley, S.P.; Stage, S.A. Sonic anemometer tilt correction algorithms. Bound. Layer Meteorol. 2001, 99, 127–150. [Google Scholar] [CrossRef]
- Moncrieff, J.B.; Massheder, J.M.; De Bruin, H.; Elbers, J.; Friborg, T.; Heusinkveld, B.; Kabat, P.; Scott, S.; Soegaard, H.; Verhoef, A. A system to measure surface fluxes of momentum, sensible heat, water vapour and carbon dioxide. J. Hydrol. 1997, 188–189, 589–611. [Google Scholar] [CrossRef]
- Webb, E.K.; Pearman, G.I.; Leuning, R. Correction of flux measurements for density effects due to heat and water vapour transfer. Q. J. R. Meteorol. Soc. 1980, 106, 85–100. [Google Scholar] [CrossRef]
- Kljun, N.; Calanca, P.; Rotach, M.W.; Schmid, H.P. A simple two-dimensional parameterisation for Flux Footprint Prediction (FFP). Geosci. Model Dev. 2015, 8, 3695–3713. [Google Scholar] [CrossRef] [Green Version]
- Kormann, R.; Meixner, F.X. An analytical footprint model for non-neutral stratification. Bound.-Layer Meteorol. 2001, 99, 207–224. [Google Scholar] [CrossRef]
- Papale, D.; Reichstein, M.; Aubinet, M.; Canfora, E.; Bernhofer, C.; Kutsch, W.; Longdoz, B.; Rambal, S.; Valentini, R.; Vesala, T.; et al. Towards a standardized processing of Net Ecosystem Exchange measured with eddy covariance technique: Algorithms and uncertainty estimation. Biogeosciences 2006, 3, 571–583. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: http://www.R-project.org/2021 (accessed on 10 August 2021).
- Reichstein, M.; Falge, E.; Baldocchi, D.; Papale, D.; Aubinet, M.; Berbigier, P.; Bernhofer, C.; Buchmann, N.; Gilmanov, T.; Granier, A. On the separation of net ecosystem exchange into assimilation and ecosystem respiration : Review and improved algorithm. Glob. Chang. Biol. 2005, 11, 1424–1439. [Google Scholar] [CrossRef]
- Falge, E.; Baldocchi, D.; Olson, R.; Anthoni, P.; Aubinet, M.; Bernhofer, C.; Burba, G.; Ceulemans, R.; Clement, R.; Dolman, H.; et al. Gap filling stratergies for defansible annual sums of net ecosystem exchange. Agric. For. Meteorol. 2001, 107, 43–69. [Google Scholar] [CrossRef] [Green Version]
- Tagesson, T.; Fensholt, R.; Cropley, F.; Guiro, I.; Horion, S.; Ehammer, A.; Ardo, J. Dynamics in carbon exchange fluxes for a grazed semi-arid savanna ecosystem in West Africa. Agric. Ecosyst. Environ. 2015, 205, 15–24. [Google Scholar] [CrossRef] [Green Version]
- Richardson, A.D.; Hollinger, D.Y. A method to estimate the additional uncertainty in gap-filled NEE resulting from long gaps in the CO 2 flux record. Agric. For. Meteorol. 2007, 147, 199–208. [Google Scholar] [CrossRef]
- Richardson, A.D.; Hollinger, D.Y.; Burba, G.G.; Davis, K.J.; Flanagan, L.B.; Katul, G.G.; Munger, J.W.; Ricciuto, D.M.; Stoy, P.C.; Suyker, A.E.; et al. A multi-site analysis of random error in tower-based measurements of carbon and energy fluxes. Agric. For. Meteorol. 2006, 136, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Campbell, G.S.; Norman, J.M. An Introduction to Environmental Biophysics, 2nd ed.; Springer: Pullman, WA, USA, 1998; Volume 6, ISBN 0387949372. [Google Scholar]
- Chi, J.; Waldo, S.; Pressley, S.; O’Keeffe, P.; Huggins, D.; Stöckle, C.; Pan, W.L.; Brooks, E.; Lamb, B. Assessing carbon and water dynamics of no-till and conventional tillage cropping systems in the inland Pacific Northwest US using the eddy covariance method. Agric. For. Meteorol. 2016, 218–219, 37–49. [Google Scholar] [CrossRef] [Green Version]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. FAO Irrigation and Drainage Paper No. 56-Crop Evapotranspiration 1998, 300. Available online: http://www.climasouth.eu/sites/default/files/FAO%2056.pdf (accessed on 29 September 2021).
