
Passive Treatment for Acid Mine Drainage Partially Restores Microbial Community Structure in Different Stream Habitats

1
Department of Biology, Grand Valley State University, Allendale, MI 49401, USA
2
Department of Biology, Slippery Rock University, Slippery Rock, PA 16057, USA
*
Author to whom correspondence should be addressed.
Water 2021, 13(22), 3300; https://doi.org/10.3390/w13223300
Submission received: 13 October 2021
/
Revised: 17 November 2021
/
Accepted: 18 November 2021
/
Published: 22 November 2021
(This article belongs to the Section Biodiversity and Functionality of Aquatic Ecosystems)
Abstract
:The assessment of the degree to which biological communities in streams impaired by acid mine drainage (AMD) are restored by passive treatment has focused primarily on eukaryotic-cell organisms and microbial processes. The responses of microbial community structure to passive treatment have received much less attention, even though functional processes such as nutrient cycling and organic matter decomposition depend on taxonomic composition. Our objective was to determine the degree to which passive treatment restored microbial communities in three types of habitats: aqueous, leaf, and sediment. To assess their recovery, we compared the community composition in these habitats based on 16S rRNA gene sequencing at three different stream sites: an untreated AMD site (U), a remediated site below AMD passive treatment (T), and an unimpaired reference site (R). The acidity, conductivity, and soluble metal concentrations at T were found to be elevated compared to R, but generally 1–2 orders of magnitude less than at U. Microbial community composition was found to be synergistically affected by habitat type and AMD impact, with the similarity among communities in the three habitats increasing with the severity of the AMD. Sediment- and leaf-associated microbial communities at U were characterized by taxa that are tolerant to severe AMD. The absence of the nitrogen oxidizing bacterium Nitrospira in sediment communities at T and U was found to correspond to higher NH4+ concentrations compared to R, possibly because of the presence of iron oxyhydroxide precipitate. In contrast, the microbial composition was found to be similar between the T and R sites for both aqueous and leaf communities, indicating that passive treatment was more able to restore these communities to the reference condition than sediment communities. The remediation of AMD streams should consider the habitat-specific responses of microbial community composition and be guided by future studies that empirically couple changes in taxonomic composition to measured functional processes.
1. Introduction
While extreme habitats pose many challenges for lifeforms, some microbes can thrive under such conditions [1]. One challenge that affects microbial communities is acid mine drainage (AMD) [2,3]. AMD can be produced in active or abandoned mining sites and is a major source of contamination for freshwater ecosystems worldwide [4]. The details of AMD formation have been described extensively [5,6]. Briefly, AMD runoff is usually highly acidic and contains high concentrations of dissolved metals. Iron, Mn, and Al are frequently the highest in concentration in coal mine drainage sites, along with elevated amounts of other toxic heavy metals [7]. Additionally, the chemical reactions of the AMD with the stream water downstream of discharges often results in the precipitation of metals, primarily Fe and Al oxyhydroxides, which bury the stream substratum. Both the chemical and physical stressors from AMD have been shown to have severe impacts on eukaryotic-cell organisms in stream food webs [8].
The passive treatment of AMD associated with coal mining to remediate impacted streams has become prevalent in the past 30 years [7]. There are several types of passive treatment, all of which rely on gravity, chemical reactions with alkaline minerals, and biological processes to neutralize acid and remove metals [9,10,11]. Streams remediated using passive treatment for AMD have shown varying degrees of biological recovery in benthic algal and macroinvertebrate communities relative to reference streams [12,13,14]. The role of microbes in contributing to AMD has been examined [2,15]; however, the effects of AMD and remediation methods on stream bacterial communities have received less attention, especially compared to eukaryotes [16]. Nutrient cycling and organic matter processing by benthic and aqueous microbial communities are critical to stream ecosystems [17], and AMD runoff has been shown to significantly impair these processes [18,19,20]. Bott et al. [21] found that some microbial processes in AMD-impacted streams were restored by passive treatment. The microbial community composition in AMD stream sites has been characterized [22,23,24], but more information is needed on the degree to which the microbial community structure impaired by AMD are restored by passive treatment.
The habitat of microbial communities in streams often corresponds to specific processes. Aqueous communities are involved in the cycling of dissolved organic matter and nutrients through the “microbial loop”, while carbon, nitrogen, and phosphorus transformations are carried out by sediment-associated microbes on the stream bed [17,25]. Organic matter from fallen tree leaves is the primary energy source for food webs in small streams with forested watersheds. The microbes colonizing the leaves are food for many aquatic macroinvertebrates, which themselves are preyed upon by fish [26]. Changes in the composition of the microbial communities in AMD streams may eliminate groups vital for the above processes. For example, leaf litter decomposition and microbial respiration rates on leaves are generally much lower in streams affected by AMD [18,27,28]. However, this relationship does not always correspond to a lower microbial biomass in AMD streams [29], suggesting that microbial composition also plays a role [30]. Finally, it is unclear whether the microbial community structure in different types of habitats respond similarly to AMD impacts and treatment remediation.
In this study, we sampled microbial communities from aqueous, sediment, and leaf habitats at three different stream sites: an untreated AMD site, a remediated site below AMD passive treatment, and an unimpaired reference site. Our objectives were (1) to determine the impact AMD has on microbial community structure in the three different types of stream habitats, (2) to ascertain the degree to which those communities are restored by passive treatment, and (3) to assess whether the similarity among the three types of communities (beta diversity) was altered by AMD and treatment. To the best of our knowledge, only one study has previously examined microbial community structure in relation to the passive treatment of AMD [16]. Our study differs by including samples from a reference stream to determine the degree of recovery and examining the microbial communities on leaves in addition to those in sediment and aqueous habitats.
