
Fast and Reliable Identification of Ammonia Phylotypes T1, T2 and T6 Using a Stereomicroscope: Implication for Large-Scale Ecological Surveys and Monitoring Programs

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
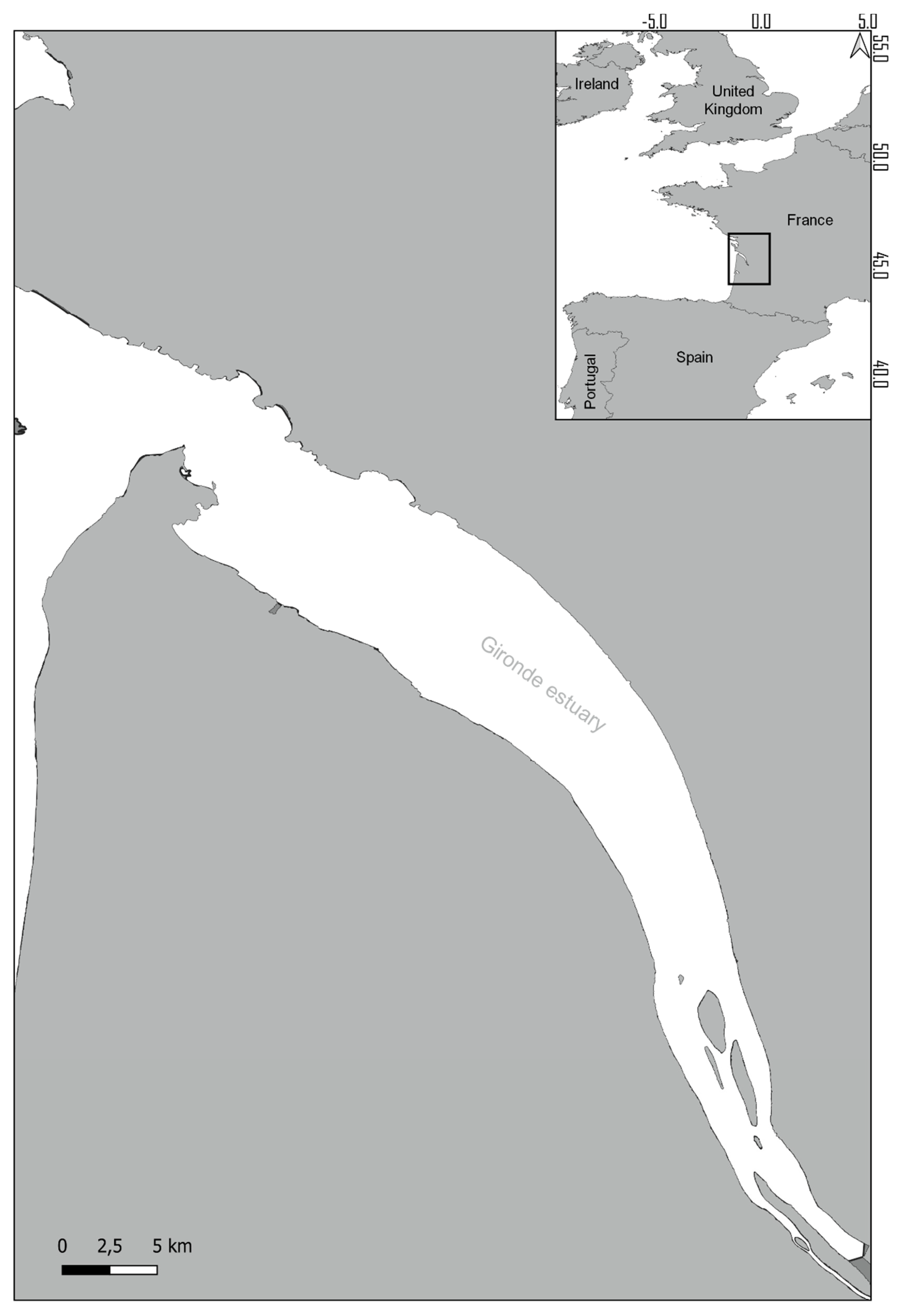
2.1. Sampling Site
