
A Case Study of a Prymnesium parvum Harmful Algae Bloom in the Ohio River Drainage: Impact, Recovery and Potential for Future Invasions/Range Expansion

 ,
,
Abstract
:1. Introduction
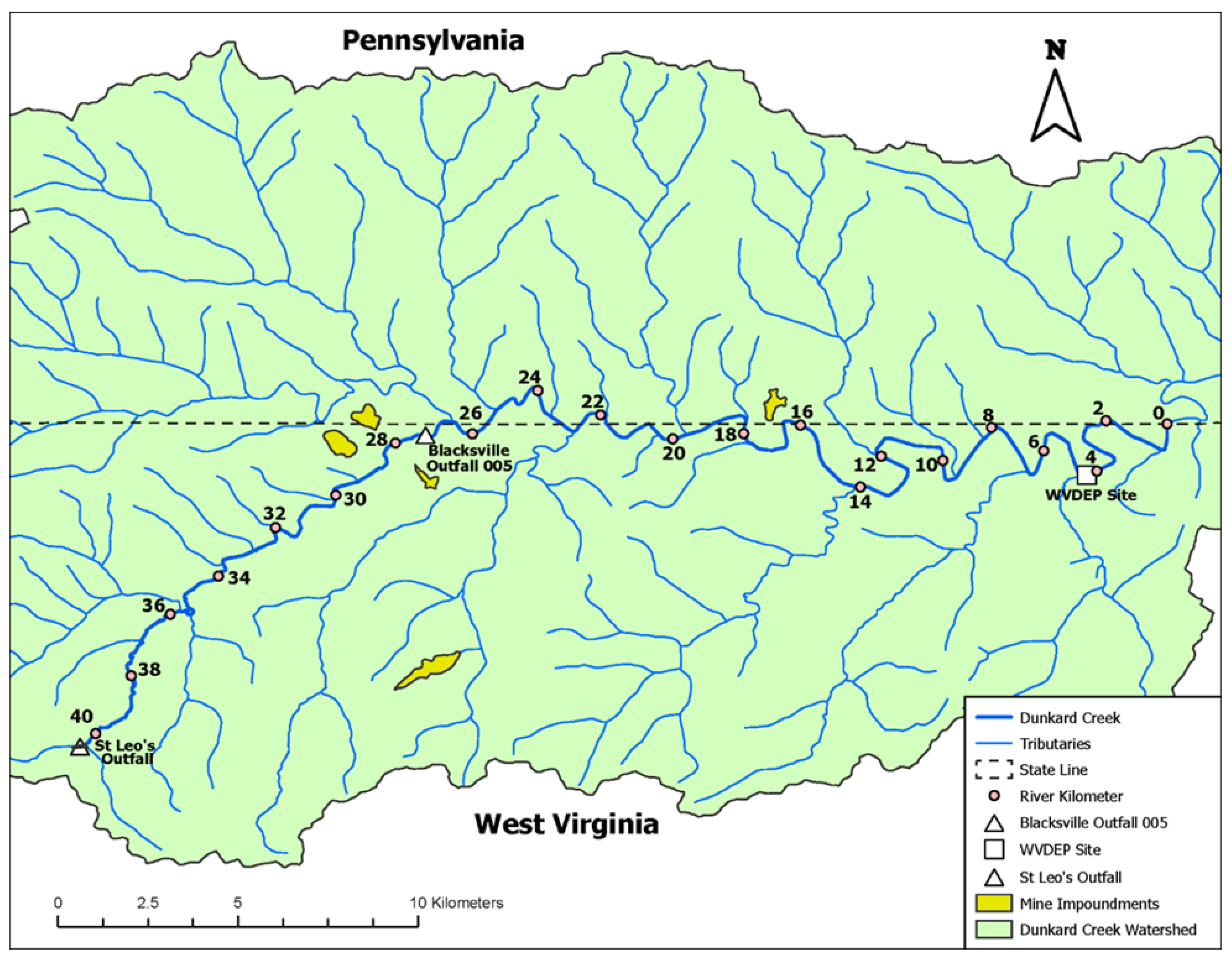
2. Dunkard Creek Case Study
2.1. Study Area
2.2. Background
2.3. Aquatic Life Sampling
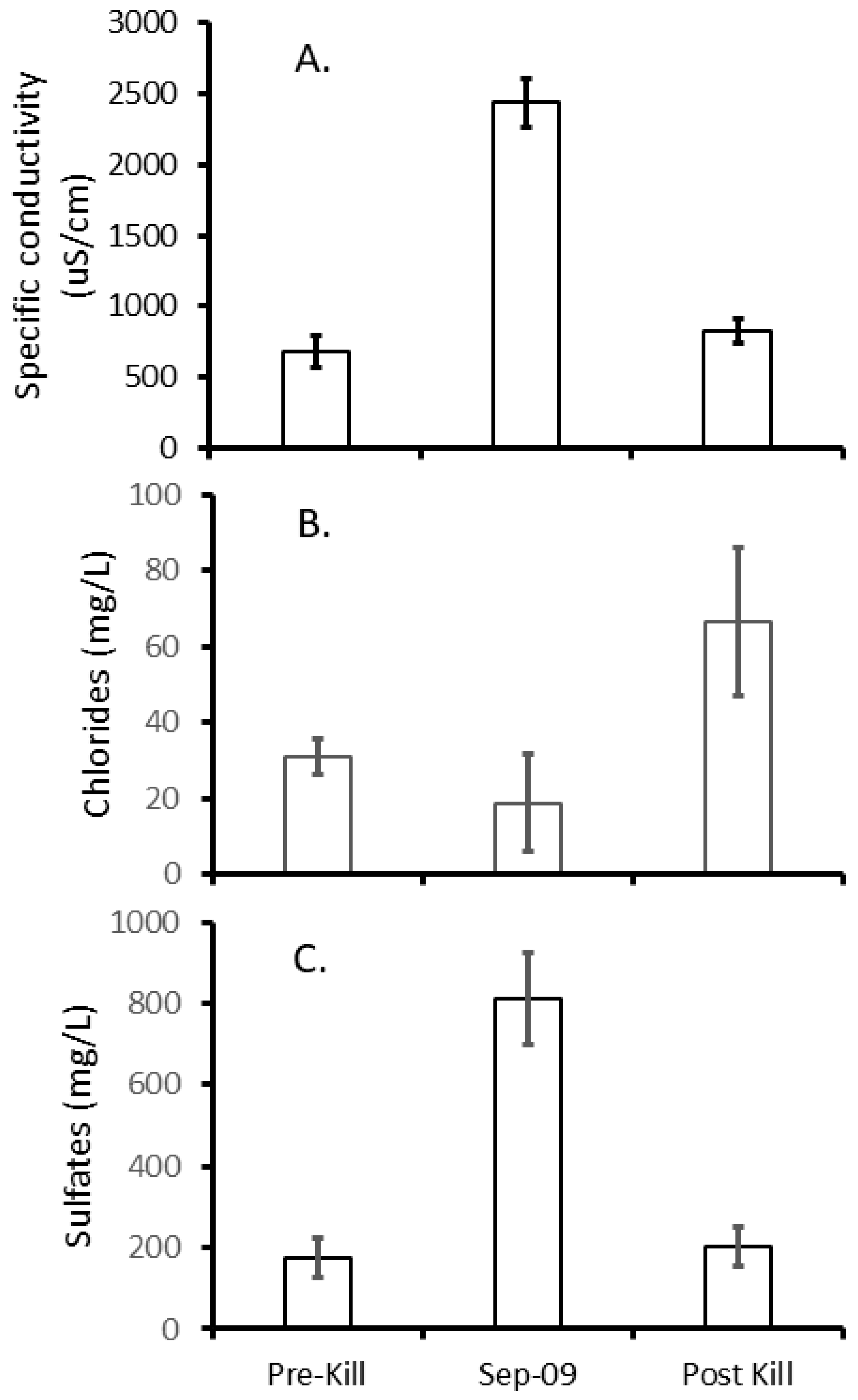
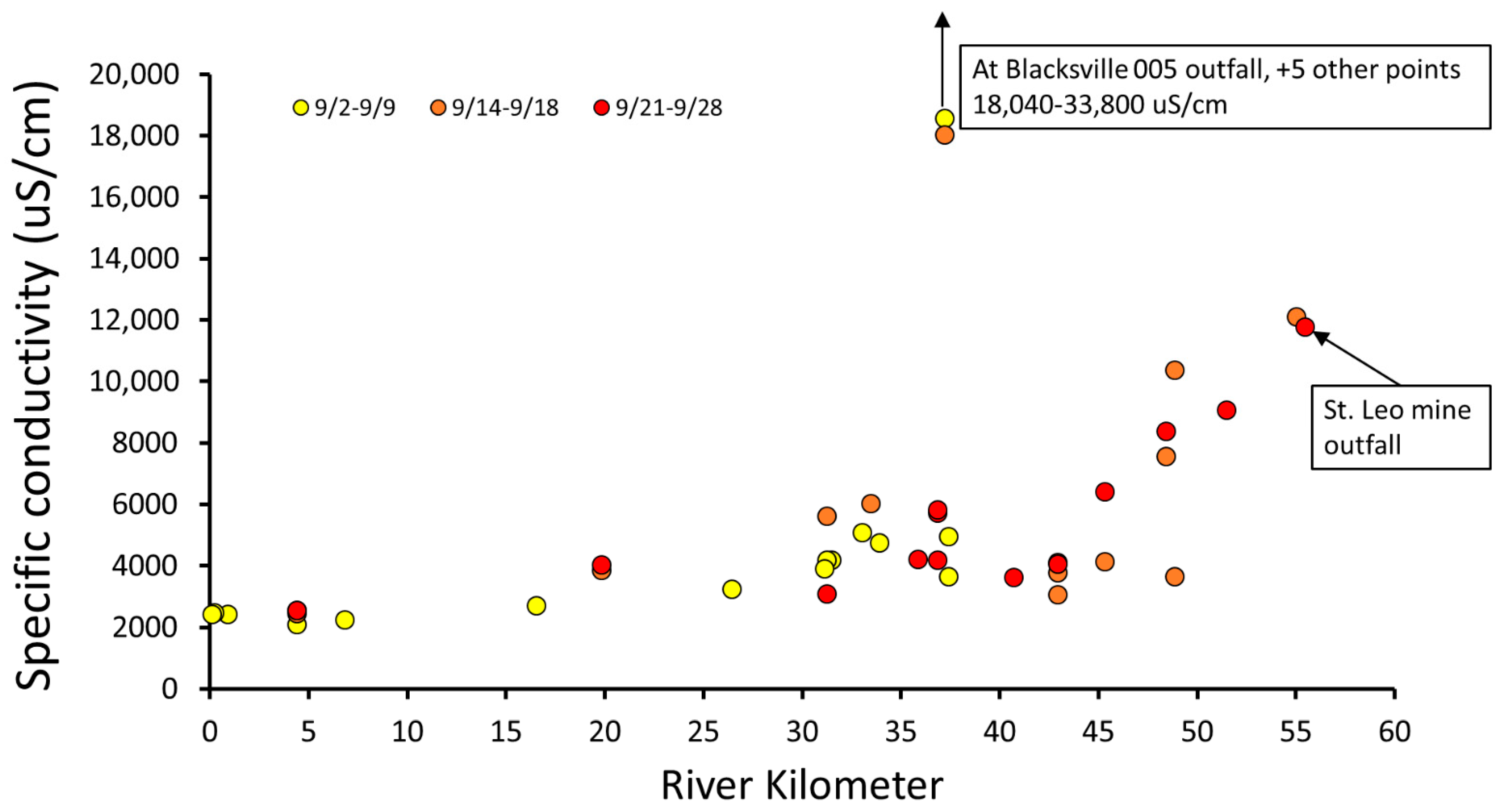
3. Results
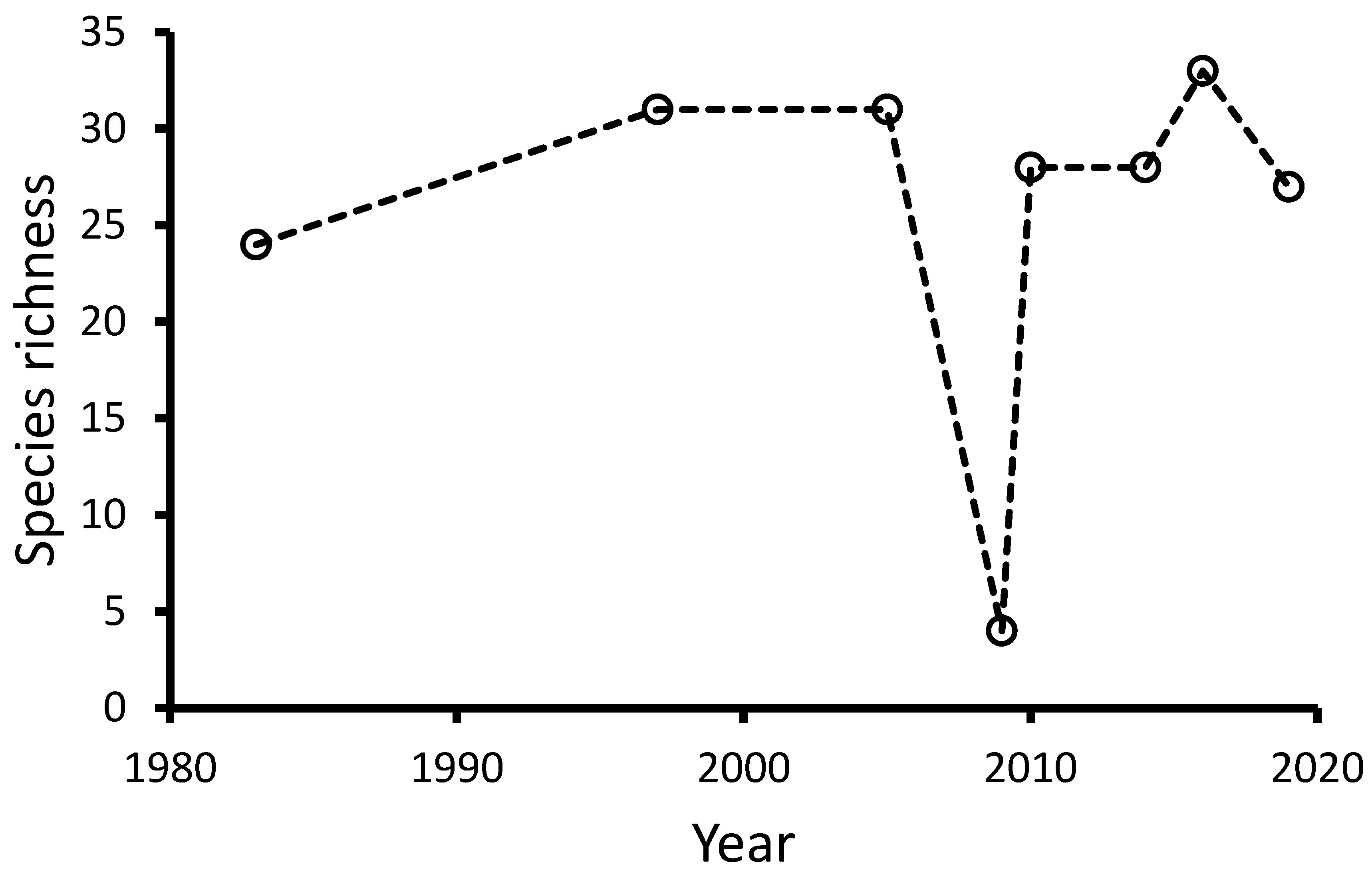
3.1. Fish Community Recovery
3.2. Mussel Community Recovery
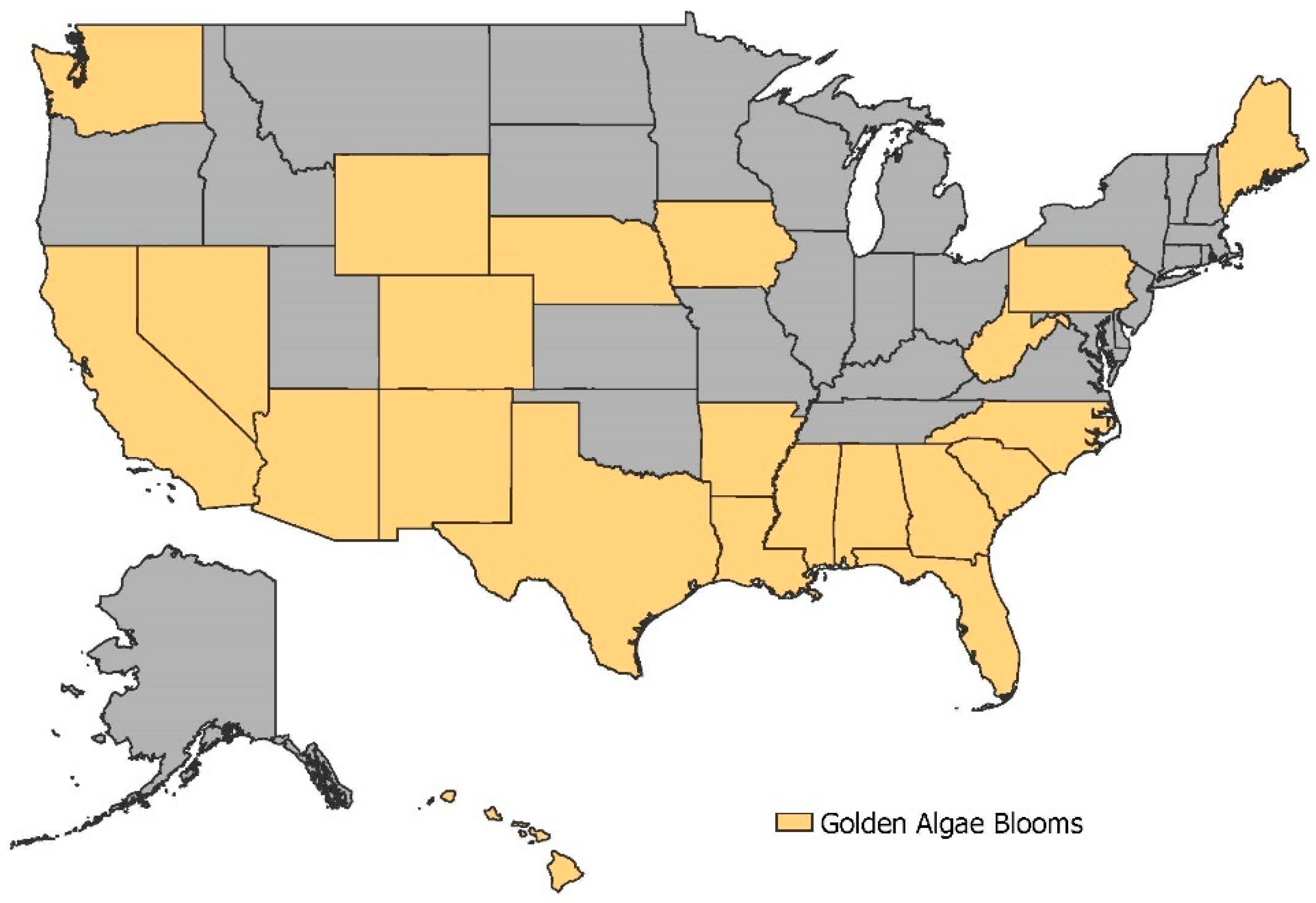
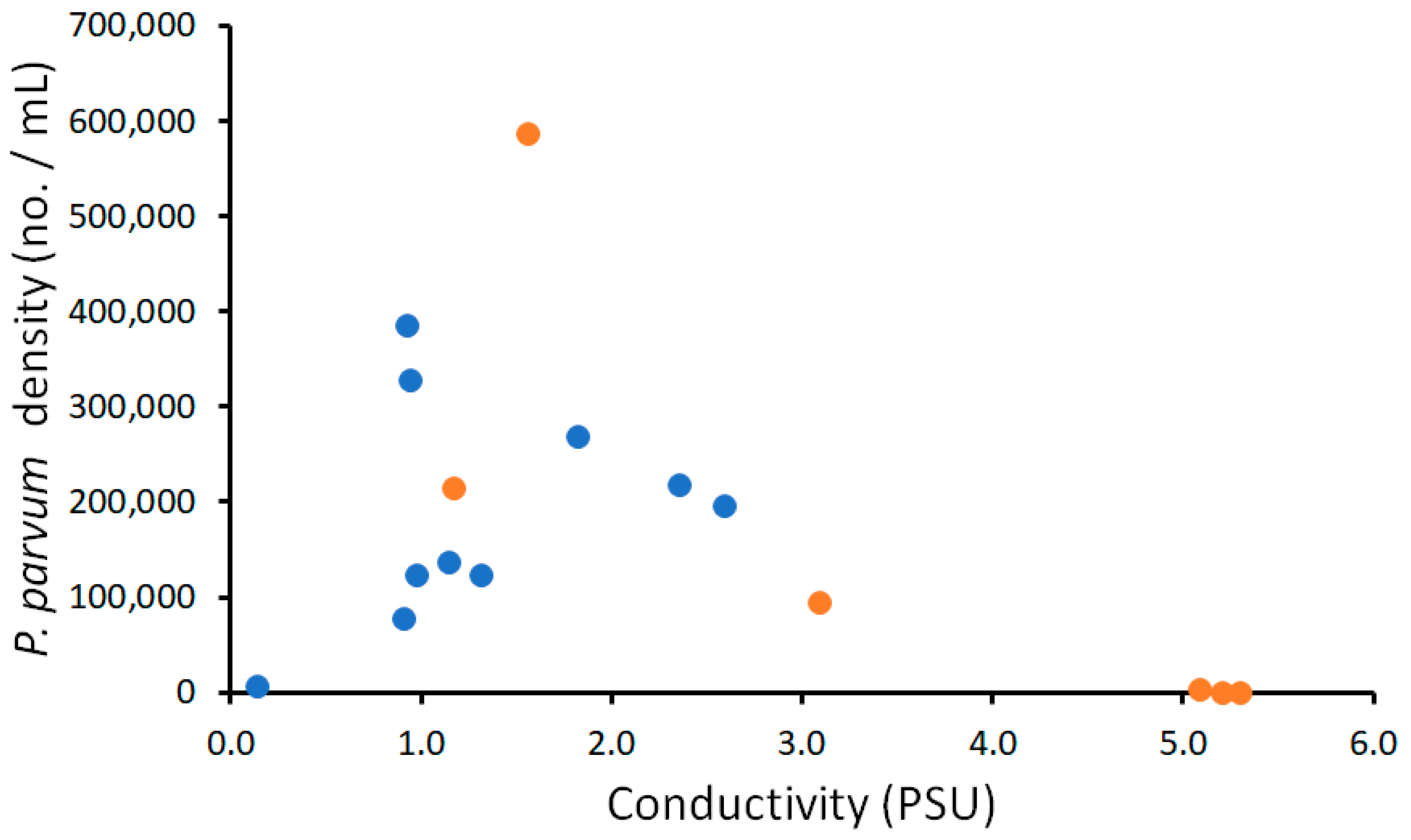
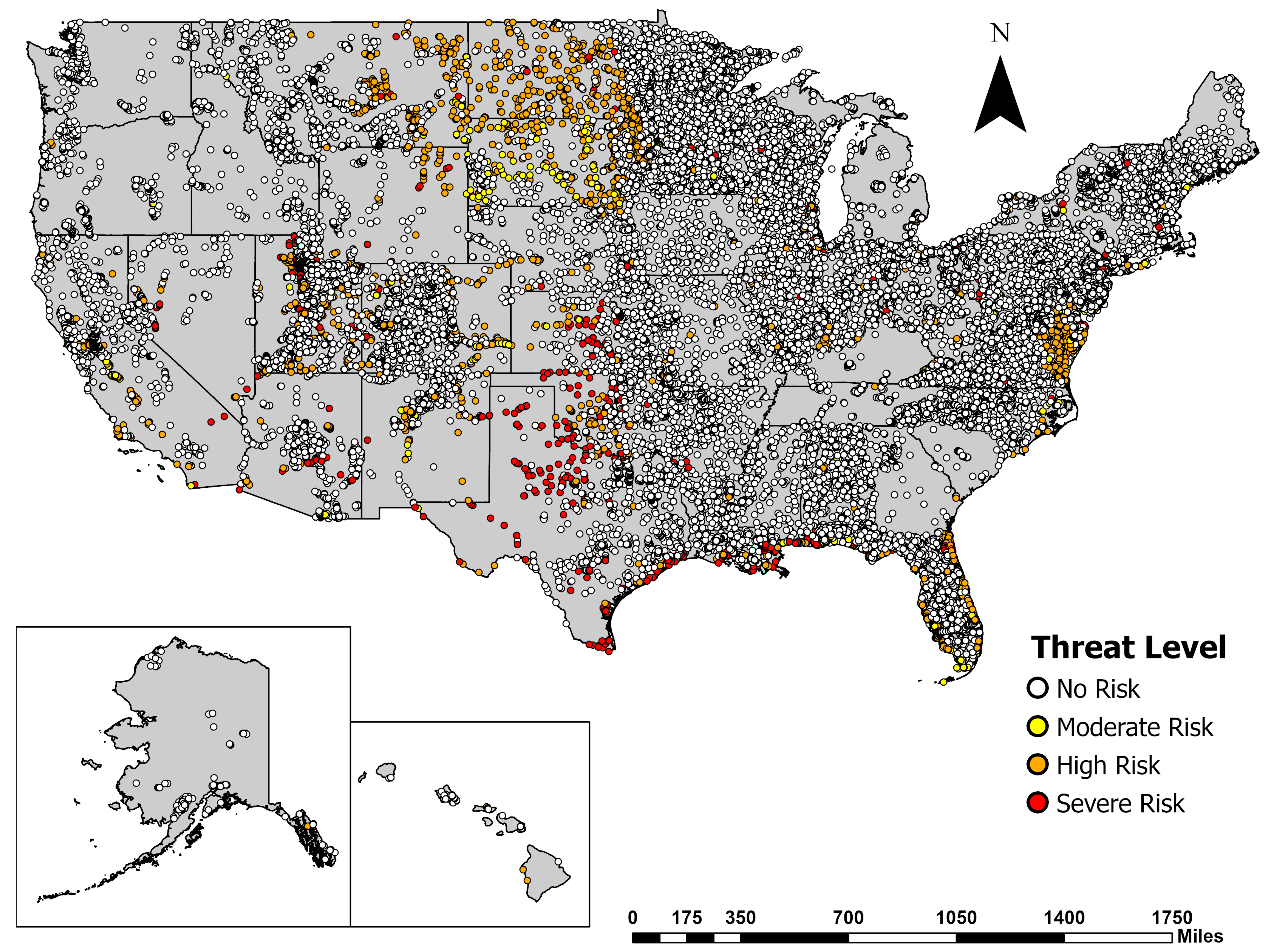
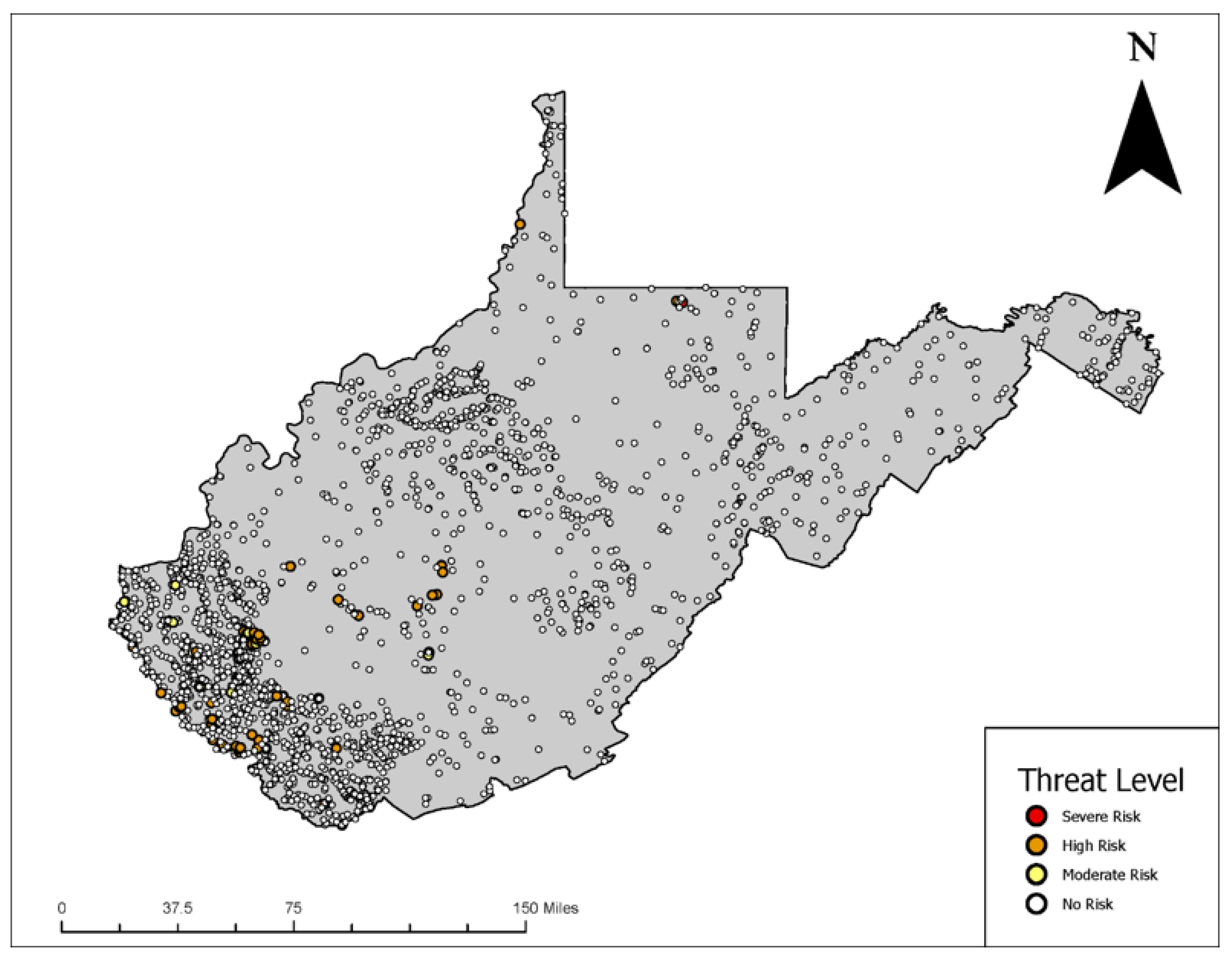
3.3. Spatial Analysis of Potential Prymnesium parvum waters
3.3.1. Water Quality Criteria and Justification
3.3.2. Spatial Mapping of Water Quality to Identify Risk Levels for the Golden Algae Prymnesium parvum
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cai, W.-J.; Hu, X.; Huang, W.-J.; Murrell, M.C.; Lehrter, J.C.; Lohrenz, S.E. Acidification of subsurface coastal waters enhanced by eutrophication. Nat. Geosci. 2011, 4, 766–770. [Google Scholar] [CrossRef]
- Lara-Martin, P.A.; Chiaia-Hernandez, A.C.; Biel-Maeso, M.; Baena-Nogueras, R.M.; Hollender, J. Tracing Urban Wastewater Contaminants into the Atlantic Ocean by Non-target Screening. Environ. Sci. Technol. 2020, 54, 3996–4005. [Google Scholar] [CrossRef]
- Rabouille, C.; Conley, D.J.; Dai, M.H.; Cai, W.-J.; Chen, C.T.A.; Lansard, B. Comparison of hypoxia among four river-dominated ocean margins: The Changjiang (Yangtze), Mississippi, Pearl, and Rhône rivers. Contin. Shelf Res. 2008, 28, 1527–1537. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Gallego, L.; Achkar, M.; Defeo, O.; Vidal, L.; Meerhoff, E.; Conde, D. Effects of land use changes on eutrophication indicators in five coastal lagoons of the Southwestern Atlantic Ocean. Est. Coast. Shelf Sci. 2017, 11, 116–126. [Google Scholar] [CrossRef]
- ASA (American Sportfishing Association). Sportfishing in America—A Reliable Economic Force; ASA: Alexandria, VA, USA, 2021; p. 18. Available online: https://asafishing.org/wp-content/uploads/2021/03/Sportfishing-in-America-economics-report-v3-16-21.pdf (accessed on 1 November 2021).
- NOAA (National Oceanic and Atmospheric Administration). Sustainable Fisheries—NOAA Fisheries Economics of the United States, 2021. Available online: https://www.fisheries.noaa.gov/national/sustainable-fisheries/fisheries-economics-united-states; https://media.fisheries.noaa.gov/2021-05/FUS-2019-infographics-webready-FINAL.pdf?null%09; (accessed on 1 November 2021).
- Israel, N.M.D.; VanLandeghem, M.W.; Denny, S.; Ingle, J.; Patino, R.A. Golden algae presence and abundance inversely related to salinity in a high-salinity river ecosystem, Pecos River, USA. Harm. Algae 2014, 39, 81–91. [Google Scholar] [CrossRef]