2. Materials and Methods
2.1. Site Characterization and Sampling
Three first- or second-order forested-stream sites located in the Western Allegheny Plateau ecoregion of Pennsylvania, United States (latitude 41°70 31.00″ N, longitude 79°510 38.00″ W) were chosen to examine the effects of AMD on microbial community composition (Table 1). Two of the sites were within the headwaters of Slippery Rock Creek, which is underlain by shale, siltstone, and sandstone, with thin limestone and coal. The area has been mined for coal for over 100 years. About 25% of the land area in the 70 km2 subwatershed is underlain by drift mines, while 22% is partially reclaimed surface mines. Although there has been no major coal mining activity in this area since the early 1990s, dozens of abandoned mines have resulted in over 70 AMD discharges into the streams. These range in pH from 3.0 to 6.1; can have net acidities greater than 1000 mg CaCO3/L; and have iron, aluminum, and manganese concentrations that can exceed 100 mg/L. One sample site originated from an untreated AMD discharge (U) in this subwatershed. From 1995 to 2003, 15 reclamation projects were carried out in the subwatershed, including the construction of 12 passive treatment systems. Treatment systems in the headwaters area were designed for the specific water chemistry of the discharges and consist of anoxic limestone drains (ALD), reducing alkalinity-producing systems, horizontal-flow limestone beds, and constructed aerobic wetlands (system types reviewed by [5,10]). A large amount of the discharge at the second stream site (T) comprised treated AMD from an upstream ALD passive-treatment system constructed in 1995. In this system, a 0.015 m3/s AMD discharge is diverted through a buried, anoxic limestone bed (1500 tons) to raise alkalinity, then flows into a series of aerobic wetlands (1675 m2) to precipitate metals. Detailed information related to the design and long-term chemical monitoring of the performance of this treatment system (SR 114) and other systems in the subwatershed can be found at the Datashed website [31]. The third stream site was in a watershed outside of the Slippery Rock Creek headwaters area. It has never had AMD impacts on its drainage, supported brook trout, and represented the reference condition (R).
The study was conducted from September to November 2019. Stream water pH, acidity, SO4, and conductivity were measured in the field when microbial samples were taken using standard methods [32]. Conductivity and SO4 are considered the best indicators of the degree of AMD impacts in streams [33]. However, to provide a more detailed description of the water chemistry, we also report ranges for metal and nutrient concentrations measured in previous studies conducted by the authors and others at the 3 stream sites [33,34]. In these previous studies, water samples for dissolved metal analysis were analyzed in the laboratory for Fe, Mn, and Al using a 710-OES inductively coupled plasma spectrophotometer (Varian/Agilent, Santa Clara, CA, USA) following standard procedures that assure accuracy and precision [32]. The total P of water samples was determined using the ascorbic acid–molybdate blue method [35]. Nitrate+Nitrite was determined by a cadmium reduction reaction [34]. Ammonia was measured by the salicylate method [36] using reagents from HACH method 8155 (HACH, Loveland, CO, USA).
Samples of microbial communities were obtained from three types of habitats within each stream site. The microbes present in water were collected and genomic DNA obtained via a water prep kit (Qiagen, Fredrick, MD, USA). DNA for the sediment community was obtained using a soil prep kit (Qiagen, Fredrick, MD, USA). Recently fallen red maple leaves (Acer rubrum) were collected at the stream sites and epiphytic microbes were processed for genomic DNA following the methods of Chen and Kuo [37] and McNamara and Leff [30]. There were 2 replicates for samples from leaves at site R and from leaves at site U. Thus, there were a total of 11 microbial community samples (3 sites × 3 habitat types, plus the 2 replicates).
2.2. Genomics and Data Analysis
Genomic DNA from the microbial community samples was sent to Genewiz (South Plainfield, NJ, USA) for 16S rRNA gene sequencing. Operational taxonomic units (OTU’s) are assumed to be derived from a single species. To obtain the classification levels and community composition information, blastn software (Qiime 1.9.1) was used to map the OTU representative sequences with a 97% similarity level to the Nt database, and the best match was found. The microbial community structure was based on the relative abundance data of classified genera. Alpha diversity (Shannon Index, H’) and richness were determined for each of the 11 microbial communities. Beta diversity among communities in the 3 habitat types within each stream site was determined based on sum-of-squares of the sample-by-community data matrix with a chord transformation using the package adespatial [38] in R [39]. Communities from replicate samples were averaged for this analysis. Non-metric multidimensional scaling (NMDS) ordination based on a Bray–Curtis resemblance matrix was performed to illustrate differences among the 11 microbial communities using the vegan package in R [40]. Data for NMDS were square-root-transformed to reduce the influence of highly abundant taxa.
3. Results
3.1. Water Chemistry
The water chemistry at the three sites reflected the impacts from the AMD and treatment (Table 1). Site R was a high-quality trout stream with circumneutral pH and no acidity. Site T was approximately 50 m downstream of the outlet for an axonic limestone drain treatment system and about 1000 m below a mine-land reclamation project. Specific information about the performance of the treatment system (SR 114) is available at the Datashed website [31]. The stream chemistry at T was slightly more acidic, had higher SO4, conductivity, and had higher Fe and Mn concentrations than R. Site U received most of its discharge from an untreated AMD source. Severe impacts at this site were indicated by extremely acidic conditions and very high conductivity and SO4. The metal concentrations were 2–3 orders of magnitude greater than those in R. The total P and NO3− concentrations were similar at all sites, but the NH4+ concentration increased by two orders of magnitude over the AMD gradient of the three sites (Table 1). The benthic substrata at sites T and U were metal oxyhydroxide precipitates, primarily orange Fe(OH)3.
3.2. Microbial Community Composition
A total of 58 genera were identified in the microbial samples. The richness among the communities ranged from 30 to 11 and was lowest for sediment communities at the U and T sites (Table 2). Community alpha diversity did not correspond to either the site or habitat type (Table 2). The beta diversity among the three habitat sites was 0.95 for R, 0.89 for T, and 0.78 for U. This indicates that the variation in the composition of the microbial communities among the three types of habitat decreased (they became more similar) with increasing AMD impact.
