
Fish Assemblages in Seagrass (Zostera marina L.) Meadows and Mussel Reefs (Mytilus edulis): Implications for Coastal Fisheries, Restoration and Marine Spatial Planning

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
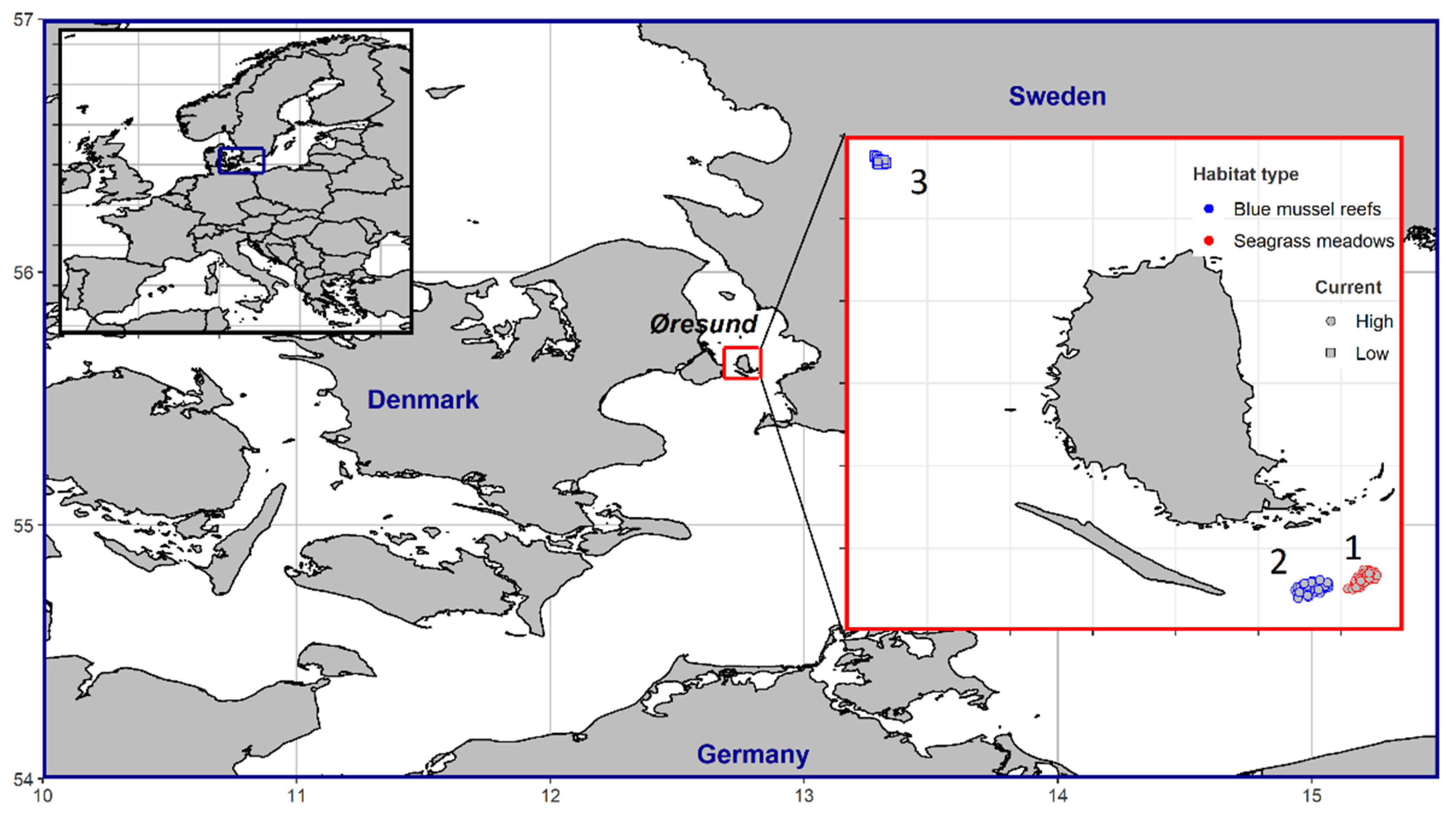
2.1. Study Area
2.2. Fish Assemblages in Z. marina Meadows and M. edulis Reefs
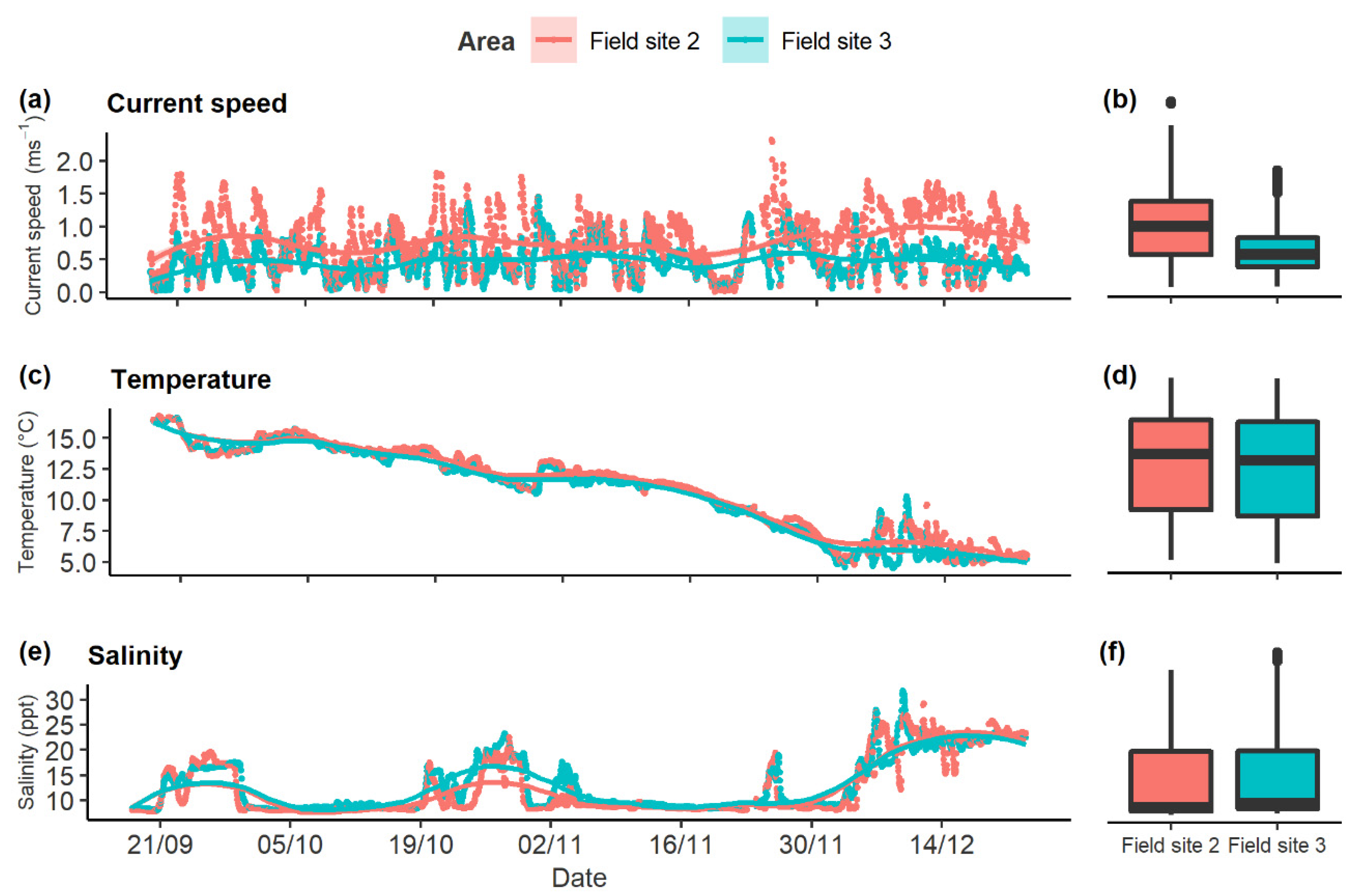
2.3. Fish Assemblages in M. edulis Reefs with High and Low Water Currents
2.4. Statistical Analysis
2.4.1. Statistical Analysis of Fyke Nets Data
2.4.2. Statistical Analysis of Video Cameras’ Data
3. Results
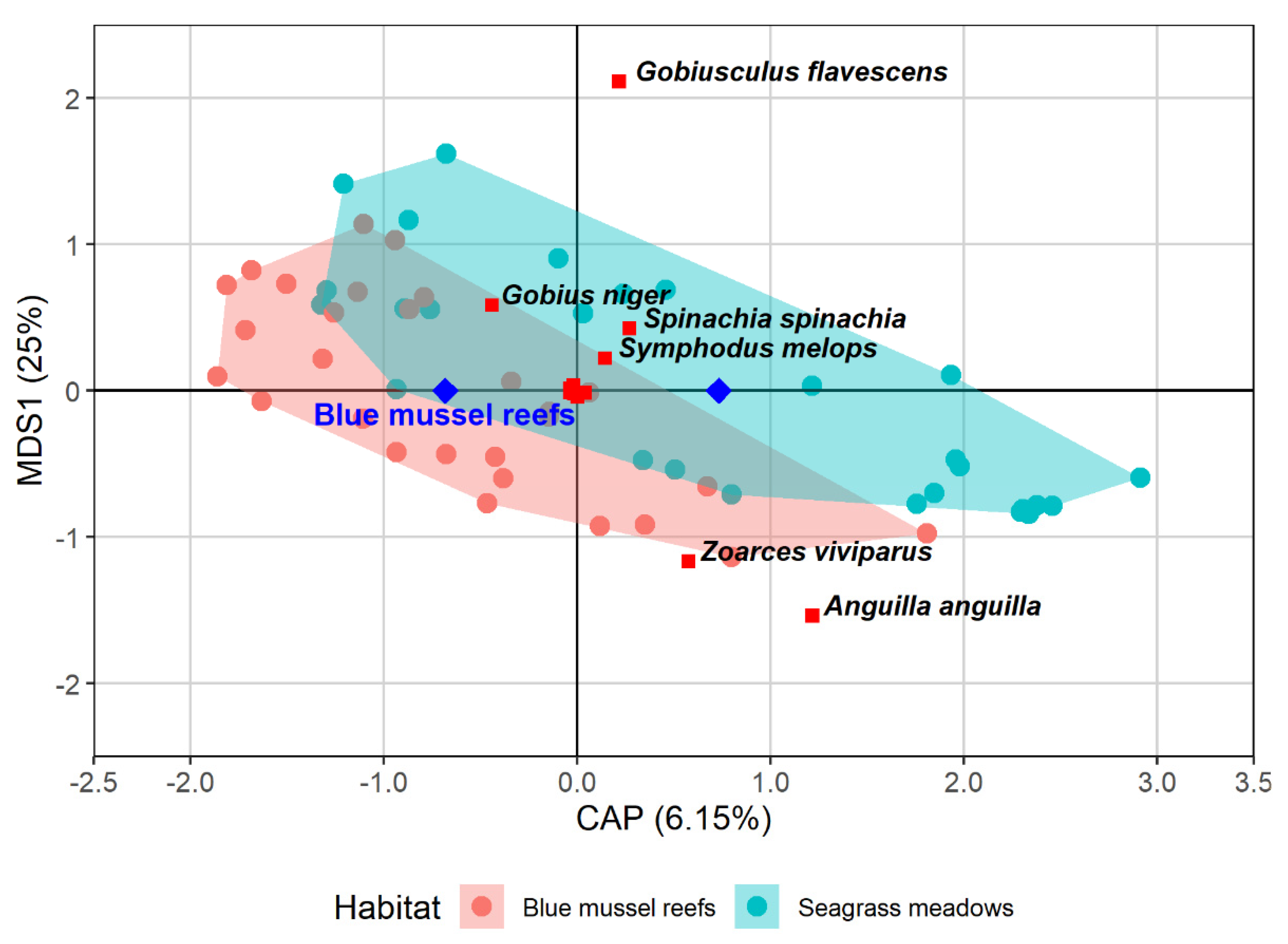
3.1. Using Fyke Nets to Estimate Fish Assemblage in Seagrass Meadows and Mussel Reefs
3.2. Using Stationary Underwater Cameras to Estimate Fish Assemblage in Two Different Mussel Reef Habitats
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van der Zee, E.M.; Angelini, C.; Govers, L.L.; Christianen, M.; Altieri, A.H.; van der Reijden, K.J.; Silliman, B.R.; van de Koppel, J.; Van der Geest, M.; van Gils, J.A.; et al. How habitat-modifying organisms structure the food web of two coastal ecosystems. Proc. R. Soc. B Biol. Sci. 2016, 283, 20152326. [Google Scholar] [CrossRef] [Green Version]
- Peterson, C.H.; Grabowski, J.H.; Powers, S.P. Estimated enhancement of fish production resulting from restoring oyster reef habitat: Quantitative valuation. Mar. Ecol. Prog. Ser. 2003, 264, 249–264. [Google Scholar] [CrossRef] [Green Version]
- Orth, R.J.; Carruthers, T.J.B.; Dennison, W.C.; Duarte, C.M.; Fourqurean, J.W.; Heck, K.L.; Hughes, A.R.; Kendrick, G.A.; Kenworthy, W.J.; Olyarnik, S.; et al. A global crisis for seagrass ecosystems. Bioscience 2006, 56, 987–996. [Google Scholar] [CrossRef] [Green Version]
- Pihl, L.; Baden, S.; Kautsky, N.; Rönnbäck, P.; Söderqvist, T.; Troell, M.; Wennhage, H. Shift in fish assemblage structure due to loss of seagrass Zostera marina habitats in Sweden. Estuar. Coast. Shelf Sci. 2006, 67, 123–132. [Google Scholar] [CrossRef]
- Andersen, J.H.; Laamanen, M.; Aigars, J.; Axe, P.; Blomqvist, M.; Carstensen, J.; Josefson, A.B.; Fleming-Lehtinen, V.; Järvinen, M.; Kaartokallio, H.; et al. HELCOM eutrophication in the Baltic Sea—An integrated thematic assessment of the effects of nutrient enrichment and eutrophication in the Baltic Sea region. In Baltic Sea Environment Proceedings (BSEP) No. 119; Helsinki Commission: Helsinki, Finland, 2009; Volume 115B, p. 148. [Google Scholar]
- Eriksson, B.K.; van der Heide, T.; van de Koppel, J.; Piersma, T.; van der Veer, H.W.; Olff, H. Major changes in the ecology of the wadden sea: Human impacts, ecosystem engineering and sediment dynamics. Ecosystems 2010, 13, 752–764. [Google Scholar] [CrossRef] [Green Version]
- Beck, M.W.; Heck, K.L.; Able, K.W.; Childers, D.L.; Eggleston, D.B.; Gillanders, B.M.; Halpern, B.; Hays, C.G.; Hoshino, K.; Minello, T.J.; et al. The identification, conservation, and management of estuarine and marine nurseries for fish and invertebrates. Bioscience 2001, 51, 633–641. [Google Scholar] [CrossRef]
- Pomeroy, L.R.; D’Elia, C.F.; Schaffner, L.C. Top-down control of phytoplankton by oysters in Chesapeake Bay, USA: Reply to Newell et al. (2007). Mar. Ecol. Prog. Ser. 2007, 341, 299–301. [Google Scholar] [CrossRef]
- Grabowski, J.H.; Peterson, C.H. Restoring oyster reefs to recover ecosystem services. In Theoretical Ecology Series; Academic Press: Cambridge, MA, USA, 2007; Volume 4, pp. 281–298. ISBN 9780123738578. [Google Scholar]