- Roelke, D.L.; Barkoh, A.; Brooks, B.W.; Grover, J.P.; Hambright, K.D.; LaCalair II, J.W.; Moeller, P.D.R.; Patino, R. A chronicle of a killer algae in the west: Ecology, assessment, and management of Prymnesium parvum blooms. Hydrobiologia 2016, 764, 29–50. [Google Scholar] [CrossRef]
- VanLandeghem, M.W.; Farooqi, M.; Southard, G.M.; Patino, R. Spatiotemporal associations of reservoir nutrient characteristics and the invasive, harmful algae Prymnesium parvum in West Texas. J. Am. Water Res. Assoc. 2015, 51, 487–501. [Google Scholar] [CrossRef]
- Guo, M.X.; Harrison, P.J.; Taylor, F.J.R. Fish kills related to Prymnesium parvum N Carter (Haptophyta) in the People’s Republic of China. J. Appl. Phyc. 1996, 8, 111–117. [Google Scholar] [CrossRef]
- Holdway, P.A.; Watson, R.A.; Moss, B. Aspects of the ecology of Prymnesium parvum (Haptophyta) and water chemistry in the Norfolk Broads, England. FW Biol. 1978, 8, 295–311. [Google Scholar] [CrossRef]
- VanLandeghem, M.W.; Farooqi, M.; Farquhar, B.; Patino, R. Impacts of Golden Alga Prymnesium parvum on Fish Populations in Reservoirs of the Upper Colorado River and Brazos River Basins, Texas. Trans. Am. Fish. Soc. 2013, 142, 581–595. [Google Scholar] [CrossRef]
- Southard, G.M.; Fries, L.T.; Barkoh, A. Prymnesium parvum: The Texas experience. J. Am. Water Res. Assoc. 2010, 46, 14–23. [Google Scholar] [CrossRef]
- Johnsen, T.M.; Eickrem, W.; Olseng, C.D.; Tollefsen, K.E.; Bjrerknes, V. Prymnesium parvum: The Norwegian Experience. J. Am. Water Res. Assoc. 2010, 46, 6–13. [Google Scholar] [CrossRef]
- Granéli, E.; Edvardsen, B.; Roelke, D.L.; Hagström, J.A. The ecophysiology and bloom dynamics of Prymnesium spp. Harm. Algae 2012, 14, 260–270. [Google Scholar] [CrossRef]
- Henrikson, J.C.; Gharfeh, M.S.; Easton, A.C.; Easton, J.D.; Glenn, K.L.; Shadfan, M.; Mooberry, S.L.; Hambright, K.D.; Cichewicz, R.H. Reassessing the ichthyotoxin profile of cultured Prymnesium parvum (golden algae) and comparing it to samples collected from recent freshwater bloom and fish kill events in North America. Toxicon 2010, 55, 1396–1404. [Google Scholar] [CrossRef] [PubMed]
- Renner, R. Salt-loving algae wipe out fish in Appalachian stream. Environ. Sci. Technol. 2009, 43, 9046–9047. [Google Scholar] [CrossRef] [Green Version]
- Hambright, K.D.; Easton, J.D.; Zamor, R.M.; Beyer, J.; Easton, A.C.; Allison, B. Regulation of growth and toxicity of a mixotrophic microbe: Implications for understanding range expansion in Prymnesium parvum. FW Sci. 2014, 33, 745–754. [Google Scholar] [CrossRef]