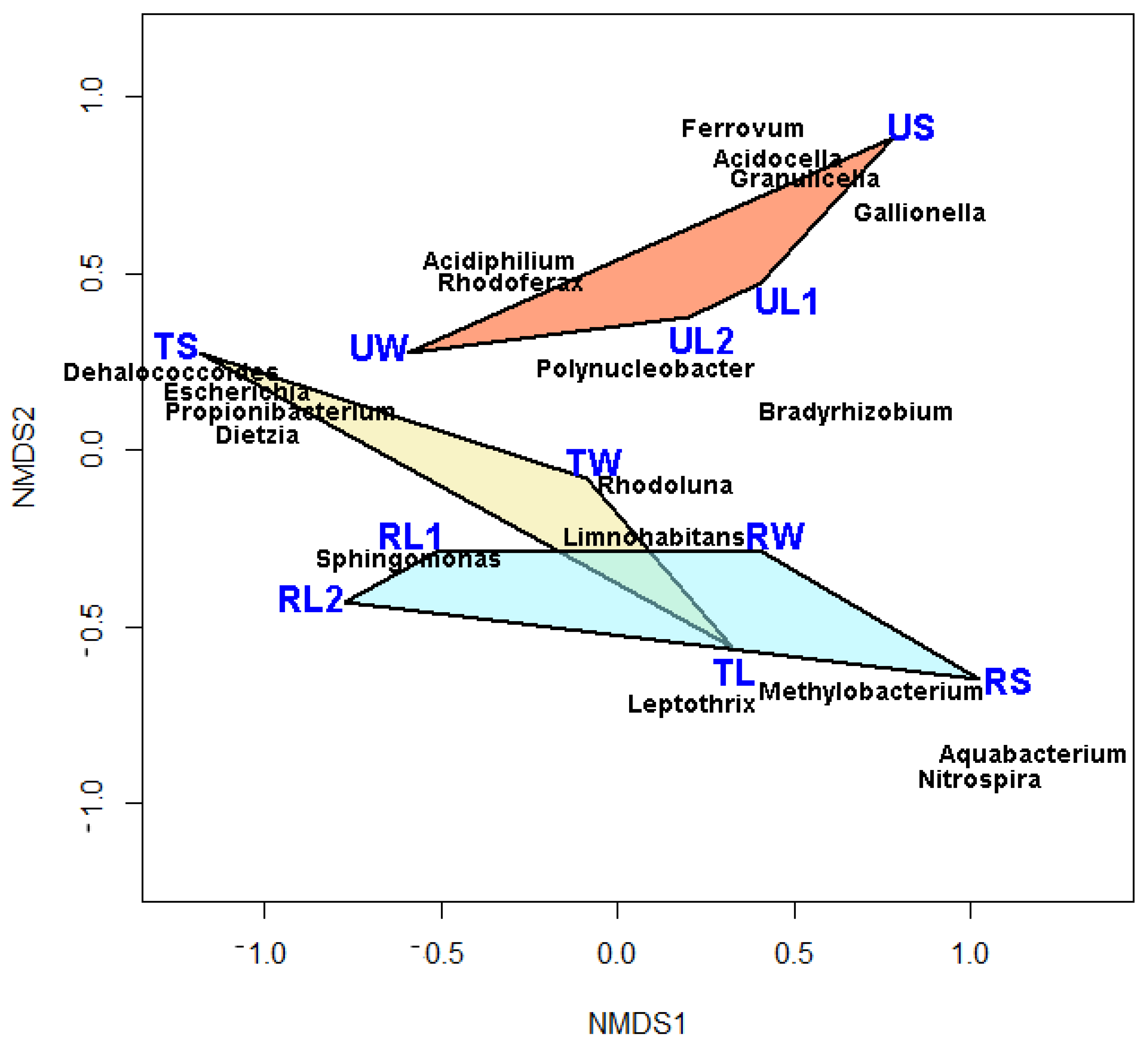
The ordination of the communities had a low stress value of 0.10, indicating a very good representation of communities in two-dimensional space. The community compositions of replicate samples were much more similar to each other than to the other samples (Figure 1). Alpha and beta proteobacteria dominated communities from water and leaf habitats at the T and R sites. Aqueous communities at these two sites were primarily members of Limnohabitans and Rhodoluna, with mostly Sphingomonas characterizing leaf communities (Figure 1 and Table 2). Most communities from U were distinct from the other two sites because of an abundance of acidophilic/acidotolerant genera, including Acidocella and Granulicella. However, there was an abundance of Sphingomonas in the untreated aqueous community, a taxon that was also common on leaves at the T and R sites. Polynucleobacter characterized leaf communities at U. Sediment communities among the three sites were most dissimilar because each was defined by distinct taxa. Nitrospira and Aquabacterium characterized sediment communities at the reference site, Propionibacterium at the treated site, and Gallionella at the untreated site (Figure 1 and Table 2).
4. Discussion
4.1. Water Chemistry
The chemical parameters at the three sites showed a gradient of AMD [5,13], with the potential impacts of acid and metals on microbial communities ranging from none at R to severe at U. DeNicola and Stapleton [13] found that the passive treatment system for AMD discharges above the sample location at site T reduced the acid and metal impacts on benthic algal communities relative to untreated sites in the same watershed. Consequently, the effects of AMD on the microbial communities at T should have been intermediate to the reference and untreated sites.
Low total P concentrations at the treated and untreated sites were probably from the P absorption onto metal oxyhydroxide precipitates on the substratum [33]. AMD-induced P limitation can inhibit microbial development [41], which may alter the community structure. We deliberately chose a reference site for the study that had a similar total P concentration in order to more clearly isolate the AMD and passive treatment effects related to pH and metals. The increase in N in the form of NH4+ with AMD impact was likely due to a loss of nitrogen-oxidizing bacteria [21], which is discussed in more detail in the next section.