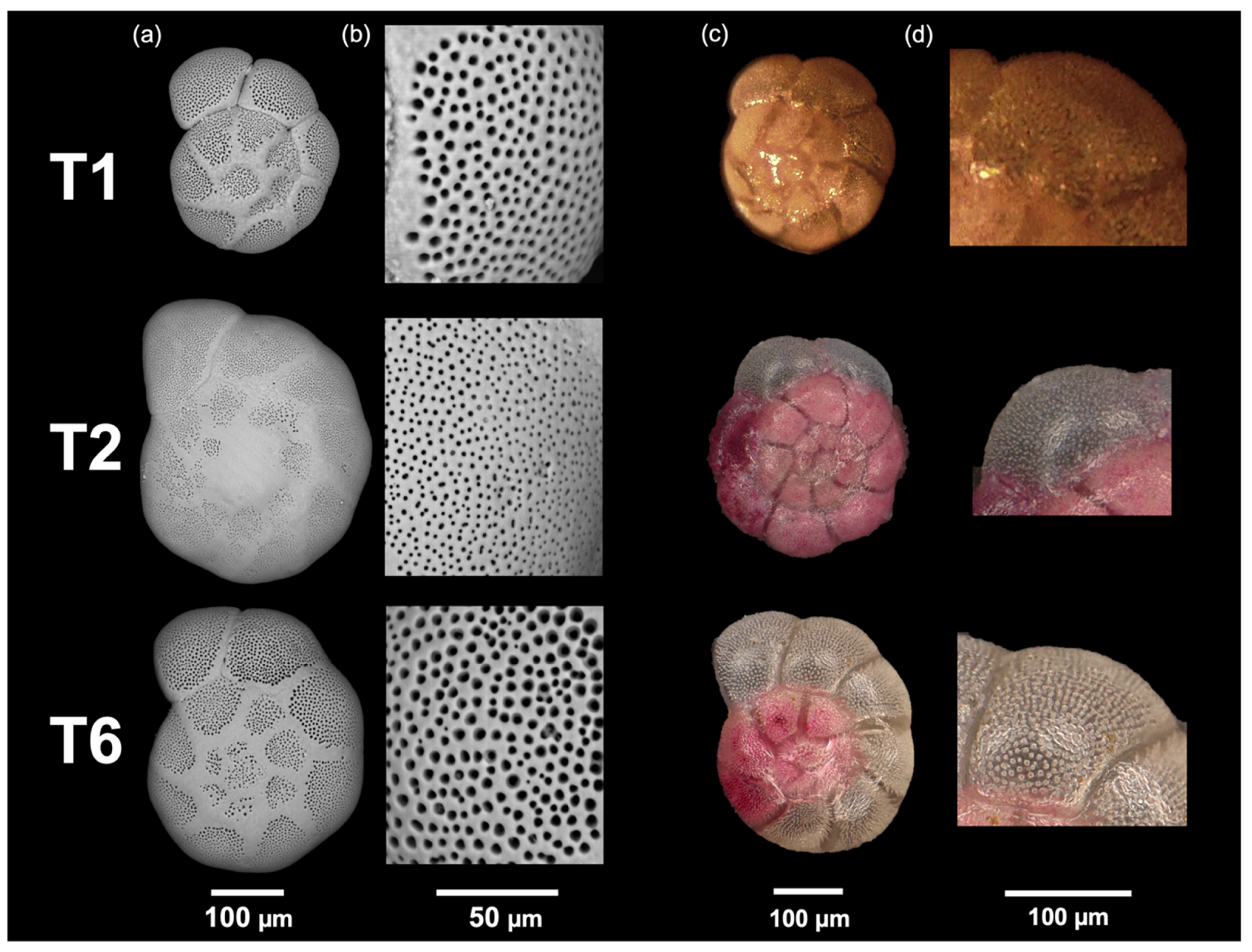
2.2. Ammonia Phylotype Identification
3. Results
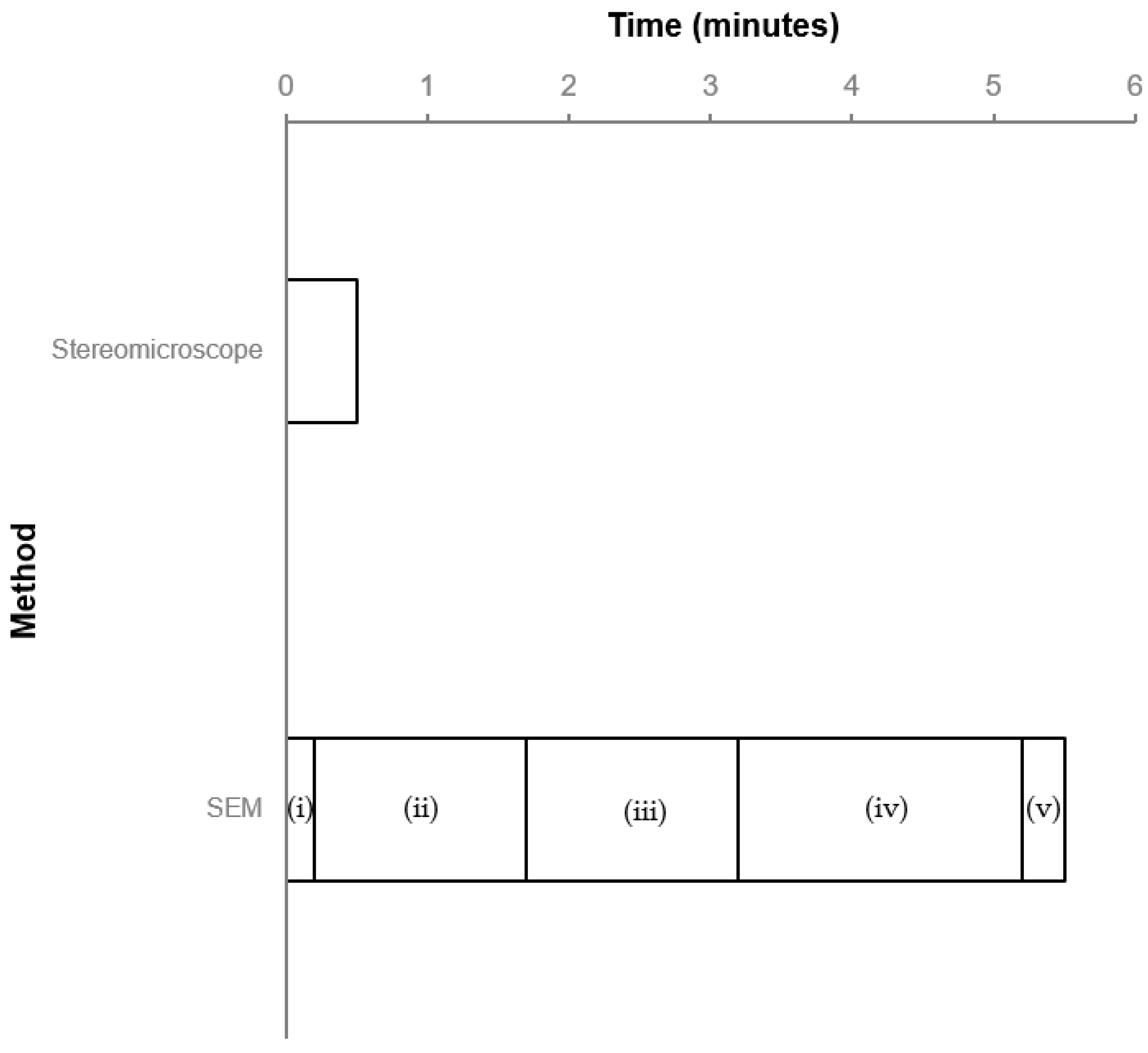
