
The Role of Climate Changes in the Spread of Freshwater Fishes: Implications for Alien Cool and Warm-Water Species in a Mediterranean Basin




1
Department of Chemistry, Biology and Biotechnologies, University of Perugia, 06100 Perugia, Italy
2
Forest, Economics and Mountain Territory Service, Umbria Region, 06100 Perugia, Italy
*
Author to whom correspondence should be addressed.
Water 2021, 13(3), 347; https://doi.org/10.3390/w13030347
Submission received: 10 December 2020
/
Revised: 8 January 2021
/
Accepted: 25 January 2021
/
Published: 30 January 2021
(This article belongs to the Special Issue Effects of Climate Change on Freshwater Biodiversity)
Abstract
:In running waters, under climate change conditions, the combined effect of water warming and decreasing flow rates may encourage colonisation by invasive cool and warm-water fish species. The aim of the study was to analyze the potential climate change effects on the spread of four invasive alien fishes in the Tiber River basin, taking into account the effects of river fragmentation. Fish and environmental data collected in 91 sites over the years 1998–2018, were used to analyze temporal changes in their habitat requirements. A multivariate analysis was conducted, and the hypothesis of a range expansion towards the upstream reaches has been tested. For Barbus barbus, Gobio gobio, Padogobius bonelli and Pseudorasbora parva population abundances and body condition were analyzed. Detectability, occupancy, local extinction and colonization probabilities were estimated. We showed that B. barbus and P. bonelli have significantly extended their range toward upstream. P. parva did not move toward higher altitudes significantly, suggesting that, at this stage, the species has probably reached an equilibrium. River fragmentation, elevation, water temperature and average current speed seem to be major determinants in colonization processes, affecting the dispersal ability of the species. Not surprisingly for species introduced in relatively recent times, the colonization probabilities were much higher than extinction probabilities. Our results provided evidence for some synergistic effects between climate changes and alien fish species invasions, in terms of species range shifts mediated by rising water temperatures, although they should be interpreted cautiously, taking into account that these species most likely were not yet stabilized.
1. Introduction
The negative impacts of alien fish invasions and climate changes on freshwater ecosystems have been the subject of several investigations [1,2,3], although often these two aspects are treated separately, overlooking their interactions [4,5,6]. Relatively few studies have been attempted on the role of climate factors in the spread of non-native freshwater fishes [7]. The progressive rise in water temperature as a result of global warming may encourage colonization by invasive alien species whose expansion would otherwise be limited by low temperatures [4,7,8].
The Mediterranean area, due to the particular climatic and geographic conditions, the richness of endemism, the high number of introduced alien species, in addition to other anthropogenic pressures, is particularly vulnerable to climate changes and to the loss of biodiversity [9,10,11,12].
Mediterranean rivers, in particular, as a result of both the effects of climate changes and alien species invasions, are strongly affected by the decline in native fish biodiversity [10,13]. Being ectothermic organisms, fish are particularly sensitive to changes in temperature resulting from global warming [14]. In running waters, the combined effect of increasing water temperature and decreasing flow rates may result in changes in the fish distribution along the longitudinal gradient [15,16,17]. Especially in the summer season, cool- and warm-water fish species may be induced to move upstream, while the cold-water species, already located near the springs, can become isolated [6,16,18,19]. Moreover, in Mediterranean watercourses numerous cool- and warm-water species are of alien origin, and can be advantaged by rising temperatures [6,11]. Many of them are pollution tolerant and may be more resilient to the indirect effects of climate change (i.e., increase of the pollutants concentration and reduced dissolved oxygen as a consequence to the decreasing flow rates and rising temperatures) then native species.
Recent studies conducted on fish communities inhabiting Italian basins provided evidence for their gradual qualitative degradation over time due to the progressive expansion of alien species [20,21,22]. At present, 60% of the fish species occurring in Italian freshwaters are of exotic origin [23]. Previous research carried out on fish communities inhabiting the Tiber river basin (central Italy) have shown that most of alien freshwater fish have been introduced over the last 30 years through restocking activities in favor of recreational fishery [20]; they have been initially released in multiple sites, and subsequently expanded their range using the connections of the hydrographic network [24]. Some of these species, as Barbus barbus (Linnaeus, 1758), Gobio gobio (Linnaeus, 1758), Padogobius bonelli (Bonaparte, 1846) and Pseudorasbora parva (Temminck & Schlegel, 1846), immediately after their introduction occurred in the period between 1993 and 1999, quickly acclimatized and are currently present with many self-sustaining populations [25,26].
Worryingly, the spread of these invasive species has contributed to the local extinction or range shift of some endemic ones. In particular, it has been shown that B. barbus represents a serious threat to the native Italian congeneric species, mainly due to introgressive hybridization phenomena leading to the loss of genetic diversity [21,27,28,29,30,31]. In the Tiber river basin, B. barbus has determined the local extinction of numerous populations of the endemic Barbus tyberinus Bonaparte, 1839 [26]. Additionally, P. bonelli exerted heavy negative impacts on a congeneric species endemic to central Italy, Padogobius nigricans (Canestrini, 1867) [32]; in this case, the main mechanisms underlying the local extinction phenomena or population declines include the greater competitiveness of the alien species for environmental resources (i.e., food and habitat) compared to the native one. As regards P. parva, the species is listed among the worst alien species for Europe, according to its negative ecological effects exerted on the colonized environments [33,34]. Even in the Tiber basin, previous studies based on the analysis of relative weight [25,35] suggested that the presence of P. parva could have exerted a negative impact on the body condition of many native species, as Sarmarutilus rubilio (Bonaparte, 1837), B. tyberinus, Squalius squalus (Bonaparte, 1837) and Telestes muticellus (Bonaparte, 1837). For G. gobio, given the relatively recent introduction of the species, which took place in 1999, no ecological effects on the endemic species with which it was found in co-occurrence have yet been tested [24]. In accordance with the literature for temperate fish [36,37], on the basis of their thermal preferences, B. barbus, G. gobio and P. bonelli could be classified as cool-water species (optimum temperature ≈17 °C), while P. parva could be considered as warm-water adapted species (optimum temperature ≈22 °C).
As reported in the literature for other areas, the detrimental impacts exerted by these four alien species could be amplified by the synergistic effect with additional anthropogenic stressors, including global warming [2,38,39]. Generally, the ability to persist of introduced species is influenced by their dispersal capacity, environmental tolerance, and food and habitat availability [19]. In recent times, there have also been highlighted some effects of global climate change, in terms of rising water temperatures, which seem to encourage their further expansion and therefore their invasiveness [40]. Warmer waters would provide new suitable habitats in which warm-water adapted species could expand their range [4]. Even in central-Italian watercourses, the progressive increase in water temperature, extended drought periods, and altered stream flow pattern seem to penalize the endemic species forcing them to go upstream [41], while at the same time it is reasonable to hypothesize the range shift of cool and warm-water alien fishes toward headwaters. In the present paper we don’t analyze the climate change effects in terms of rising temperatures and decreasing precipitations because this aspect has already described in Carosi et al. [41].
Another aspect to be taken into account in relation to the spread of alien fish is the high river fragmentation affecting the Tiber river basin. Under global warming conditions, it is important to analyze the role of connectivity in the freshwater fish range shifts put in place in order to face rising water temperatures. In fact, interruptions in river continuity may on one hand restrict the expansion of alien species [20], and on the other may preclude native species from accessing more suitable habitats, often located in the upper stream reaches [42].
The main aim of the present study was to analyze the potential climate change effects on the spread of four invasive alien fishes in the Tiber River basin using a spatio-temporal approach and taking into account the effects of river fragmentation. On the basis of long-term fish and environmental data series, temporal changes in their habitat use were analyzed, and the hypothesis of a range expansion towards upstream reaches has been tested.
2. Materials and Methods
2.1. Study Area
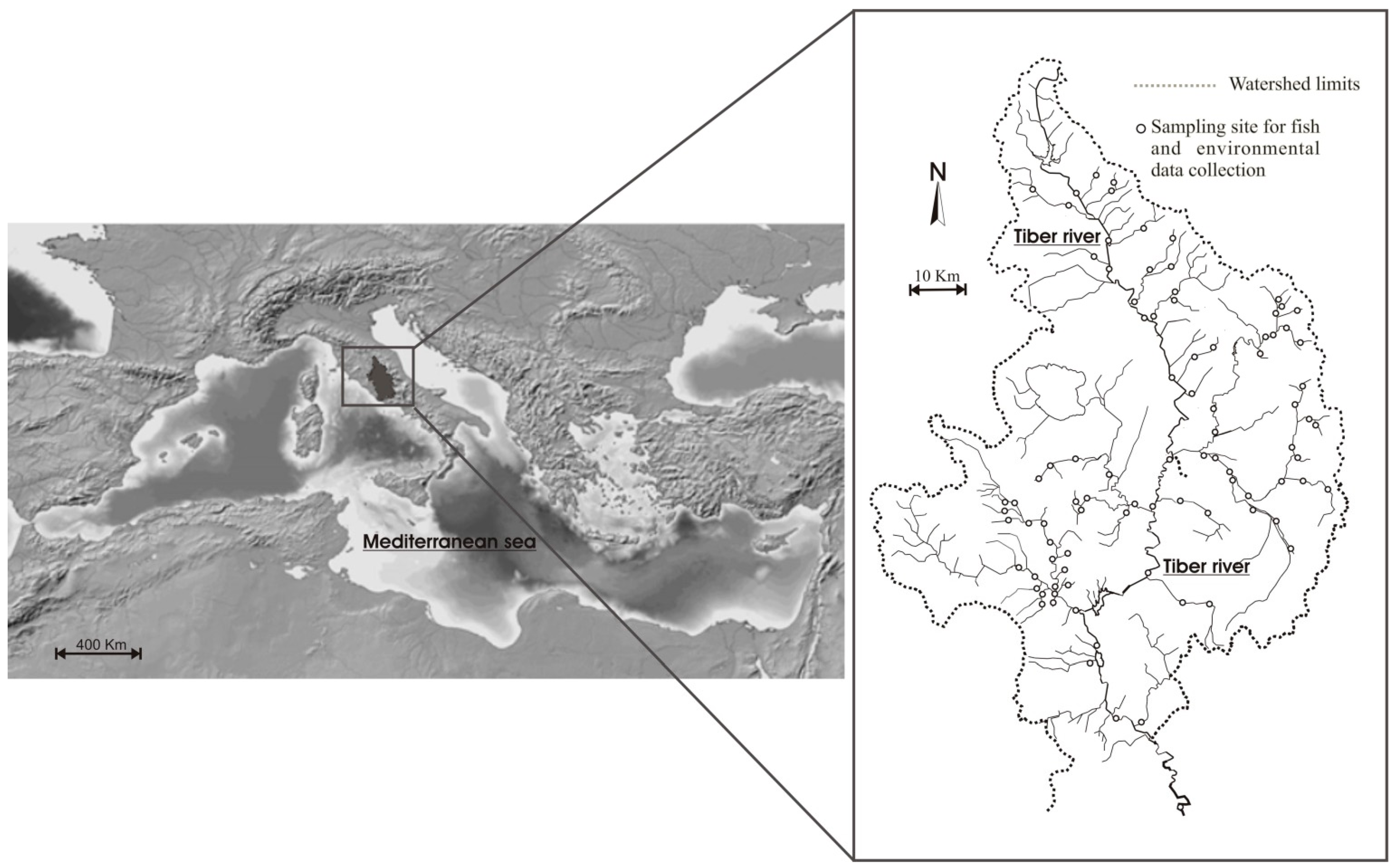
The study area includes the northern portion of the Tiber river basin, for a total extension of 5133 km−2, equal to 30% of the total catchment area (Figure 1). It is located within the Mediterranean region, where the climate is characterized by dry periods in summer and wet periods in winter [43]. These meteorological features make aquatic environments particularly vulnerable to potential climatic changes effects, in terms of rising water temperatures and decreasing precipitations [11,44]. In fact, the geological characteristics of the upper Tiber river basin, which is predominantly impermeable, make the flow rates of the watercourses closely linked to rainfalls [45]. The high-water demand for irrigation purposes contributes to aggravate the water shortage, particularly during drought periods. A recent study provided evidence for some effects of climate changes within the Tiber river basin, in terms of decreasing flow rates over the last 100 years, and increasing water temperatures over the last 15 years, with a magnitude of 0.4 °C [41]. As mentioned above, the whole hydrographic network is highly fragmented due to the presence of many weirs and dams [41].
