
Effects of Imazapyr on Spartina alterniflora and Soil Bacterial Communities in a Mangrove Wetland

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
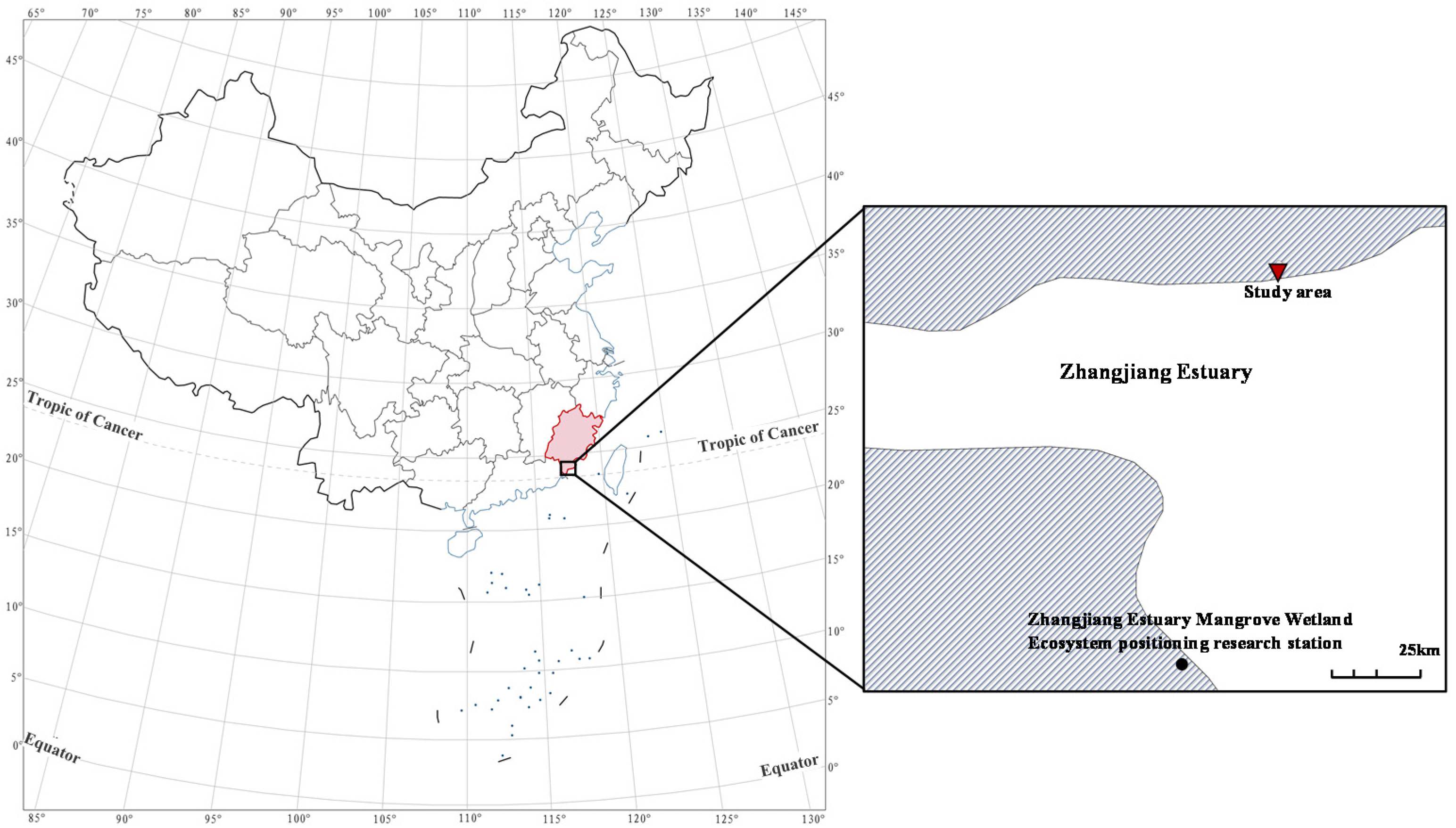
2.1. Overview of the Study Area
2.2. Site Setting and Experimental Methods
2.3. Determination Methods of Residues and Soil Physicochemical Properties
2.4. Soil DNA Extraction, PCR Amplification, and Sequencing
2.5. Data Statistics and Analysis
3. Results and Discussion
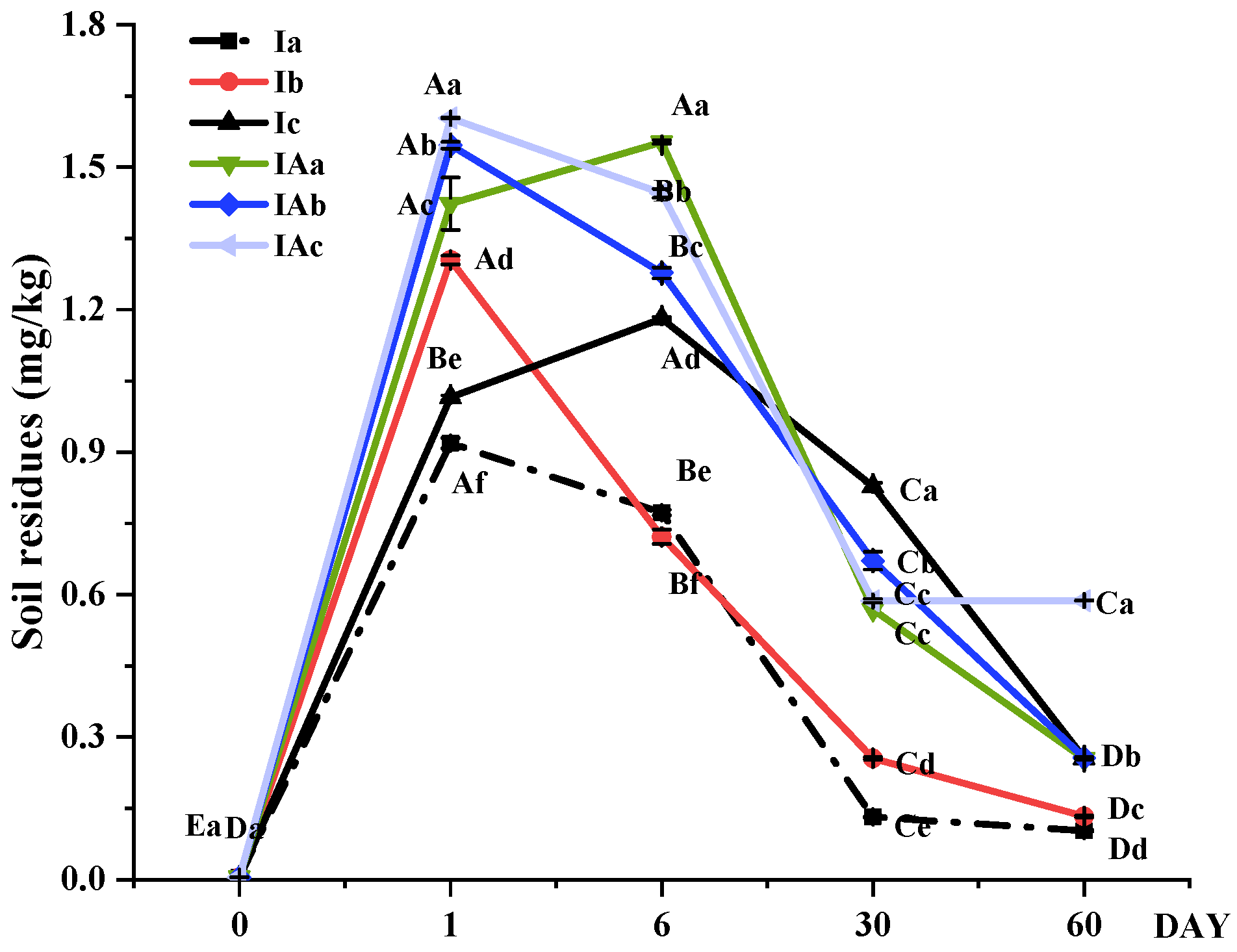
3.1. Effects of Imazapyr on Plants
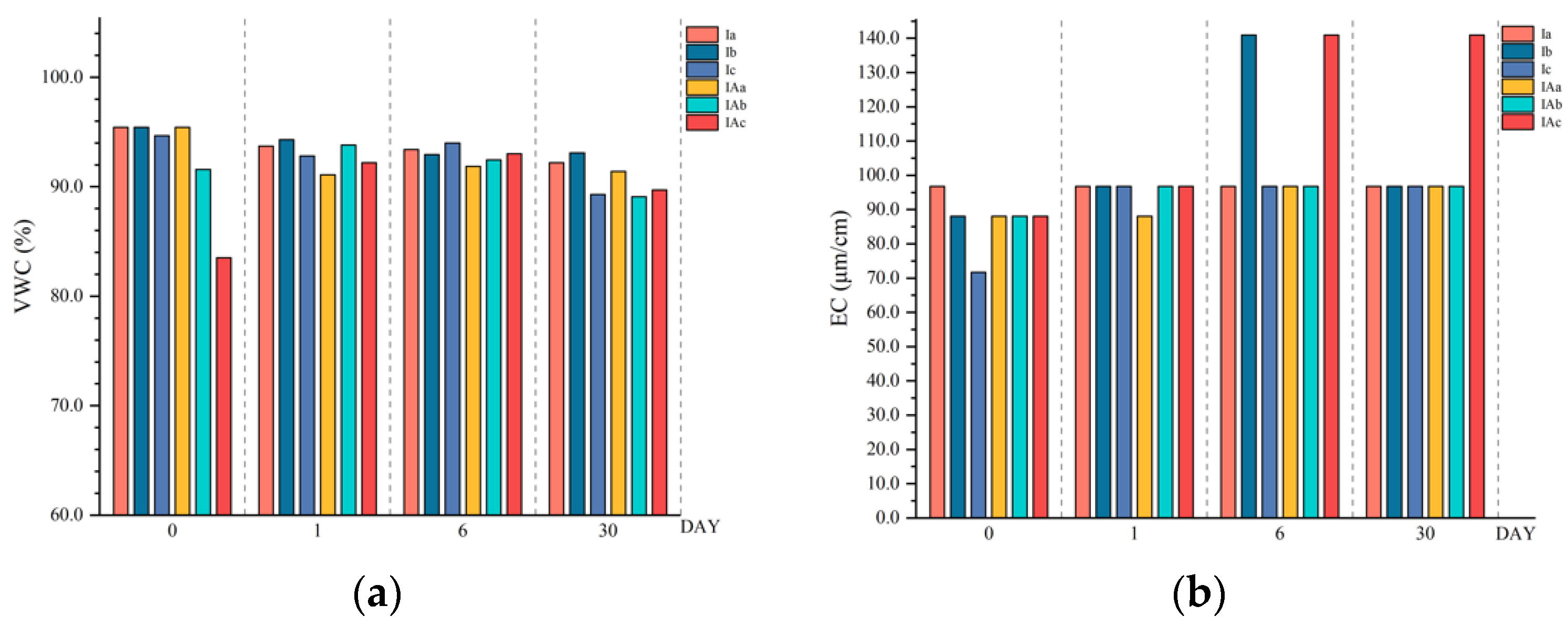
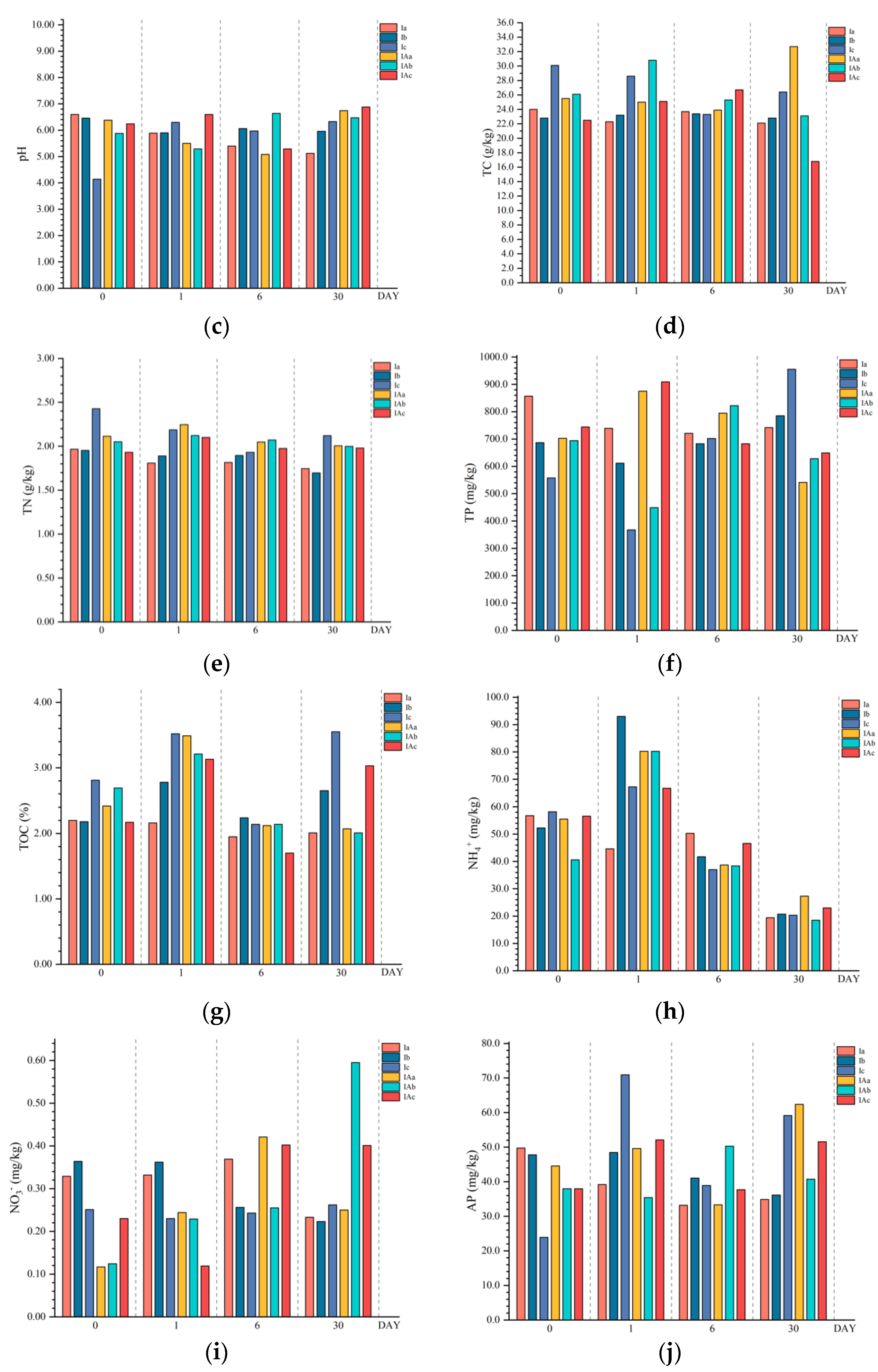
3.2. Effects of Imazapyr and Its Additives on the Physicochemical Properties of the Topsoil
3.3. Effects of Imazapyr and Its Additives on Soil Bacterial Abundance and Diversity
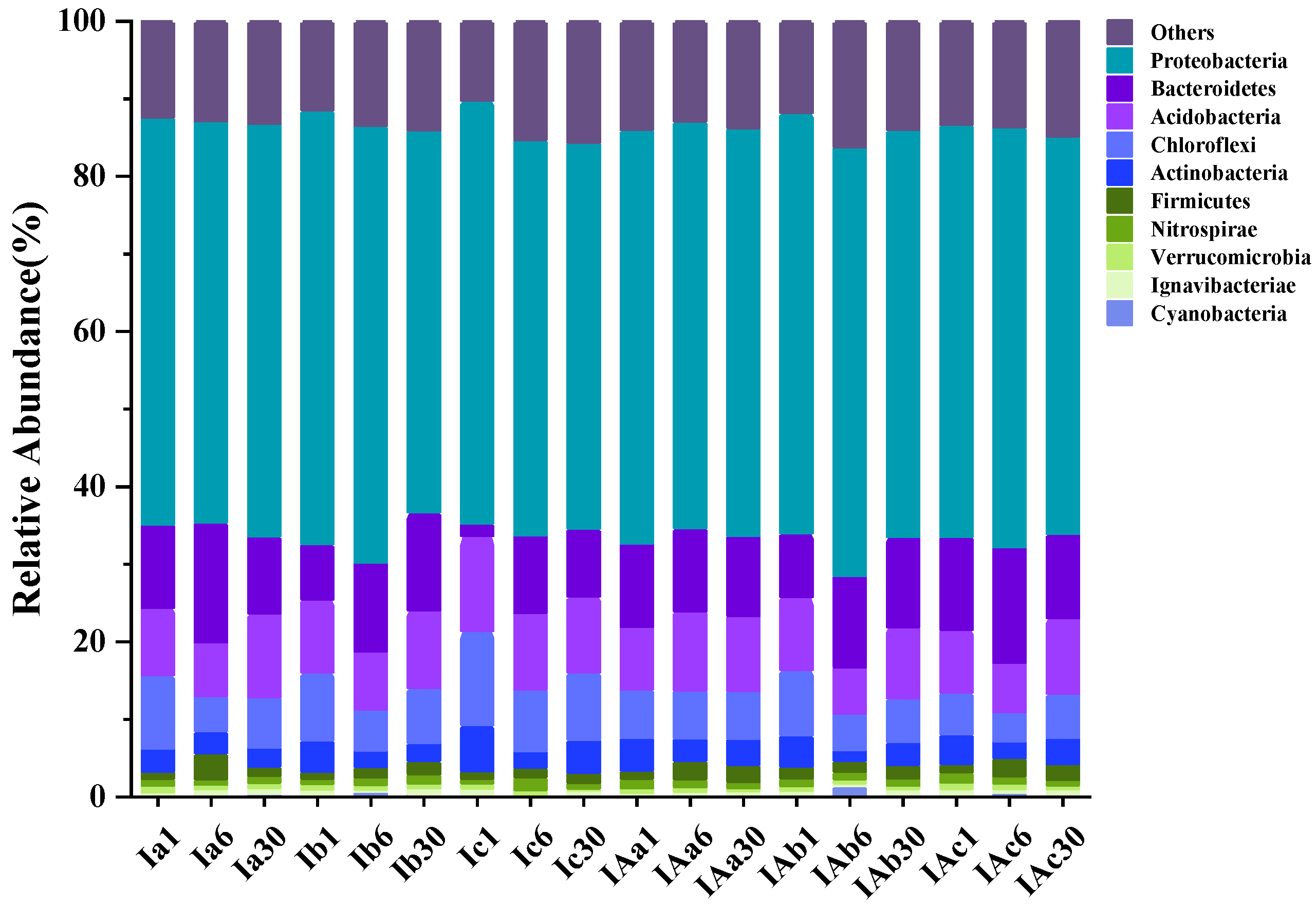
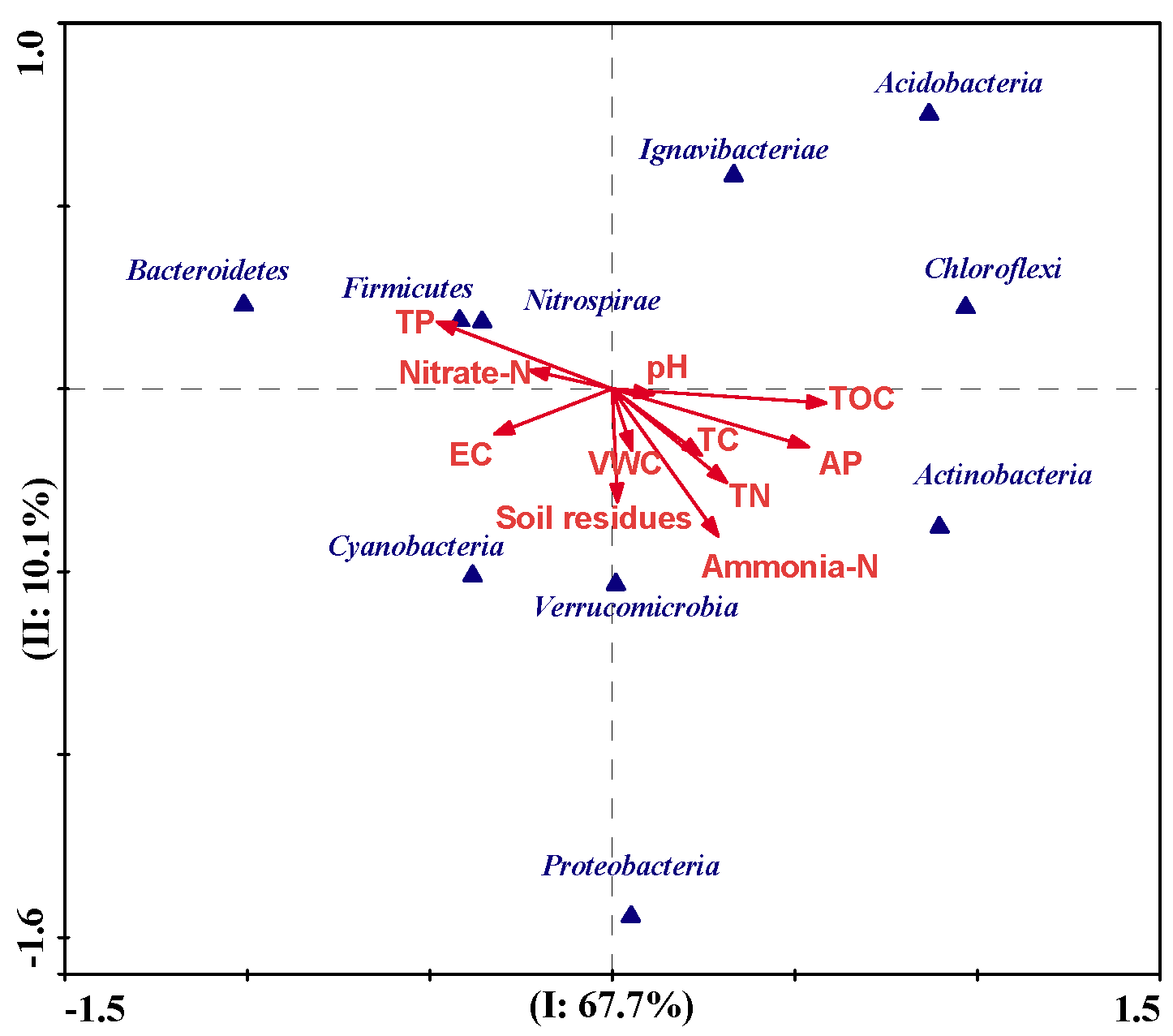
3.4. Effects of Imazapyr and Its Additives on the Structure of Soil Bacterial Communities
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, B. Spartina alterniflora invasions in the Yangtze River estuary, China: An overview of current status and ecosystem effects. Ecol. Eng. 2009, 35, 511–520. [Google Scholar] [CrossRef]
- Xie, W.; Gao, S. Invasive Spartina alterniflora-induced factors affecting epibenthos distribution in coastal salt marsh, China. Acta Oceanol. Sin. 2013, 32, 81–88. [Google Scholar] [CrossRef]
- Qi, X.; Liu, H.; Lin, Z.; Liu, X.; Gong, H. Impacts of Age and Expansion Direction of Invasive Spartina alterniflora on Soil Organic Carbon Dynamics in Coastal Salt Marshes Along Eastern China. Estuaries Coasts 2019, 42, 1858–1867. [Google Scholar]
- Schmidt, R.; Mitchell, J.; Scow, K. Cover cropping and no-till increase diversity and symbiotroph:saprotroph ratios of soil fungal communities. Soil Biol. Biochem. 2018, 129, 99–109. [Google Scholar] [CrossRef] [Green Version]
- An, S.Q.; Gu, B.H.; Zhou, C.; Wang, Z.S.; Liu, Y.H. Spartina Invasion in China: Implications for Invasive Species Management and Future Research. Weed Res. 2007, 47, 183–191. [Google Scholar] [CrossRef]