4.2. Microbial Community Composition
Although replication was very limited, the similarity between replicate samples indicated a reasonable level of precision in the study. The ordination results showed that microbial community composition was influenced by both habitat type and AMD impacts. The aqueous community at R was dominated by Limnohabitans, an important freshwater planktonic genus commonly found in freshwater lakes and streams. This genus is believed to have a major role in nutrient cycling and as a food source for protozoans [42]. The common planktonic genus Rhodoluna also was abundant at R and likely involved in nutrient cycling [43]. The absence of these two taxa in the aqueous community at U indicates a serious impairment of the processes associated with the microbial loop. Their relatively high abundance in the aqueous communities at the T site indicates that the passive treatment resulted in some recovery of water column processes. The aqueous community at U was dominated by Sphingomonas. This genus is common on decomposing leaf litter and was the most abundant taxon on the leaves at both T and R (see discussion below).
The sediment communities at R primarily comprised Aquabacterium and Nitrospira. Aquabacterium is commonly found in uncontaminated freshwater biofilms [44]. Nitrospira is a ubiquitous chemolithoautotrophic genus that is a key nitrogen oxidizing bacterium in many ecosystems [45]. Nitrification is severely reduced and NH4+ correspondingly elevated in AMD-impacted streams [21]. The absence of Nitrospira in our U site is likely why the NH4+ concentrations were three orders of magnitude greater than at R. Nitrospira also was much less abundant and the NH4+ concentrations were one order of magnitude higher at T than R. This indicates that passive treatment at T did not restore the impacts associated with nitrification, possibly because the substrata was predominantly iron oxyhydroxides. Schlief [19] found that the presence of metal precipitates at treated and untreated AMD sites produced both chemical and physical impacts on benthic microbial communities. The lack of a natural substrata at T may also explain the dominance of Escherichia and Propionibacterium in the sediment community. Escherichia tend to come from autochthonous or allochthonous inputs of animal waste, while Propionibacterium is an actinomycete found in soils [46,47]. Both these taxa could be introduced from surrounding soils and take hold due to limited competition on the poor substrata at T. The iron-oxidizing chemolithoautotrophic Gallionella and the heterotrophic Acidocella were the abundant genera in the sediment community at U. Both have been reported at severely impacted AMD sites [24,48]. Another prevalent genus was Bradyrhizobium. While one might not think of this nitrogen-fixing genus being present in an AMD contaminated site, it has been identified in other sites impacted by mining [49,50]. Since members of this genus are also known to be associated with plants [51], it may have been introduced from riparian vegetation at U.
Various studies have found that proteobacteria, such as the genera Burkholderia, Granulicella, Pseudomonas, and Sphingomonus, are common in the microbial communities of uncontaminated leaf litter [52,53], including decomposing maple leaves [54,55]. Therefore, the dominance of Sphingomonas on maple leaves at R suggests a natural, non-impacted environment. This also was the most abundant taxon on the leaves at T, suggesting that treatment made conditions there favorable for the breakdown of microbial leaf. Sphingomonas was the most abundant taxon in the aqueous community at U but comprised a relatively small portion of the leaf community. Instead, the leaf community at U more closely resembled that of the sediment community at that site characterized by Acidocella, Gallionella, and Granulicella. Due to the acidic nature of this site (see Table 1), any bacteria present would need to be well adapted to tolerate such conditions. Therefore, it is no surprise to see acidophilic/acidotolerant genera dominating. Additionally, Polynucleobacter was abundant. All of these are typical of sites impacted by mining but not commonly associated with the decomposition of organic matter [22,24,48,56,57]. This corroborates Schilief’s [19] suggestion that the heavy precipitation of metal oxyhydroxides on leaves in highly impacted AMD sites creates an environment similar to sediment substrata, and may reduce leaf litter decomposition by forming a barrier to colonization of the leaf surface. Correspondingly, several studies have found that the processing of this important energy source in streams is greatly reduced by AMD runoff [19,20,27,58].
Nitrification is an important part of a normal, healthy habitat, supported by the presence of Nitrospira at site R. In order to restore nitrification to an AMD site, one must also restore the nitrifying bacteria. No nitrifying bacteria were identified at sites U or T. This is likely a result of the conditions at these sites. Ammonia-oxidizing bacteria (AOB), which are involved in the first step of the nitrification process, are highly sensitive to, and have decreased growth rates, in response to environmental conditions, such as pH and metal concentrations [59]. As shown above in Table 1, both T and U had an increased concentration of metals, possibly inhibiting AOBs. At U, there is also a very low pH. Due to these two factors, it would be expected that the growth of AOB is significantly depressed, if not inhibited completely. Further study is required to determine what approach to restoring this critical group of bacteria is most effective.
The richness and alpha diversity of the microbial communities in this study were generally not useful metrics for distinguishing the habitat type or the effects of AMD treatment. This contrasts with Ly et al. [16], who found less diverse microbial communities at sites with raw AMD. However, our beta diversity trends corresponded to the findings of Ly et al. [16] in that the similarity of communities among different habitats increased with AMD impact. At U, the communities in the three habitats were constrained to a pool of taxa tolerant to AMD stressors, making the communities more similar. Conversely, the microbial communities in the three habitats at R were quite different because the growth of specialized taxa was not eliminated by AMD stressors. The beta diversity at T was intermediate between R and U, reflecting a reduction in the AMD stress that resulted in a partial recovery of separate community structures among the habitats.
It is clear that microbial community composition is highly impacted by AMD. Although passive treatment at site T in this study did not completely reduce the stream acidity and soluble metal concentrations to the reference condition, it was successful in shifting the water and leaf-litter community composition towards those of the reference condition. Contrary to this, treatment did not appear to restore the sediment community composition towards those of the reference site. An interesting question to address in future research is whether treatment strategies that include removing legacy (prior to treatment) metal oxyhydroxides from the stream substratum may improve the restoration of sediment communities. Despite being built in 1995, the monitoring of the treatment system at site T indicates that it continues to perform well [31]; however, eventually the limestone in the ALD systems needs to be replaced and the wetlands need to be dredged of precipitates in order to continue treatment into the future [10]. In this study, the implications of how the recovery of microbial communities due to passive treatment may affect functional processes were based on what is known about the ecophysiology of the taxa. Subsequent studies that empirically couple the recovery of microbial community composition with measured functional processes would more conclusively help restoration ecologists to design and build systems that remediate AMD streams to more natural conditions.