- Larsen, A.; Bryant, S. Growth Rate and Toxicity of Prymnesium parvum and Prymnesium patelliferum (Haptophyta) in Response to Changes in Salinity, Light, and Temperature. Sarsia 1998, 83, 409–418. [Google Scholar] [CrossRef]
- Hambright, K.D.; Zamor, R.M.; Easton, J.D.; Glenn, K.L.; Remmel, E.J.; Easton, A.C. Temporal and spatial variability of an invasive protist in a North American subtropical reservoir. Harm. Algae 2010, 9, 568–577. [Google Scholar] [CrossRef]
- Baker, J.W.; Grover, J.P.; Brooks, B.W.; Ureña-Boeck, F.; Roelke, D.L.; Errera, R.; Kiesling, R.L. Growth and toxicity of Prymnesium parvum (Haptophyta) as a function of salinity, light, and temperature. J. Phycol. 2007, 43, 219–227. [Google Scholar] [CrossRef]
- Patino, R.; Dawson, D.; VanLandeghem, M.M. Retrospective Analysis of Associations between Water Quality and Toxic Blooms of Golden Alga (Prymnesium parvum) in Texas Reservoirs: Implications for Understanding Dispersal Mechanisms and Impacts of Climate Change. Harm. Algae 2014, 33, 1–11. [Google Scholar] [CrossRef]
- Roelke, D.L.; Grover, J.P.; Brooks, B.W.; Glass, J.; Buzan, D.; Southard, G.M.; Fries, L.; Gable, G.M.; Schwierzke-Wade, L.; Byrd, M.; et al. A decade of fish-killing Prymnesium parvum blooms in Texas: Roles of inflow and salinity. J. Plankt. Res. 2011, 33, 243–253. [Google Scholar] [CrossRef] [Green Version]
- Genitsaris, S.; Kormas, K.A.; Moustaja-Gouni, M. Microscopic eukaryotes living in a dying lake (Lake Koronia, Greece). FEMS Microb. Ecol. 2009, 69, 75–83. [Google Scholar] [CrossRef] [Green Version]
- Granéli, E.; Salomon, P.S. Factors influencing Allelopathy and Toxocity in Prymnesium parvum. J. Am. Water Res. Assoc. 2010, 46, 108–120. [Google Scholar] [CrossRef]
- Prosser, K.N.; Valenti, T.W., Jr.; Hayden, N.J.; Neisch, M.T.; Hewitt, N.C.; Umphres, G.D.; Gable, G.M.; Grover, J.O.; Roelke, D.L.; Brooks, B.W. Low pH preempts bloom development of a toxic haptophyte. Harm. Algae 2012, 20, 156–164. [Google Scholar] [CrossRef]
- Sager, D.R.; Barkoh, A.; Buzan, D.L.; Fries, L.T.; Glass, J.A.; Kurten, G.L.; Ralph, J.J.; Singhurst, E.J.; Southard, G.M.; Swanson, E. Toxic Prymnesium parvum: A potential threat to U.S. reservoirs. Am. Fish. Soc. Symp. 2008, 62, 261–273. [Google Scholar]
- James, T.L.; De La Cruz, A. Prymnesium parvum Carter (Chrysophyceae) as a suspect of mass mortalities of fish and shellfish communities in western Texas. TX J. Sci. 1989, 41, 429–430. [Google Scholar]
- Brooks, B.W.; Grover, J.P.; Roelke, D.L. Prymnesium parvum, an emerging threat to inland waters. Environ. Tox. Chem. 2011, 30, 1955–1964. [Google Scholar] [CrossRef] [PubMed]