- Seitz, R.D.; Wennhage, H.; Bergström, U.; Lipcius, R.N.; Ysebaert, T. Ecological value of coastal habitats for commercially and ecologically important species. ICES J. Mar. Sci. J. Cons. 2013. [Google Scholar] [CrossRef] [Green Version]
- Kuusemäe, K.; Rasmussen, E.K.; Canal-Vergés, P.; Flindt, M.R. Modelling stressors on the eelgrass recovery process in two Danish estuaries. Ecol. Modell. 2016, 333, 11–42. [Google Scholar] [CrossRef]
- Whitfield, A.K. The role of seagrass meadows, mangrove forests, salt marshes and reed beds as nursery areas and food sources for fishes in estuaries. Rev. Fish Biol. Fish. 2016, 1–36. [Google Scholar] [CrossRef]
- Lefcheck, J.S.; Hughes, B.B.; Johnson, A.J.; Pfirrmann, B.W.; Rasher, D.B.; Smyth, A.R.; Williams, B.L.; Beck, M.W.; Orth, R.J. Are coastal habitats important nurseries? A meta-analysis. Conserv. Lett. 2019, 12, e12645. [Google Scholar] [CrossRef]
- Bromley, C.; McGonigle, C.; Ashton, E.C.; Roberts, D. Restoring degraded European native oyster, Ostrea edulis, habitat: Is there a case for harrowing? Hydrobiologia 2016, 768, 151–165. [Google Scholar] [CrossRef] [Green Version]
- Norling, P.; Lindegarth, M.; Lindegarth, S.; Strand, Å. Effects of live and post-mortem shell structures of invasive Pacific oysters and native blue mussels on macrofauna and fish. Mar. Ecol. Prog. Ser. 2015, 518, 123–138. [Google Scholar] [CrossRef] [Green Version]
- Gain, I.E.; Brewton, R.A.; Reese Robillard, M.M.; Johnson, K.D.; Smee, D.L.; Stunz, G.W. Macrofauna using intertidal oyster reef varies in relation to position within the estuarine habitat mosaic. Mar. Biol. 2016, 164, 8. [Google Scholar] [CrossRef]
- Kristensen, L.D.; Stenberg, C.; Støttrup, J.; Poulsen, L.K.; Christensen, H.T.; Dolmer, P.; Landes, A.; Røjbek, M.; Thorsen, S.W.; Holmer, M.; et al. Establishment of blue mussel beds to enhance fish habitats. Appl. Ecol. Environ. Res. 2015, 13, 783–798. [Google Scholar] [CrossRef]
- Norling, P.; Kautsky, N. Patches of the mussel Mytilus sp. are islands of high biodiversity in subtidal sediment habitats in the Baltic Sea. Aquat. Biol. 2008, 4, 75–87. [Google Scholar] [CrossRef] [Green Version]
- Koivisto, M.E. Blue Mussel Beds as Biodiversity Hotspots on the Rocky Shores of the Northern Baltic Sea. Ph.D. Thesis, University of Helsinki, Helsinki, Finland, 2011. [Google Scholar]
- Reusch, T.; Chapman, A.; Groger, J. Blue mussels Mytilus edulis do not interfere with eelgrass Zostera marina but fertilize shoot growth through biodeposition. Mar. Ecol. Prog. Ser. 1994, 108, 265–282. [Google Scholar] [CrossRef]
- Schubert, P.; Hukriede, W.; Karez, R.; Reusch, T. Mapping and modeling eelgrass Zostera marina distribution in the western Baltic Sea. Mar. Ecol. Prog. Ser. 2015, 522, 79–95. [Google Scholar] [CrossRef] [Green Version]
- Cullen-Unsworth, L.; Unsworth, R. Seagrass meadows, ecosystem services, and sustainability. Environ. Sci. Policy Sustain. Dev. 2013, 55, 14–28. [Google Scholar] [CrossRef]
- Cavanagh, R.D.; Broszeit, S.; Pilling, G.M.; Grant, S.M.; Murphy, E.J.; Austen, M.C. Valuing biodiversity and ecosystem services: A useful way to manage and conserve marine resources? Proc. R. Soc. B Biol. Sci. 2016, 283, 20161635. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez, J.L.; Jones, C.G.; Byers, J.E.; Arkema, K.K.; Berkenbusch, K.; Commito, A.; Duarte, C.M.; Hacker, S.D.; Lambrinos, J.G.; Hendriks, I.E.; et al. Physical ecosystem engineers and the functioning of estuaries and coasts. In Treatise on Estuarine and Coastal Science; Wolanski, E., McLusky, D., Eds.; Academic Press: Waltham, MA, USA, 2012; Volume 7, pp. 53–81. ISBN 9780080878850. [Google Scholar]
