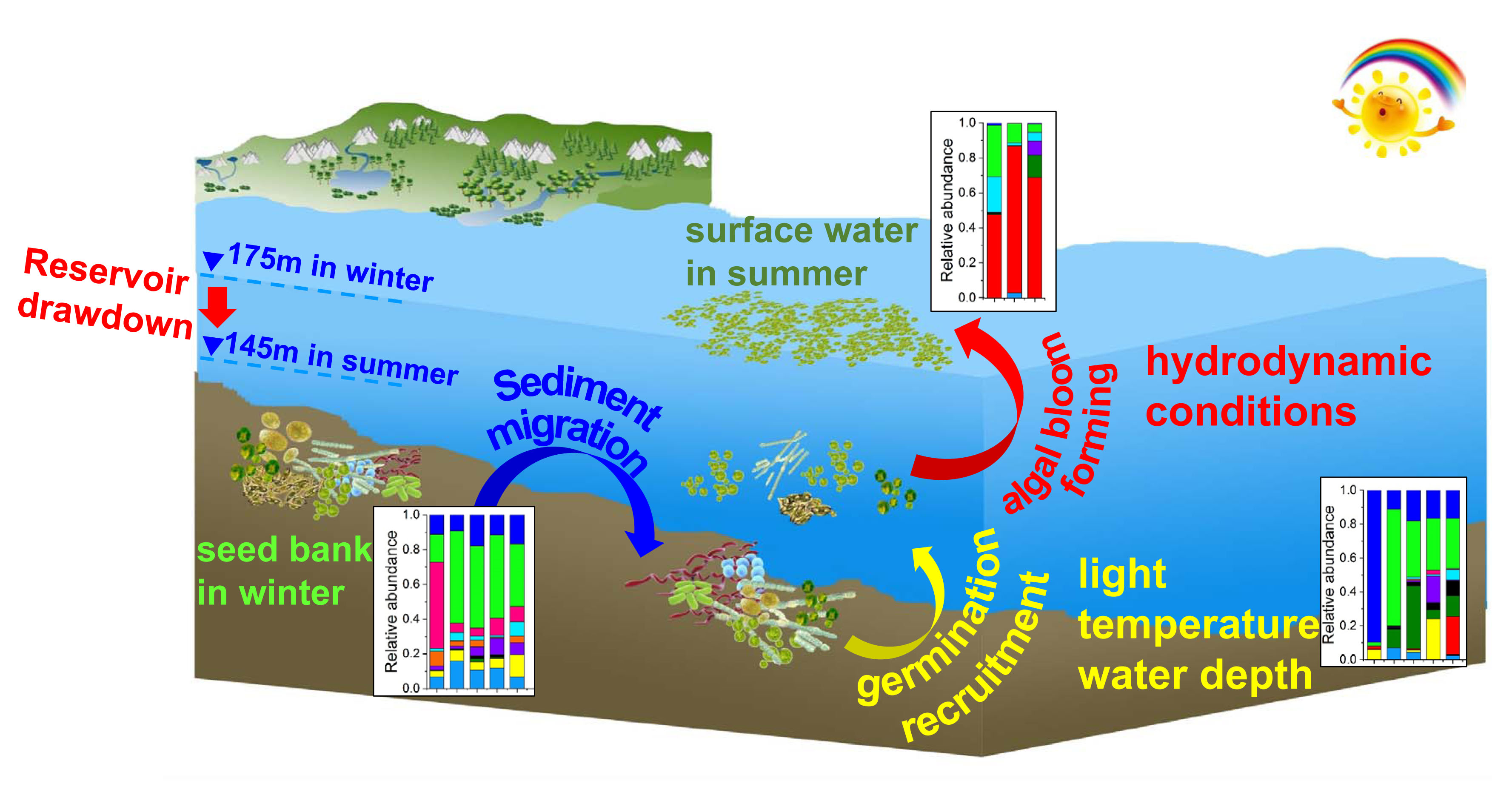
Spatio-Temporal Variations in Phytoplankton Communities in Sediment and Surface Water as Reservoir Drawdown—A Case Study of Pengxi River in Three Gorges Reservoir, China

Abstract
:
1. Introduction
2. Materials and Methods
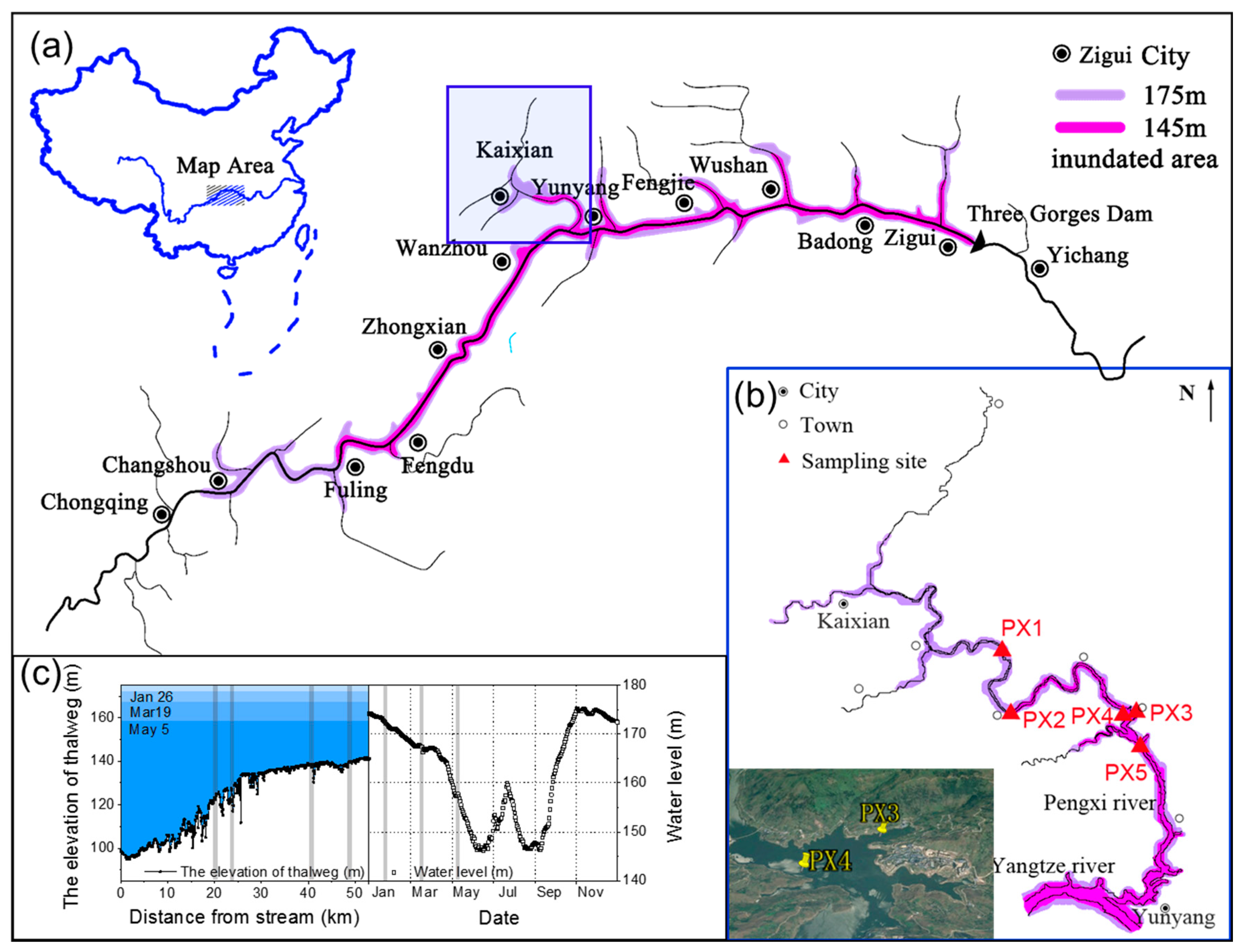
2.1. Study Area and Sample Collection
2.2. Analysis of Physicochemical Factors
2.3. DNA Extraction and Polymerase Chain Reaction (PCR)
2.4. Illumina MiSeq Sequencing and Analysis
2.5. Statistics Analysis
3. Results
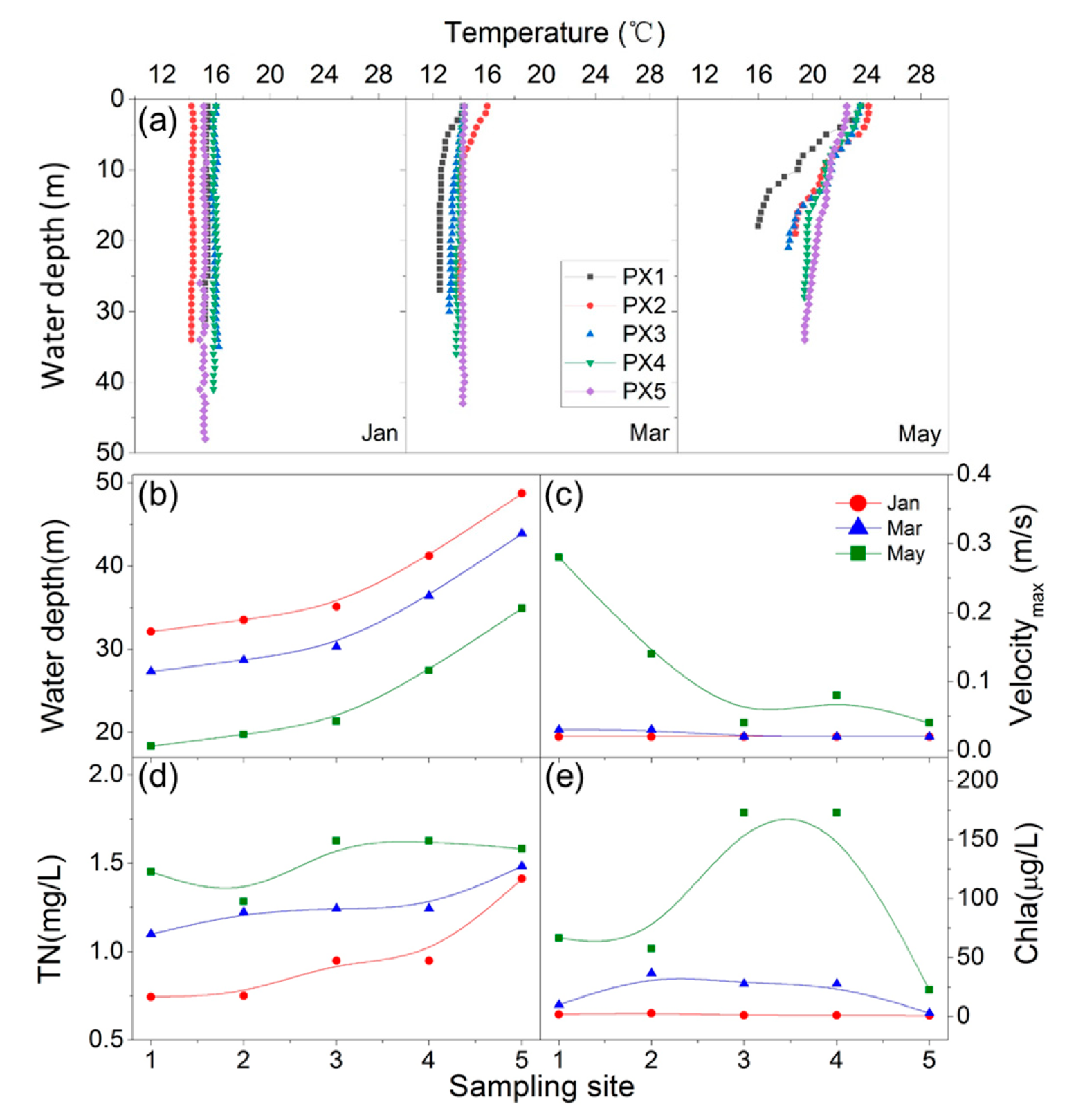
3.1. Environmental Parameters
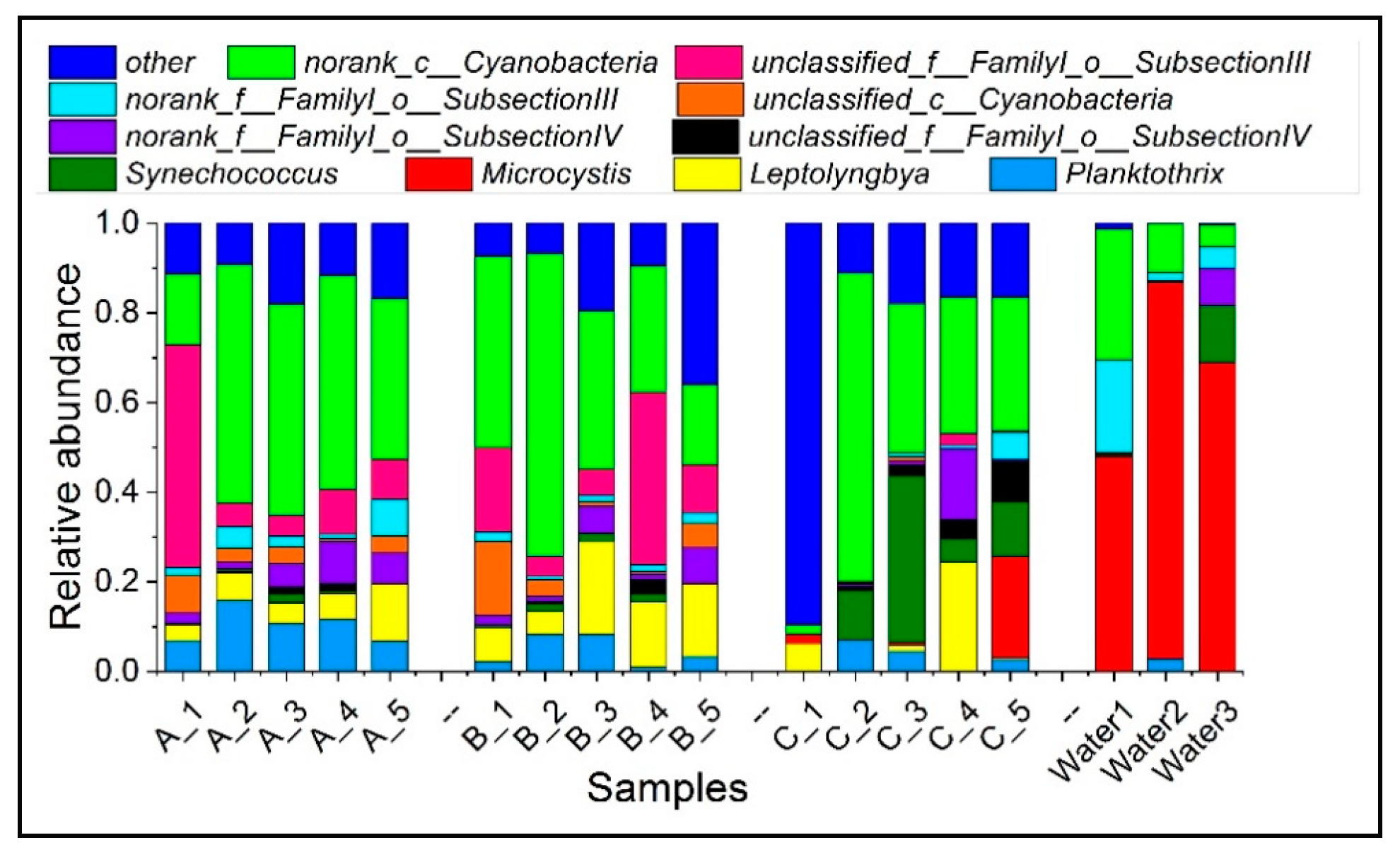
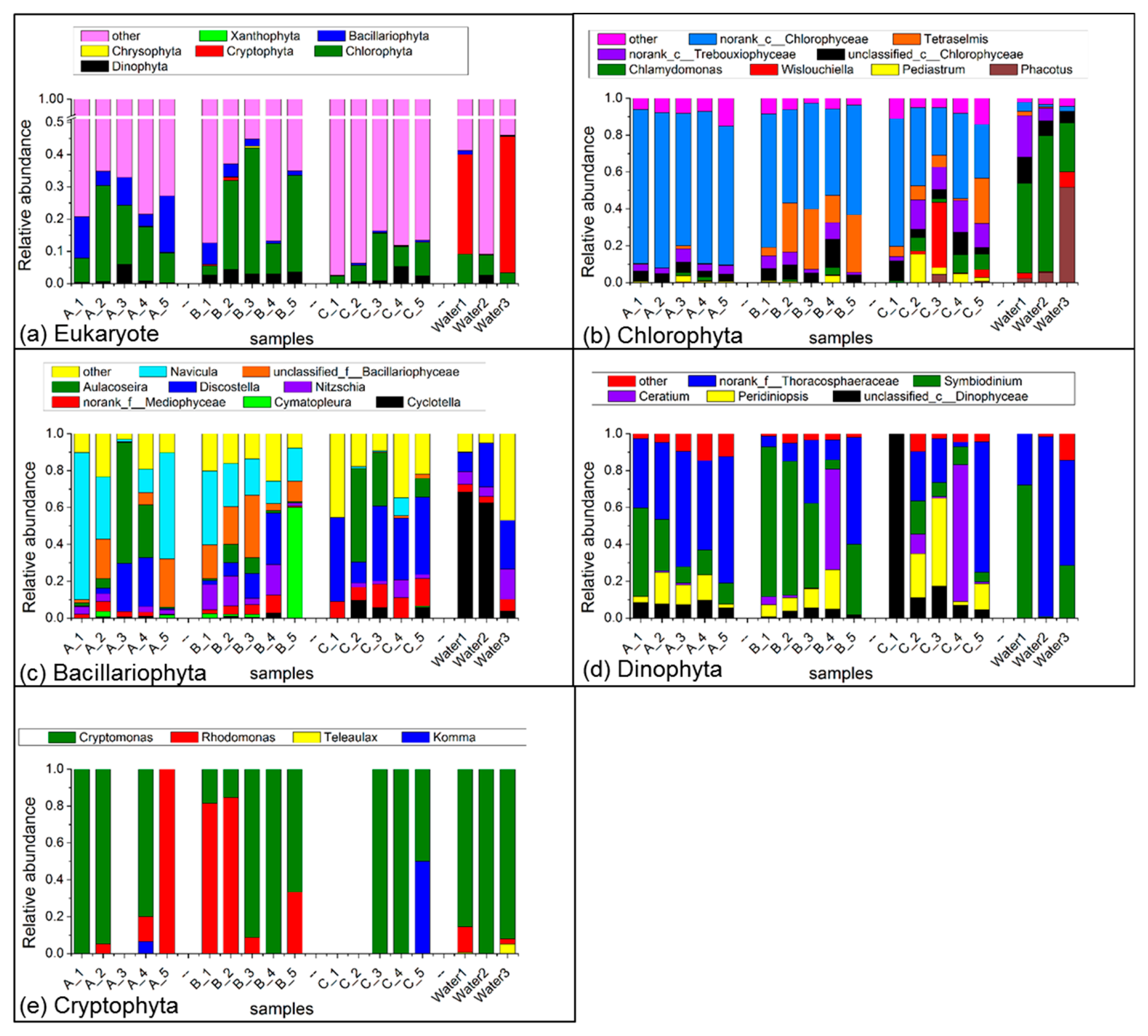
3.2. Phytoplankton Communities in Sediment
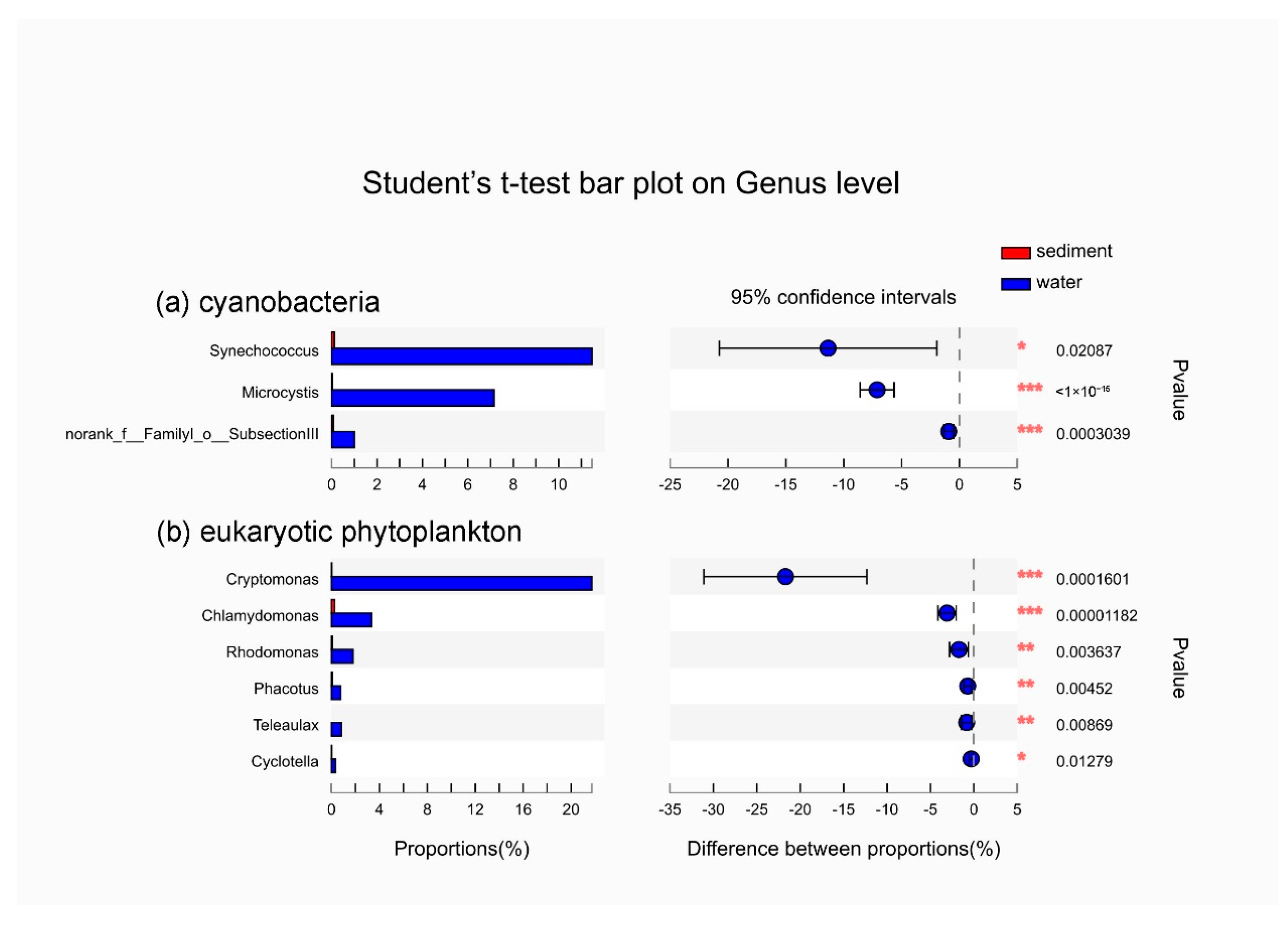
3.3. The Dominant Species of Phytoplankton in Sediment and Surface Water during Algal Blooms Period
4. Discussion
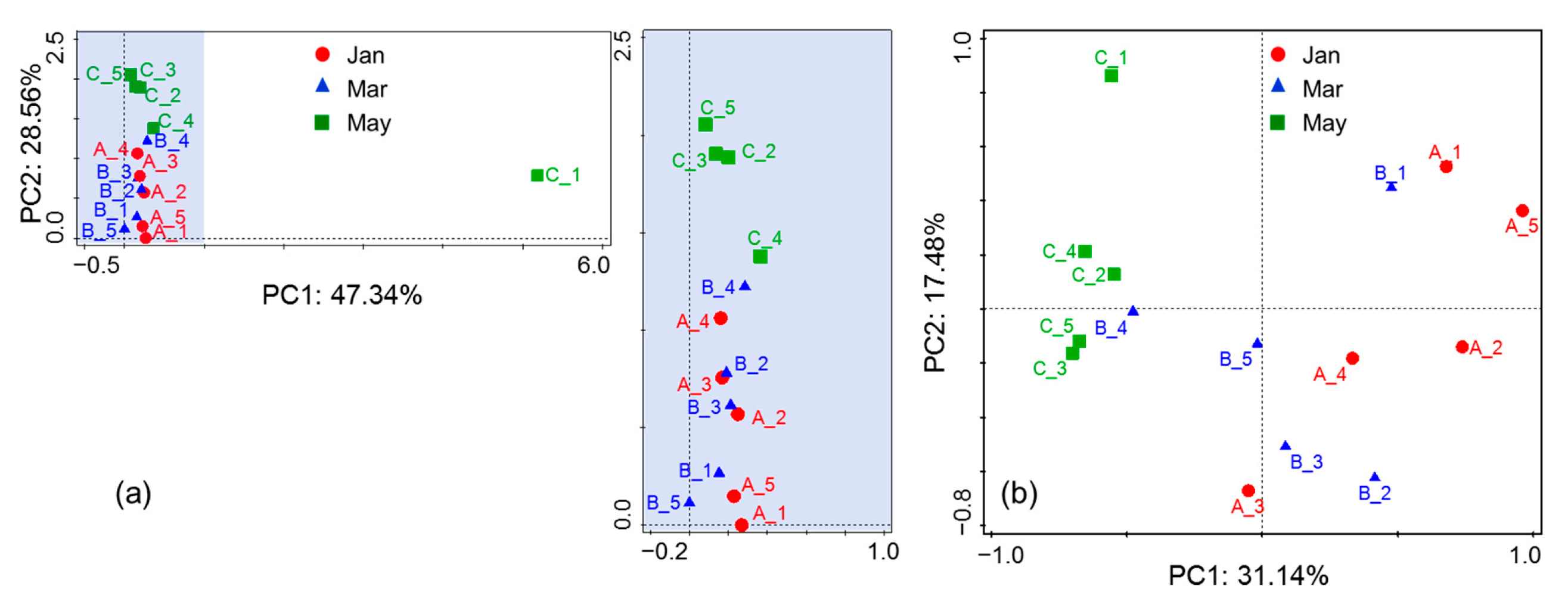
4.1. Variations in Phytoplankton Communities in Sediment
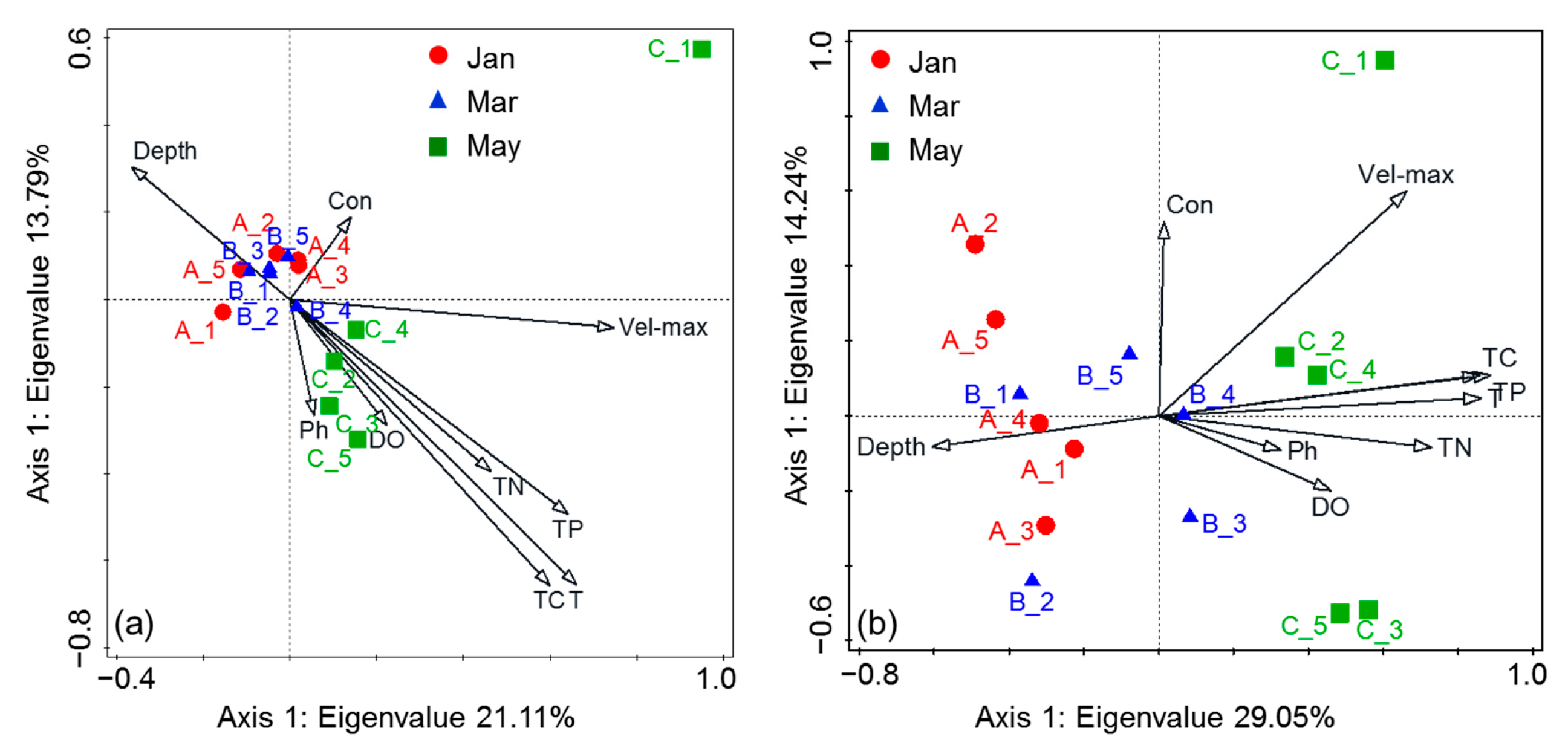
4.2. The Factor Influencing Species in Surface Water during Algal Blooms
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hallegraeff, G.M. A review of harmful algal blooms and their apparent globle increase. Phycologia 1993, 32, 79–99. [Google Scholar] [CrossRef] [Green Version]
- Duan, H.; Ma, R.; Xu, X.; Kong, F.; Zhang, S.; Kong, W.; Hao, J.; Shang, L. Two-Decade Reconstruction of Algal Blooms in China’s Lake Taihu. Environ. Sci. Technol. 2009, 43, 3522–3528. [Google Scholar] [CrossRef] [PubMed]
- Falconer, I.R. An overview of problems caused by toxic blue-green algae (cyanobacteria) in drinking and recreational water. Environ. Toxicol. 1999, 14, 5–12. [Google Scholar] [CrossRef]
- Lambert, T.W.; Holmes, C.F.B.; Hrudey, S.E. Microcystin class of toxins: Health effects and safety of drinking water supplies. Environ. Rev. 1994, 2, 167–186. [Google Scholar] [CrossRef]
- Seegers, B.N.; Birch, J.M.; Marin, R.; Scholin, C.A.; Caron, D.A.; Seubert, E.L.; Howard, M.D.A.; Robertson, G.L.; Jones, B.H. Subsurface seeding of surface harmful algal blooms observed through the integration of autonomous gliders, moored environmental sample processors, and satellite remote sensing in southern California. Limnol. Oceanogr. 2015, 60, 754–764. [Google Scholar] [CrossRef] [Green Version]