- Griffith, M.B. Natural variation and current reference for specific conductivity and major ions in wadeable streams of the conterminous USA. Freshw. Sci. 2014, 33, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Lutz-Carrillo, D.J.; Southard, G.M.; Fries, L.T. Global genetic relationships among isolates of golden alga (Prymnesium parvum). J. Am. Water Res. Assoc. 2010, 46, 24–32. [Google Scholar] [CrossRef]
- Hallegraeff, G.; Gollasch, S. Anthropogenic introductions of microalgae. In Ecology of Harmful Algae. Ecological Studies (Analysis and Synthesis); Granéli, E., Turner, J.T., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; Volume 189. [Google Scholar] [CrossRef]
- Kristiansen, J. Dispersal of freshwater algae—A review. Hydrobiologia 1996, 336, 151–157. [Google Scholar] [CrossRef]
- United States Environmental Protection Agency (USEPA). A Field-Based Aquatic Life Benchmark for Conductivity in Central Appalachian Streams; National Center for Environmental Assessment, Office of Research and Development: Cincinnati, OH, USA, 2011; EPA/600/R-10/023F. Available online: https://cfpub.epa.gov/ncea/risk/recordisplay.cfm?deid=220171 (accessed on 1 November 2021).
- Reynolds, L. Update on Dunkard Creek—November 23, 2009. USEPA Region 3, Wheeling, WV. 2009; p. 17. Available online: https://archive.epa.gov/region03/archive/web/pdf/dunkard.pdf (accessed on 1 November 2021).
- Rodgers, J.H., Jr.; Johnson, B.M.; Bishop, W.M. Comparison of three algaecides for controlling the density of Prymnesium parvum. J. Am. Water Res. Assoc. 2010, 46, 153–160. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1111/j.1752-1688.2009.00399.x (accessed on 1 November 2021). [CrossRef]
- Urban, C. Dunkard Creek Aquatic Life Kill Re-Assessment; Pennsylvania Fish & Boat Commission, Division of Environmental Services, Natural Diversity Section: Corry, PA, USA, 2016; p. 41. [Google Scholar]
- Holton, G.H.; Sullivan, C.R., Jr. West Virginia’s Electrical Fish-Collecting Methods. Prog. Fish-Cult. 1954, 16, 10–18. [Google Scholar] [CrossRef]
- Clayton, J.L.; Everhart, M.E. Dunkard Creek Mussel Restoration; West Virginia Division of Natural Resources, Wildlife Resources Section, Wildlife Diversity Unit: Elkins, WV, USA, 2016; p. 8. [Google Scholar]
- Smith, D.R.; Villella, R.F.; Laramie, D.P. Survey protocol for assessment of endangered freshwater mussels in the Allegheny River, Pennsylvania. J. N. Am. Benth. Soc. 2001, 20, 118–132. [Google Scholar] [CrossRef]
- Kalkhoff, S.J.; Hubbard, L.E.; Tomer, M.D.; James, D.E. Effect of variable annual precipitation and nutrient input on nitrogen and phosphorus transport from two Midwestern agricultural watersheds. Sci. Total Environ. 2016, 559, 53–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minaudo, C.; Meybeck, M.; Moatar, F.; Grassama, N.; Curie, F. Eutrophication mitigation in rivers: 30 years of trends in spatial and seasonal patterns of biochemistry of the Loire River (1980–2012). Biogeosciences 2015, 12, 2549–2563. [Google Scholar] [CrossRef] [Green Version]