- Zhang, R.S.; Shen, Y.M.; Lu, L.Y.; Yan, S.G.; Wang, Y.H.; Li, J.L.; Zhang, Z.L. Formation of Spartina alterniflora salt marshes on the coast of Jiangsu Province, China. Ecol. Eng. 2004, 23, 95–105. [Google Scholar] [CrossRef]
- Chen, Z.Y.; Li, B.; Chen, J.K. Ecological consequences and management of Spartina spp. Invasions in coastal ecosystems. Biodivers. Sci. 2004, 12, 280–289. [Google Scholar]
- Zhang, Y.; Huang, G.; Wang, W.; Lin, C.G. Interactions between mangroves and exotic Spartina in an anthropogenically disturbed estuary in southern China. Ecology 2012, 93, 588–597. [Google Scholar] [CrossRef] [Green Version]
- Grosholz, E.D.; Levin, L.A.; Tyler, A.C.; Neira, C. Changes in Community Structure and Ecosystem Function Following Spartina Alterniflora Invasion of Pacific Estuaries. In Human Impacts on Salt Marshes: A Global Perspective; University of California Press: Berkeley, CA, USA, 2009; pp. 23–40. [Google Scholar]
- Page, H.M.; Lastra, M.; Rodil, I.F.; Briones, M.; Garrido, J. Effects of non-native Spartina patens on plant and sediment organic matter carbon incorporation into the local invertebrate community. Biol. Invasions 2010, 12, 3825–3838. [Google Scholar] [CrossRef] [Green Version]
- Tang, L.; Yang, G.; Wang, C.H.; Zhao, B.; Li, B. A plant invader declines through its modification to habitats: A case study of a 16-year chronosequence of Spartina alterniflora invasion in a salt marsh. Ecol. Eng. 2012, 49, 181–185. [Google Scholar] [CrossRef]
- Donato, D.C.; Kauffman, J.B.; Murdiyarso, D.; Kurnianto, S.; Stidham, M.; Kanninen, M. Mangroves among the most carbon-rich forests in the tropics. Nat. Geosci. 2011, 4, 293–297. [Google Scholar] [CrossRef]
- Riddin, T.; Wyk, E.V.; Adams, J. The rise and fall of an invasive estuarine grass. S. Afr. J. Bot. 2016, 107, S0254629916336523. [Google Scholar] [CrossRef]
- Knott, C.A.; Webster, E.P.; Nabukalu, P. Control of smooth cordgrass (Spartina alterniflora) seedlings with four herbicides. J. Aquat. Plant Manag. 2013, 51, 132–135. [Google Scholar]
- Yuan, L.; Zhang, L.; Xiao, D.; Huang, H. The application of cutting plus waterlogging to control Spartina alterniflora on saltmarshes in the Yangtze Estuary, China. Estuar. Coast. Shelf Sci. 2011, 92, 103–110. [Google Scholar] [CrossRef]