- Verspagen, J.M.H.; Snelder, E.O.F.M.; Visser, P.M.; Huisman, J.; Mur, L.R.; Ibelings, B.W. Recruitment of benthic microcystis (cyanophyceae) to the water column: Internal buoyancy changes or resuspension? J. Phycol. 2004, 40, 260–270. [Google Scholar] [CrossRef]
- Boero, F.; Belmonte, G.; Fanelli, G.; Piraino, S.; Rubino, F. The continuity of living matter and the discontinuities of its constituents: Do plankton and benthos really exist? Trends Ecol. Evol. 1996, 11, 177–180. [Google Scholar] [CrossRef]
- Karlsson-Elfgren, I.; Rengefors, K.; Gustafsson, S. Factors regulating recruitment from the sediment to the water column in the bloom-forming cyanobacterium Gloeotrichia echinulata. Freshw. Biol. 2004, 49, 265–273. [Google Scholar] [CrossRef] [Green Version]
- Head, R.M.; Jones, R.I.; Bailey-Watts, A.E. An assessment of the influence of recruitment from the sediment on the development of planktonic populations of cyanobacteria in a temperate mesotrophic lake. Freshw. Biol. 1999, 41, 759–769. [Google Scholar] [CrossRef]
- Anderson, D.M.; Chisholm, S.W.; Watras, C.J. Importance of life cycle events in the population dynamics of Gonyaulax tamarensis. Mar. Biol. 1983, 76, 179–189. [Google Scholar] [CrossRef]
- Anglès, S.; Garcés, E.; Reñé, A.; Sampedro, N. Life-cycle alternations in Alexandrium minutum natural populations from the NW Mediterranean Sea. Harmful Algae 2012, 16, 1–11. [Google Scholar] [CrossRef]
- McQuoid, M.R.; Godhe, A. Recruitment of coastal planktonic diatoms from benthic versus pelagic cells: Variations in bloom development and species composition. Limnol. Oceanogr. 2004, 49, 1123–1133. [Google Scholar] [CrossRef]
- Hansson, L.-A. Algal recruitment from lake sediments in relation to grazing, sinking, and dominance patterns in the phytoplankton community. Limnol. Oceanogr. 1996, 41, 1312–1323. [Google Scholar] [CrossRef]
- Stahl-Delbanco, A.; Hansson, L.A. Effects of bioturbation on recruitment of algal cells from the "seed bank" of lake sediments. Limnol. Oceanogr. 2002, 47, 1836–1843. [Google Scholar] [CrossRef]
- Rengefors, K.; Gustafsson, S.; Ståhl-Delbanco, A. Factors regulating the recruitment of cyanobacterial and eukaryotic phytoplankton from littoral and profundal sediments. Aquat. Microb. Ecol. 2004, 36, 213–226. [Google Scholar] [CrossRef]