- Romeo, E.; Garnier, J.; Lassaletta, L.; Billen, G.; Le Gendre, R.; Riou, P.; Cugier, P. Large-scale patterns of river inputs in southwestern Europe: Seasonal and interannual variations and potential eutrophication effects at the coastal zone. Biogeochemistry 2012, 113, 481–505. [Google Scholar] [CrossRef] [Green Version]
- Granéli, E.; Johansson, N. Effects of the toxic haptophyte Prymnesium parvum on the survival and feeding of a ciliate: The influence of different nutrient conditions. Mar. Ecol. Prog. Ser. 2003, 254, 49–56. [Google Scholar] [CrossRef]
- McLaughlin, J.J.A. Euryhaline chrysomonads: Nutrition and toxigenesis in Prymnesium parvum, with notes on Isochrysis galbana and Monochrysis lutheri. J. Protozool. 1958, 5, 75–81. [Google Scholar] [CrossRef]
- Gordon, N.; Colorni, A. Prymnesium parvum, an Ichthyotoxic Alga in an Ornamental Fish Farm in Southern Israel. Isra. J. Aquacul. 2008, 60, 5–8. [Google Scholar]
- Lindholm, T.; Virtanen, T. A bloom of Prymnesium parvum Carter in a small coastal inlet in Dragsfjard, Southwest Finland. Environ. Tox. Wat. Qual. 1992, 7, 165–170. [Google Scholar] [CrossRef]
- Vasas, G.; Marta, M.; Borics, G.; Gonda, S.; Mathe, C.; Jambrok, K.; Nagy, Z.L. Occurrence of toxic Prymnesium parvum blooms with high protease activity is related to fish mortality in Hungarian ponds. Harm. Algae 2012, 17, 102–110. [Google Scholar] [CrossRef]
- Farrow, G.A. Note on the Association of Prymnesium with Fish Mortalities. Water Res. 1969, 3, 375–379. [Google Scholar] [CrossRef]
- Zamor, R.M.; Franssen, N.R.; Porter, C.; Patton, T.M.; Hambright, K.D. Rapid recovery of a fish assemblage following an ecosystem disruptive algal bloom. Freshw. Sci. 2014, 33, 390–401. [Google Scholar] [CrossRef]
- Larimore, R.W.; Childers, W.F.; Heckrotte, C. Destruction and re-establishment of stream fish and invertebrates affected by drought. Trans. Am. Fish. Soc. 1959, 88, 261–285. [Google Scholar] [CrossRef]
- Matthews, W.J.; Marsh-Mathews, E. Effects of drought on fish across axis of space, time and ecological complexity. Freshw. Biol. 2003, 48, 1232–1253. [Google Scholar] [CrossRef]
- Roghair, C.N.; Dolloff, C.A.; Underwood, M.K. Response of a brook trout population and instream habitat to a catastrophic flood and debris flow. Trans. Am. Fish. Soc. 2002, 131, 718–730. [Google Scholar] [CrossRef]
- Wellman, D.I. Post-Flood Recovery and Distributions of Fishes in the New River Gorge National River, West Virginia. Master’s Thesis, West Virginia University, Morgantown, WV, USA, 2004. Available online: https://researchrepository.wvu.edu/cgi/viewcontent.cgi?article=3005&context=etd (accessed on 9 November 2021).