2.2. Fish and Environmental Data Collection
Fish and environmental data were collected during three census periods, 1998–2004, 2005–2011 and 2012–2018, in 91 sampling sites spread over a total of 50 watercourses. Each sampling site was sampled once for each census period, in the autumn season. The fish fauna was sampled by two-pass electrofishing applying the removal method [46,47] (for more details, see [25]). All captured fishes were identified and counted in order to estimate the density (individual m‒2) for each population. For all specimens, total length (TL) and weight (W) were recorded. To characterize river stretches, 21 environmental parameters that play a crucial role in the distribution and life-history strategies of freshwater fish species were used. Conductivity (μS cm‒1), pH (units), water temperature (°C) and dissolved oxygen (mg L‒1) were measured during the field activities simultaneously with fish samplings, using electronic meters manufactured by YSI Incorporated (model 55/12, Yellow Springs, OH, USA), Hanna Instruments (HI9033, Woonsocket, RI, USA) and WTW GmbH (pH 320, Weilheim, Germany). Chlorides, sulphates, phosphates, total phosphorous, COD, NNO2, NNO3, ammonia were subsequently measured in the laboratory following the environmental protection agencies methods [48,49,50]. Hydraulic variables were determined at cross sections within each sampling stretch, using an OTT MF-pro electromagnetic current meter (OTT Hydromet, Kempten, Germany). Morphological parameters, as watershed area (km‒2), distance from the water source (km), and elevation (m a.s.l.) were calculated from digital maps (geographic information system (GIS)). The location of dams and weirs within the Tiber river basin was provided by the Forest, Economics and Mountain Territory Service of the Umbria Region, Perugia, Italy. The river fragmentation degree has been codified as the number of weirs with height > 80 cm occurring downstream of each sampling station, up to the main channel. To evaluate the water quality on the basis of the macrobenthic fauna composition, the Extended Biotic Index (EBI) and the IBE Quality class were used [51] (for more details, see [25]).
2.3. Body Condition
For B. barbus, G. gobio, P. bonelli and P. parva the relative condition factor (Kn) was calculated for all individual following [52]. Primarily, the length–weight relationship equation was estimated for the total sample, applying the least-squares method [53], using the logarithmic equation:
log10W(g) = a + b log10TL (cm)
The values of the regression coefficients a and b were used to estimate theoretically expected values of body weight using the equation:
W = a × TLb
Then, the relative condition factor (Kn) was calculated as follows:
Kn = 100W/(a × TLb)
This approach is based on the assumption that high Kn values characterize fish in good condition and good health, while low Kn values are typical of fish in poor body condition [54].
2.4. Statistical Analysis
To analyze the relationship between the time of introduction and spatial distribution of B. barbus, G. gobio, P. bonelli and P. parva, the Pearson correlation between the number of invaded sites and the species residence time (i.e., number of years since their first detection in the study area) was calculated. To test the ecological preferences of B. barbus, G. gobio, P. bonelli and P. parva a one-way ANOVA was performed to compare average values of environmental variables among the three sampling periods, considering only the sites in which the species were detected. Data used for the Pearson correlation and one-way ANOVA were tested for normality using the Shapiro-Wilk test.
To assess the changes over time of population densities and body condition for B. barbus, G. gobio, P. bonelli and P. parva, a one-way repeated-measures ANOVA was carried out. Data were tested for sphericity using the Mauchly test, and the Greenhouse-Geisser and Huynh-Feldt adjustments were used. ANOVA was processed with Dell STATISTICA 13 software (Dell Inc., Aliso Viejo, CA, USA) for Windows.
In order to extract the environmental variables that drive the fish species densities over the three census periods (1998–2004, 2005–2011 and 2012–2018), Canonical Correspondence Analysis (CCA) [55] was performed. The software CANOCO statistical package 4.5 for Windows (Microcomputer Power, Ithaca, NY, USA), was used to carry out the multivariate analysis. CCA was conducted using environmental variables previous selected by a Principal Component Analysis. The environmental variables more strongly associated with each of the first six components have been selected, and the final environmental matrix included 13 environmental parameters and 91 observations (sampling sites). The fish matrix included 4 variables (fish species) and 91 observations (sampling sites). Given that environmental and fish data were collected as repeated surveys of the same sites, according to Lepš and Šmilauer [56] the analysis was performed using time (i.e., census period) as a covariate. Since the CCA is a unimodal gradient analysis method, to avoid biases the sampling sites have been chosen to be distant more than one kilometer from dams or weirs. All variables (N) used in the analysis were transformed [log10 (N + 1)] to normalize their distributions [57] and standardized to a mean of 0 and standard deviation of 1. The Monte Carlo test based on 999 permutations was used to test the significance of the analysis, and a Generalized Linear Models (GLM) was used to fit the relationship between the fish species densities and elevation.
2.5. Detection Probability, Occupancy, Colonization and Extinction Probabilities
For B. barbus, G. gobio, P. bonelli and P. parva a multi-season model was used to estimate the detection probabilities (pi = probability that the species will be detected at site i at time t, given its presence) [58], occupancy (ψi = probability that the species is present at site i) [59], colonization (ɣ = probability that a site unoccupied at time t is occupied in t + 1), local extinction (ε = probability a site occupied in t is unoccupied in t + 1) [60]. The analysis was carried out using presence-absence data collected in 91 sampling sites across three census periods (t1 = 1998–2004, t2 = 2005–2011, t3 = 2012–2018). Akaike’s information criterion [61] was used to select the best candidate model. Following Burnham and Anderson [62], all models within 2 AIC units were considered as a set of parsimonious models. The analysis was processed using the program PRESENCE (version 2.13.6, Vermont Cooperative Fish and Wildlife Research Unit, Burlington, VT, USA) for Windows.
Five covariates that was believed to play a key role as climate drivers for the spread a distribution of the species were considered in the analysis. Since they do not vary over time, elevation and river fragmentation degree were used as site covariates. Water temperature, average current speed and flow rate, which were all point estimated, were used as sample covariates. Covariates for which 95% confidence intervals (α = 0.05) overlapped zero and with β/SE absolute values < 1.4 were considered [63]. The analysis was performed using the PRESENCE program for Windows.
3. Results
3.1. Distribution Pattern
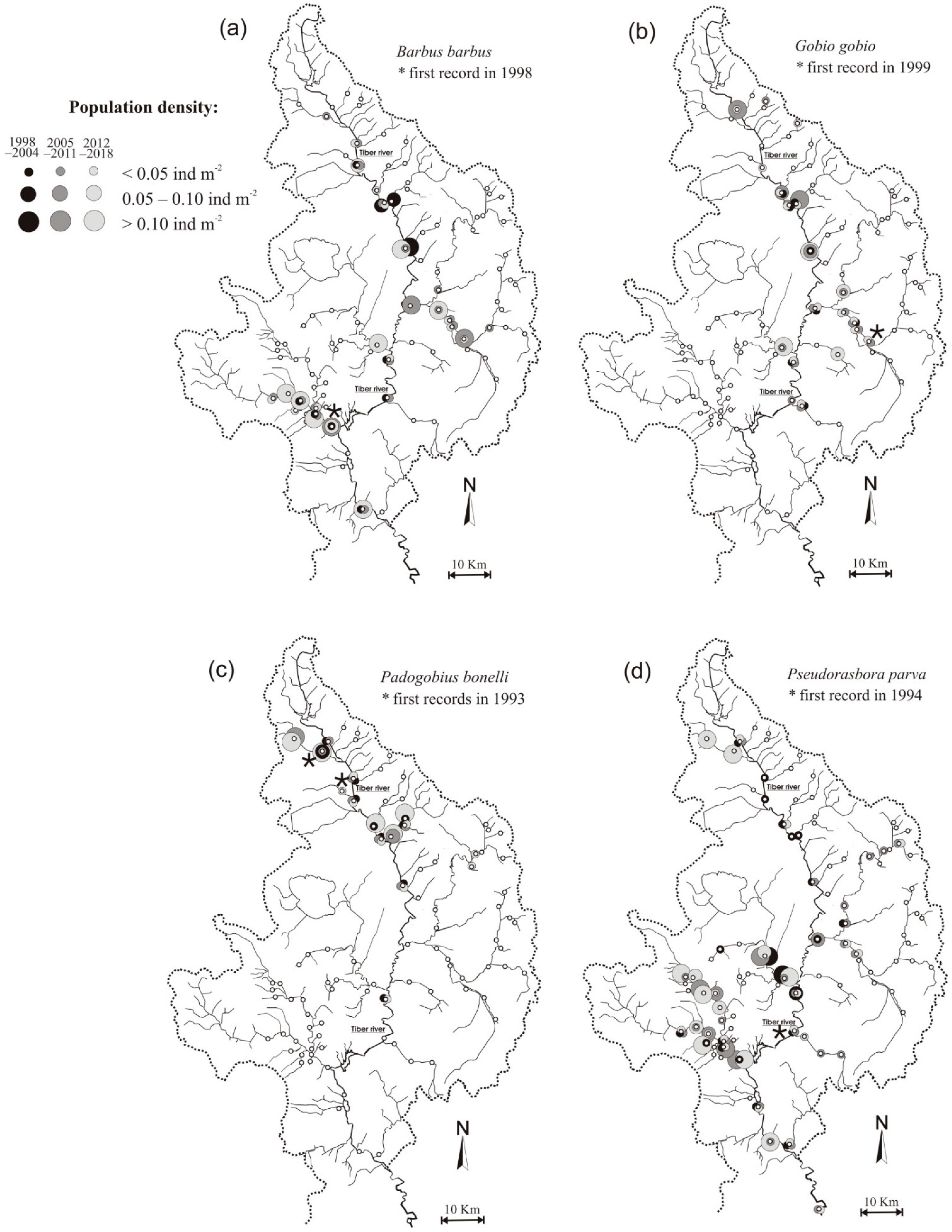
Following its introduction occurred in 1998, B. barbus has colonized the main channel of the Tiber river and its main left and right tributaries, reaching the greater population abundances especially in more recent times (Figure 2a). The distribution of G. gobio was restricted to the middle and upper stretches of the Tiber river, and some of its left tributaries, where the introduction of the species took place over 20 years ago; also, in this case the highest population density values could be observed in more recent times (Figure 2b). The populations of P. bonelli were mainly circumscribed in the upper part of the Tiber river basin, where its introduction occurred 27 years ago, even if the sporadic presence of the species has been recently detected in the mountain portion of a left tributary, the Chiascio river; also, for P. bonelli it was possible to notice how the most abundant populations distinguish, above all, the most recent census period (Figure 2c). As has been observed for B. barbus, also P. parva, following its introduction occurred in 1994 in the downstream reaches of the Tiber river, has rapidly colonized the whole hydrographic network, with abundant populations that already be found starting from the years 2005–2011 (Figure 2d).