- Brunberg, A.K.; Blomqvist, P. Recruitment of Microcystis (Cyanophyceae) from lake sediments: The importance of littoral inocula. J. Phycol. 2003, 39, 58–63. [Google Scholar] [CrossRef]
- Karlsson-Elfgren, I.; Brunberg, A.-K. The importance of shallow sediments in the recruitment of anabaena and aphanizomenon (cyanophyceae). J. Phycol. 2004, 40, 831–836. [Google Scholar] [CrossRef]
- Misson, B.; Latour, D. Influence of light, sediment mixing, temperature and duration of the benthic life phase on the benthic recruitment of Microcystis. J. Plankton Res. 2011, 34, 113–119. [Google Scholar] [CrossRef]
- Schöne, K.; Jähnichen, S.; Ihle, T.; Ludwig, F.; Benndorf, J. Arriving in better shape: Benthic Microcystis as inoculum for pelagic growth. Harmful Algae 2010, 9, 494–503. [Google Scholar] [CrossRef]
- Brosnahan, M.L.; Ralston, D.K.; Fischer, A.D.; Solow, A.R.; Anderson, D.M. Bloom termination of the toxic dinoflagellate Alexandrium catenella: Vertical migration behavior, sediment infiltration, and benthic cyst yield. Limnol. Oceanogr. 2017, 62, 2829–2849. [Google Scholar] [CrossRef] [Green Version]
- Ramm, J.; Rücker, J.; Knie, M.; Nixdorf, B. Lost in the dark: Estimation of the akinete pool for the recruitment of Nostocales populations (cyanobacteria) in a temperate deep lake. J. Plankton Res. 2017, 39, 392–403. [Google Scholar] [CrossRef] [Green Version]
- Villena, M.-J.; Romo, S. Effects of Nutrients, Fish, Charophytes and Algal Sediment Recruitment on the Phytoplankton Ecology of a Shallow Lake. Int. Rev. Hydrobiol. 2007, 92, 626–639. [Google Scholar] [CrossRef]
- Borges, H.; Wood, S.A.; Puddick, J.; Blaney, E.; Hawes, I.; Dietrich, D.R.; Hamilton, D.P. Intracellular, environmental and biotic interactions influence recruitment of benthicMicrocystis(Cyanophyceae) in a shallow eutrophic lake. J. Plankton Res. 2016, 38, 1289–1301. [Google Scholar] [CrossRef] [Green Version]
- Tan, X.; Kong, F.; Zhang, M.; Yü, Y. Recruitment of Phytoplankton from Winter Sediment of Lake Taihu: A Laboratory Simulation. J. Freshw. Ecol. 2009, 24, 339–341. [Google Scholar] [CrossRef] [Green Version]