5. Conclusions
Microbial community composition was synergistically affected by both habitat type and AMD impact, with the similarity among communities in the three habitats increasing with the severity of the AMD. Our results indicate that passive treatment restored some of these microbial communities toward the reference condition better than others. Additionally, the ecophysiological requirements of the taxa suggested that restoration would potentially improve some habitat-specific functional processes. For example, the taxa corresponding to the microbial loop in the water column and leaf decomposition were similar at the T and R sites, but the taxa associated with nitrification in sediment communities at T did not recover. Thus, achieving the complete restoration of functional processes in AMD-impaired streams requires a better understanding of how stressors affect taxonomic composition within different types of habitats, which can then be used to establish thresholds for rehabilitation.
Author Contributions
Conceptualization, C.M. and D.M.D.; methodology, C.M. and D.M.D.; field sampling, A.N.N.; formal analysis, A.N.N., C.M., D.M.D.; investigation, A.N.N., C.M., D.M.D.; writing—original draft preparation, A.N.N. and D.M.D.; writing—review and editing, C.M.; supervision, C.M. and D.M.D.; funding acquisition, C.M., A.N.N. and D.M.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by a Slippery Rock University Faculty Student Research Grant Awarded to Chris Maltman and Dean DeNicola.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rothschild, L.; Mancinelli, R. Life in extreme environments. Nature 2001, 409, 1092–1101. [Google Scholar] [CrossRef] [PubMed]
- Baker, B.; Banfield, J. Microbial communities in acid mine drainage. FEMS Microbiol. Ecol. 2003, 44, 139–152. [Google Scholar] [CrossRef] [Green Version]
- Aguinaga, O.E.; McMahon, A.; White, K.N.; Dean, A.P.; Pittman, J.K. Microbial community shifts in response to acid mine drainage pollution within a natural wetland ecosystem. Front. Microbiol. 2018, 9, 1445. [Google Scholar] [CrossRef] [PubMed]
- Reid, W.V.; Mooney, H.A.; Cropper, A.; Capistrano, D.; Carpenter, S.R.; Chopra, K.; Dasgupta, P.; Dietz, T.; Duraiappah, A.K.; Hassan, R.; et al. Ecosystems and Human Well-Being-SYNTHESIS: A Report of the Millennium Ecosystem Assessment; Sarukhan, J., Whyte, A., Board, M., Eds.; Island Press: Washington, DC, USA, 2005; Volume 5. [Google Scholar]
- Younger, P.L.; Banwart, S.A.; Hedin, R.S. Mine water hydrology. In Mine Water; Springer: Dordrecht, The Netherlands, 2002; Volume 5, pp. 127–270. [Google Scholar] [CrossRef]
- Simate, G.S.; Ndlovu, S. Acid mine drainage: Challenges and opportunities. J. Environ. Chem. Eng. 2014, 2, 1785–1803. [Google Scholar] [CrossRef]
- Watzlaf, G.R.; Schroeder, K.T.; Kleinmann, R.L.; Kairies, C.L.; Nairn, R.W. The Passive Treatment of Coal Mine Drainage; National Energy Technology Laboratory: Pittsburgh, PA, USA, 2004; pp. 1–72. [Google Scholar]
- Hogsden, K.L.; Harding, J.S. Anthropogenic and natural sources of acidity and metals and their influence on the structure of stream food webs. Environ. Pollut. 2012, 162, 466–474. [Google Scholar] [CrossRef]
- Johnson, D.B.; Hallberg, K.B. Pitfalls of passive mine water treatment. Rev. Environ. Sci. Biotechnol. 2002, 1, 335–343. [Google Scholar] [CrossRef]
- Ziemkiewicz, P.F.; Skousen, J.G.; Simmons, J. Long-term performance of passive acid mine drainage treatment systems. Mine Water Environ. 2003, 22, 118–129. [Google Scholar] [CrossRef]
- Skousen, J.; Zipper, C.E.; Rose, A.; Ziemkiewicz, P.F.; Nairn, R.; McDonald, L.M.; Kleinmann, R.L. Review of passive systems for acid mine drainage treatment. Mine Water Environ. 2017, 36, 133–153. [Google Scholar] [CrossRef] [Green Version]
- Nelson, S.M.; Roline, R.A. Recovery of a stream macroinvertebrate community from mine drainage disturbance. Hydrobiologia 1996, 339, 73–84. [Google Scholar] [CrossRef]
- DeNicola, D.M.; Stapleton, M.G. Benthic diatoms as indicators of long-term changes in a watershed receiving passive treatment for acid mine drainage. Hydrobiologia 2014, 732, 29–48. [Google Scholar] [CrossRef]
- Williams, K.M.; Turner, A.M. Acid mine drainage and stream recovery: Effects of restoration on water quality, macroinvertebrates, and fish. Knowl. Manag. Aquat. Ecosyst. 2015, 416, 18. [Google Scholar] [CrossRef] [Green Version]
- Rawat, N.; Singh, G. The role of micro-organisms in the formation of acid mine drainage in the north-eastern coal field of India. Int. J. Mine Water 1982, 2, 29–36. [Google Scholar] [CrossRef]
- Ly, T.; Wright, J.R.; Weit, N.; McLimans, C.J.; Ulrich, N.; Tokarev, V.; Valkanas, M.M.; Trun, N.; Rummel, S.; Grant, C.J.; et al. Microbial communities associated with passive acidic abandoned coal mine remediation. Front. Microbiol. 2019, 10, 1955. [Google Scholar] [CrossRef] [PubMed]