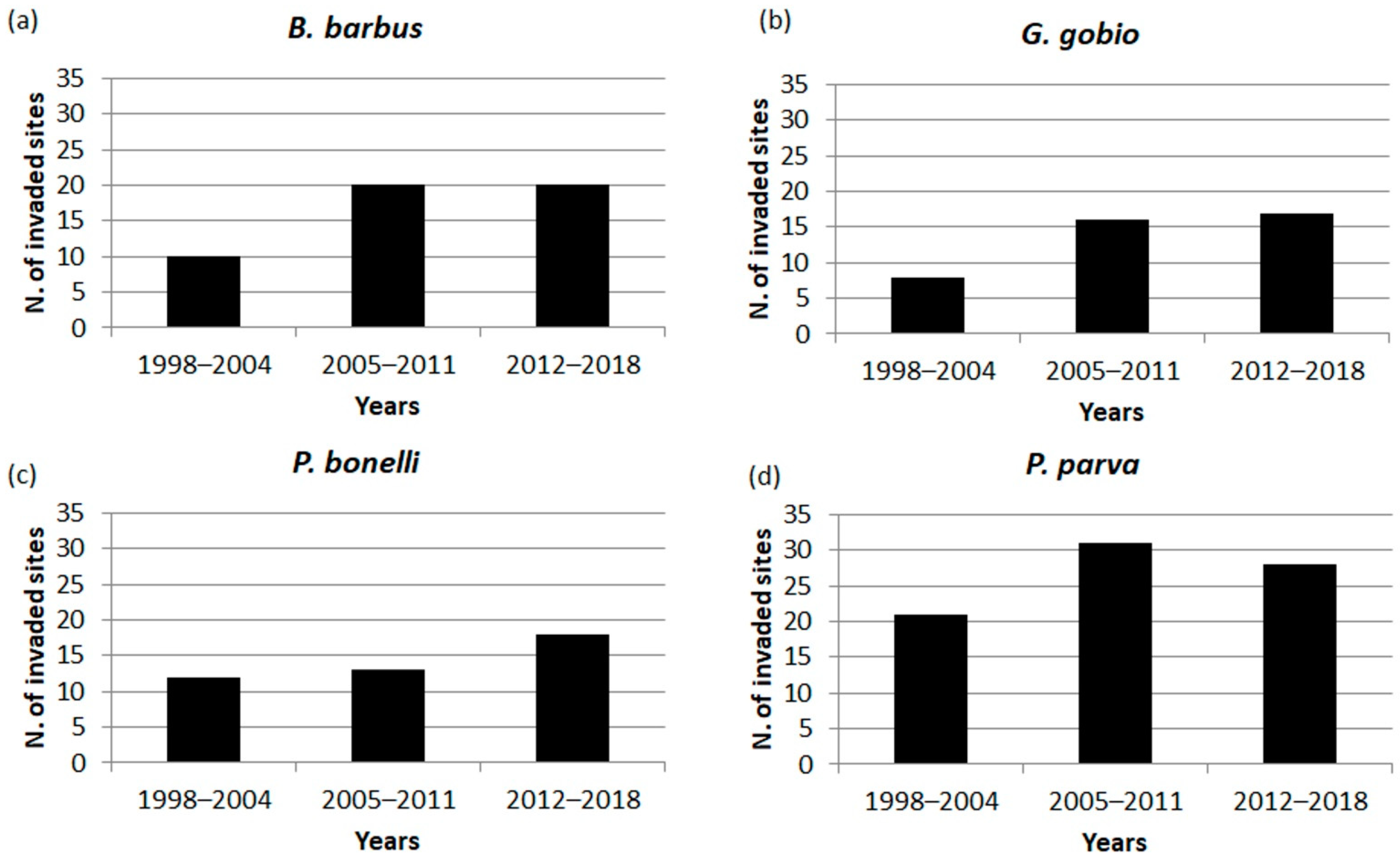
For all the four species, the number of invaded sites was closely related to residence time (r = 0.79; p = 0.001). For B. barbus the number of colonized sites over time was stable in the last two census period and stood at a total of 20 sites (Figure 3a). The invaded sites progressively increased for G. gobio, until reaching a total of 17 sites in the most recent years (Figure 3b). For P. bonelli the number of occurring sites remained quite stable and equal to 12 and 13 in the first two census periods, respectively, while an increase of 5 sites was observed in more recent years (Figure 3c). P. parva was characterized by the greater number of colonized sites compared to other species, with 31 sites invaded in the period 2005–2011; a slight decrease in the last census period was observed, when the number of invaded sites was 28 (Figure 3d).
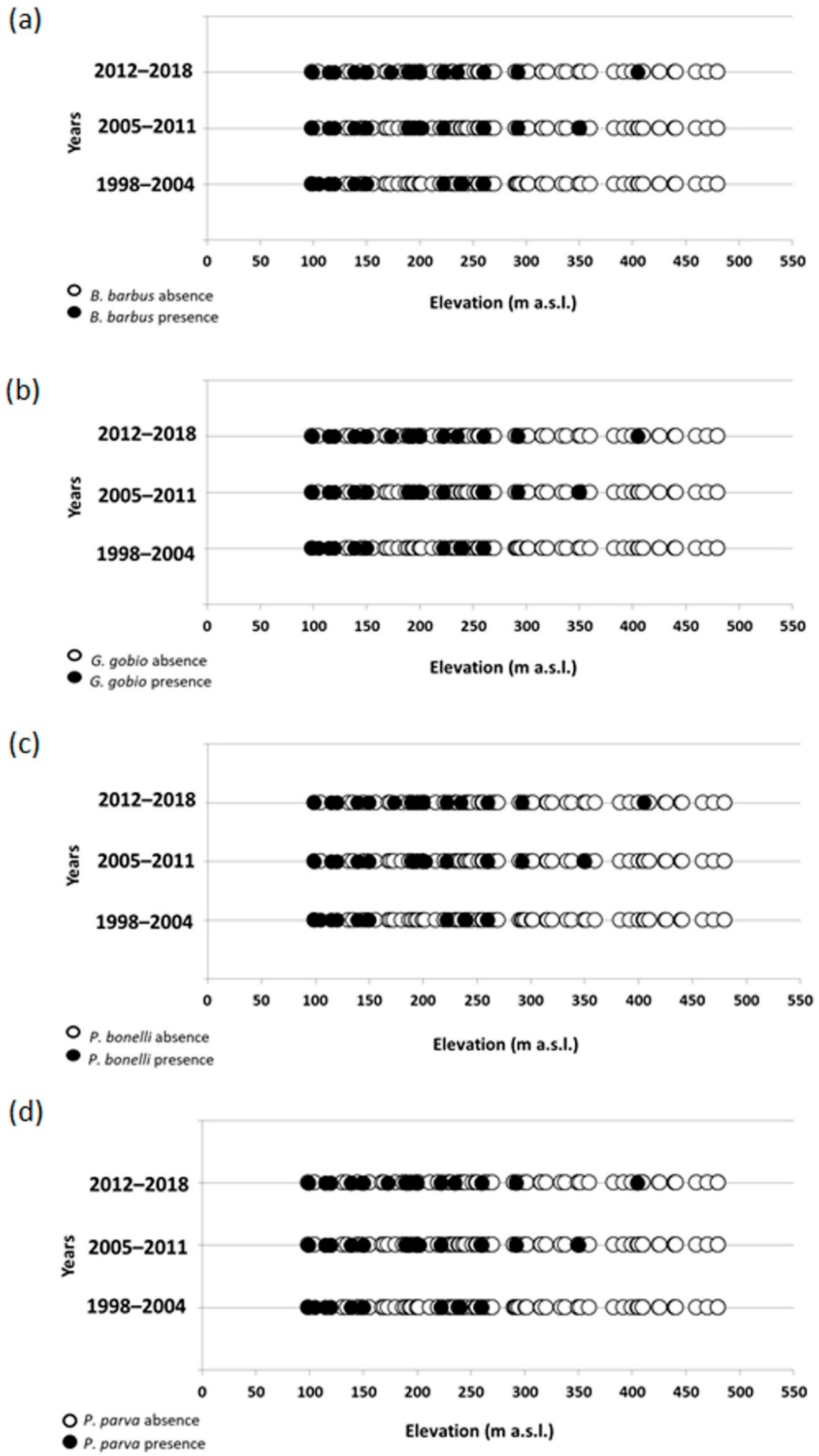
For B. barbus the occupancy-elevation plot showed a prevalent distribution of the species in sites located at lower altitudes, between 100 and 260 m a.s.l.; a tendency of the species to colonize sites located at higher altitudes over time was also observed, with a difference between the maximum altitudes at which the species was recorded in the 1st (245 m a.s.l.) and 3rd census period (316 m a.s.l.) equal to 71 m a.s.l. (Figure 4a). G. gobio was distributed mainly in the sites of altitude between 150 and 250 m a.s.l., even if also in this case during the last two census periods the species colonized sites located at altitudes over the 400 m a.s.l. (Figure 4b). For P. bonelli, the altitude range of the colonized sites fell mainly between 210 and 320, even if in more recent years the presence of the species has been detected at higher altitudes, up to 460 m a.s.l.; in this case the difference between the maximum altitudes at which the species was recorded in the 1st and the last census period was equal to 140 m a.s.l. (Figure 4c). As for B. barbus, although the distribution of P. parva seems to involve more the downstream reaches located in the plain areas, at altitudes between 80 and 250 m a.s.l., in the last two census periods the species colonized sites located at 410 m a.s.l. (Figure 4d).
3.2. Population Status
For all species there was an increasing trend over time of the mean population density values, even if, except for P. bonelli, in all other cases the results were not statistically significant at the rmANOVA (Figure 5). B. barbus and P. bonelli were characterized by large population size especially in the last sampling period, when the mean abundance values exceeded 0.4 ind m−2 for both species (Figure 5a,c). In the case of G. gobio and P. parva the mean density values were always below 0.13 ind m−2 (Figure 5b,d).