- Cao, H.-S.; Tao, Y.; Kong, F.-X.; Yang, Z. Relationship between Temperature and Cyanobacterial Recruitment from Sediments in Laboratory and Field Studies. J. Freshw. Ecol. 2008, 23, 405–412. [Google Scholar] [CrossRef] [Green Version]
- Carey, C.C.; Weathers, K.C. Spatial and temporal variability in recruitment of the cyanobacterium Gloeotrichia echinulata in an oligotrophic lake. Freshw. Sci. 2014, 33, 577–592. [Google Scholar] [CrossRef]
- Feng, B.; Wang, C.; Wu, X.; Tian, C.; Tian, Y.; Xiao, B. Involvement of microcystins, colony size and photosynthetic activity in the benthic recruitment of Microcystis. Environ. Biol. Fishes 2019, 31, 223–233. [Google Scholar] [CrossRef]
- Zou, W.; Wang, Z.; Song, Q.; Tang, S.; Peng, Y. Recruitment-promoting of dormant Microcystis aeruginosa by three benthic bacterial species. Harmful Algae 2018, 77, 18–28. [Google Scholar] [CrossRef]
- Karlson, A.M.; Nascimento, F.J.A.; Suikkanen, S.; Elmgrenc, R. Benthic fauna affects recruitment from sediments of the harmful cyanobacterium Nodularia spumigena. Harmful Algae 2012, 20, 126–131. [Google Scholar] [CrossRef]
- Hamre, K.D.; Gerling, A.; Munger, Z.; Doubek, J.; McClure, R.; Cottingham, K.L.; Carey, C. Spatial variation in dinoflagellate recruitment along a reservoir ecosystem continuum. J. Plankton Res. 2017, 39, 715–728. [Google Scholar] [CrossRef]
- Su, Y.; You, X.; Lin, H.; Zhuang, H.; Weng, Y.; Zhang, D. Recruitment of cyanobacteria from the sediments in the eutrophic Shanzi Reservoir. Environ. Technol. 2015, 37, 641–651. [Google Scholar] [CrossRef] [PubMed]
- Stone, R.; Jia, H. Hydroengineering: Going Against the Flow. Science 2006, 313, 1034–1037. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, C.; Reidy, C.A. Fragmentation and flow regulation of the world’s large river systems. Science 2005, 308, 405–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stone, R. China’s environmental challenges: Three gorges dam: Into the unknown. Science 2008, 321, 628–632. [Google Scholar] [CrossRef]
- Becker, V.; Huszar, V.L.M.; Naselli-Flores, L.; Padisák, J. Phytoplankton equilibrium phases during thermal stratification in a deep subtropical reservoir. Freshw. Biol. 2008, 53, 952–963. [Google Scholar] [CrossRef]
- Conley, D.J.; Stålnacke, P.; Pitkänen, H.; Wilander, A. The transport and retention of dissolved silicate by rivers in Sweden and Finland. Limnol. Oceanogr. 2000, 45, 1850–1853. [Google Scholar] [CrossRef]