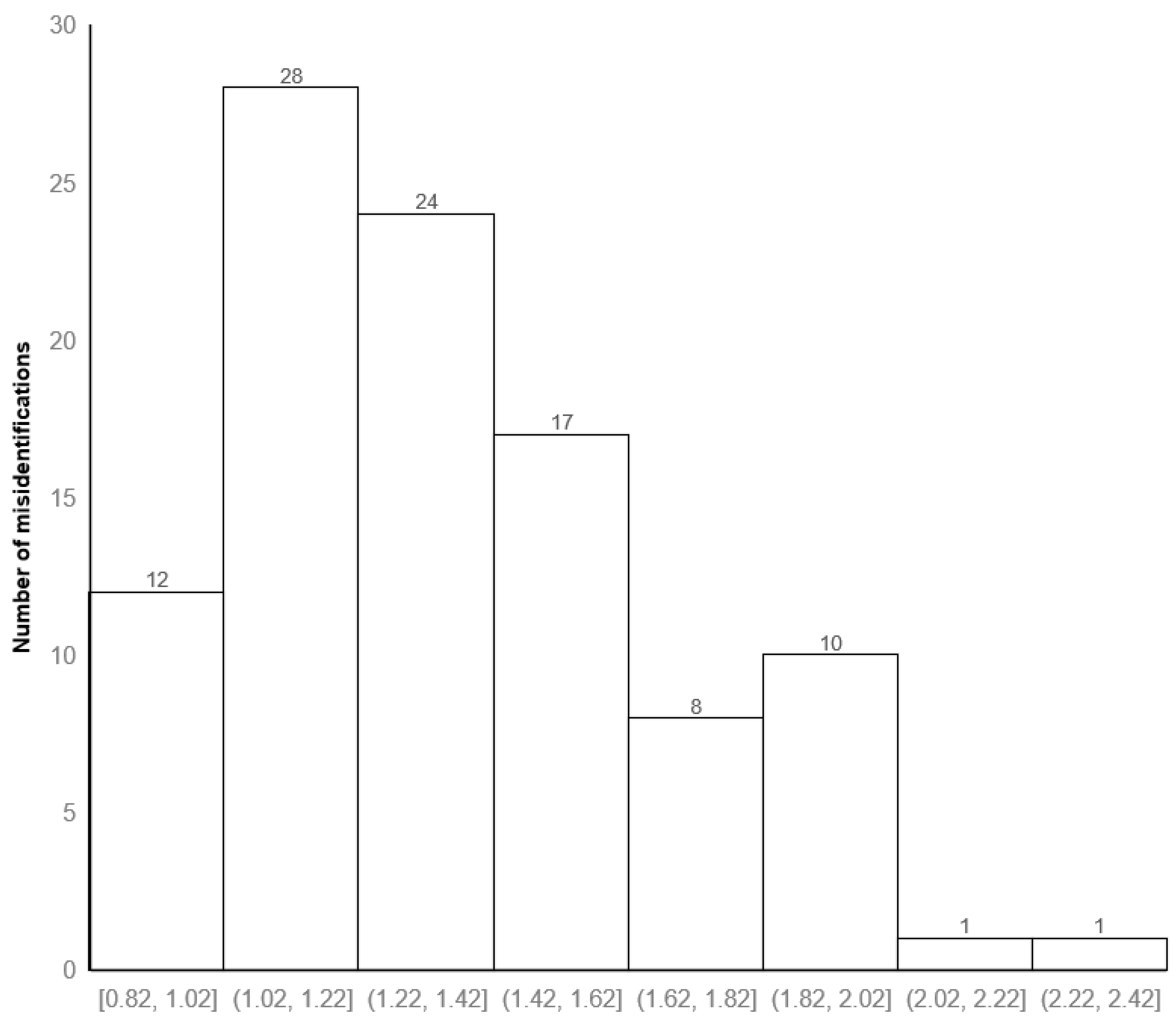
3.1. Identification Time
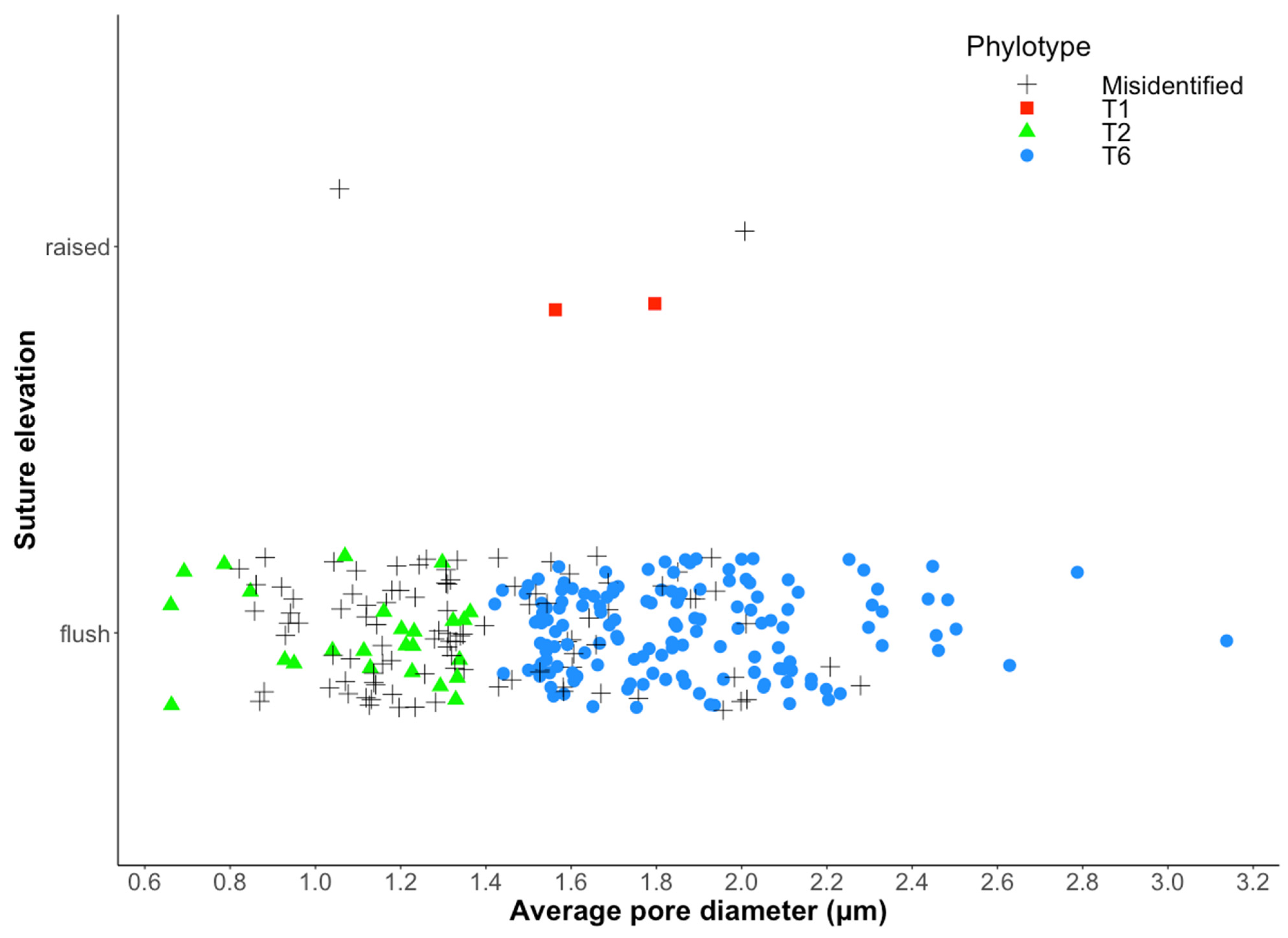
3.2. Correct Assignation Rate between the Two Identification Methods
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brünnich, M.T. Zoologiae Fundamenta. Hafniae Lipsiae Grund. I Dye. 1771.