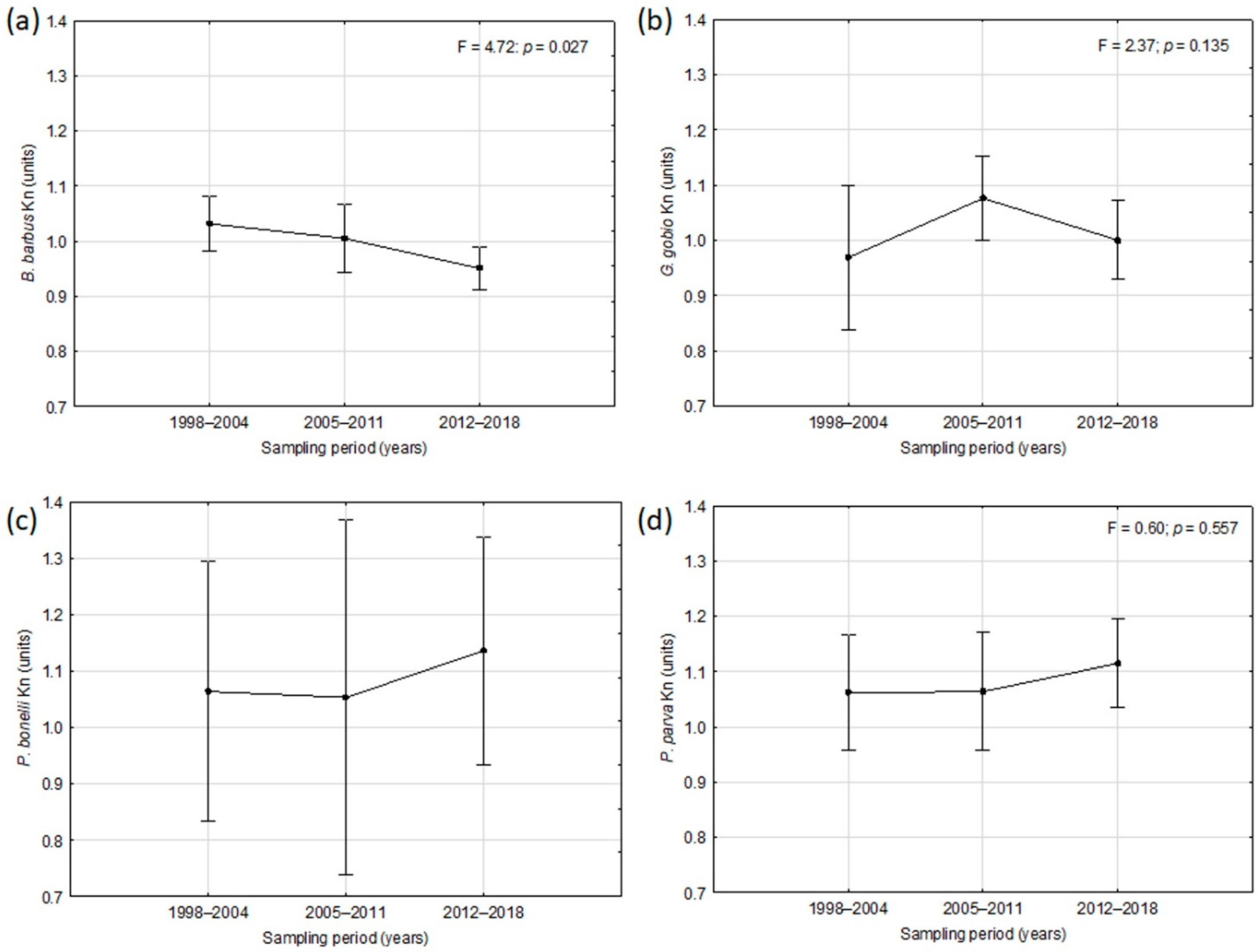
As regards the body condition, a progressive decreasing trend over time of the mean Kn values was observed for B. barbus, even if the average values always remained almost close to unity, that could be considered the optimal value for this parameter (Figure 6a). On the contrary, an increasing trend in the last period of the condition factor was observed for P. bonelli (Figure 6c) and P. parva (Figure 6d), for which in the two last sampling periods the values remained always above 1.0 unit. For G. gobio a fluctuating trend was observed, even if also in this case the Kn mean values remained close to 1.0 (Figure 6b). Except for B. barbus, for all the other species the results were not statistically significant at the rmANOVA.
3.3. Ecological Requirements
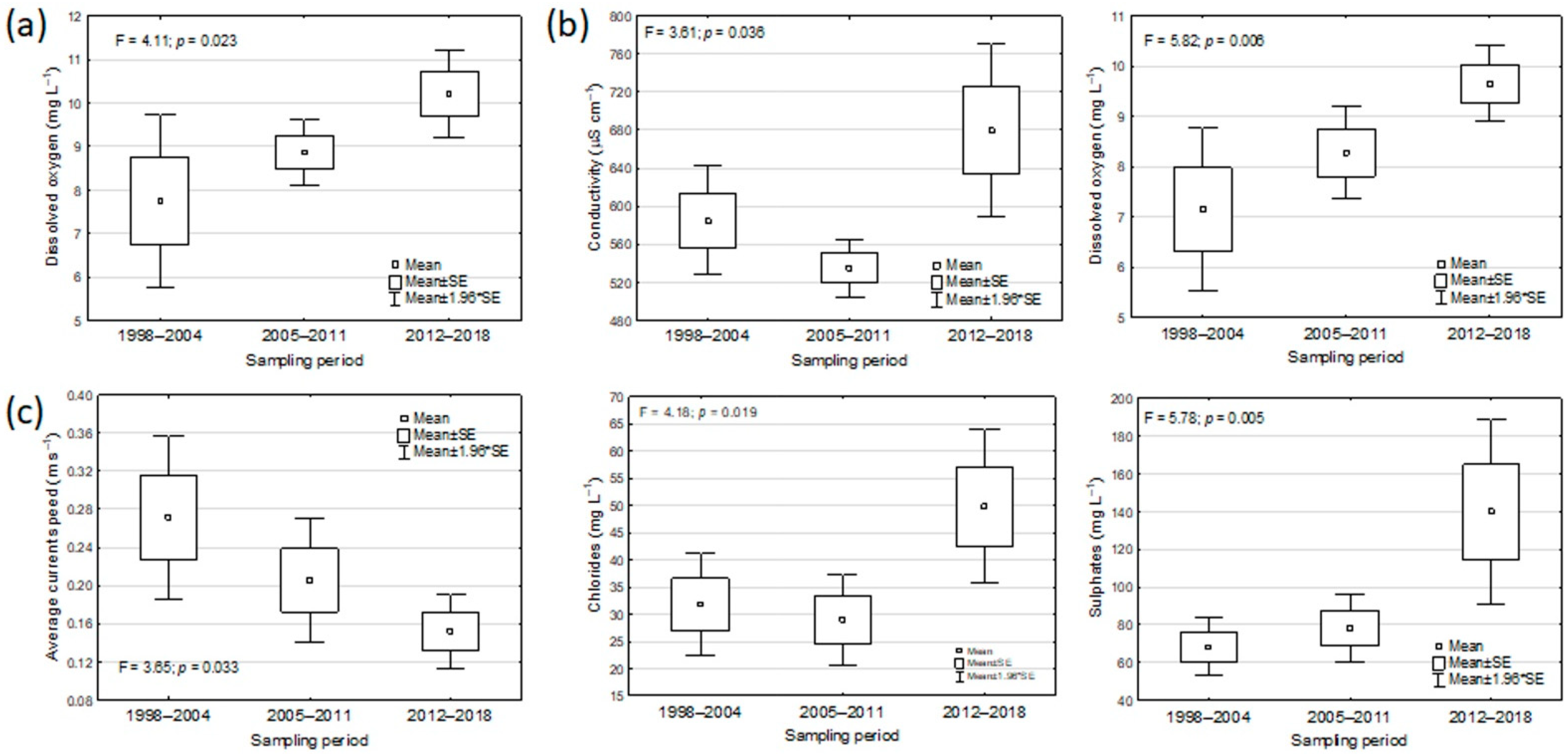
In B. barbus and P. bonelli occurring sites, a significant increase over time in the mean value of the dissolved oxygen concentration was detected (ANOVA B. barbus: F = 4.11, p = 0.023; P. bonelli: F = 5.82, p = 0.006), reaching in the last census values of around 10 mg L−1 for both species (Figure 7a,b). At the sites where the presence of P. parva had been detected, the results showed a progressive significant decrease over time in the average current speed (ANOVA: F = 3.65, p = 0.033), while especially in the last census period, a notable increase in the average values of chlorides and sulphates was observed (Figure 7c). These results, together with the significant increase in conductivity in P. bonelli occurring sites (Figure 6b), could reflect a worsening of water quality over time resulting from the indirect effects of reduced flow rates and current speed in the occurring sites, in terms of greater pollutants concentration (Figure 7c and Figure S1c).
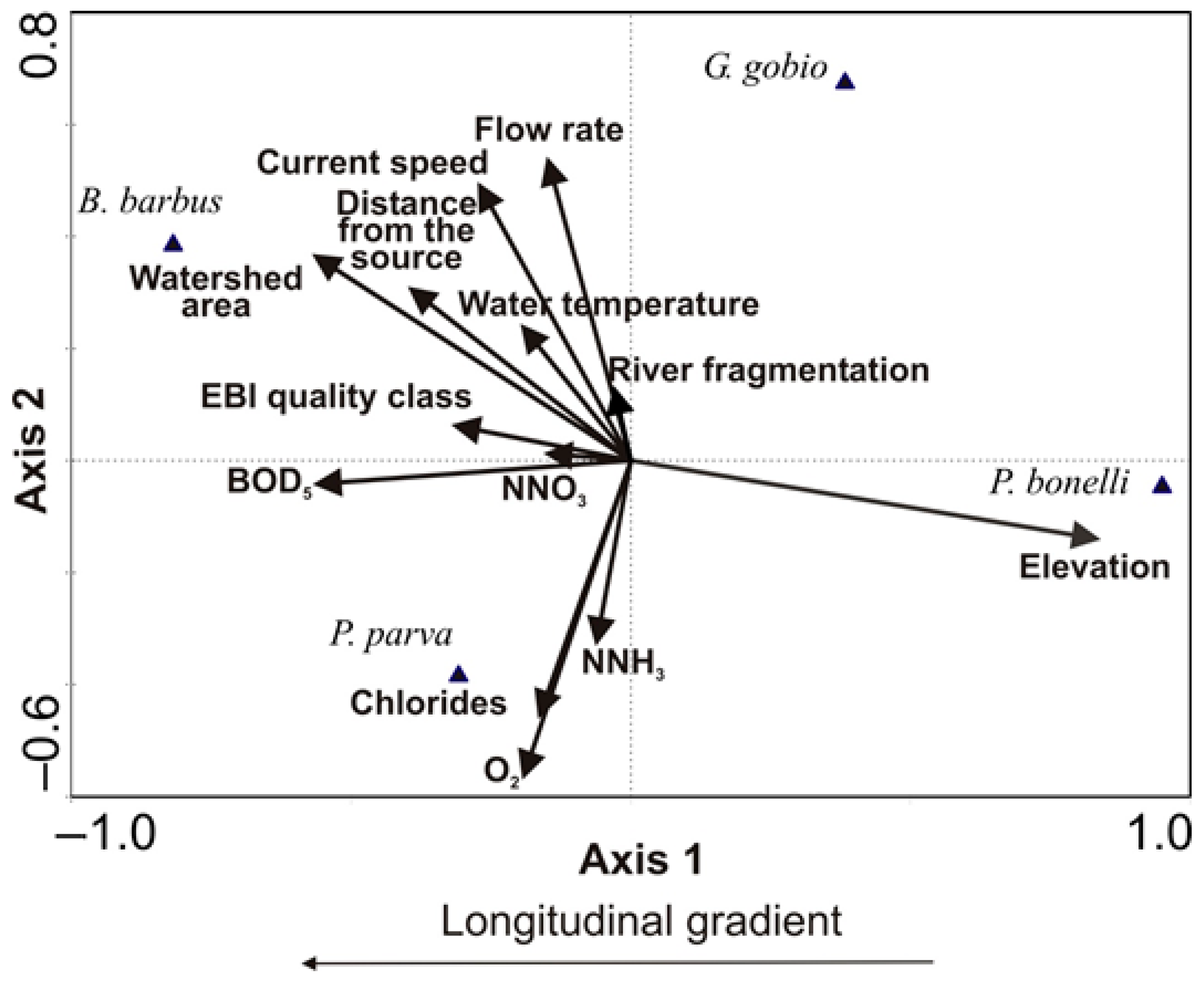
The axes 1 and 2 of CCA explained 92.8% of the total variability and the analysis was highly statistically significant at the Monte Carlo test (F = 8.50, p = 0.001, total inertia = 2.028). Altitude and river fragmentation degree were significantly positively related to the first axis of CCA, which reflected the upstream-downstream gradient of the watercourses (Table 1). BOD5, chlorides, dissolved oxygen, NNH3, NNO3, and watershed area showed a significant inverse relationship to Axis 2 (Table 1). The scattered distribution of points indicating B. barbus, G. gobio, P. bonelli and P. parva positions, suggested that the 4 species inhabit different river stretches with different environmental characteristics within the study area (Figure 8). For P. bonelli the projection point showed the close relation of the species with altitude, while G. gobio was placed in the river stretches located a little more downstream, where the flow rates are higher. P. parva and B. barbus preferentially colonized the downstream reaches of larger rivers, where the water quality is rather poor, and the water temperature is higher.