- Wu, J.G.; Huang, J.H. Three-Gorges Dam—Experiment in habitat fragmentation? Science 2003, 300, 1239–1240. [Google Scholar] [CrossRef]
- Huang, Z.L.; Wu, B.F. The overview of the project and environment. In Research on Ecological and Environmental Monitoring System of the Three Gorges Project, 1st ed.; Li, F., Gai, Y., Eds.; Science Press: Beijing, China, 2006. [Google Scholar]
- Zhou, G.J.; Kuang, Q.J. Investigation of algal water-blooms in the Three Gorges Reservoir and its toxicological study. Acta Hydrobiol. Sin. 2006, 1, 37. [Google Scholar]
- Zheng, B.H.; Cao, Y.J. Analysis of Algal Blooms in Da-ning River of Three Gorges Reservoir. Environ. Sci. 2009, 30, 3218. [Google Scholar]
- Zhang, S.; Xiao, Y.; Li, Z.; Wang, S.; Guo, J.; Lu, L. Turbulence exerts nutrients uptake and assimilation of bloom-forming Dolichospermum through modulating morphological traits: Field and chemostat culture studies. Sci. Total. Environ. 2019, 671, 329–338. [Google Scholar] [CrossRef]
- Ran, X.; Bouwman, A.F.; Yu, Z.; Liu, J. Implications of eutrophication for biogeochemical processes in the Three Gorges Reservoir, China. Reg. Environ. Chang. 2019, 19, 55–63. [Google Scholar] [CrossRef]
- Liu, X.; Qian, K.; Chen, Y. Effects of water level fluctuations on phytoplankton in a Changjiang River floodplain lake (Poyang Lake): Implications for dam operations. J. Great Lakes Res. 2015, 41, 770–779. [Google Scholar] [CrossRef]
- Li, Z.; Guo, J.S. 1D hydrodynamic model on Pengxi (Xiaojiang) River backwater area in the Three Gorges Reservoir. J. Chongqing Uni. 2012, 5, 143. [Google Scholar]
- Xiao, Y.; Li, Z.; Guo, J.; Fang, F.; Smith, V.H. Succession of phytoplankton assemblages in response to large-scale reservoir operation: A case study in a tributary of the Three Gorges Reservoir, China. Environ. Monit. Assess. 2016, 188, 153. [Google Scholar] [CrossRef]
- Bråte, J.; Logares, R.; Berney., C.; Ree, D.K.; Klaveness, D.; Jakobsen, K.S.; Shalchian-Tabrizi, K. Freshwater Perkinsea and marine-freshwater colonizations revealed by pyrosequencing and phylogeny of environmental rDNA. ISME J. 2010, 4, 1144–1153. [Google Scholar] [CrossRef] [Green Version]