- Allan, J.D.; Castillo, M.M.; Capps, K. Stream Ecology: Structure and Function of Running Waters, 2nd ed.; Springer: Dordrecht, The Netherlands, 2007. [Google Scholar]
- Niyogi, D.K.; Lewis, W.M., Jr.; McKnight, D.M. Litter breakdown in mountain streams affected by mine drainage: Biotic mediation of abiotic controls. Ecol. Appl. 2001, 11, 506–516. [Google Scholar] [CrossRef]
- Schlief, J. Leaf associated microbial activities in a stream affected by acid mine drainage. Int. Rev. Hydrobiol. 2004, 89, 467–475. [Google Scholar] [CrossRef]
- Baeseman, J.L.; Smith, R.L.; Silverstein, J. Denitrification potential in stream sediments impacted by acid mine drainage: Effects of pH, various electron donors, and iron. Microb. Ecol. 2006, 51, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Bott, T.L.; Jackson, J.K.; McTammany, M.E.; Newbold, J.D.; Rier, S.T.; Sweeney, B.W.; Battle, J.M. Abandoned coal mine drainage and its remediation: Impacts on stream ecosystem structure and function. Ecol. Appl. 2012, 22, 2144–2163. [Google Scholar] [CrossRef]
- Lear, G.; Niyogi, D.; Harding, J.; Dong, Y.; Lewis, G. Biofilm bacterial community structure in streams affected by acid mine drainage. Appl. Environ. Microbiol. 2009, 75, 3455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuang, J.L.; Huang, L.N.; Chen, L.X.; Hua, Z.S.; Li, S.J.; Hu, M.; Li, J.T.; Shu, W.S. Contemporary environmental variation determines microbial diversity patterns in acid mine drainage. ISME J. 2013, 7, 1038–1050. [Google Scholar] [CrossRef] [Green Version]
- Méndez-García, C.; Peláez, A.I.; Mesa, V.; Sánchez, J.; Golyshina, O.V.; Ferrer, M. Microbial diversity and metabolic networks in acid mine drainage habitats. Front. Microbiol. 2015, 6, 475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Findlay, S. Stream microbial ecology. J. N. Am. Benthol. Soc. 2010, 29, 170–181. [Google Scholar] [CrossRef] [Green Version]
- Webster, J.R.; Benfield, E.F. Vascular plant breakdown in freshwater ecosystems. Annu. Rev. Ecol. Evol. Syst. 1986, 17, 567–594. [Google Scholar] [CrossRef]
- Simmons, J.A.; Lawrence, E.R.; Jones, T.G. Treated and untreated acid mine drainage effects on stream periphyton biomass, leaf decomposition, and macroinvertebrate diversity. J. Freshwater Ecol. 2005, 20, 413–424. [Google Scholar] [CrossRef] [Green Version]
- Johnson, K.S.; Thompson, P.C.; Gromen, L.; Bowman, J. Use of leaf litter breakdown and macroinvertebrates to evaluate gradient of recovery in an acid mine impacted stream remediated with an active alkaline doser. Environ. Monit. Assess. 2014, 186, 4111–4127. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, J.; Odum, W.E.; Mills, A. Leaf litter decomposition in a reservoir affected by acid mine drainage. Oikos 1983, 41, 165–172. [Google Scholar] [CrossRef]
- McNamara, C.J.; Leff, L.G. Bacterial community composition in biofilms on leaves in a northeastern Ohio stream. J. N. Am. Benthol. Soc. 2004, 23, 677–685. [Google Scholar] [CrossRef]
- Datashed. Available online: https://www.datashed.org/ (accessed on 3 November 2021).
- American Public Health Association. Standard Methods for the Evaluation of Water and Wastewater; American Public Health Association: Washington, DC, USA, 1998; p. 766. [Google Scholar]
- DeNicola, D.M.; Lellock, A.J. Nutrient limitation of algal periphyton in streams along an acid mine drainage gradient. J. Phycol. 2015, 51, 739–749. [Google Scholar] [CrossRef] [PubMed]
- DeNicola, D.M.; Stapleton, M.G.; Onufrak, A.J. Examination of effects of acid and metals on benthic algae in streams using chemical diffusing substrata. Ecol. Indic. 2018, 88, 17–26. [Google Scholar] [CrossRef]
- Lind, O.T. Handbook of Common Methods in Limnology, 2nd ed.; Kendall/Hunt: Dubuque, IA, USA, 1985; p. 199. [Google Scholar]
- Bower, C.E.; Holm-Hansen, T. A salicylate-hypochloritemethod for determining ammonia in seawater. Can. J. Fish. Aquat. Sci. 1980, 37, 794–798. [Google Scholar] [CrossRef]
- Chen, W.P.; Kuo, T.T. A simple and rapid method for the preparation of gram-negative bacterial genomic DNA. Nucleic Acids Res. 1993, 21, 2260. [Google Scholar] [CrossRef]
- Dray, S.; Bauman, D.; Blanchet, G.; Borcard, D.; Clappe, S.; Guenard, G.; Jombart, T.; Larocque, G.; Legendre, P.; Madi, N.; et al. Adespatial: Multivariate Multiscale Spatial Analysis [R Package Version 0.0-9]. 2019. Available online: https://cran.r-project.org/web/packages/adespatial (accessed on 13 October 2021).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.; O’Hara, R.B.; Simpson, G.; Solymos, P.; et al. Vegan [R Package Version 2.5-6]. 2020. Available online: https://cran.r-project.org/package=vegan (accessed on 13 October 2021).