- Hovel, K.A.; Warneke, A.M.; Virtue-Hilborn, S.P.; Sanchez, A.E. Mesopredator foraging success in eelgrass (Zostera marina L.): Relative effects of epiphytes, shoot density, and prey abundance. J. Exp. Mar. Bio. Ecol. 2016, 474, 142–147. [Google Scholar] [CrossRef]
- Bertelli, C.M.; Unsworth, R.K.F. Protecting the hand that feeds us: Seagrass (Zostera marina) serves as commercial juvenile fish habitat. Mar. Pollut. Bull. 2014, 83, 425–429. [Google Scholar] [CrossRef]
- Furness, E.; Unsworth, R.K.F. Demersal fish assemblages in NE Atlantic seagrass and Kelp. Diversity 2020, 12, 366. [Google Scholar] [CrossRef]
- Nordlund, L.M.; Unsworth, R.K.F.; Gullström, M.; Cullen-Unsworth, L.C. Global significance of seagrass fishery activity. Fish Fish. 2018, 19, 399–412. [Google Scholar] [CrossRef]
- Heck, K.L.; Able, K.W.; Fahay, M.P.; Roman, C.T. Fishes and decapod crustaceans of cape cod eelgrass meadows: Species composition, seasonal abundance patterns and comparison with unvegetated substrates. Estuaries 1989, 12, 59. [Google Scholar] [CrossRef]
- Unsworth, R.K.F.; Nordlund, L.M.; Cullen-Unsworth, L.C. Seagrass meadows support global fisheries production. Conserv. Lett. 2019, 12, e12566. [Google Scholar] [CrossRef]
- Layman, C.A.; Jud, Z.R.; Albrey Arrington, D.; Sabin, D. Using fish behavior to assess habitat quality of a restored oyster reef. Ecol. Restor. 2014, 32, 140–143. [Google Scholar] [CrossRef]
- Mark, L.; Harvey, C.J.; Anderson, L.E.; Guerry, A.D.; Ruckelshaus, M.H. The role of eelgrass in marine community interactions and ecosystem services: Results from Ecosystem-Scale Food Web Models. Ecosystems 2013, 16, 237–251. [Google Scholar] [CrossRef]
- Fariñas-Franco, J.M.; Roberts, D. Early faunal successional patterns in artificial reefs used for restoration of impacted biogenic habitats. Hydrobiologia 2014, 727, 75–94. [Google Scholar] [CrossRef]
- Grabowski, J.H. Habitat complexity disrupts predator-prey interactions but not the trophic cascade on oyster reefs. Ecology 2004, 85, 995–1004. [Google Scholar] [CrossRef] [Green Version]
- McDermott, S.; Burdick, D.; Grizzle, R.; Greene, J. Restoring ecological functions and increasing community awareness of an urban tidal pond using blue mussels. Ecol. Restor. 2008, 26, 254–262. [Google Scholar] [CrossRef]
- Gratwicke, B.; Speight, M.R. The relationship between fish species richness, abundance and habitat complexity in a range of shallow tropical marine habitats. J. Fish Biol. 2005, 66, 650–667. [Google Scholar] [CrossRef]
- Hosack, G.R.; Dumbauld, B.R.; Ruesink, J.L.; Armstrong, D.A. Habitat associations of estuarine species: Comparisons of intertidal mudflat, seagrass (Zostera marina), and oyster (Crassostrea gigas) habitats. Estuaries Coasts 2006, 29, 1150–1160. [Google Scholar] [CrossRef]
- Borland, H.P.; Gilby, B.L.; Henderson, C.J.; Leon, J.X.; Schlacher, T.A.; Connolly, R.M.; Pittman, S.J.; Sheaves, M.; Olds, A.D. The influence of seafloor terrain on fish and fisheries: A global synthesis. Fish Fish. 2021, 22, 707–734. [Google Scholar] [CrossRef]