- Zhou, C.; Yu, J.-J.; Fu, L.; Cui, Y.-J.; Liu, D.; Jiang, W.; Haffner, D.; Zhang, L. Temporal and Spatial Distribution of Environmental Factors and Phytoplankton During Algal Bloom Season in Pengxi River, Three Gorges Reservoir. Environ. Sci. 2016, 37, 873–883. [Google Scholar]
- Huang, Y.; Yasarer, L.M.W.; Li, Z.; Sturm, B.S.M.; Zhang, Z.; Guo, J.; Shen, Y. Air–water CO2 and CH4 fluxes along a river–reservoir continuum: Case study in the Pengxi River, a tributary of the Yangtze River in the Three Gorges Reservoir, China. Environ. Monit. Assess. 2017, 189, 223. [Google Scholar] [CrossRef]
- Shin, H.H.; Li, Z. Seasonal production of dinoflagellate cysts in relation to environmental characteristics ih jiiihae-Masan Bay, Korea: One-year sedimerit trap observation. Estuar. Coast. Shelf Sci. 2018, 215, 83–93. [Google Scholar] [CrossRef]
- Rolland, D.C.; Vincent, W.F. Characterization of phytoplankton seed banks in the sediments of a drinking water reservoir. Lake Reserv. Manag. 2014, 30, 371–380. [Google Scholar] [CrossRef]
- Valdespino-Castillo, P.M.; Merino-Ibarra, M.; Jiménez-Contreras, J.; Castillo-Sandoval, F.S.; Ramírez-Zierold, J.A. Community metabolism in a deep (stratified) tropical reservoir during a period of high water-level fluctuations. Environ. Monit. Assess. 2014, 186, 6505–6520. [Google Scholar] [CrossRef] [Green Version]
- Jäger, C.G.; Diehl, S.; Schmidt, G.M. Influence of water-column depth and mixing on phytoplankton biomass, community composition, and nutrients. Limnol. Oceanogr. 2008, 53, 2361–2373. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Liu, D.; Johnson, D.M.; Yi, Z.; Huang, Y. Effects of vertical mixing on phytoplankton blooms in Xiangxi Bay of Three Gorges Reservoir: Implications for management. Water Res. 2012, 46, 2121–2130. [Google Scholar] [CrossRef] [PubMed]
- Oliver, R.L.; Ganf, G.G. Freshwater Blooms. In The Ecology of Cyanobacteria II: Their Diversity in Time and Space, 1st ed.; Whitton, B.A., Potts, M., Eds.; Springer: Dordrecht, The Netherlands, 2002; pp. 149–194. [Google Scholar]
- Li, Z.; Fang, F. Spring algal bloom and nutrients characteristics in Xiaojiang River backwater area, Three Gorges Reservoir. J. Lake Sci. 2009, 21, 36. [Google Scholar]