- Hu, Z.J.; Ge, Z.M.; Ma, Q.; Zhang, Z.T.; Tang, C.D.; Cao, H.B.; Zhang, T.Y.; Li, B.; Zhang, L.Q. Revegetation of a native species in a newly formed tidal marsh under varying hydrological conditions and planting densities in the Yangtze Estuary. Ecol. Eng. 2015, 83, 354–363. [Google Scholar] [CrossRef]
- Strong, D.R.; Ayres, D.A. Control and consequences of Spartina spp. invasions with focus upon San Francisco Bay. Biol. Invasions 2016, 18, 2237–2246. [Google Scholar] [CrossRef] [Green Version]
- Jie, W.; Gu, Y.; You, H. Research Progress on the Control Measures and Utilization Status of Spartina alterniflora. Genom. Appl. Biol. 2017, 36, 3152–3156. [Google Scholar]
- Zhao, Z.-Y.; Xu, Y.; Yuan, L.; Li, W.; Zhu, X.-J.; Zhang, L.-Q. Emergency control of Spartina alterniflora re-invasion with a chemical method in Chongming Dongtan, China. Water Sci. Eng. 2020, 13, 24–33. [Google Scholar] [CrossRef]
- Patten, K. Persistence and non-target impact of imazapyr associated with smooth cordgrass control in an estuary. J. Aquat. Plant Manag. 2003, 41, 1–6. [Google Scholar]
- Patten, K. Smooth Cordgrass (Spartina alterniflora) Control with Imazapyr1. Weed Technol. 2002, 16, 826–832. [Google Scholar] [CrossRef]
- Kaiser, K.; Wemheuer, B.; Korolkow, V.; Wemheuer, F.; Nacke, H.; Schöning, I.; Schrumpf, M.; Daniel, R. Driving forces of soil bacterial community structure, diversity, and function in temperate grasslands and forests. Sci. Rep. 2016, 6, 33696. [Google Scholar] [CrossRef] [Green Version]
- Nottingham, A.T.; Noah, F.; Turner, B.L.; Jeanette, W.; Ostle, N.J.; Mcnamara, N.P.; Bardgett, R.D.; Leff, J.W.; Norma, S.; Miles, S. Microbes follow Humboldt: Temperature drives plant and soil microbial diversity patterns from the Amazon to the Andes. Ecology 2018, 99, 2455–2466. [Google Scholar] [CrossRef] [Green Version]
- Bonfante, P.; Anca, I.A. Plants, Mycorrhizal Fungi, and Bacteria: A Network of Interactions. Annu. Rev. Microbiol. 2009, 63, 363–383. [Google Scholar] [CrossRef] [Green Version]
- Kuzyakov, Y.; Blagodatskaya, E. Microbial hotspots and hot moments in soil: Concept & review. Soil Biol. Biochem. 2015, 83, 184–199. [Google Scholar]