- Olmsted, L.L.; Cloutman, D.G. Repopulation after a fish kill in Mud Creek, Washington County, Arkansas following a pesticide pollution. Trans. Am. Fish. Soc. 1974, 1, 79–87. [Google Scholar] [CrossRef]
- Niemi, G.J.; DeVore, P.; Detebeck, N.; Taylor, D.; Lima, A.; Pastor, J.; Yount, J.D.; Naiman, R.J. Overview of case studies on recovery of aquatic systems from disturbance. Environ. Mgt. 1990, 14, 571–587. [Google Scholar] [CrossRef]
- Meade, R. Fish and Invertebrate Recolonization in a Missouri Prairie Stream after an Acute Pollution Event. N. Am. J. Fish Mgt. 2004, 24, 7–19. [Google Scholar] [CrossRef]
- Warner, K.; Fenderson, O.C. Effects of DDT spraying for forest insects on Maine trout streams. J. Wild. Mgt. 1962, 26, 86–93. [Google Scholar] [CrossRef]
- Detenbeck, N.E.; Devore, P.W.; Niemi, G.J.; Lima, A. Recovery of temperate-stream fish communities from disturbance: A review of case studies and synthesis of theory. Environ. Mgt. 1992, 16, 33–53. [Google Scholar] [CrossRef]
- Pearsons, T.N.; Li, H.W.; Lamberti, G.I. Influence of habitat complexity on resistance to flooding and resilience of stream fish assemblages. Trans. Am. Fish. Soc. 1992, 121, 427–436. [Google Scholar] [CrossRef]
- Haag, W.R. North American Freshwater Mussels: Natural History, Ecology and Conservation; Cambridge University Press: New York, NY, USA, 2012; pp. 1–535. [Google Scholar]
- Illinois Natural History Museum. Freshwater Mussel Host Database; Illinois Natural History Museum: Champaign, IL, USA, 2017; Available online: https://mollusk.inhs.illinois.edu/57-2/ (accessed on 30 June 2021).
- Ford, D.E.; Oliver, A.M. The known and potential hosts of Texas mussels: Implications for future research and conservation efforts. Freshw. Mollusk Biol. Conserv. 2015, 18, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Haag, W.R.; Warren, M.L., Jr. Host fishes and infection strategies of freshwater mussels in large Mobile Basin streams, USA. J. NA Benthol. Soc. 2003, 22, 78–91. [Google Scholar] [CrossRef]
- Haag, W.R.; Warren, M.L., Jr.; Shillingsford, M. Host fishes and host-attracting behavior of Lampsilis altilis and Villosa vibex (Bivalvia: Unionidae). Am. Mid. Nat. 1999, 14, 149–157. [Google Scholar] [CrossRef]
- Keller, A.E.; Ruessler, D.S. Determination or verification of host fish for nine species of unionid mussels. Am. Mid. Nat. 1997, 138, 402–407. [Google Scholar] [CrossRef]
- Ingersoll, C.G.; Kernaghan, N.J.; Gross, T.S.; Bishop, C.D.; Wang, N.; Roberts, A. Laboratory toxicity testing with freshwater mussels. In Freshwater Bivalve Ecotoxicology; Farris, J.L., van Hassel, J.H., Eds.; SETAC: Pensacola, FL, USA, 2006; pp. 95–134. [Google Scholar]
- Augspurger, T.; Keller, A.E.; Black, M.C.; Cope, G.; Dwyer, F.J. Derivation of water quality guidance for protection of freshwater mussels (Unionidae) from ammonia exposure. Environ. Tox. Chem. 2003, 22, 2569–2575. [Google Scholar] [CrossRef] [PubMed]
- Prosser, R.S.; Rochfort, Q.; McKinnis, R.; Exall, K.; Gillis, P.L. Assessing the toxicity and risk of salt-impacted winter road runoff to the early life stages of freshwater mussels in the Canadian province of Ontario. Environ. Poll. 2017, 230, 589–597. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, P.J.; Neves, R.J.; Cherry, D.S.; Farris, J.L. Sensitivity of glochidial stages of freshwater mussels (Bivalvia: Unionidae) to copper. Environ. Tox. Chem. 2009, 16, 2384–2392. [Google Scholar] [CrossRef]
- Keller, A.E.; Zam, S.G. The acute toxicity of selected metals to the freshwater mussels, Anodonta imbecilis. Environ. Tox. Chem. 1991, 10, 539–546. [Google Scholar] [CrossRef]
- Patnode, K.A.; Hittle, E.; Anderson, R.M.; Zimmerman, L.; Fulton, J.W. Effects of high salinity wastewater discharges on Unionid mussels in the Allegheny River, Pennsylvania. J. Fish Wild. Mgt. 2015, 6, 55–70. [Google Scholar] [CrossRef] [Green Version]
- Latimer, J.S.; Zheng, J. The sources, transport, and fate of PAHs in the marine environment. In PAHs: An Ecotoxicological Perspective; Peter, E., Douben, T., Eds.; John Wiley & Sons Ltd.: London, UK, 2003; pp. 9–33. [Google Scholar]
- West Virginia Department of Environmental Protection. Ambient Water Quality Data Report; West Virginia Department of Environmental Protection: Charleston, WV, USA, 2021. Available online: https://apps.dep.wv.gov/dwwm/wqdatac/ (accessed on 30 June 2021).
- Goudreau, S.E.; Neves, R.J.; Sheehan, R.K. Effects of wastewater treatment plant effluents on freshwater mollusks in the upper Clinch River, Virginia, USA. Hydrobiologia 1993, 252, 211–230. [Google Scholar] [CrossRef]
- Myers-Kinzie, M. Factors Affecting Survival and Recruitment of Unionid Mussels in Small Midwestern Streams. Ph.D. Dissertation, Purdue University, West Lafayette, IN, USA, 1998. [Google Scholar]
- Imlay, M.J. Effects of potassium on survival and distribution of freshwater mussels. Malacologia 1973, 12, 97–113. [Google Scholar]
- Cope, W.G.; Bregeron, C.M.; Archambault, J.M.; Jones, J.W.; Beay, B.; Lazaro, P.R.; Shea, D.; Callihan, J.L.; Rogers, J.J. Understanding the influence of multiple pollutant stressors on the decline of freshwater mussels in a biodiversity hotspot. Sci. Total Environ. 2021, 773, 144757. [Google Scholar] [CrossRef] [PubMed]
- United States Environmental Protection Agency (USEPA). Update on Dunkard Creek. US EPA Region 3; Environmental Analysis and Innovation Division, Office of Monitoring and Assessment: Wheeling, WV, USA, 2009. Available online: https://dep.wv.gov/WWE/hot_topics/Documents/Dunkard%20Fish%20Kill%20Documents/Other%20docs%20Not%20sampling%20docs/mtmorrriswvdep.pdf (accessed on 9 November 2021).
- Klose, K.; Cooper, S.D.; Leydecker, A.D.; Kreitler, J. Relationships among catchment land use and concentrations of nutrients, algae, and dissolved oxygen in a southern California river. Freshw. Sci. 2012, 31, 908–927. [Google Scholar] [CrossRef]
- Morgan, A.M.; Royer, T.V.; David, M.B.; Gentry, L.E. Relationships among nutrients, chlorophyll-a, and dissolved oxygen in agricultural streams in Illinois. J. Environ. Qual. 2006, 35, 1110–1117. [Google Scholar] [CrossRef] [Green Version]
- Adelman, I.R.; Smith, L.L.J.; Siesennop, G.D. Acute toxicity of sodium chloride, pentachlorophenol, guthion, and hexavalent chromium to fathead minnows (Pimephales promelas) and goldfish (Carassius auratus). J. Fish. Res. Board Can. 1976, 33, 203–208. [Google Scholar] [CrossRef]
- Birge, W.J.; Black, J.A.; Westerman, A.G.; Short, T.M.; Taylor, S.B.; Bruser, D.M.; Wallingford, E.D. Recommendations on Numerical Values for Regulating Iron and Chloride Concentrations for the Purpose of Protecting Warm Water Species of Aquatic Life in the Commonwealth of Kentucky; Memorandum of Agreement; Kentucky Natural Resources and Environmental Protection Cabinet: Lexington, KY, USA, 1985; Volume 5429, p. 74.