- Bey, C.R.; Sullivan, S.M.P. Associations between stream hydrogeomorphology and co-dependent mussel-fish assemblages: Evidence from an Ohio, USA river system. Aquat. Conserv. Mar. Freshw. Ecosyst. 2015, 25, 555–568. [Google Scholar] [CrossRef]
- Kerschbaumer, P.; Tritthart, M.; Keckeis, H. Abundance, distribution, and habitat use of fishes in a large river (Danube, Austria): Mobile, horizontal hydroacoustic surveys vs. a standard fishing method. ICES J. Mar. Sci. 2020, 77, 1966–1978. [Google Scholar] [CrossRef]
- Eggertsen, L.; Hammar, L.; Gullström, M. Effects of tidal current-induced flow on reef fish behaviour and function on a subtropical rocky reef. Mar. Ecol. Prog. Ser. 2016, 559. [Google Scholar] [CrossRef]
- Schmiing, M.; Afonso, P.; Tempera, F. Predictive habitat modelling of reef fishes with contrasting trophic ecologies. Mar. Ecol. Prog. Ser. 2013, 474, 201–216. [Google Scholar] [CrossRef] [Green Version]
- Alexander, S.; Aronson, J.; Whaley, O.; Lamb, D. The relationship between ecological restoration and the ecosystem services concept. Ecol. Soc. 2016, 21, 34. [Google Scholar] [CrossRef] [Green Version]
- Pinnix, W.D.; Shaw, T.A.; Acker, K.C.; Hetrick, N.J. Fish Communities in Eelgrass, Oyster Culture and Mudflat Habitats of North Humboldt Bay, California, Final Report; USFWS Arcata Fisheries Technical Report TR2005-02; Arcata Fish and Wildlife Office: Arcata, CA, USA, 2005; Volume 95521. [Google Scholar]
- Brown, T.; Bergstrom, J.; Loomis, J. Defining, valuing, and providing ecosystem goods and services. Nat. Resour. J. 2007, 47, 329–376. [Google Scholar]
- McCormick, H.; Salguero-Gómez, R.; Mills, M.; Davis, K. Using a residency index to estimate the economic value of coastal habitat provisioning services for commercially important fish species. Conserv. Sci. Pract. 2021, 3, e363. [Google Scholar] [CrossRef]
- Watermeyer, K.E.; Hutchings, L.; Jarre, A.; Shannon, L.J. Patterns of distribution and spatial indicators of ecosystem change based on key species in the southern Benguela. PLoS ONE 2016, 11, e0158734. [Google Scholar] [CrossRef] [Green Version]
- Long, R.D.; Charles, A.; Stephenson, R.L. Key principles of marine ecosystem-based management. Mar. Policy 2015, 57, 53–60. [Google Scholar] [CrossRef]
- Boström, C.; Baden, S.; Bockelmann, A.C.; Dromph, K.; Fredriksen, S.; Gustafsson, C.; Krause-Jensen, D.; Möller, T.; Nielsen, S.L.; Olesen, B.; et al. Distribution, structure and function of Nordic eelgrass (Zostera marina) ecosystems: Implications for coastal management and conservation. Aquat. Conserv. Mar. Freshw. Ecosyst. 2014, 24, 410–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, A.; Lyngby, J.E. Environmental management and monitoring at the Øresund fixed link. Terra Aqua 1999, 74, 10–20. [Google Scholar]
- Stenberg, C.; Støttrup, J.; Dahl, K.; Lundsteen, S.; Göke, C.; Andersen, O.N. Ecological Benefits from Restoring a Marine Cavernous Boulder Reef in Kattegat, Denmark; National Institute of Aquatic Resources, Danmarks Tekniske Universitet: Lyngby, Denmark, 2015. [Google Scholar]
- Kleiven, A.R.; Fernandez-Chacon, A.; Nordahl, J.-H.; Moland, E.; Espeland, S.H.; Knutsen, H.; Olsen, E.M. Harvest pressure on coastal Atlantic cod (Gadus morhua) from recreational fishing relative to commercial fishing assessed from tag-recovery data. PLoS ONE 2016, 11, e0149595. [Google Scholar]