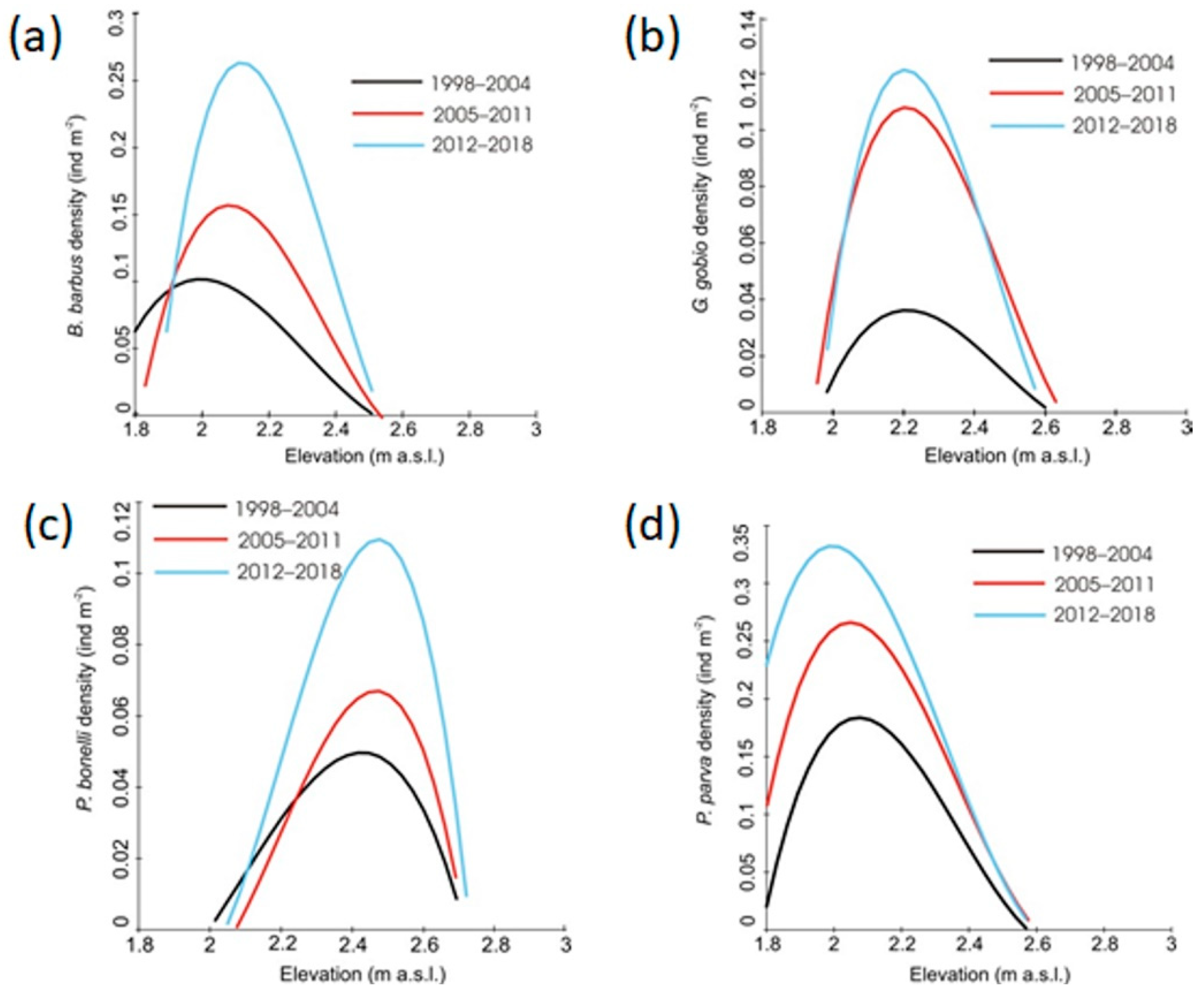
For all the four species the GLM results showed a progressive increase of population abundances over the three census periods (Figure 9). For B. barbus and P. bonelli a gradual shift toward higher altitudes has been also observed (Figure 9a,c). For G. gobio and P. parva, the analysis did not reveal significant changes in the elevation preferences of the species (Figure 9b,d).
3.4. Detectability, Occupancy, Colonization, and Local Extinction Probabilities
The detection probability was quite high and never lower than 0.74, even if perfect detection (1.00) was never achieved; the highest value was observed for G. gobio (0.86 ± 0.06), while the lowest detection probability was observed for P. parva (0.74 ± 0.06). Occupancy probability values were not very high but still noteworthy, considering that they referred to alien species introduced in recent decades; the occupancy probability varied from 0.14 ± 0.04 for B. barbus to 0.31 ± 0.07 for P. parva. Colonization probabilities ranged from 0.12 ± 0.04 for G. gobio to 0.26 ± 0.07 for P. parva; these values were considerably higher than the local extinction probabilities, which for G. gobio and P. bonelli were 0.00, while the highest value referred to P. parva (0.18 ± 0.10) (Table 2).
The lowest values of detectability were observed during the first census period for all species (Table 3). For B. barbus, the detection probabilities over time varied from 0.53 ± 0.06 (years 1998–2004) to 0.80 ± 0.08 (years 2012–2018). Additionally, for G. gobio and P. bonelli an increase over time of detectability was observed, with mean values varying from 0.42 ± 0.01 and 0.67 ± 0.11 (years 1998–2004) to 0.88 ± 0.08 and 0.93 ± 0.07 (years 2012–2018), respectively. For P. parva the detection probabilities ranged from 0.53 ± 0.06 (years 1998–2004) to 0.71 ± 0.08 (years 2012–2018) (Table 3).
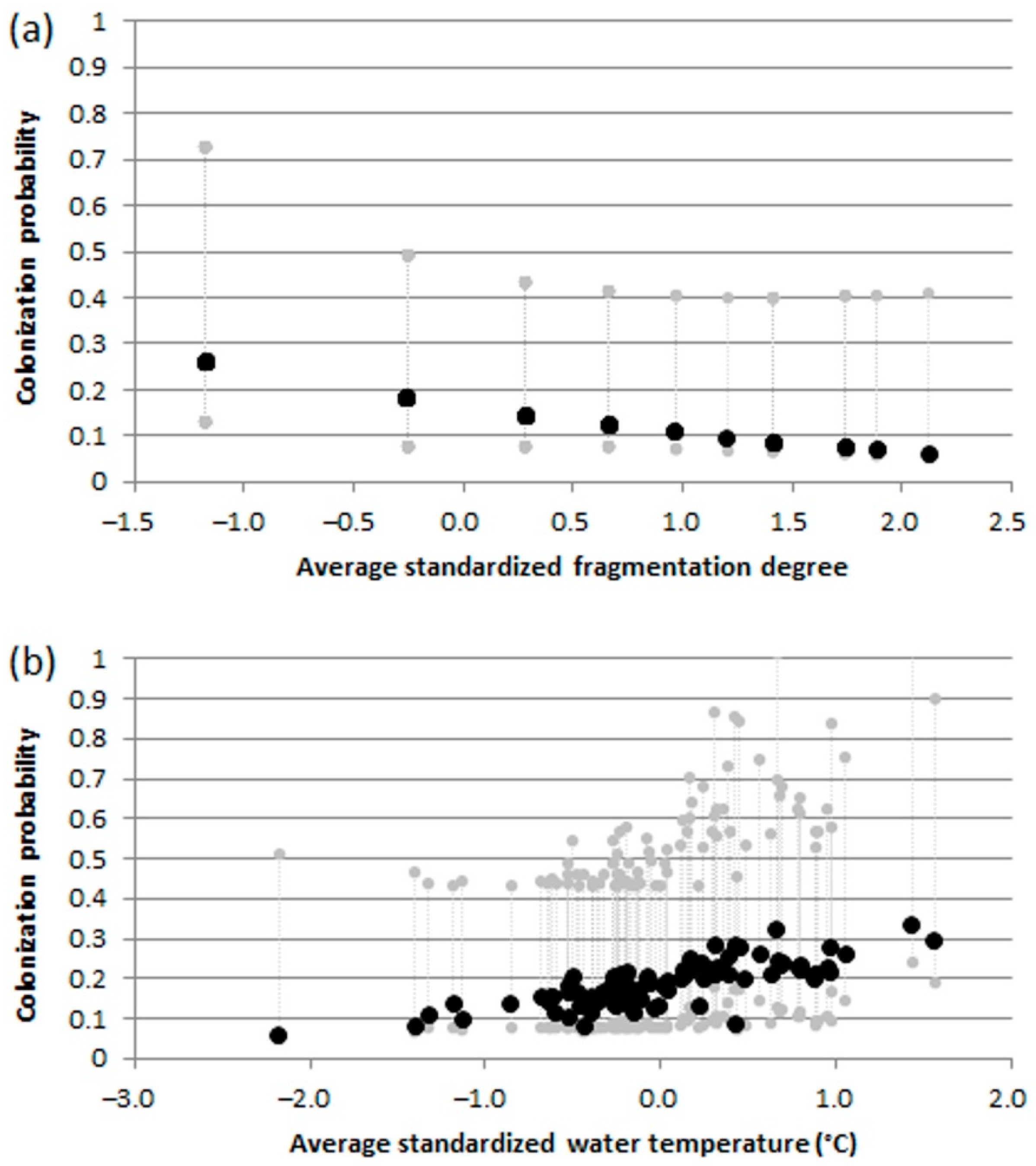
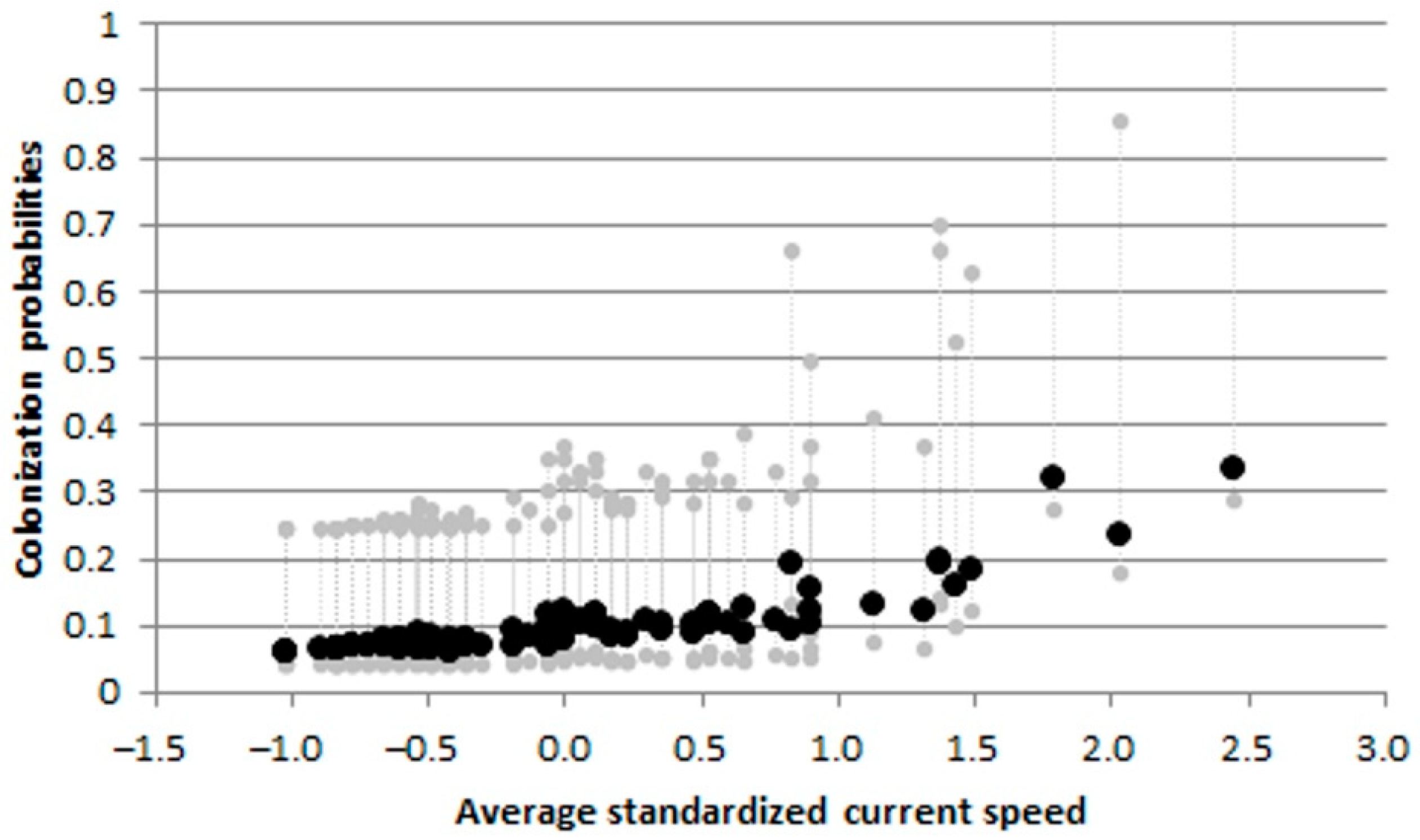
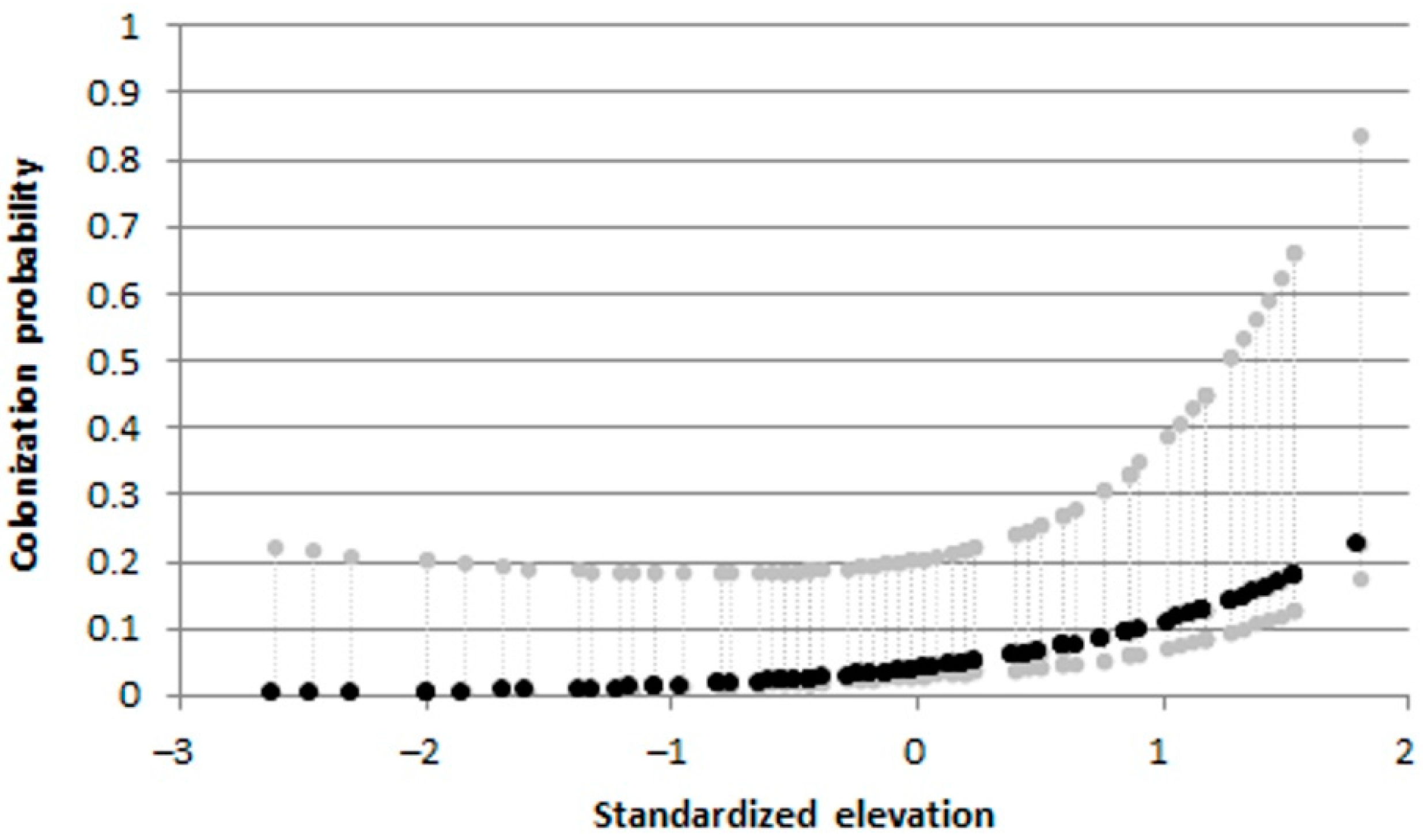
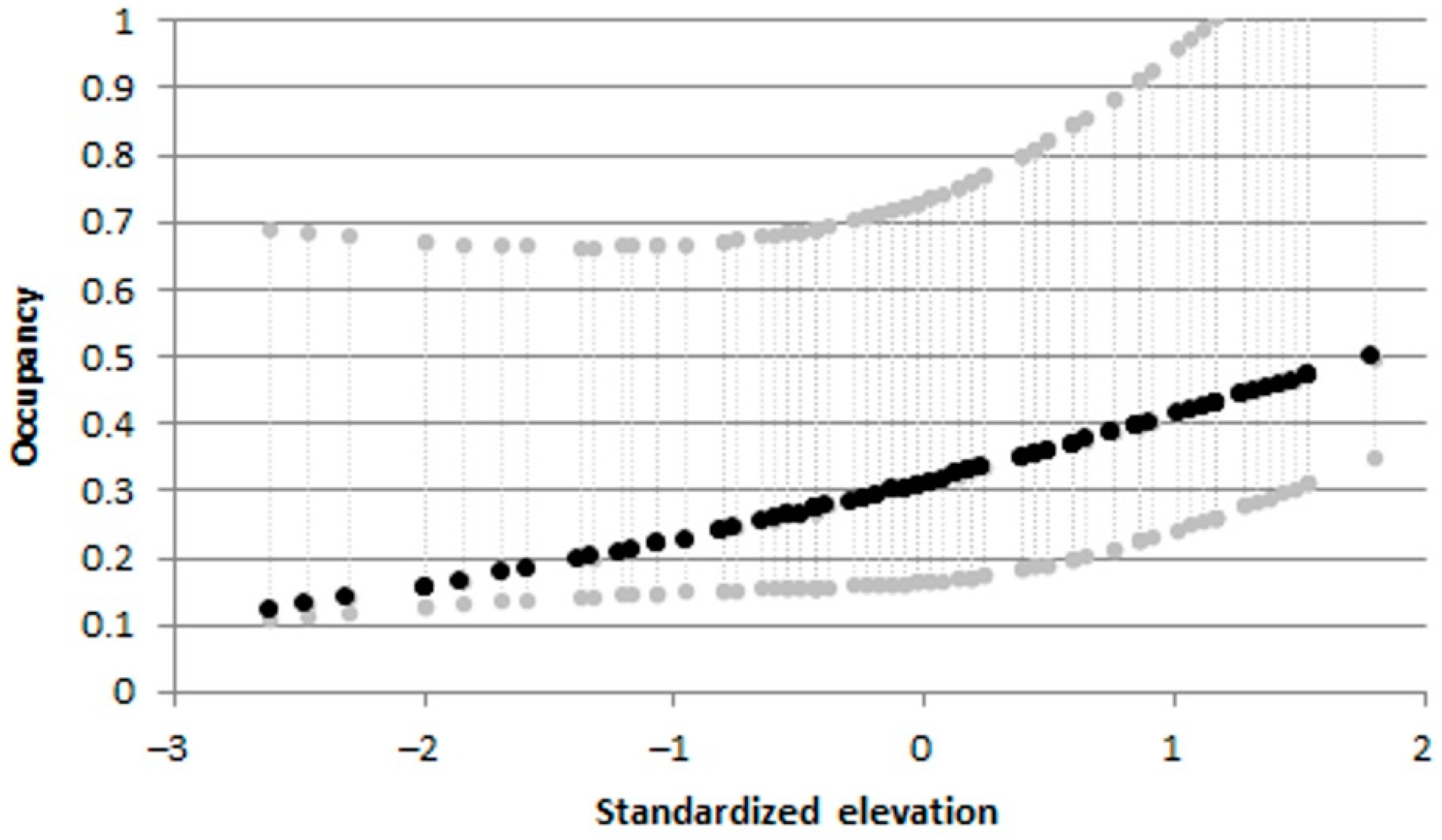
The best multi-sampling period models, within two AIC units of the top model, for estimating occupancy, colonization, local extinction, and detection, included: elevation, fragmentation degree, water temperature and average current speed as informative variables (Table S1). For B. barbus, colonization probabilities progressively decreased with increasing river fragmentation degree (Figure 10a) and were highest at the warmer sites (Figure 10b). For G. gobio the colonization probabilities increased according to the increasing average current speed (Figure 11). As regards P. bonelli, colonization probability increased with elevation, while occupancy increased with river fragmentation degree (Figure 12). For P. parva the occupancy probability increased with elevation (Figure 13).