- Hayward, B.W.; Holzmann, M.; Pawlowski, J.; Parker, J.H.; Kaushik, T.; Toyofuku, M.S.; Tsuchiya, M. Molecular and Morphological Taxonomy of Living Ammonia and Related Taxa (Foraminifera) and Their Biogeography. Micropaleontology 2021, 67, 109–313. [Google Scholar]
- Hayward, B.W.; Holzmann, M.; Grenfell, H.R.; Pawlowski, J.; Triggs, C.M. Morphological Distinction of Molecular Types in Ammonia—towards a Taxonomic Revision of the World’s Most Commonly Misidentified Foraminifera. Mar. Micropaleontol. 2004, 50, 237–271. [Google Scholar] [CrossRef]
- Holzmann, M.; Pawlowski, J. Taxonomic Relationships in the Genus Ammonia (Foraminifera) Based on Ribosomal DNA Sequences. J. Micropalaeontol. 2000, 19, 11. [Google Scholar] [CrossRef] [Green Version]
- Richirt, J.; Schweizer, M.; Mouret, A.; Quinchard, S.; Saad, S.A.; Bouchet, V.M.P.; Wade, C.M.; Jorissen, F.J. Biogeographic Distribution of Three Phylotypes (T1, T2 and T6) of Ammonia (Foraminifera, Rhizaria) around Great Britain: New Insights from Combined Molecular and Morphological Recognition. J. Micropalaeontol. 2021, 40, 61–74. [Google Scholar] [CrossRef]
- Alve, E. Benthic Foraminiferal Responses to Estuarine Pollution; a Review. J. Foraminifer. Res. 1995, 25, 190–203. [Google Scholar] [CrossRef]
- Bouchet, V.M.P.; Debenay, J.-P.; Sauriau, P.-G.; Radford-Knoery, J.; Soletchnik, P. Effects of Short-Term Environmental Disturbances on Living Benthic Foraminifera during the Pacific Oyster Summer Mortality in the Marennes-Oléron Bay (France). Mar. Environ. Res. 2007, 64, 358–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armynot du Châtelet, E.; Debenay, J.-P.; Soulard, R. Foraminiferal Proxies for Pollution Monitoring in Moderately Polluted Harbors. Environ. Pollut. 2004, 127, 27–40. [Google Scholar] [CrossRef]
- Mojtahid, M.; Geslin, E.; Coynel, A.; Gorse, L.; Vella, C.; Davranche, A.; Zozzolo, L.; Blanchet, L.; Bénéteau, E.; Maillet, G. Spatial Distribution of Living (Rose Bengal Stained) Benthic Foraminifera in the Loire Estuary (Western France). J. Sea Res. 2016, 118, 1–16. [Google Scholar] [CrossRef]
- Nikulina, A.; Polovodova, I.; Schönfeld, J. Foraminiferal Response to Environmental Changes in Kiel Fjord, SW Baltic Sea. eEarth 2008, 3, 37–49. [Google Scholar] [CrossRef] [Green Version]
- Cushman, J.A. Recent Foraminifera from Porto Rico; Carnegie Institution of Washington: Washington, DC, USA, 1926; Volume 342. [Google Scholar]
- Richirt, J.; Schweizer, M.; Bouchet, V.M.P.; Mouret, A.; Quinchard, S.; Jorissen, F.J. Morphological Distinction of Three Ammonia Phylotypes Occurring Along European Coasts. J. Foraminifer. Res. 2019, 49, 76–93. [Google Scholar] [CrossRef]