- Mangeks, G. Behaviour of the imidazolinone herbicides in soil—Areview of the literature. In The Imidazolinone Herbicides; Shaner, D.L., O’Conner, S.L., Eds.; CRC Press: Boca Raton, FL, USA, 1991. [Google Scholar]
- Lonarevi, I.; Pajin, B.; Petrovi, J.; Zari, D.; Omorjan, R. The impact of sunflower and rapeseed lecithin on the rheological properties of spreadable cocoa cream. J. Food Eng. 2016, 171, 67–77. [Google Scholar] [CrossRef]
- Bedos, C.; Cellier, P.; Calvet, R.; Barriuso, E.; Gabrielle, B. Mass transfer of pesticides into the atmosphere by volatilization from soils and plants: Overview. Agronomie 2002, 22, 21–33. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, W.; Sheng, Y.; Chen, J.; Qu, M. Degradation characteristics of imazapyr in farmland soil. Acta Agric. Shanghai 2014, 30, 79–81. [Google Scholar]
- Luo, G.; Xue, C.; Jiang, Q.; Xiao, Y.; Zhang, F.; Guo, S.; Shen, Q.; Ling, N. Soil Carbon, Nitrogen, and Phosphorus Cycling Microbial Populations and Their Resistance to Global Change Depend on Soil C:N:P Stoichiometry. mSystems 2020, 5, e00162-20. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.D.; Wang, H.L.; Fan, D.F. Degradation and Metabolism of Imazapyr by Microorganisms in Soils. Res. Environ. Sci. 2004, 17, 42–45. [Google Scholar]
- Debra, E. Reregistration Eligibility Decision (RED) Document for Imazapyr; United States Environmental Protection Agency: Washington, DC, USA, 2006; Volume 6. [Google Scholar]
- Patten, K. Evaluating Imazapyr in Aquatic Environments, Searching for Ways to Stem the Tide of Aquatic Weeds. Agrichem. Environ. News 2003, 205, 1–7. [Google Scholar]
- Olofson, P.; Pless, P.; Env, D. Use of Imazapyr Herbicide to Control Invasive Cordgrass (Spartina spp.) in the San Francisco Estuary; Leson & Associates: Berkeley, CA, USA, 2005; Volume 4. [Google Scholar]
- Liu, F.; Mo, X.; Kong, W.; Song, Y. Soil bacterial diversity, structure, and function of Suaeda salsa in rhizosphere and non-rhizosphere soils in various habitats in the Yellow River Delta, China. Sci. Total Environ. 2020, 740, 140144. [Google Scholar] [CrossRef]