4. Discussion
As reported for other European countries [38] a close positive relationship between the time of introduction and spatial distribution of the alien species was observed in the Tiber river basin. Each species showed its own invasion pattern, in which the environmental factors that are major determinants under climate change conditions seem to be mostly represented by river fragmentation, water temperature and hydrological parameters. Most alien species are expanding their range because probably in the last twenty years they have not yet completed their dispersal process. In addition to the dispersal capability of the species, some effects of climate change, in terms of water temperature increase, seemed to favor especially cool and warm-water species, playing a key role in their expansion.
B. barbus, being among the species introduced in more recent years (1998), after having rapidly invaded numerous sites, and having formed ever more abundant populations over time [26], seemed to be still expanding its range. For the species was observed a tendency to progressively move towards the river stretches located at higher altitudes, probably favored by the water warming consequent to climate change. Moreover, the high density population values reached in the last census period may in part have contributed to the spread of the species, due to intraspecific competition phenomena [64]. Moreover, as this is a species of particular interest for sport fishing [65], secondary introductions in multiple sites through illegal stocking could have encouraged the expansion of the species [38,43]. However, a brake on the expansion of a vagile species such as B. barbus can be represented by river fragmentation, as also reported in the literature for some large rivers of England and Belgium [65,66]. Thus, the presence of obstacles to migration for reproductive purposes could affect B. barbus response to climate change [67], especially in a dendritic hydrographic network as the Tiber river basin, where a single barrier to migration and dispersal could create an extended isolated area [68].