- Drerup, S.A.; Vis, M.L. Extracellular enzyme activity suggests phosphorus limitation of biofilm productivity in acid mine drainage remediated streams. Hydrobiologia 2017, 784, 225–236. [Google Scholar] [CrossRef]
- Kasalický, V.; Jezbera, J.; Hahn, M.W.; Šimek, K. The diversity of the Limnohabitans genus, an important group of freshwater bacterioplankton, by characterization of 35 isolated strains. PLoS ONE 2013, 8, e58209. [Google Scholar] [CrossRef]
- Hahn, M.W.; Schmidt, J.; Taipale, S.J.; Doolittle, W.F.; Koll, U. Rhodoluna lacicola gen. nov., sp. nov., a planktonic freshwater bacterium with stream-lined genome. IJSEM 2014, 64, 3254. [Google Scholar] [CrossRef] [PubMed]
- Kalmbach, S.; Manz, W.; Bendinger, B.; Szewzyk, U. In situ probing reveals Aquabacterium commune as a widespread and highly abundant bacterial species in drinking water biofilms. Water Res. 2000, 34, 575–581. [Google Scholar] [CrossRef]
- Koch, H.; Lücker, S.; Albertsen, M.; Kitzinger, K.; Herbold, C.; Spieck, E.; Nielsen, P.H.; Wagner, M.; Daims, H. Expanded metabolic versatility of ubiquitous nitrite-oxidizing bacteria from the genus Nitrospira. Proc. Natl. Acad. Sci. USA 2015, 112, 11371–11376. [Google Scholar] [CrossRef] [Green Version]
- Korajkic, A.; Wanjugi, P.; Brooks, L.; Cao, Y.; Harwood, V.J. Persistence and decay of fecal microbiota in aquatic habitats. Microbiol. Mol. Biol. Rev. 2019, 83, e00005-19. [Google Scholar] [CrossRef]
- Hayashi, S.; Furusaka, C. Enrichment of Propionibacterium in paddy soil by addition of various organic substances. Antonie Van Leeuwenhoek 1980, 46, 313–320. [Google Scholar] [CrossRef]
- Fabisch, M.; Freyer, G.; Johnson, C.A.; Büchel, G.; Akob, D.M.; Neu, T.R.; Küsel, K. Dominance of ‘Gallionella capsiferriformans’ and heavy metal association with Gallionella-like stalks in metal-rich pH 6 mine water discharge. Geobiology 2016, 14, 68–90. [Google Scholar] [CrossRef]
- Percak-Dennett, E.M. Microbial Iron Redox Cycling in Terrestrial Environments. Ph.D. Thesis, The University of Wisconsin-Madison, Madison, WI, USA, 2014. [Google Scholar]
- Giddings, L.A.; Chlipala, G.; Kunstman, K.; Green, S.; Morillo, K.; Bhave, K.; Peterson, H.; Driscoll, H.; Maienschein-Cline, M. Characterization of an acid rock drainage microbiome and transcriptome at the Ely Copper Mine Superfund site. PLoS ONE 2020, 15, e0237599. [Google Scholar] [CrossRef]
- Ormeño-Orrillo, E.; Martínez-Romero, E. A genomotaxonomy view of the Bradyrhizobium genus. Front. Microbiol. 2019, 10, 1334. [Google Scholar] [CrossRef]
- Tláskal, V.; Voříšková, J.; Baldrian, P. Bacterial succession on decomposing leaf litter exhibits a specific occurrence pattern of cellulolytic taxa and potential decomposers of fungal mycelia. FEMS Microbiol. Ecol. 2016, 92, fiw177. [Google Scholar] [CrossRef]
- Gołębiewski, M.; Tarasek, A.; Sikora, M.; Deja-Sikora, E.; Tretyn, A.; Niklińska, M. Rapid microbial community changes during initial stages of pine litter decomposition. Microb. Ecol. 2019, 77, 56–75. [Google Scholar] [CrossRef] [Green Version]
- Mcnamara, C.J.; Leff, L.G. Response of biofilm bacteria to dissolved organic matter from decomposing maple leaves. Microb. Ecol. 2004, 48, 324–330. [Google Scholar] [CrossRef]
- Newman, M.M.; Liles, M.R.; Feminella, J.W. Litter breakdown and microbial succession on two submerged leaf species in a small forested stream. PLoS ONE 2015, 10, e0130801. [Google Scholar] [CrossRef]
- Edberg, F.; Andersson, A.F.; Holmström, S.J. Bacterial community composition in the water column of a lake formed by a former uranium open pit mine. Microb. Ecol. 2012, 64, 870–880. [Google Scholar] [CrossRef]
- Ettamimi, S.; Carlier, J.D.; Cox, C.J.; Elamine, Y.; Hammani, K.; Ghazal, H.; Costa, M.C. A meta-taxonomic investigation of the prokaryotic diversity of water bodies impacted by acid mine drainage from the São Domingos mine in southern Portugal. Extremophiles 2019, 23, 821–834. [Google Scholar] [CrossRef]
- Ferreira, V.; Koricheva, J.; Duarte, S.; Niyogi, D.K.; Guérold, F. Effects of anthropogenic heavy metal contamination on litter decomposition in streams–A meta-analysis. Environ. Pollut. 2016, 210, 261–270. [Google Scholar] [CrossRef]
- Li, X.; Kapoor, V.; Impellitteri, C.; Chandrran, K.; Santo Domingo, J. Measuring nitrification inhibition by metals in wastewater treatment systems: Current state of science and fundamental research needs. Crit. Rev. Environ. Sci. Technol. 2016, 46, 249–289. [Google Scholar] [CrossRef]
Figure 1.
NMDS ordination biplot for microbial communities from 3 types of habitats at 3 sites along an AMD gradient. Stress = 0.10. Only the most abundant genera are shown. Community codes: U = untreated site; T = treated site; R = reference site; W = water habitat; L = leaf habitat; S = sediment habitat; 1 and 2 indicate replicate samples. Communities from the 3 habitats are connected for each respective site.
Figure 1.
NMDS ordination biplot for microbial communities from 3 types of habitats at 3 sites along an AMD gradient. Stress = 0.10. Only the most abundant genera are shown. Community codes: U = untreated site; T = treated site; R = reference site; W = water habitat; L = leaf habitat; S = sediment habitat; 1 and 2 indicate replicate samples. Communities from the 3 habitats are connected for each respective site.


{kind=link}
Table 1.
Water chemistry parameters for the 3 stream sites. pH, SO4, conductivity and acidity were measured concurrently with microbial samples in this study. * Three samples in 2016. # Two samples in 2014. @ Three samples in 2013. $ 18–31 samples in 2002–2004 (EPA Storet Data base).
Table 1.