- Mimmo, T.; Del Buono, D.; Terzano, R.; Tomasi, N.; Vigani, G.; Crecchio, C.; Pinton, R.; Zocchi, G.; Cesco, S. Rhizospheric organic compounds in the soil-microorganism-plant system: Their role in iron availability. Eur. J. Soil Sci. 2014, 65, 629–642. [Google Scholar] [CrossRef]
- Cao, M.; Cui, L.; Sun, H.; Zhang, X.; Zheng, X.; Jiang, J. Effects of Spartina alterniflora Invasion on Soil Microbial Community Structure and Ecological Functions. Microorganisms 2021, 9, 138. [Google Scholar] [CrossRef]
- Ma, J.C.; Ibekwe, A.M.; Yang, C.H.; Crowley, D.E. Bacterial diversity and composition in major fresh produce growing soils affected by physiochemical properties and geographic locations. Sci. Total Environ. 2016, 563–564, 199–209. [Google Scholar] [CrossRef] [Green Version]
- Yan, R.; Feng, W. Effect of vegetation on soil bacteria and their potential functions for ecological restoration in the Hulun Buir Sandy Land, China. J. Arid Land 2020, 12, 473–494. [Google Scholar] [CrossRef]
- Jing, C.L.; Xu, Z.C.; Zou, P.; Tang, Q.; Li, Y.Q.; You, X.W.; Zhang, C.S. Coastal halophytes alter properties and microbial community structure of the saline soils in the Yellow River Delta, China. Appl. Soil Ecol. 2019, 134, 1–7. [Google Scholar] [CrossRef]
- Tan, L.; Gu, S.; Li, S.; Ren, Z.; Deng, Y.; Gong, Z.; Xiao, W.; Hu, Q. Responses of Microbial Communities and Interaction Networks to Different Management Practices in Tea Plantation Soils. Sustainability 2019, 11, 4428. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.M.; Wang, N.; Hu, Y.B.; Sun, G.Y. Changes in soil physicochemical properties and soil bacterial community in mulberry (Morus alba L.)/alfalfa (Medicago sativa L.) intercropping system. Microbiologyopen 2018, 7, e00555. [Google Scholar] [CrossRef]
- Liu, Y.; Jiang, L.; Shao, Z. Advances in sulfur-oxidizing bacterial taxa and their sulfur oxidation pathways. Acta Microbiol. Sin. 2018, 58, 191–201. [Google Scholar]
- Yu, H.; Kashima, H.; Regan, J.M.; Hussain, A.; Elbeshbishy, E.; Lee, H.-S. Kinetic study on anaerobic oxidation of methane coupled to denitrification. Enzym. Microb. Technol. 2017, 104, 47–55. [Google Scholar] [CrossRef]