Also G. gobio resulted in rapid expansion in space and time, with a number of colonized sites progressively increasing over time, and with more abundant populations showing good body condition. The average current speed plays a key role in colonization processes, as there was a positive relationship with the colonization probabilities. Being a species introduced in the Tiber river basin in fairly recent times (first record in 1999), further research based on long-term data series are needed to highlight the ecological effects of G. gobio invasion, and to evaluate the resilience of the species to global warming.
For P. bonelli our results highlighted a resilience to climate change effects, showing a range shift towards upstream reaches, probably favored by water warming. In fact, elevation exerted a positive effect on colonization probabilities. Especially in the last census period, the number of colonized sites and the density of the populations have increased considerably, in combination with a good body condition. Contrary to B. barbus, P. bonelli is not of interest to sport fishing; however, the species may have been the subject of human-mediated involuntary translocations among different watercourses, especially during transfers of fish trapped in pools during drought periods, made by unauthorized volunteers. This would explain the appearance of the species in recent times in the Chiascio river, where P. bonelli could not have arrived using the natural connections of the hydrographic network. As reported for Neogobius melanostomus (Pallas, 1814) in the Greath Lakes, the secondary human-made traslocations within a basin could play a key role in the secondary diffusive spread of an invasive species [64]. However, as in our case these are occasional and unpredictable events, involving few individuals, it is difficult to document this kind of secondary introductions, and it is difficult to establish how much they may have influenced the expansion of the species in the Tiber river basin.
As for B. barbus and P. bonelli, also in the case of P. parva it is possible to hypothesize a “step and stone diffusion” dispersal pattern [25,69]; in particular, starting from the two first introduction points, the species could have colonized the hydrographic network using the natural connections and spreading both downstream and upstream, thanks to its pollution and thermal tolerance, also aided by involuntary secondary releases. Being a species of less recent introduction, P. parva seemed to have reached, at this stage, a temporary equilibrium in terms of distribution in the Tiber river basin. The stop in the increase in average populations density and the decrease in colonized sites in the last census period, provided some evidence that P. parva seems to have almost completed the dispersal process in the study area. However, being a warm-water species, this balance could be destabilized in the future due to the effects of climate change that tend to benefit species with such thermal preferences [38]. Our results confirmed that one of the major drivers for P. parva is represented by the average current speed, as was also demonstrated in a previous study [25]; in fact, as reported in the literature, the species inhabits lowland areas with low current speed, where it must not incur an excessive expenditure of energy for swimming, avoiding metabolic stress [70].
Investigations on the relationships between habitat and fish species are fundamental to preserve native biodiversity and to manage multi-stressed freshwater ecosystems [71]. This is the case of the Tiber river basin, where previous research has shown that changes in the composition of fish communities along the longitudinal gradient of watercourses were associated with a progressive increasing perturbation proceeding from upstream to downstream [20,72], as also highlighted by Saez-Gomez et al. [71] for other highly stressed Mediterranean basins. The 4 species considered, although belonging to the barbel zone [73], have shown different ecological requirements, with P. bonelli seeming to prefer environments with cleaner waters located at higher altitudes, while P. parva showed greater tolerance with respect to environmental degradation in terms of water pollution. G. gobio was placed in river stretches showing intermediate characteristics. The increase over time in the concentration of dissolved oxygen in the sites where B. barbus and P. bonelli were present, could be linked to the range shift of the two species that are moving upstream. The progressive increase in chlorides and sulphates in P. parva occurring sites could reflect a worsening of water quality as an indirect effect of the progressive decrease in the average current speed.
The GLM analysis showed for B. barbus and P. bonelli range shifts towards sites located at higher altitude. These results match the hypothesis that rising water temperatures could favor the upstream colonization by cool-water adapted species, although the response curves could be influenced by the fact that the species have not yet completed the expansion process in the invaded area [43].
As might have been expected in the case of species introduced in recent decades, the colonization probabilities for the 4 species were much higher than extinction ones. The colonization processes were driven by elevation for P. bonelli and P. parva, while average current speed exerted a positive effect on colonization processes for G. gobio. Water temperature and river fragmentation play a key role in B. barbus colonization processes. In particular, for this species, colonization probabilities increased with water temperature and decreased with river fragmentation degree. This latter result confirmed that barriers (including human-made weirs and drought river stretches) could hinder the expansion of alien species [68], but at the same time should be considered that they prevent the range shift of native species to achieve thermal optimum, leading to a loss of biodiversity [43,74]. However, in the ecological evaluation of these relationships between colonization and environmental variables should be taken into account that these species were probably not yet stabilized and have not yet reached a stable equilibrium [75].
5. Conclusions
Our results provided some evidence for synergistic effects between climate change effects and alien cool-water adapted species invasions. The range shifts towards upstream reaches of B. barbus and P. bonelli allowed us to consolidate the hypothesis of a further expansion of these invasive species, favored by rising temperatures that provide new suitable habitats in which to live and reproduce. However, these results should be interpreted cautiously, taking into account that these species most likely were not yet stabilized [7,43]. The interruptions of the river connectivity can counteract the dispersal ability and resilience of invasive species by limiting the colonization probabilities. Effective conservation policy and management measures to be taken to contrast the introduction and expansion of alien fish species (i.e., early detection, eradication and removal activities, preventing secondary introductions through environmental education) [64] should be accompanied by interventions aimed at mitigating the global climate change effects on multi-stressed freshwater ecosystems (i.e., riparian restoration, maintenance of ecological flow) [43].
Supplementary Materials
The following are available online at https://www.mdpi.com/2073-4441/13/3/347/s1, Figure S1: Ecological requirements: trend over time of the mean values of elevation, water temperature and average current speed for: (a) Barbus barbus, (b) Gobio gobio, (c) Padogobius bonelli, (d) Pseudorasbora parva, Table S1: Models within two Akaike information criterion (AIC) units of the top model for estimating occupancy (ψ), colonization (ɣ), local extinction (ε), and detection (p) probabilities for: (a) Barbus barbus, (b) Gobio gobio, (c) Padogobius bonelli, (d) Pseudorasbora parva.
Author Contributions
Conceptualization, A.C. and M.L.; methodology, A.C. and M.L.; investigation, A.C., L.G. and M.L.; data curation, A.C. and L.G.; writing—original draft preparation, A.C.; writing—review and editing, A.C. and M.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
The authors wish to thank all of the people who joined in the field activities during the project. They also thank the anonymous reviewers and the editorial staff for helpful comments on the original version of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Strayer, D.L. Alien species in fresh waters: Ecological effects, interactions with other stressors, and prospects for the future. Freshw. Biol. 2010, 55, 152–174. [Google Scholar] [CrossRef]
- Ricciardi, A.; Hoopes, M.F.; Marchetti, M.P.; Lockwood, J.L. Progress toward understanding the ecological impacts of nonnative species. Ecol. Monogr. 2013, 83, 263–282. [Google Scholar] [CrossRef] [Green Version]
- Moyle, P.B.; Kiernan, J.D.; Crain, P.K.; Quiñones, R.M. Climate change vulnerability of native and alien freshwater fishes of California: A systematic assessment approach. PLoS ONE 2013, 8, e63883. [Google Scholar] [CrossRef] [PubMed]
- Walther, G.-R.; Roques, A.; Hulme, P.E.; Sykes, M.T.; Pyšek, P.; Kühn, I.; Zobel, M.; Bacher, S.; Botta-Dukát, Z.; Bugmann, H.; et al. Alien species in a warmer world: Risks and opportunities. Trends Ecol. Evol. 2009, 24, 686–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, A.L.; Hewitt, N.; Klenk, N.; Bazely, D.R.; Yan, N.; Wood, S.; Henriques, I.; MacLellan, J.I.; Lipsig-Mummé, C. Effects of climate change on the distribution of invasive alien species in Canada: A knowledge synthesis of range change projections in a warming world. Environ. Rev. 2012, 20, 1–16. [Google Scholar] [CrossRef]
- Rolls, R.J.; Hayden, B.; Kahilainen, K.K. Conceptualising the interactive effects of climate change and biological invasions on subarctic freshwater fish. Ecol. Evol. 2017, 7, 4109–4128. [Google Scholar] [CrossRef] [PubMed]
- Bellard, C.; Jeschke, J.M.; Leroy, B.; Mace, G.M. Insights from modeling studies on how climate change affects invasive alien species geography. Ecol. Evol. 2018, 8, 5688–5700. [Google Scholar] [CrossRef]
- Graham, C.T.; Harrod, C. Implications of climate change for the fishes of the British Isles. J. Fish Biol. 2009, 74, 1143–1205. [Google Scholar] [CrossRef]
- Cuttelod, A.; García, N.; Malak, A.D.; Temple, H.; Katariya, V. The Mediterranean: A biodiversity hotspot under threat. In Wildlife in a Changing World—An Analysis of the 2008 IUCN Red List of Threatened Species; Vié, J.C., Hilton-Taylor, C., Stuart, S.N., Eds.; IUCN: Gland, Switzerland, 2009; pp. 89–104. [Google Scholar]
- Hermoso, V.; Clavero, M. Threatening processes and conservation management of endemic freshwater fish in the Mediterranean basin: A review. Mar. Freshw. Res. 2011, 62, 244–254. [Google Scholar] [CrossRef]
- Erol, A.; Randhir, T.O. Climatic change impacts on the ecohydrology of Mediterranean watersheds. Clim. Chang. 2012, 114, 319–341. [Google Scholar] [CrossRef]