Water chemistry parameters for the 3 stream sites. pH, SO4, conductivity and acidity were measured concurrently with microbial samples in this study. * Three samples in 2016. # Two samples in 2014. @ Three samples in 2013. $ 18–31 samples in 2002–2004 (EPA Storet Data base).
| Parameter | Reference (R) | Treated (T) | Untreated (U) |
|---|---|---|---|
| Discharge (m3/s) | 0.17 | 0.07 | 0.01 |
| pH | 7.2 | 6.6 | 3.1 |
| SO4 (mg/L) | 39 | 90 | 770 |
| Conductivity (µs/cm) | 136 | 223 | 1727 |
| Acidity (mg CaCO3/L) | 0 | 10 | 149 |
| Soluble Fe (mg/L) | 0.02–0.10 $ | 0.31–0.52 @ | 6.63–11.72 @ |
| Soluble Mn (mg/L) | 0.02–0.09 $ | 1.07–1.12 @ | 25.52–45.89 @ |
| Soluble Al (mg/L) | 0.01–0.12 $ | 0.05–0.05 @ | 5.73–13.89 @ |
| Metal Precipitate | No | Yes | Yes |
| Total P (µg/L) | 3–5 * | <2–17 # | <2–8 # |
| NO3 + NO2-N (mg/L) | 0.21–0.73 * | 0.2–0.4 # | 0.6–0.9 # |
| NH4-N (mg/L) | 0.006–0.012 * | 0.06–0.08 # | 0.25–0.58 # |
Table 2.
Relative abundance heat map of the most-abundant-classified genera in microbial communities in water, on leaves and in sediment at 3 sites comprising an AMD gradient (values were averaged for replicates from the same community). Colors indicate magnitude of values: green = lowest, yellow = 50th percentile, and red = highest. Shannon Diversity and richness are given for each community. U = untreated site; T = treated site; R = reference site; W = water habitat; L = leaf habitat; S = sediment habitat.
Table 2.
Relative abundance heat map of the most-abundant-classified genera in microbial communities in water, on leaves and in sediment at 3 sites comprising an AMD gradient (values were averaged for replicates from the same community). Colors indicate magnitude of values: green = lowest, yellow = 50th percentile, and red = highest. Shannon Diversity and richness are given for each community. U = untreated site; T = treated site; R = reference site; W = water habitat; L = leaf habitat; S = sediment habitat.
| Taxon | U-W | U-L | U-S | T-W | T-L | T-S | R-W | R-L | R-S |
|---|---|---|---|---|---|---|---|---|---|
| Sphingomonas | 0.49 | 0.06 | 0.00 | 0.09 | 0.39 | 0.00 | 0.02 | 0.83 | 0.01 |
| Escherichia | 0.08 | 0.00 | 0.00 | 0.00 | 0.00 | 0.22 | 0.00 | 0.02 | 0.00 |
| Polynucleobacter | 0.07 | 0.32 | 0.00 | 0.06 | 0.00 | 0.00 | 0.10 | 0.01 | 0.00 |
| Propionibacterium | 0.06 | 0.01 | 0.00 | 0.02 | 0.01 | 0.39 | 0.00 | 0.04 | 0.00 |
| Acidiphilium | 0.03 | 0.01 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Acidocella | 0.02 | 0.20 | 0.16 | 0.01 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Ferrovum | 0.02 | 0.01 | 0.07 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Dehalococcoides | 0.00 | 0.00 | 0.13 | 0.00 | 0.01 | 0.00 | 0.00 | 0.00 | 0.00 |
| Rhodoluna | 0.00 | 0.00 | 0.02 | 0.30 | 0.01 | 0.00 | 0.15 | 0.01 | 0.00 |
| Granulicella | 0.00 | 0.15 | 0.02 | 0.01 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Dietzia | 0.04 | 0.00 | 0.00 | 0.00 | 0.00 | 0.14 | 0.00 | 0.01 | 0.00 |
| Gallionella | 0.00 | 0.07 | 0.27 | 0.00 | 0.01 | 0.00 | 0.00 | 0.00 | 0.02 |
| Limnohabitans | 0.00 | 0.01 | 0.00 | 0.37 | 0.02 | 0.00 | 0.39 | 0.02 | 0.00 |
| Bradyrhizobium | 0.00 | 0.01 | 0.18 | 0.02 | 0.04 | 0.02 | 0.02 | 0.00 | 0.10 |
| Methylobacterium | 0.00 | 0.00 | 0.00 | 0.00 | 0.07 | 0.00 | 0.00 | 0.00 | 0.01 |
| Rhodoferax | 0.00 | 0.00 | 0.02 | 0.01 | 0.00 | 0.03 | 0.00 | 0.00 | 0.00 |
| Leptothrix | 0.00 | 0.00 | 0.00 | 0.01 | 0.02 | 0.00 | 0.01 | 0.01 | 0.06 |
| Nitrospira | 0.00 | 0.00 | 0.00 | 0.00 | 0.02 | 0.00 | 0.00 | 0.00 | 0.20 |
| Aquabacterium | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.26 |
| Richness | 19 | 30 | 11 | 30 | 27 | 11 | 26 | 22 | 21 |
| Shannon Diversity | 2.02 | 2.14 | 1.99 | 1.93 | 2.51 | 1.75 | 2.14 | 0.86 | 2.36 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Neff, A.N.; DeNicola, D.M.; Maltman, C. Passive Treatment for Acid Mine Drainage Partially Restores Microbial Community Structure in Different Stream Habitats. Water 2021, 13, 3300. https://doi.org/10.3390/w13223300
AMA Style
Neff AN, DeNicola DM, Maltman C. Passive Treatment for Acid Mine Drainage Partially Restores Microbial Community Structure in Different Stream Habitats. Water. 2021; 13(22):3300. https://doi.org/10.3390/w13223300
Chicago/Turabian StyleNeff, Alexis N., Dean M. DeNicola, and Chris Maltman. 2021. "Passive Treatment for Acid Mine Drainage Partially Restores Microbial Community Structure in Different Stream Habitats" Water 13, no. 22: 3300. https://doi.org/10.3390/w13223300
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.