- Bottrill, D.; Ogbourne, S.M.; Citerne, N.; Smith, T.; Farrar, M.B.; Hu, H.-W.; Omidvar, N.; Wang, J.; Burton, J.; Kämper, W.; et al. Short-term application of mulch, roundup and organic herbicides did not affect soil microbial biomass or bacterial and fungal diversity. Chemosphere 2020, 244, 125436. [Google Scholar] [CrossRef] [PubMed]
- García-Delgado, C.; Barba-Vicente, V.; Marín-Benito, J.M.; Mariano Igual, J.; Sánchez-Martín, M.J.; Sonia Rodríguez-Cruz, M. Influence of different agricultural management practices on soil microbial community over dissipation time of two herbicides. Sci. Total Environ. 2019, 646, 1478–1488. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.X.; Laborda, P.; Xie, X.L.; Zhou, R.; Chen, Y.; Li, T.; Pu, Z.H.; Wang, Y.L.; Deng, Z.F. Spartina alterniflora invasion alters soil microbial metabolism in coastal wetland of China. Estuar. Coast. Shelf Sci. 2020, 245, 106982. [Google Scholar] [CrossRef]
- Zeleke, J.; Sheng, Q.; Wang, J.G.; Huang, M.Y.; Xia, F.; Wu, J.H.; Quan, Z.X. Effects of Spartina alterniflora invasion on the communities of methanogens and sulfate-reducing bacteria in estuarine marsh sediments. Front. Microbiol. 2013, 4, 243. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tested Herbicide | Active Ingredient Concentration | Absorption/Action Site | Production Company |
|---|---|---|---|
| 25% imazapyr water agent | 22.62% | Leaves | Beijing Top Green Ecological Technology Co., Ltd. |
| AGE 809 (Lecithin) | 50% | Leaves | Beijing Top Green Ecological Technology Co., Ltd. |
| Sample number | Treatment method | ||
| Ia | 25% imazapyr 3035 mL/acre | ||
| Ib | 25% imazapyr 6070 mL/acre | ||
| Ic | 25% imazapyr 9105 mL/acre | ||
| IAa | 25% imazapyr 6070 mL/acre + AGE 809 1299 mL/acre | ||
| IAb | 25% imazapyr 6070 mL/acre + AGE 809 2604 mL/acre | ||
| IAc | 25% imazapyr 6070 mL/acre + AGE 809 5202 mL/acre | ||
| VWC | EC | PH | TC | TN | TP | TOC | AP | NH4+-N | NO3−-N | |
|---|---|---|---|---|---|---|---|---|---|---|
| Plant density | −0.222 | −0.609 ** | −0.158 | 0.634 ** | 0.209 | 0.274 | 0.716 ** | 0.118 | −0.182 | 0.471 |
| residues | −0.071 | 0.568 * | 0.27 | −0.363 | −0.251 | −0.12 | −0.299 | 0.171 | −0.343 | −0.028 |