- Kerwath, S.E.; Winker, H.; Götz, A.; Attwood, C.G. Marine protected area improves yield without disadvantaging fishers. Nat. Commun. 2013, 4, 2347. [Google Scholar] [CrossRef] [PubMed]
- Reubens, J.T.; Braeckman, U.; Vanaverbeke, J.; Van Colen, C.; Degraer, S.; Vincx, M. Aggregation at windmill artificial reefs: CPUE of Atlantic cod (Gadus morhua) and pouting (Trisopterus luscus) at different habitats in the Belgian part of the North Sea. Fish. Res. 2013, 139, 28–34. [Google Scholar] [CrossRef]
- Adams, G.D.; Flores, D.; Flores, O.G.; Aarestrup, K.; Svendsen, J.C. Spatial ecology of blue shark and shortfin mako in southern Peru: Local abundance, habitat preferences and implications for conservation. Endanger. Species Res. 2016, 31, 19–32. [Google Scholar] [CrossRef] [Green Version]
- Rhodes, N.; Wilms, T.; Baktoft, H.; Ramm, G.; Bertelsen, J.L.; Flávio, H.; Støttrup, J.G.; Kruse, B.M.; Svendsen, J.C. Comparing methodologies in marine habitat monitoring research: An assessment of species-habitat relationships as revealed by baited and unbaited remote underwater video systems. J. Exp. Mar. Biol. Ecol. 2020, 526, 151315. [Google Scholar] [CrossRef]
- Cappo, M.; Speare, P.; De’Ath, G. Comparison of baited remote underwater video stations (BRUVS) and prawn (shrimp) trawls for assessments of fish biodiversity in inter-reefal areas of the Great Barrier Reef Marine Park. J. Exp. Mar. Biol. Ecol. 2004, 302, 123–152. [Google Scholar] [CrossRef]
- Becker, A.; Taylor, M.D.; Lowry, M.B. Monitoring of reef associated and pelagic fish communities on Australia’s first purpose built offshore artificial reef. ICES J. Mar. Sci. 2016, 74, 277–285. [Google Scholar] [CrossRef]
- Ellis, D.; DeMartini, E. Evaluation of a video camera technique for indexing abundances of juvenile pink snapper, Pristipomoides filamentosus, and other Hawaiian insular shelf fishes. Fish. Bull. 1995, 93, 67–77. [Google Scholar]
- Priede, I.G.; Merrett, N.R. Estimation of abundance of abyssal demersal fishes; a comparison of data from trawls and baited cameras. J. Fish Biol. 1996, 49, 207–216. [Google Scholar] [CrossRef]
- Sogard, S.M.; Powell, G.V.N.; Holmquist, J.G. Epibenthic fish communities on Florida Bay banks: Relations with physical parameters and seagrass cover. Mar. Ecol. Prog. Ser. 1987, 40, 25–39. [Google Scholar] [CrossRef]
- Aller, E.A.; Gullström, M.; Eveleens Maarse, F.K.J.; Gren, M.; Nordlund, L.M.; Jiddawi, N.; Eklöf, J.S. Single and joint effects of regional- and local-scale variables on tropical seagrass fish assemblages. Mar. Biol. 2014, 161, 2395–2405. [Google Scholar] [CrossRef]
- Minello, T.J.; Able, K.W.; Weinstein, M.P. Salt marshes as nurseries for nekton: Testing hypotheses on density, growth and survival through meta-analysis. Mar. Ecol. Prog. Ser. 2003, 246, 39–59. [Google Scholar] [CrossRef]
- Oksanen, A.J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; Mcglinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan Community Ecology Package, R Package Version 2.5-7; 28 November 2020. Available online: https://mran.microsoft.com/package/vegan (accessed on 14 November 2021).
- Audebert, C.; Even, G.; Cian, A.; Loywick, A.; Merlin, S.; Viscogliosi, E.; Chabé, M. Colonization with the enteric protozoa Blastocystis is associated with increased diversity of human gut bacterial microbiota. Sci. Rep. 2016, 6, 25255. [Google Scholar] [CrossRef]