- Rueden, C.T.; Schindelin, J.; Hiner, M.C.; De Zonia, B.E.; Walter, A.E.; Arena, E.T.; Eliceiri, K.W. ImageJ2: ImageJ for the next Generation of Scientific Image Data. BMC Bioinform. 2017, 18, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Petersen, J.; Riedel, B.; Barras, C.; Pays, O.; Guihéneuf, A.; Mabilleau, G.; Schweizer, M.; Meysman, F.J.R.; Jorissen, F.J. Improved Methodology for Measuring Pore Patterns in the Benthic Foraminiferal Genus Ammonia. Mar. Micropaleontol. 2016, 128, 1–13. [Google Scholar] [CrossRef]
- Bernhard, J.M.; Ostermann, D.R.; Williams, D.S.; Blanks, J.K. Comparison of Two Methods to Identify Live Benthic Foraminifera: A Test between Rose Bengal and CellTracker Green with Implications for Stable Isotope Paleoreconstructions. Paleoceanogr. Paleoclimatology 2006, 21, PA4210. [Google Scholar] [CrossRef]
- Schönfeld, J.; Alve, E.; Geslin, E.; Jorissen, F.; Korsun, S.; Spezzaferri, S. The FOBIMO (FOraminiferal BIo-MOnitoring) Initiative—Towards a Standardised Protocol for Soft-Bottom Benthic Foraminiferal Monitoring Studies. Mar. Micropaleontol. 2012, 94–95, 1–13. [Google Scholar] [CrossRef]
- Schweizer, M.; Polovodova, I.; Nikulina, A.; Schönfeld, J. Molecular Identification of Ammonia and Elphidium Species (Foraminifera, Rotaliida) from the Kiel Fjord (SW Baltic Sea) with RDNA Sequences. Helgol. Mar. Res. 2011, 65, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Pawlowski, J.; Holzmann, M. Diversity and Geographic Distribution of Benthic Foraminifera: A Molecular Perspective. Biodivers. Conserv. 2008, 17, 317–328. [Google Scholar] [CrossRef] [Green Version]
- Polovodova, I.; Nikulina, A.; Schönfeld, J.; Dullo, W.-C. Recent Benthic Foraminifera in the Flensburg Fjord (Western Baltic Sea). J. Micropalaeontol. 2009, 28, 131–142. [Google Scholar] [CrossRef] [Green Version]
- Bird, C.; Schweizer, M.; Roberts, A.; Austin, W.E.N.; Knudsen, K.L.; Evans, K.M.; Filipsson, H.L.; Sayer, M.D.J.; Geslin, E.; Darling, K.F. The Genetic Diversity, Morphology, Biogeography, and Taxonomic Designations of Ammonia (Foraminifera) in the Northeast Atlantic. Mar. Micropaleontol. 2020, 155, 101726. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pavard, J.-C.; Richirt, J.; Courcot, L.; Bouchet, P.; Seuront, L.; Bouchet, V.M.P. Fast and Reliable Identification of Ammonia Phylotypes T1, T2 and T6 Using a Stereomicroscope: Implication for Large-Scale Ecological Surveys and Monitoring Programs. Water 2021, 13, 3563. https://doi.org/10.3390/w13243563
Pavard J-C, Richirt J, Courcot L, Bouchet P, Seuront L, Bouchet VMP. Fast and Reliable Identification of Ammonia Phylotypes T1, T2 and T6 Using a Stereomicroscope: Implication for Large-Scale Ecological Surveys and Monitoring Programs. Water. 2021; 13(24):3563. https://doi.org/10.3390/w13243563
Chicago/Turabian StylePavard, Jean-Charles, Julien Richirt, Lucie Courcot, Pascal Bouchet, Laurent Seuront, and Vincent M. P. Bouchet. 2021. "Fast and Reliable Identification of Ammonia Phylotypes T1, T2 and T6 Using a Stereomicroscope: Implication for Large-Scale Ecological Surveys and Monitoring Programs" Water 13, no. 24: 3563. https://doi.org/10.3390/w13243563