| Tag Number | OTU Number | Chao1 | ACE | Shannon | Simpson | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ia1 | 43,745 ± 3597.43bc | 4395.67 ± 141.21fg | 6636.99 ± 229.62e | 8051.70 ± 22,770cd | 6.80 ± 0.026g | 0.0035 ± 0.00009d | ||||||
| Ia6 | 38,803.33 ± 1069.46c | B | 4784.67 ± 47.26de | AB | 7534.57 ± 170.51bc | A | 9228.64 ± 188.63ab | AB | 7.03 ± 0.041d | A | 0.0027 ± 0.00016g | BC |
| Ia30 | 42,417 ± 2856.91c | 4749 ± 85.06de | 7267.80 ± 133.38c | 8440.74 ± 958.35bc | 6.95 ± 0.015ef | 0.0031 ± 0.00012ef | ||||||
| Ib1 | 47,514 ± 3408.79b | 4480 ± 39.05f | 6560.89 ± 164.63e | 7595.95 ± 917.97d | 6.73 ± 0.026h | 0.0042 ± 0.00008b | ||||||
| Ib6 | 39,802.67 ± 2017.67c | B | 5172.67 ± 41.96b | AB | 7791.87 ± 143.42a | AB | 8988.58 ± 974.66bc | B | 7.15 ± 0.012b | AB | 0.0025 ± 0.00002h | B |
| Ib30 | 39,864.67 ± 1915.39c | 4252.33 ± 31.39g | 6396.81 ± 204.36e | 7664.25 ± 272.59cd | 6.84 ± 0.023g | 0.0036 ± 0.00012d | ||||||
| Ic1 | 54,614 ± 2091.4a | 2971.67 ± 34.06h | 3902.45 ± 127.42f | 4048.71 ± 153.72f | 6.13 ± 0.014i | 0.0075 ± 0.00012a | ||||||
| Ic6 | 40,877 ± 778.77c | A | 4964 ± 32.19cd | B | 7597.27 ± 148.29b | B | 9102.58 ± 85.52b | C | 7.05 ± 0.015d | B | 0.0028 ± 0.00006fg | A |
| Ic30 | 47,561.33 ± 1875.13b | 4822.67 ± 109.18d | 7168.43 ± 122.28cd | 7203.39 ± 155.02d | 6.96 ± 0.020ef | 0.0033 ± 0.00013e | ||||||
| IAa1 | 43,478 ± 5648.47bc | 4646.67 ± 102.72e | 7150.67 ± 141.16cd | 8569.66 ± 93.28bc | 6.98 ± 0.023e | 0.0027 ± 0.00016g | ||||||
| IAa6 | 42,171 ± 1547.36c | B | 4806 ± 167.52de | AB | 7410.37 ± 274.34bc | A | 8989.56 ± 423.11bc | AB | 6.98 ± 0.025e | A | 0.0028 ± 0.00006fg | BC |
| IAa30 | 41,204.67 ± 1845.65c | 4537.67 ± 37.17ef | 6943.70 ± 51.28d | 8416.68 ± 204.54bc | 6.93 ± 0.037f | 0.0031 ± 0.00027ef | ||||||
| IAb1 | 51,154.67 ± 3123.97ab | 4601.67 ± 49.52ef | 6525.03 ± 229.82e | 6731.94 ± 203.99e | 6.80 ± 0.019g | 0.0039 ± 0.00016c | ||||||
| IAb6 | 38,562.67 ± 1270.07c | B | 5542.67 ± 15.17a | A | 8030.69 ± 85.36a | A | 8370.64 ± 126.30c | B | 7.30 ± 0.027a | A | 0.0022 ± 0.00009i | BC |
| IAb30 | 40,284 ± 2776.52c | 4659.67 ± 152.68e | 7292.71 ± 116.20c | 8820.81 ± 279.42bc | 6.96 ± 0.041ef | 0.0029 ± 0.00014f | ||||||