- Ståhl-Delbanco, A.; Hansson, L.; Gyllström, M. Recruitment of resting stages may induce blooms of Microcystis at low N: P ratios. J. Plankton Res. 2003, 25, 1099–1106. [Google Scholar] [CrossRef]
- Xiao, M.; Li, M.; Reynolds, C.S. Colony formation in the cyanobacteriumMicrocystis. Biol. Rev. 2018, 93, 1399–1420. [Google Scholar] [CrossRef] [Green Version]
- Chia National Environmental Monitoring Center. Available online: http://www.cnemc.cn/jcbg/zjsxgcstyhjjcbg/ (accessed on 18 December 2020).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Time | Sampling Site | T (°C) | Conductivity (μS/cm) | pH | DO (mg/L) | TC (mg/L) | TOC (mg/L) | IC (mg/L) | TN (mg/L) | TP (mg/L) | Chla (μg/L) | Average Flow Velocity (m/s) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| January | PX1 | 15.4 | 388.6 | 7.87 | 7.89 | 37.98 | 1.31 | 36.68 | 0.745 | 0.024 | 1.78 | 0.02 |
| PX2 | 14.2 | 473.0 | 8.36 | 7.71 | 32.09 | 3.17 | 28.92 | 0.752 | 0.029 | 2.63 | 0.02 | |
| PX3 | 16.0 | 365.7 | 7.80 | 6.46 | 30.87 | 3.40 | 27.47 | 0.949 | 0.041 | 0.93 | 0.02 | |
| PX4 | 16.0 | 365.7 | 7.80 | 6.46 | 30.87 | 3.40 | 27.47 | 0.949 | 0.041 | 0.93 | 0.02 | |
| PX5 | 15.1 | 359.6 | 8.38 | 7.90 | 30.14 | 2.51 | 27.63 | 1.413 | 0.045 | 0.70 | 0.02 | |
| March | PX1 | 14.2 | 448.5 | 8.57 | 9.95 | 33.79 | 5.31 | 28.49 | 1.100 | 0.036 | 9.94 | 0.02 |
| PX2 | 16.0 | 437.1 | 7.69 | 15.56 | 33.31 | 7.49 | 25.53 | 1.224 | 0.032 | 36.62 | 0.02 | |
| PX3 | 14.3 | 382.3 | 8.18 | 10.96 | 32.55 | 6.58 | 25.97 | 1.244 | 0.047 | 27.69 | 0.02 | |
| PX4 | 14.3 | 382.3 | 8.18 | 10.96 | 32.55 | 6.58 | 25.97 | 1.244 | 0.047 | 27.69 | 0.02 | |
| PX5 | 14.3 | 379.1 | 8.15 | 9.26 | 31.29 | 3.66 | 27.63 | 1.484 | 0.048 | 2.82 | 0.01 | |
| May | PX1 | 23.6 | 471.0 | 7.68 | 10.08 | 68.39 | 8.07 | 60.33 | 1.451 | 0.127 | 66.66 | 0.07 |
| PX2 | 24.1 | 410.7 | 8.28 | 12.41 | 79.54 | 9.01 | 70.54 | 1.284 | 0.087 | 57.59 | 0.05 | |
| PX3 | 23.5 | 395.5 | 9.69 | 15.80 | 61.15 | 15.17 | 45.97 | 1.627 | 0.142 | 172.86 | 0.01 | |
| PX4 | 23.5 | 395.5 | 9.69 | 15.80 | 61.15 | 15.17 | 45.97 | 1.627 | 0.142 | 172.86 | 0.02 | |
| PX5 | 22.5 | 372.5 | 7.73 | 7.29 | 66.79 | 6.46 | 60.34 | 1.582 | 0.085 | 22.72 | 0.01 |
| Cyanobacteria | Eukaryotic Phytoplankton | |||||||
|---|---|---|---|---|---|---|---|---|
| Sediment | Water | Sediment | Water | |||||