- Mauder, M.; Foken, T.; Cuxart, J. Surface-Energy-Balance Closure over Land: A Review; Springer: Garmisch-Partenkirchen, Germany, 2020; Volume 177, ISBN 0123456789. [Google Scholar]
- Callañaupa Gutierrez, S.; Segura Cajachagua, H.; Saavedra Huanca, M.; Flores Rojas, J.; Silva Vidal, Y.; Cuxart, J. Seasonal variability of daily evapotranspiration and energy fluxes in the Central Andes of Peru using eddy covariance techniques and empirical methods. Atmos. Res. 2021, 261, 105760. [Google Scholar] [CrossRef]
- Wilson, K.; Goldstein, A.; Falge, E.; Aubinet, M.; Baldocchi, D.; Berbigier, P.; Bernhofer, C.; Ceulemans, R.; Dolman, H.; Field, C.; et al. Energy balance closure at FLUXNET sites. Agric. For. Meteorol. 2002, 113, 223–243. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Huang, Y.; Wang, G. Detecting drought-induced GPP spatiotemporal variabilities with sun-induced chlorophyll fluorescence during the 2009/2010 droughts in China. Ecol. Indic. 2021, 121, 107092. [Google Scholar] [CrossRef]
- Fu, Z.; Ciais, P.; Bastos, A.; Stoy, P.C.; Yang, H.; Green, J.K.; Wang, B.; Yu, K.; Huang, Y.; Knohl, A.; et al. Sensitivity of gross primary productivity to climatic drivers during the summer drought of 2018 in Europe: Sensitivity of GPP to climate drivers. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Şaylan, L.; Kimura, R.; Munkhtsetseg, E.; Kamichika, M. Seasonal variation of carbon dioxide fluxes over irrigated soybean (Glycine max L.). Theor. Appl. Climatol. 2011, 105, 277–286. [Google Scholar] [CrossRef]
- Labedzki, L.; Bąk, B. Meteorological and agricultural drought indices used in drought monitoring in Poland: A review. Meteorol. Hydrol. Water Manag. 2014, 2, 13. [Google Scholar]
- Aubinet, M.; Moureaux, C.; Bodson, B.; Dufranne, D.; Heinesch, B.; Suleau, M.; Vancutsem, F.; Vilret, A. Carbon sequestration by a crop over a 4-year sugar beet/winter wheat/seed potato/winter wheat rotation cycle. Agric. For. Meteorol. 2009, 149, 407–418. [Google Scholar] [CrossRef]
- Ruidisch, M.; Nguyen, T.T.; Li, Y.L.; Geyer, R.; Tenhunen, J. Estimation of annual spatial variations in forest production and crop yields at landscape scale in temperate climate regions. Ecol. Res. 2015, 30, 279–292. [Google Scholar] [CrossRef]
- Ciais, P.; Reichstein, M.; Viovy, N.; Granier, A.; Ogée, J.; Allard, V.; Aubinet, M.; Buchmann, N.; Bernhofer, C.; Carrara, A.; et al. Europe-wide reduction in primary productivity caused by the heat and drought in 2003. Nature 2005, 437, 529–533. [Google Scholar] [CrossRef]
- Van der Molen, M.K.; Dolman, A.J.; Ciais, P.; Eglin, T.; Gobron, N.; Law, B.E.; Meir, P.; Peters, W.; Phillips, O.L.; Reichstein, M.; et al. Drought and ecosystem carbon cycling. Agric. For. Meteorol. 2011, 151, 765–773. [Google Scholar] [CrossRef]
- Stocker, B.D.; Zscheischler, J.; Keenan, T.F.; Prentice, I.C.; Seneviratne, S.I.; Peñuelas, J. Drought impacts on terrestrial primary production underestimated by satellite monitoring. Nat. Geosci. 2019, 12, 264–270. [Google Scholar] [CrossRef]
- Dufranne, D.; Moureaux, C.; Vancutsem, F.; Bodson, B.; Aubinet, M. Comparison of carbon fluxes, growth and productivity of a winter wheat crop in three contrasting growing seasons. Agric. Ecosyst. Environ. 2011, 141, 133–142. [Google Scholar] [CrossRef]