- Trama, F.B. The Acute Toxicity of Some Common Salts of Sodium, Potassium and Calcium to the Common Bluegill (Lepomis macrochirus Rafinesque). Proc. Acad. Nat. Sci. Phil. 1954, 106, 195–205. Available online: https://www.jstor.org/stable/4064450 (accessed on 9 November 2021).
- Wallen, I.E.; Greer, W.C.; Lasater, R. Toxicity to Gambusia affinis of certain pure chemicals in turbid waters. Sewage Ind. Wastes 1957, 29, 695–711. [Google Scholar]
- Reed, P.; Evans, R. Acute toxicity of chlorides, sulfates, and total dissolved solids to some fishes in Illinois. Illinois Dept. Energy Nat. Res. 1981, 283, 51. Available online: https://www.ideals.illinois.edu/bitstream/handle/2142/72701/ISWSCR-283.pdf?sequence=2 (accessed on 9 November 2021).
- Kimmel, W.G.; Argent, D.G. Status of fish and macroinvertebrate communities in a watershed experiencing high rates of fossil fuel extraction: Tenmile Creek, a major Monongahela River tributary. Wat. Air Soil Poll. 2012, 223, 4647–4657. [Google Scholar] [CrossRef]
- Kimmel, W.G.; Argent, D.G. Stream fish community responses to a gradient of specific conductance. Wat. Air Soil Poll. 2010, 206, 49–56. [Google Scholar] [CrossRef]
- Kimmel, W.G.; Argent, D.G. Scuds (Gammaridae) and Darters (Percidae) Dominate Aquatic Communities in a Stream Exhibiting Levels of Specific Conductance Exceeding 4,000 μS/cm. Int. J. Environ. Monit. Anal. 2018, 6, 47–52. [Google Scholar] [CrossRef]
- Cañedo-Argüelles, M.; Kefford, B.J.; Piscart, C.; Prat, N.; Schafer, R.B.; Schulz, C.-J. Salinization of rivers: An urgent ecological issue. Environ. Poll. 2013, 173, 157–167. [Google Scholar] [CrossRef]
- Evans, D.M.; Zipper, C.E.; Donovan, P.F.; Daniels, W.L. Long-term trends of specific conductance in waters discharged by coal-mine valley fills in Central Appalachia, USA. J. Am. Water Res. Assoc. 2014, 50, 1449–1460. [Google Scholar] [CrossRef]
- Kaushal, S.S.; Likens, G.E.; Pace, M.L.; Utz, R.M.; Shahan, H.; Gorman, J.; Grese, M. Freshwater salinization syndrome on a continental scale. Proc. Natl. Acad. Sci. USA 2018, 115, E574–E583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, J.; Fanelli, R.M.; Sekellick, A.J. High-Frequency Data Reveal Deicing Salts Drive Elevated Specific Conductance and Chloride along with Pervasive and Frequent Exceedances of the U.S. Environmental Protection Agency Aquatic Life Criteria for Chloride in Urban Streams. Environ. Sci. Technol. 2020, 54, 778–789. [Google Scholar] [CrossRef] [PubMed]
- Novotny, E.V.; Murphy, D.; Stefan, H.G. Increase of urban lake salinity by road deicing salt. Sci. Total Environ. 2008, 406, 131–144. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RKM | Location | Temp_C | Sp.Cond (uS/cm) | DO_mg/L | DO % Sat | pH |
|---|---|---|---|---|---|---|
| 5.1 | Dunkard Creek upstream of Dolls Run | 19.7 | 2257 | 8.67 | 95.3 | 8.28 |
| 12.0 | Dunkard Creek in Pentress | 20.1 | 2714 | 13.93 | 154.8 | 8.37 |
| 19.0 | Dunkard Creek in Blacksville | 19.5 | 3259 | 9.52 | 104.7 | 8.2 |
| 22.7 | Dunkard Creek downstream of Miracle Run | 19.4 | 3911 | 8.85 | 97.3 | 8.13 |
| 24.0 | Dunkard Creek upstream of Morris Run | 20.67 | 5085 | 10.36 | 117.4 | 8.39 |
| n/a | Hoovers Run (tributary to Dunkard) | 18.8 | 770 | 8.64 | 92.8 | 8.45 |
| n/a | PA Fork Dunkard at T309 Bridge | 18.95 | 672 | 7.88 | 84.7 | 8.02 |
| 27.1 | WV Fork Dunkard downstream of Consol Outfall | 21.93 | 18,570 | 13.45 | 165.8 | 8.17 |
| 27.0 | Consol Outfall 005 WV 0064602 | 22.64 | 25,250 | 8.34 | 105.3 | 8.55 |
| 26.9 | WV Fork Dunkard upstream of Consol Outfall | 20.62 | 4957 | 11.54 | 130.7 | 8.13 |
| Species | Common Name | Year | ||||||
|---|---|---|---|---|---|---|---|---|
| 1919 | 1969–1980 | 1992–1997 | 2000–2007 | 2009 | 2010–2018 | Stocked 2010–2018 | ||
| Amblema plicata | Threeridge | X | X | X | X | X | S | |
| Cyclonaias tuberculata | Purple Wartyback | X | ||||||
| Epioblasma triquetra | Snuffbox | X | X | X | X | X | ||
| Eurynia dilatata | Lady Finger | X | X | X | X | X | ||
| Fusconaia flava | Wabash Pigtoe | X | X | X | X | X | S | |
| Lampsilis cardium | Plain Pocketbook | X | X | X | X | X | S | |
| Lampsilis fasciola | Wavyrayed Lampmussel | X | S | |||||
| Lampsilis siliquoidea | Fatmucket | X | X | X | X | X | S | |
| Lasmigona costata | Fluted-Shell | X | X | X | X | X | S | |
| Leptodea fragilis | Fragile Papershell | X | S | |||||
| Ligumia recta | Black Sandshell | X | ||||||
| Pleurobema clava | Clubshell | X | ||||||
| Pleurobema sintoxia | Round Pigtoe | X | X | X | X | S | ||
| Potamilus alatus | Pink Heelsplitter | X | X | S | ||||
| Ptychobranchus fasciolaris | Kidney Shell | X | X | X | X | X | S | |
| Pyganodon grandis | Giant Floater | X | X | X | X | X | S | |
| Simpsonaias ambigua | Salamander Mussel | X | X | X | ||||
| Strophitus undulatus | Creeper | X | X | X | X | X | S | |
| Tritogonia verrucosa | Buckhorn | X | X | X | S | |||
| Utterbackia imbecillis | Paper Pondshell | X | X | S | ||||
| Villosa iris | Rainbow-Shell | X | X | X | X | S | ||
| Species Richness | 16 | 15 | 15 | 11 | 16 | 0 | 15 | |
| Taxa | Common Name | 1983–2005 | 2009 | 2010–2019 | Total |
|---|---|---|---|---|---|
| Darters | |||||
| Etheostoma blennioides | Greenside Darter | 1 | 0 | 1 | 2508 |
| Etheostoma caeruleum | Rainbow Darter | 1 | 0 | 1 | 463 |
| Etheostoma flabellare | Fantail Darter | 1 | 0 | 1 | 324 |
| Etheostoma nigrum | Johnny Darter | 1 | 0 | 1 | 161 |
| Etheostoma variatum | Variegate Darter | 1 | 0 | 1 | 1800 |
| Etheostoma zonale | Banded Darter | 1 | 0 | 1 | 1049 |
| Percina caprodes | Logperch | 1 | 0 | 1 | 275 |
| Percina maculata | Blackside Darter | 1 | 0 | 1 | 74 |
| Minnows | |||||
| Campostoma anomalum | Central Stoneroller | 1 | 1 | 1 | 5808 |
| Cyprinella spiloptera | Spotfin Shiner | 1 | 0 | 1 | 1140 |
| Ericymba buccata | Silverjaw Minnow | 1 | 0 | 1 | 329 |
| Labidesthes sicculus | Brook Silverside | 1 | 0 | 1 | 85 |
| Luxilus chrysocephalus | Striped Shiner | 1 | 1 | 1 | 1796 |
| Notemigonus crysloeucas | Golden Shiner | 0 | 0 | 0 | 1 |
| Notropis atherinoides | Emerald Shiner | 1 | 0 | 1 | 388 |
| Notropis photogenis | Silver Shiner | 1 | 0 | 1 | 26 |