- Matteodo, M.; Ammann, K.; Verrecchia, E.P.; Vittoz, P. Snowbeds are more affected than other subalpine-alpine plant communities by climate change in the Swiss Alps. Ecol. Evol. 2016, 6, 6969–6982. [Google Scholar] [CrossRef]
- Wang, Y.; Naumann, U.; Wright, S.T.; Warton, D.I. mvabund–an R package for model-based analysis of multivariate abundance data. Methods Ecol. Evol. 2012, 3, 471–474. [Google Scholar] [CrossRef]
- Warton, D.I.; Wright, S.T.; Wang, Y. Distance-based multivariate analyses confound location and dispersion effects. Methods Ecol. Evol. 2012, 3, 89–101. [Google Scholar] [CrossRef]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Sequeira, A.M.M.; Mellin, C.; Lozano-Montes, H.M.; Meeuwig, J.J.; Vanderklift, M.A.; Haywood, M.D.E.; Babcock, R.C.; Caley, M.J. Challenges of transferring models of fish abundance between coral reefs. PeerJ 2018, 6, e4566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, C.L.; Summerside, M.; Johnson, P.T.J. An effective method for ecosystem-scale manipulation of bird abundance and species richness. Ecol. Evol. 2019, 9, 9748–9758. [Google Scholar] [CrossRef] [Green Version]
- Christ, A. Mixed effects models and extensions in Ecology with R. J. Stat. Softw. 2009, 32, 1–3. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2020. [Google Scholar]
- Katara, I.; Peden, W.J.; Bannister, H.; Ribeiro, J.; Fronkova, L.; Scougal, C.; Martinez, R.; Downie, A.; Sweeting, C.J. Conservation hotspots for fish habitats: A case study from English and Welsh waters. Reg. Stud. Mar. Sci. 2021, 44, 101745. [Google Scholar] [CrossRef]
- Bergström, L.; Karlsson, M.; Pihl, L. Comparison of Gill Nets and Fyke Nets for the Status Assessment of Coastal fish Communities; WATERS Report no. 2013:7; Havsmiljöinstitutet: Gothenburg, Sweden, 2013; Deliverable 3.4-2. [Google Scholar]
- KA, M.; Tomas, F.; Waldbusser, G. On the edge: Assessing fish habitat use across the boundary between Pacific oyster aquaculture and eelgrass in Willapa Bay, Washington, USA. Aquac. Environ. Interact. 2020, 12, 541–557. [Google Scholar]
- Bobsien, I.C.; Brendelberger, H. Comparison of an enclosure drop trap and a visual diving census technique to estimate fish populations in eelgrass habitats. Limnol. Oceanogr. Methods 2006, 4, 130–141. [Google Scholar] [CrossRef]
- Bergström, L.; Sundqvist, F.; Bergström, U. Effects of an offshore wind farm on temporal and spatial patterns in the demersal fish community. Mar. Ecol. Prog. Ser. 2013, 485, 199–210. [Google Scholar] [CrossRef] [Green Version]
- Thormar, J.; Hasler-Sheetal, H.; Baden, S.; Boström, C.; Clausen, K.K.; Krause-Jensen, D.; Olesen, B.; Rasmussen, J.R.; Svensson, C.J.; Holmer, M. Eelgrass (Zostera marina) food web structure in different environmental settings. PLoS ONE 2016, 11, e0146479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly, J.R.; Proctor, H.; Volpe, J.P. Intertidal community structure differs significantly between substrates dominated by native eelgrass (Zostera marina L.) and adjacent to the introduced oyster Crassostrea gigas (Thunberg) in British Columbia, Canada. Hydrobiologia 2008, 596, 57–66. [Google Scholar] [CrossRef]
- Schwartzbach, A.; Munk, P.; Sparholt, H.; Christoffersen, M. Marine mussel beds as attractive habitats for juvenile European eel (Anguilla anguilla); A study of bottom habitat and cavity size preferences. Estuar. Coast. Shelf Sci. 2020, 246, 107042. [Google Scholar] [CrossRef]