| IAc1 | 40,916.33 ± 120.38c | 5042.33 ± 120.38c | 7656.86 ± 145.03b | 9470.94 ± 140.69ab | 7.11 ± 0.026c | 0.0024 ± 0.00011hi | ||||||
| IAc6 | 39,968.67 ± 1809.1c | B | 5241.33 ± 143.20b | A | 8003.24 ± 239.65a | A | 9835.43 ± 317.38a | A | 7.19 ± 0.050b | A | 0.0022 ± 0.00015i | C |
| IAc30 | 43,549.33 ± 3172.85bc | 4955.67 ± 131.96cd | 7381.83 ± 271.23bc | 8824.52 ± 208.37bc | 7.04 ± 0.007d | 0.0027 ± 0.00001gh | ||||||
| Soil Factors | Importance Ranking | Explanatory Quantity | P | F |
|---|---|---|---|---|
| TOC | 1 | 47.6 | 0.002 | 14.546 |
| TP | 2 | 27.7 | 0.024 | 6.141 |
| AP | 3 | 22.8 | 0.03 | 4.721 |
| NH4+-N | 4 | 20.9 | 0.042 | 4.227 |
| TN | 5 | 18.3 | 0.048 | 3.583 |
| TC | 6 | 11.5 | 0.19 | 2.078 |
| EC | 7 | 5.2 | 0.352 | 0.881 |
| NO3−-N | 8 | 3.8 | 0.47 | 0.626 |
| Soil residues | 9 | 3.6 | 0.496 | 0.605 |
| VWC | 10 | 0.6 | 0.0818 | 0.092 |
| pH | 11 | 0.3 | 0.091 | 0.04 |
| Soil Factors | Importance Ranking | Explanatory Quantity | P | F |
|---|---|---|---|---|
| TOC | 1 | 29 | 0.004 | 6.525 |
| AP | 2 | 24.9 | 0.024 | 5.299 |
| TP | 3 | 20.7 | 0.026 | 4.166 |
| NH4+-N | 4 | 12.6 | 0.108 | 2.296 |
| TN | 5 | 10.9 | 0.128 | 1.948 |
| EC | 6 | 9.4 | 0.172 | 1.652 |
| TC | 7 | 6 | 0.314 | 1.028 |
| NO3−-N | 9 | 4.8 | 0.396 | 0.807 |
| Soil residues | 8 | 3.4 | 0.582 | 0.571 |
| VWC | 10 | 2.7 | 0.66 | 0.446 |
| pH | 11 | 1.4 | 0.872 | 0.232 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mo, X.; Dong, P.; Xie, L.; Xiu, Y.; Wang, Y.; Wu, B.; Liu, J.; Song, X.; Zhang, M.; Zhang, Z. Effects of Imazapyr on Spartina alterniflora and Soil Bacterial Communities in a Mangrove Wetland. Water 2021, 13, 3277. https://doi.org/10.3390/w13223277
Mo X, Dong P, Xie L, Xiu Y, Wang Y, Wu B, Liu J, Song X, Zhang M, Zhang Z. Effects of Imazapyr on Spartina alterniflora and Soil Bacterial Communities in a Mangrove Wetland. Water. 2021; 13(22):3277. https://doi.org/10.3390/w13223277
Chicago/Turabian StyleMo, Xue, Panpan Dong, Lumeng Xie, Yujiao Xiu, Yanqi Wang, Bo Wu, Jiakai Liu, Xiuhua Song, Mingxiang Zhang, and Zhenming Zhang. 2021. "Effects of Imazapyr on Spartina alterniflora and Soil Bacterial Communities in a Mangrove Wetland" Water 13, no. 22: 3277. https://doi.org/10.3390/w13223277