| Sampling site | January | March | May | May | January | March | May | May |
| PX1 | 11.91% | 6.06% | 0.18% | 9.18% | 20.57% | 10.22% | 2.62% | 41.30% |
| PX2 | 1.37% | 4.68% | 2.17% | 7.93% | 34.57% | 33.97% | 6.20% | 9.14% |
| PX3 | 3.23% | 2.41% | 2.44% | 56.93% | 32.38% | 43.45% | 16.29% | 45.99% |
| PX4 | 0.67% | 1.80% | 0.36% | - 1 | 21.42% | 13.06% | 11.33% | - 1 |
| PX5 | 2.22% | 1.26% | 2.46% | - 1 | 26.91% | 32.78% | 13.31% | - 1 |
| Algae Species | Temperature | Light | Depth of Water | Disturbance of Sediment | Biological Factors | Other Condition | Reference |
|---|---|---|---|---|---|---|---|
| Microcystis | higher than 20 °C | 2000 lx | positive effect | [31] | |||
| Oscillatoria | higher than 20 °C | positive effect | |||||
| Chlorophytes and Diatoms | higher than 9 °C | [24] | |||||
| Cyanobacteria | higher than 12.5 °C | ||||||
| Microcystis and Aphanizomenon gracile | 16–25 °C | between 50 and 100 μmol m−2 s−1 | allopathic interactions, ammonium | [23] | |||
| Dolichospermum; Aphanizomenon; and Cylindrospermopsis raciborskii | 8–12 °C | >0.1 mol m−2 d−1 | <12 m | oxic conditions | [21] | ||
| Microcystis aeruginosa, Viridis, Wesenbergii, Aphanizomenon flos-aquae, Anabaena Circinalis | 1–2 m was better than 6–7 m | [16,17] | |||||
| Microcystis | positive effect | photosynthetic activity, colony size, and intracellular microcystin content | [27] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ouyang, W.; Li, Z.; Yang, J.; Lu, L.; Guo, J. Spatio-Temporal Variations in Phytoplankton Communities in Sediment and Surface Water as Reservoir Drawdown—A Case Study of Pengxi River in Three Gorges Reservoir, China. Water 2021, 13, 340. https://doi.org/10.3390/w13030340
Ouyang W, Li Z, Yang J, Lu L, Guo J. Spatio-Temporal Variations in Phytoplankton Communities in Sediment and Surface Water as Reservoir Drawdown—A Case Study of Pengxi River in Three Gorges Reservoir, China. Water. 2021; 13(3):340. https://doi.org/10.3390/w13030340
Chicago/Turabian StyleOuyang, Wenjuan, Zhe Li, Jixiang Yang, Lunhui Lu, and Jinsong Guo. 2021. "Spatio-Temporal Variations in Phytoplankton Communities in Sediment and Surface Water as Reservoir Drawdown—A Case Study of Pengxi River in Three Gorges Reservoir, China" Water 13, no. 3: 340. https://doi.org/10.3390/w13030340