- Christoffersen, M.; Svendsen, J.C.; Kuhn, J.A.; Nielsen, A.; Martjanova, A.; Støttrup, J.G. Benthic habitat selection in juvenile European eel Anguilla anguilla: Implications for coastal habitat management and restoration. J. Fish Biol. 2018, 93, 996–999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helsinki Commission. HELCOM Red List of Baltic Sea Species in Danger of Becoming Extinct; Baltic Sea Environmental Proceedings No 140; Helsinki Commission: Helsinki, Finland, 2013; Volume 140, ISSN 0357-2994. [Google Scholar]
- Carl, H.; Møller, P.R. Fisk. In Moeslund, J.E. m.fl. (red.): Den Danske Rødliste 2019; Nationalt Center for Miljø og Energi: Roskilde, Denmark, 2019. [Google Scholar]
- Helsinki Commission. HELCOM Red List of Threatened and Declining Species of Lampreys and Fishes of the Baltic Sea; Baltic Sea Environmental Proceedings No 109; Helsinki Commission: Helsinki, Finland, 2006; ISSN 0357-294. [Google Scholar]
- Thedinga, J.F.; Johnson, S.W.; Neff, A.D. Diel differences in fish assemblages in nearshore eelgrass and kelp habitats in Prince William Sound, Alaska. Environ. Biol. Fishes 2011, 90, 61–70. [Google Scholar] [CrossRef]
- Pessanha, A.L.M.; Araújo, F.G.; De Azevedo, M.C.C.; Gomes, I.D. Diel and seasonal changes in the distribution of fish on a southeast Brazil sandy beach. Mar. Biol. 2003, 143, 1047–1055. [Google Scholar] [CrossRef]
- Magnhagen, C. Reproduction under predation risk in the sand goby, Pomatoschistus minutes, and the black goby, Gobius niger: The effect of age and longevity. Behav. Ecol. Sociobiol. 1990, 26, 331–335. [Google Scholar] [CrossRef]
- Fausch, K.D. Profitable stream positions for salmonids: Relating specific growth rate to net energy gain. Can. J. Zool. 1984, 62, 441–451. [Google Scholar] [CrossRef]
- Hamner, W.M.; Jones, M.S.; Carleton, J.H.; Hauri, I.R.; Williams, D.M. Zooplankton, planktivorous fish, and water currents on a windward reef face: Great Barrier Reef, Australia. Bull. Mar. Sci. 1988, 42, 459–479. [Google Scholar]
- Baktoft, H.; Jacobsen, L.; Skov, C.; Koed, A.; Jepsen, N.; Berg, S.; Boel, M.; Aarestrup, K.; Svendsen, J.C. Phenotypic variation in metabolism and morphology correlating with animal swimming activity in the wild: Relevance for the OCLTT (oxygen- and capacity-limitation of thermal tolerance), allocation and performance models. Conserv. Physiol. 2016, 4, cov055. [Google Scholar] [CrossRef] [Green Version]
- Kullander, S.O.; Nyman, L.; Jilg, K.; Delling, B. Nationalnyckeln till Sveriges flora och fauna. Strålfeniga fiskar. Actinopterygii; SLU Artdatabanken: Uppsala, Sweden, 2012; ISBN 978-91-88506-80-1. [Google Scholar]
- Carla, F.; Olsen, E.; Knutsen, H.; Albretsen, J.; Moland, E. Temperature-associated habitat selection in a cold-water marine fish. J. Anim. Ecol. 2015, 85, 628–637. [Google Scholar] [CrossRef] [Green Version]
- International Council for the Exploration of the Sea. ICES Report of the ICES Advisory Committee 2012; ICES Advise 2012; International Council for the Exploration of the Sea: Copenhagen, Denmark, 2012; p. 389. [Google Scholar]
- Svendsen, J.C.; Genz, J.; Anderson, W.G.; Stol, J.A.; Watkinson, D.A.; Enders, E.C. Evidence of circadian rhythm, oxygen regulation capacity, metabolic repeatability and positive correlations between forced and spontaneous maximal metabolic rates in lake sturgeon Acipenser fulvescens. PLoS ONE 2014, 9, e94693. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Family | Fish Species Latin Name | Common Name | CPUE Mussel | CPUE Seagrass | p | Dev |