- Geruo, A.; Velicogna, I.; Kimball, J.S.; Du, J.; Kim, Y.; Colliander, A.; Njoku, E. Satellite-observed changes in vegetation sensitivities to surface soil moisture and total water storage variations since the 2011 Texas drought. Environ. Res. Lett. 2017, 12. [Google Scholar] [CrossRef] [Green Version]
- Berninger, F. Effects of drought and phenology on GPP in Pinus sylvestris: A simulation study along a geographical gradient. Funct. Ecol. 1997, 11, 33–42. [Google Scholar] [CrossRef]
- Doughty, R.; Xiao, X.; Wu, X.; Zhang, Y.; Bajgain, R.; Zhou, Y.; Qin, Y.; Zou, Z.; McCarthy, H.; Friedman, J.; et al. Responses of gross primary production of grasslands and croplands under drought, pluvial, and irrigation conditions during 2010–2016, Oklahoma, USA. Agric. Water Manag. 2018, 204, 47–59. [Google Scholar] [CrossRef]
- Xie, X.; Li, A.; Tan, J.; Jin, H.; Nan, X.; Zhang, Z.; Bian, J.; Lei, G. Assessments of gross primary productivity estimations with satellite data-driven models using eddy covariance observation sites over the northern hemisphere. Agric. For. Meteorol. 2020, 280, 107771. [Google Scholar] [CrossRef]
- Zhao, L. Improved determination of daytime net ecosystem exchange of carbon dioxide at croplands. Biogeosciences Discuss. 2012, 9, 2883–2919. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Gamon, J.A. The need for a common basis for defining light-use efficiency: Implications for productivity estimation. Remote Sens. Environ. 2015, 156, 196–201. [Google Scholar] [CrossRef] [Green Version]
- Emmel, C.; D’Odorico, P.; Revill, A.; Hörtnagl, L.; Ammann, C.; Buchmann, N.; Eugster, W. Canopy photosynthesis of six major arable crops is enhanced under diffuse light due to canopy architecture. Glob. Chang. Biol. 2020, 26, 5164–5177. [Google Scholar] [CrossRef]
- Quan, N. Greenhouse Gas Exchange above Potato and Pea Fields in the Lower Fraser Valley in British Columbia, Canada, THE UNIVERSITY OF BRITISH COLUMBIA. 2021. Available online: https://open.library.ubc.ca/soa/cIRcle/collections/ubctheses/24/items/1.0395480 (accessed on 2 September 2021).
- Kamanga, R.M.; Mbega, E.; Ndakidemi, P. Drought Tolerance Mechanisms in Plants: Physiological Responses Associated with Water Deficit Stress in Solanum lycopersicum. Adv. Crop Sci. Technol. 2018, 06, 1–8. [Google Scholar] [CrossRef]
- Varone, L.; Ribas-Carbo, M.; Cardona, C.; Gallé, A.; Medrano, H.; Gratani, L.; Flexas, J. Stomatal and non-stomatal limitations to photosynthesis in seedlings and saplings of Mediterranean species pre-conditioned and aged in nurseries: Different response to water stress. Environ. Exp. Bot. 2012, 75, 235–247. [Google Scholar] [CrossRef]
- Mafakheri, A.; Siosemardeh, A.; Bahramnejad, B.; Struik, P.C.; Sohrabi, E. Effect of drought stress on yield, proline and chlorophyll contents in three chickpea cultivars. Aust. J. Crop Sci. 2010, 4, 580–585. [Google Scholar]
- Li, W.; Zhang, S.; Shan, L. Responsibility of non-stomatal limitations for the reduction of photosynthesis-response of photosynthesis and antioxidant enzyme characteristics in alfalfa (Medicago sativa L.) seedlings to water stress and rehydration. Front. Agric. China 2007, 1, 255–264. [Google Scholar] [CrossRef]
- Beauclaire, Q.; Gourlez, L.; Motte, D.; Bernard, H.; Bernard, L. Proofs of non-stomatal limitations of potato photosynthesis during drought by using in-situ eddy covariance data 2021, 1–2. Available online: file:///C:/Users/MDPI/AppData/Local/Temp/EGU2020-5183-print.pdf (accessed on 29 September 2021).