| Notropis rubellus | Rosyface Shiner | 1 | 0 | 1 | 1394 |
| Notropis stramineus | Sand Shiner | 1 | 0 | 1 | 4019 |
| Notropis volucellus | Mimic Shiner | 1 | 0 | 1 | 3703 |
| Pimephales notatus | Bluntnose Minnow | 1 | 1 | 1 | 9598 |
| Semotilus atromaculatus | Creek Chub | 1 | 0 | 1 | 273 |
| Suckers | |||||
| Carpiodes cyprinus | Quillback | 1 | 0 | 1 | 2 |
| Catostomus commersoni | White Sucker | 1 | 0 | 1 | 423 |
| Hypentelium nigricans | Northern Hogsucker | 1 | 1 | 1 | 1839 |
| Moxostoma anisurum | Silver Redhorse | 0 | 0 | 1 | 203 |
| Moxostoma duquesnei | Black Redhorse | 1 | 0 | 1 | 167 |
| Moxostoma erythrurum | Golden Redhorse | 1 | 0 | 1 | 357 |
| Game Fishes | |||||
| Ambloplites rupestris | Rock Bass | 1 | 0 | 1 | 88 |
| Ameiurus natalis | Yellow Bullhead | 1 | 0 | 1 | 48 |
| Esox masquinongy | Muskellunge | 1 | 0 | 1 | 2 |
| Ictalurus punctatus | Channel Catfish | 0 | 0 | 1 | 16 |
| Lepomis cyanellus | Green Sunfish | 1 | 0 | 1 | 34 |
| Lepomis gibbosus | Pumpkinseed | 1 | 0 | 1 | 24 |
| Lepomis macrochirus | Bluegill | 1 | 0 | 1 | 120 |
| Micropterus dolomieu | Smallmouth Bass | 1 | 0 | 1 | 234 |
| Micropterus salmoides | Largemouth Bass | 0 | 0 | 1 | 17 |
| Pylodictus olivaris | Flathead Catfish | 1 | 0 | 0 | 11 |
| Other | |||||
| Noturus flavus | Stonecat | 1 | 0 | 1 | 299 |
| Taxa | Common Name | 2005 Base Abundance | 1983 | 1997 | 2009 | 2010 | 2014 | 2016 | 2019 |
|---|---|---|---|---|---|---|---|---|---|
| Darters | |||||||||
| Etheostoma blennioides | Greenside Darter | 82 | 1.37 | 6.98 | 0.00 | 2.66 | 2.33 | 7.37 | 8.89 |
| Etheostoma caeruleum | Rainbow Darter | 49 | 0.39 | 2.71 | 0.00 | 0.88 | 0.82 | 2.16 | 1.49 |
| Etheostoma flabellare | Fantail Darter | 7 | 17.86 | 17.86 | 0.00 | 2.43 | 1.43 | 2.29 | 3.43 |
| Etheostoma nigrum | Johnny Darter | 6 | 4.00 | 2.17 | 0.00 | 4.17 | 3.17 | 3.50 | 8.83 |
| Etheostoma variatum | Variegate Darter | 78 | 0.32 | 5.27 | 0.00 | 1.71 | 1.08 | 5.53 | 8.18 |
| Etheostoma zonale | Banded Darter | 97 | 0.11 | 4.64 | 0.00 | 0.05 | 0.28 | 1.07 | 3.66 |
| Percina caprodes | Logperch | 20 | 5.15 | 5.35 | 0.00 | 0.60 | 0.20 | 1.10 | 0.35 |
| Percina maculata | Blackside Darter | 5 | 3.40 | 3.60 | 0.00 | 1.60 | 2.40 | 1.80 | 1.00 |
| Minnows | |||||||||
| Campostoma anomalum | Central Stoneroller | 1902 | 0.12 | 0.62 | 0.00 | 0.28 | 0.22 | 0.43 | 0.39 |
| Cyprinella spiloptera | Spotfin Shiner | 100 | 0.00 | 3.15 | 0.00 | 0.21 | 0.41 | 3.72 | 2.91 |
| Ericymba buccata | Silverjaw Minnow | 12 | 0.00 | 3.92 | 0.00 | 4.25 | 3.67 | 12.83 | 1.75 |
| Labidesthes sicculus | Brook Silverside | 4 | 0.00 | 18.00 | 0.00 | 0.00 | 0.25 | 2.00 | 0.00 |
| Luxilus chrysocephalus | Striped Shiner | 121 | 3.98 | 3.18 | 0.09 | 0.70 | 1.03 | 1.88 | 2.97 |
| Notemigonus crysloeucas | Golden Shiner | 0 | -- | -- | -- | -- | -- | *1 | -- |
| Notropis atherinoides | Emerald Shiner | 0 | *385 | -- | -- | *3 | -- | -- | -- |
| Notropis photogenis | Silver Shiner | 0 | -- | -- | -- | -- | -- | *2 | *24 |
| Notropis rubellus | Rosyface Shiner | 135 | 0.00 | 3.25 | 0.00 | 0.23 | 1.01 | 2.52 | 2.32 |
| Notropis stramineus | Sand Shiner | 121 | 6.10 | 5.63 | 0.00 | 0.77 | 7.37 | 7.88 | 4.47 |
| Notropis volucellus | Mimic Shiner | 66 | 0.00 | 29.12 | 0.00 | 1.88 | 1.45 | 8.14 | 14.52 |
| Pimephales notatus | Bluntnose Minnow | 1860 | 0.71 | 1.17 | 0.00 | 0.12 | 0.77 | 0.83 | 0.55 |
| Semotilus atromaculatus | Creek Chub | 17 | 3.76 | 3.71 | 0.00 | 3.00 | 2.41 | 0.94 | 1.24 |
| Suckers | |||||||||
| Carpiodes cyprinus | Quillback | 1 | 0.00 | 0.00 | 0.00 | 1.00 | 0.00 | 0.00 | 0.00 |
| Catostomus commersoni | White Sucker | 34 | 0.94 | 0.74 | 0.00 | 4.15 | 3.79 | 1.15 | 0.68 |
| Hypentelium nigricans | Northern Hogsucker | 240 | 0.58 | 0.89 | 0.00 | 0.74 | 1.63 | 1.17 | 1.64 |
| Moxostoma anisurum | Silver Redhorse | 2 | 0.00 | 0.50 | 0.00 | 50.50 | 46.00 | 0.50 | 3.00 |
| Moxostoma duquesnei | Black Redhorse | 15 | 0.00 | 1.07 | 0.00 | 0.27 | 0.67 | 7.60 | 0.53 |
| Moxostoma erythrurum | Golden Redhorse | 50 | 0.32 | 0.54 | 0.00 | 0.90 | 1.66 | 1.90 | 0.82 |
| Game Fishes | |||||||||
| Ambloplites rupestris | Rock Bass | 8 | 4.00 | 4.00 | 0.00 | 0.00 | 0.38 | 1.38 | 0.25 |
| Ameiurus natalis | Yellow Bullhead | 7 | 1.71 | 2.29 | 0.00 | 0.14 | 0.14 | 1.57 | 0.00 |
| Esox masquinongy | Muskellunge | 1 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 1.00 | 0.00 |
| Ictalurus punctatus | Channel Catfish | 0 | -- | -- | -- | -- | *1 | *4 | *11 |
| Lepomis cyanellus | Green Sunfish | 1 | 12.00 | 14.00 | 0.00 | 0.00 | 0.00 | 7.00 | 0.00 |
| Lepomis gibbosus | Pumpkinseed | 0 | |||||||
| Lepomis macrochirus | Bluegill | 1 | 112.00 | 0.00 | 0.00 | 1.00 | 1.00 | 4.00 | 1.00 |
| Micropterus dolomieu | Smallmouth Bass | 31 | 1.61 | 1.29 | 0.00 | 0.39 | 0.45 | 2.06 | 0.74 |
| Micropterus salmoides | Largemouth Bass | 0 | -- | -- | -- | *2 | *12 | *3 | -- |
| Pylodictus olivaris | Flathead Catfish | 0 | -- | *11 | -- | -- | -- | -- | -- |
| Other | |||||||||
| Noturus flavus | Stonecat | 5 | 2.40 | 47.00 | 0.00 | 0.60 | 0.80 | 4.00 | 4.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hartman, K.J.; Wellman, D.I., Jr.; Kingsbury, J.W.; Cincotta, D.A.; Clayton, J.L.; Eliason, K.M.; Jernejcic, F.A.; Owens, N.V.; Smith, D.M. A Case Study of a Prymnesium parvum Harmful Algae Bloom in the Ohio River Drainage: Impact, Recovery and Potential for Future Invasions/Range Expansion. Water 2021, 13, 3233. https://doi.org/10.3390/w13223233
Hartman KJ, Wellman DI Jr., Kingsbury JW, Cincotta DA, Clayton JL, Eliason KM, Jernejcic FA, Owens NV, Smith DM. A Case Study of a Prymnesium parvum Harmful Algae Bloom in the Ohio River Drainage: Impact, Recovery and Potential for Future Invasions/Range Expansion. Water. 2021; 13(22):3233. https://doi.org/10.3390/w13223233
Chicago/Turabian StyleHartman, Kyle J., David I. Wellman, Jr., Joseph W. Kingsbury, Daniel A. Cincotta, Janet L. Clayton, Kevin M. Eliason, Frank A. Jernejcic, Nathaniel V. Owens, and Dustin M. Smith. 2021. "A Case Study of a Prymnesium parvum Harmful Algae Bloom in the Ohio River Drainage: Impact, Recovery and Potential for Future Invasions/Range Expansion" Water 13, no. 22: 3233. https://doi.org/10.3390/w13223233