|---|---|---|---|---|---|---|
| Anguillidae | Anguilla anguilla | European eel | 0.3 | 2.429 | <0.001 *** | 12.15 |
| Labridae | Ctenolabrus rupestris | Goldsinny wrasse | 0.067 | 0 | 0.18 | 2.64 |
| Gadidae | Gadus morhua | Atlantic cod | 0.067 | 0 | 0.14 | 2.64 |
| Gobiidae | Gobius niger | Black goby | 1.2 | 0.429 | 0.01 * | 6.45 |
| Gobiidae | Gobiusculus flavescens | Two-spotted goby | 1.333 | 1.714 | 0.61 | 0.28 |
| Cottidae | Myoxocephalus scorpius | Shorthorn sculpin | 0 | 0.071 | 0.13 | 2.91 |
| Pleuronectidae | Platichthys flesus | European flounder | 0.067 | 0.036 | 0.68 | 0.27 |
| Pleuronectidae | Pleuronectes platessa | European plaice | 0.033 | 0.036 | 0.86 | 0 |
| Gasterosteidae | Pungitius pungitius | Ninespine stickleback | 0.033 | 0 | 0.47 | 1.32 |
| Gasterosteidae | Spinachia spinachia | Sea stickleback | 0.133 | 0.607 | 0.11 | 2.96 |
| Labridae | Symphodus melops | Corkwing wrasse | 0.033 | 0.286 | 0.06 | 4.1 |
| Syngnathidae | Syngnathus typhle | Broadnosed pipefish | 0.033 | 0 | 0.44 | 1.32 |
| Cottidae | Taurulus bubalis | Longspined bullhead | 0.067 | 0.036 | 0.65 | 0.27 |
| Zoarcidae | Zoarces viviparus | Eelpout | 1.6 | 2.607 | 0.02 * | 5.79 |
| Family | Fish Species | Common Name | Mean Abundance in High Current Mussel Reef | CI for High Current | Mean Abundance in Low Current Mussel Reef | CI for Low Current | p |
|---|---|---|---|---|---|---|---|
| Gadidae | Gadus morhua | Atlantic cod | 0.000 | 0 | 0.621 | 0.446–0.864 | <0.001 *** |
| Gasterosteidae | Gasterosteus aculeatus | Three-spined stickleback | 0.000 | 0–0.038 | 0.000 | 0–0.004 | 0.485 |
| Gobiidae | Gobius niger | Black goby | 0.379 | 0.154–0.931 | 0.023 | 0.006–0.087 | <0.001 *** |
| Gobiidae | Gobiusculus flavescens | Two-spotted goby | 10.135 | 3.112–33.002 | 21.221 | 6.107–73.736 | 0.3876 |
| Gobiidae | Other gobies | 0.084 | 0.012–0.609 | 0.283 | 0.022–3.607 | 0.2511 | |
| Labridae | Ctenolabrus rupestris | Goldsinny wrasse | 0.017 | 0.003–0.086 | 0.515 | 0.157–1.694 | <0.001 *** |
| Labridae | Other wrasses | 0.000 | 0–0.006 | 0.000 | 0–0.005 | 0.7568 | |
| Pleuronectidae | Flatfish | 0.010 | 0.001–0.166 | 0.041 | 0.002–0.945 | 0.0745 | |
| Gasterosteidae | Spinachia spinachia | Sea stickleback | 0.058 | 0.022–0.149 | 0.006 | 0.001–0.048 | 0.043 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Orfanidis, G.A.; Touloumis, K.; Stenberg, C.; Mariani, P.; Støttrup, J.G.; Svendsen, J.C. Fish Assemblages in Seagrass (Zostera marina L.) Meadows and Mussel Reefs (Mytilus edulis): Implications for Coastal Fisheries, Restoration and Marine Spatial Planning. Water 2021, 13, 3268. https://doi.org/10.3390/w13223268
Orfanidis GA, Touloumis K, Stenberg C, Mariani P, Støttrup JG, Svendsen JC. Fish Assemblages in Seagrass (Zostera marina L.) Meadows and Mussel Reefs (Mytilus edulis): Implications for Coastal Fisheries, Restoration and Marine Spatial Planning. Water. 2021; 13(22):3268. https://doi.org/10.3390/w13223268
Chicago/Turabian StyleOrfanidis, Georgios A., Konstantinos Touloumis, Claus Stenberg, Patrizio Mariani, Josianne Gatt Støttrup, and Jon C. Svendsen. 2021. "Fish Assemblages in Seagrass (Zostera marina L.) Meadows and Mussel Reefs (Mytilus edulis): Implications for Coastal Fisheries, Restoration and Marine Spatial Planning" Water 13, no. 22: 3268. https://doi.org/10.3390/w13223268