- Wang, J.; Xiao, X.; Bajgain, R.; Starks, P.; Steiner, J.; Doughty, R.B.; Chang, Q. Estimating leaf area index and aboveground biomass of grazing pastures using Sentinel-1, Sentinel-2 and Landsat images. ISPRS J. Photogramm. Remote Sens. 2019, 154, 189–201. [Google Scholar] [CrossRef] [Green Version]
- Duursma, R.A.; Kolari, P.; Permki, M.; Pulkkinen, M.; Mkel, A.; Nikinmaa, E.; Hari, P.; Aurela, M.; Berbigier, P.; Bernhofer, C.; et al. Contributions of climate, leaf area index and leaf physiology to variation in gross primary production of six coniferous forests across Europe: A model-based analysis. Tree Physiol. 2009, 29, 621–639. [Google Scholar] [CrossRef] [Green Version]
- Ramezani, M.R.; Massah Bavani, A.R.; Jafari, M.; Binesh, A.; Peters, S. Investigating the leaf area index changes in response to climate change (case study: Kasilian catchment, Iran). SN Appl. Sci. 2020, 2, 1–11. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| a Carbon Fluxes | Sprouting | Vegetative | Tuberization | Tuber Bulking | |||||
|---|---|---|---|---|---|---|---|---|---|
| Rainfed | Irrigated | Rainfed | Irrigated | Rainfed | Irrigated | Rainfed | Irrigated | ||
| NEE | Mean | 4.39 (±0.39) | 2.92 (±0.30) | 1.93 (±0.27) | −1.96 (±0.34) | −0.03 (±0.29) | −5.55 (±0.46) | 0.35 (±0.27) | −3.02 (±0.39) |
| Max. | 4.13 (±0.37) | 1.95 (±0.30) | −0.30 (±0.30) | −7.07 (±0.50) | −0.52 (±0.30) | −8.36 (±0.50) | −0.26 (±0.28) | −4.82 (±0.44) | |
| sum | 57.04 (±5.11) | 46.7 (±4.80) | 88.8 (±12.5) | −76.6 (±13.3) | 0.72 (7.80) | −260.9 (±21.82) | 5.67 (±4.38) | −75.6 (±9.83) | |
| GPP | Mean | −0.60 (±0.06) | −1.77 (±0.01) | −2.06 (±0.03) | −6.88 (±0.15) | −3.20 (±0.08) | −11.3 (±0.28) | −3.01 (±0.07) | −8.47 (±0.19) |
| Max. | −0.80 (±0.05) | −2.61 (±0.02) | −3.46 (±0.09) | −13.4 (±0.34) | −3.87 (±0.11) | −13.8 (±0.37) | −3.57 (±0.09) | −11.16(±0.29) | |
| sum | −7.82 (±0.75) | −28.28 (±0.09) | −94.8 (±1.19) | −268 (±6.02) | −86.4 (±2.17) | −529.9 (±13.05) | −48.2 (±1.14) | −211.8 (±4.860) | |
| RECO | Mean | 4.99 (±0.34) | 4.69 (±0.31) | 3.99 (±0.25) | 4.91 (±0.19) | 3.23 (±0.21) | 5.72 (±0.19) | 3.37 (±0.20) | 5.45 (±0.20) |
| Max. | 5.26 (±0.38) | 5.06 (±0.36) | 4.97 (±0.31) | 6.29(±0.16) | 4.26 (±0.19) | 7.75(±0.16) | 3.80 (±0.18) | 7.38 (±0.18) | |
| sum | 64.9 (±4.36) | 75.04 (±4.89) | 184 (±11.3) | 191.5 (±7.33) | 87.1 (±5.63) | 268.9 (±8.77) | 53.9 (±3.24) | 136.2 (±4.96) | |
| Growth Stage | Rainfed (Days) | Irrigated (Days) |
|---|---|---|
| Vegetative | 67.95 | 89.11 |
| Tuberization | 78.73 | 117.03 |
| Tuber Bulking | 29.25 | 85.52 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez-Maldonado, F.E.; Castaño-Marín, A.M.; Góez-Vinasco, G.A.; Marin, F.R. Gross Primary Production of Rainfed and Irrigated Potato (Solanum tuberosum L.) in the Colombian Andean Region Using Eddy Covariance Technique. Water 2021, 13, 3223. https://doi.org/10.3390/w13223223
Martínez-Maldonado FE, Castaño-Marín AM, Góez-Vinasco GA, Marin FR. Gross Primary Production of Rainfed and Irrigated Potato (Solanum tuberosum L.) in the Colombian Andean Region Using Eddy Covariance Technique. Water. 2021; 13(22):3223. https://doi.org/10.3390/w13223223
Chicago/Turabian StyleMartínez-Maldonado, Fabio Ernesto, Angela María Castaño-Marín, Gerardo Antonio Góez-Vinasco, and Fabio Ricardo Marin. 2021. "Gross Primary Production of Rainfed and Irrigated Potato (Solanum tuberosum L.) in the Colombian Andean Region Using Eddy Covariance Technique" Water 13, no. 22: 3223. https://doi.org/10.3390/w13223223