- Marr, S.M.; Olden, J.D.; Leprieur, F.; Arismendi, I.; Ćaleta, M.; Morgan, D.L.; Nocita, A.; Šanda, R.; Tarkan, S.A.; García-Berthou, E. A global assessment of freshwater fish introductions in mediterranean-climate regions. Hydrobiologia 2013, 719, 317–329. [Google Scholar] [CrossRef] [Green Version]
- Senent-Aparicio, J.; Pérez-Sánchez, J.; Carrillo-García, J.; Soto, J. Using SWAT and Fuzzy TOPSIS to Assess the Impact of Climate Change in the Headwaters of the Segura River Basin (SE Spain). Water 2017, 9, 149. [Google Scholar] [CrossRef] [Green Version]
- Isaak, D.J.; Rieman, B.E. Stream isotherm shifts from climate change and implications for distributions of ectothermic organisms. Glob. Chang. Biol. 2013, 19, 742–751. [Google Scholar] [CrossRef]
- Bernardo, J.M.; Ilhéu, M.; Matono, P.; Costa, A.M. Interannual variation of fish assemblage structure in a Mediterranean river: Implications of streamflow on the dominance of native or exotic species. River Res. Applic. 2003, 19, 521–532. [Google Scholar] [CrossRef]
- Ficke, A.D.; Myrick, C.A.; Hansen, L.J. Potential impacts of global climate change on freshwater fisheries. Rev. Fish Biol. Fish. 2007, 17, 581–613. [Google Scholar] [CrossRef]
- Bond, N.; Thomson, J.; Reich, P.; Stein, J. Using species distribution models to infer potential climate change-induced range shifts in freshwater fish in south-eastern Australia. Mar. Freshw. Res. 2011, 62, 1043–1061. [Google Scholar] [CrossRef] [Green Version]
- Moss, B.; Hering, D.; Green, A.J.; Aidoud, A.; Becares, E.; Beklioglu, M.; Bennion, H.; Boix, D.; Brucet, S.; Carvalho, L.; et al. Climate Change and the Future of Freshwater Biodiversity in Europe: A Primer for Policy-Makers. Freshw Rev. 2009, 2, 103–130. [Google Scholar] [CrossRef]
- Olden, J.D.; Kennard, M.J.; Leprieur, F.; Tedesco, P.A.; Winemiller, K.O.; García-Berthou, E. Conservation biogeography of freshwater fishes: Recent progress and future challenges. Divers. Distrib. 2010, 16, 496–513. [Google Scholar] [CrossRef]
- Carosi, A.; Ghetti, L.; Forconi, A.; Lorenzoni, M. Fish community of the river Tiber basin (Umbria-Italy): Temporal changes and possible threats to native biodiversity. Knowl. Manag. Aquat. Ecosyst. 2015, 416, 22. [Google Scholar] [CrossRef] [Green Version]
- Piccoli, F.; Burgazzi, G.; Laini, A.; Ferrari, C.; Voccia, A.; Filonzi, L.; Bolpagni, R.; Nonnis Marzano, F. Barbel species arrangement in a regional Natura 2000 network (Emilia Romagna, Northern Italy): An altitudinal perspective. J. Limnol. 2017, 76, 140–147. [Google Scholar] [CrossRef] [Green Version]
- Gavioli, A.; Milardi, M.; Castaldelli, G.; Fano, E.A.; Soininen, J. Diversity patterns of native and exotic fish species suggest homogenization processes, but partly fail to highlight extinction threats Divers. Distrib. 2019, 25, 983–994. [Google Scholar] [CrossRef] [Green Version]
- Lorenzoni, M.; Borghesan, F.; Carosi, A.; Ciuffardi, L.; De Curtis, O.; Delmastro, G.; Di Tizio, L.; Franzoi, P.; Maio, G.; Mojetta, A.; et al. The check-list of the Italian freshwater fish fauna. Ital. J. Freshw. Ichthyol. 2019, 5, 239–254. [Google Scholar]
- Carosi, A.; Ghetti, L.; Cauzillo, C.; Pompei, L.; Lorenzoni, M. Occurrence and distribution of exotic fishes in the Tiber River basin (Umbria, central Italy). J. Appl. Ichthyol. 2017, 33, 274–283. [Google Scholar] [CrossRef] [Green Version]
- Carosi, A.; Ghetti, L.; Lorenzoni, M. Status of Pseudorasbora parva in the Tiber river basin (Umbria, central Italy) 20 years after its introduction. Knowl. Manag. Aquat. Ecosyst. 2016, 417, 11. [Google Scholar] [CrossRef] [Green Version]
- Carosi, A.; Ghetti, L.; La Porta, G.; Lorenzoni, M. Ecological effects of the European Barbus barbus (L., 1758) (Cyprinidae) invasion on the native barbel populations in the Tiber River basin. Eur. J. Zool. 2017, 84, 420–435. [Google Scholar] [CrossRef] [Green Version]
- Meraner, A.; Venturi, A.; Ficetola, G.F.; Rossi, S.; Candiotto, A.; Gandolfi, A. Massive invasion of alien Barbus barbus and introgressive hybridization with endemic Barbus plebejus in Northern Italy: Where, how and why? Mol. Ecol. 2013, 22, 5295–5312. [Google Scholar] [CrossRef]
- Buonerba, L.; Zaccara, S.; Delmastro, G.B.; Lorenzoni, M.; Salzburger, W.; Gante, H.F. Intrinsic and extrinsic factors act at different spatial and temporal scales to shape population structure, distribution and speciation in Italian Barbus (Osteichthyes: Cyprinidae). Mol. Phylogenet. Evol. 2015, 89, 115–129. [Google Scholar] [CrossRef]
- Geiger, M.F.; Schreiner, C.; Delmastro, G.B.; Herder, F. Combining geometric morphometrics with molecular genetics to investigate a putative hybrid complex: A case study with barbels Barbus spp. (Teleostei: Cyprinidae). J. Fish Biol. 2016, 88, 1038–1055. [Google Scholar] [CrossRef]
- Zaccara, S.; Quadroni, S.; Vanetti, I.; Carosi, A.; La Porta, G.; Crosa, G.; Carosi, A.; Lorenzoni, M. Morphologic and genetic variability in the Barbus fishes (Teleostei, Cyprinidae) of Central Italy. Zool. Scr. 2019, 48, 289–301. [Google Scholar] [CrossRef]
- Zaccara, S.; Quadroni, S.; De Santis, V.; Vanetti, I.; Carosi, A.; Crosa, G.; Britton, J.R.; Lorenzoni, M. Genetic and phenotypic displacement of an endemic Barbus complex by invasive European barbel Barbus barbus in central Italy. Biol. Invasions 2020, 1–15. [Google Scholar] [CrossRef]
- Pompei, L.; Carosi, A.; Forconi, A.; Giannetto, D.; Lorenzoni, M. Distribution and abundance of Padogobius nigricans (Canestrini, 1867) and P. bonelli (Bonaparte, 1846) in the river Tiber basin (Umbria, central Italy). Ital. J. Freshw. Ichthyol. 2016, 3, 28–35. [Google Scholar]
- DAISIE. Species accounts of 100 of the most invasive alien species in Europe. In Handbook of Alien Species in Europe; Springer: Dordrecht, Holland, the Netherlands, 2008; pp. 269–474. [Google Scholar]
- Nentwig, W.; Bacher, S.; Kumschick, S.; Pyšek, P.; Vilà, M. More than “100 worst” alien species in Europe. Biol. Invasions 2018, 20, 1611–1621. [Google Scholar] [CrossRef] [Green Version]
- Giannetto, D.; Carosi, A.; Franchi, E.; Ghetti, L.; Pedicillo, G.; Pompei, L.; Lorenzoni, M. Assessing the impact of non-native freshwater fishes on native species using relative weight. Knowl. Manag. Aquat. Ecosyst. 2012, 404, 3. [Google Scholar] [CrossRef] [Green Version]
- Magnuson, J.J.; Crowder, L.B.; Medvick, P.A. Temperature as an Ecological Resource. Am. Zool. 1979, 19, 331–343. [Google Scholar] [CrossRef] [Green Version]
- Blanck, A.; Tedesco, P.A.; Lamouroux, N. Relationships between life-history strategies of European freshwater fish species and their habitat preferences. Freshw. Biol. 2007, 52, 843–859. [Google Scholar] [CrossRef]
- Rabitsch, W.; Milasowszky, N.; Nehring, S.; Wiesner, C.; Wolter, C.; Essl, F. The times are changing: Temporal shifts in patterns of fish invasions in central European fresh waters. J. Fish Biol. 2013, 82, 17–33. [Google Scholar] [CrossRef]
- Stefani, F.; Schiavona, A.; Tirozzi, P.; Gomarasca, S.; Marziali, L. Functional response of fish communities in a multistressed freshwater world. Sci. Total Environ. 2020, 740, 139902. [Google Scholar] [CrossRef]
- Rahel, F.J.; Olden, J.D. Assessing the Effects of Climate Change on Aquatic Invasive Species. Conserv. Biol. 2008, 22, 521–533. [Google Scholar] [CrossRef]
- Carosi, A.; Padula, R.; Ghetti, L.; Lorenzoni, M. Endemic freshwater fish range shifts related to global climate changes: A long-term study provides some observational evidence for the Mediterranean area. Water 2019, 11, 2349. [Google Scholar] [CrossRef] [Green Version]
- Radinger, J.; García Berthou, E. The role of connectivity in the interplay between climate change and the spread of alien fish in a large Mediterranean river. Glob Change Biol. 2020, 26, 6383–6389. [Google Scholar] [CrossRef]
- Ulbrich, U.; May, W.; Li, L.; Lionello, P.; Pinto, J.G.; Somot, S. The Mediterranean Climate Change under Global Warming. In Mediterranean Climate Variability; Lionello, P., Malanotte-Rizzoli, P., Boscolo, R., Eds.; Elsevier B.V.: Amsterdam, The Netherlands, 2006; pp. 399–416. [Google Scholar]
- Giorgi, F.; Lionello, P. Climate change projections for the Mediterranean region. Glob. Planet. Chang. 2008, 63, 90–104. [Google Scholar] [CrossRef]
- Lorenzoni, M.; Barocco, R.; Carosi, A.; Giannetto, D.; Pompei, L. The fish fauna of the Apennine streams related to changes in the regime of wet depositions. Biol. Ambient. 2014, 28, 67–73. [Google Scholar]
- Moran, P.A.P. A mathematical theory of animal trapping. Biometrika 1951, 38, 307–311. [Google Scholar] [CrossRef]
- Zippin, C. An evaluation of the removal method of estimating animal populations. Biometrics 1956, 12, 163–189. [Google Scholar] [CrossRef]
- Agenzia per la Protezione dell’Ambiente e per i servizi Tecnici (APAT); Consiglio Nazionale delle Ricerche (CNR); Istituto di Ricerca sulle Acque (IRSA). Analytical Methods for Water. Manuals and Guidelines. 2003. Available online: http://www.isprambiente.gov.it/it/pubblicazioni/manuali-e-linee-guida/metodi-analitici-per-le-acque (accessed on 26 October 2019).
- American Public Health Association (APHA); American Waters Works Association (AWWA); Water Pollution Control Federation (WPCF). Standard Methods for the Examination of Water and Wastewater, 20th ed.; Clesceri, L.S., Greenberg, A.E., Eaton, A.D., Eds.; American Public Health Association: Washington, DC, USA, 1998. [Google Scholar]
- American Public Health Association (APHA); American Waters Works Association (AWWA); Water pollution Control Federation (WPCF). Standard Methods for the Examination of Water and Wastewater, 21st ed.; Greenberg, A.E., Clesceri, L.S., Eaton, A.D., Eds.; American Public Health Association: Washington, DC, USA, 2005. [Google Scholar]
- Ghetti, P.F. I macroinvertebrati Nell’analisi Di Qualità Dei Corsi D’acqua; Bertelli: Trento, Italy, 1986. [Google Scholar]
- Le Cren, E.D. The length relationship and seasonal cycle in gonad weight and condition in the perch (Perca fluviatilis). J. Anim. Ecol. 1951, 20, 210–218. [Google Scholar] [CrossRef] [Green Version]
- Ricker, W.E. Computation and interpretation of biological statistics of fish populations. J. Fish. Res. Board Can. 1975, 191, 1–382. [Google Scholar]
- Blackwell, B.G.; Brown, M.L.; Willis, D.W. Relative weight (Wr) status and current use in fisheries assessment and management. Rev. Fish. Sci. 2000, 81, 1–44. [Google Scholar] [CrossRef]
- Ter Braak, C.J.F. Canonical correspondence analysis: A new eigenvector technique for multivariate direct gradient analysis. Ecology 1986, 67, 1167–1179. [Google Scholar] [CrossRef] [Green Version]
- Lepš, J.; Šmilauer, P. Multivariate Analysis of Ecological Data; University of South Bohemia: Ceské Budejovice, Czech Republic, 1999. [Google Scholar]
- Brown, M.L.; Austen, D.J. Data management and statistical techniques. In Fisheries Techniques; Murphy, B.R., Willis, D.W., Eds.; American Fisheries Society: Bethesda, MD, USA, 1996; pp. 17–61. [Google Scholar]
- Eby, L.A.; Helmy, O.; Holsinger, L.M.; Young, M.K. Evidence of Climate-Induced Range Contractions in Bull Trout Salvelinus confluentus in a Rocky Mountain Watershed, U.S.A. PLoS ONE 2014, 9, e98812. [Google Scholar] [CrossRef]
- MacKenzie, D.I.; Nichols, J.D.; Lachman, G.B.; Droege, S.J.; Royle, A.; Langtimm, C.A. Estimating site occupancy rates when detection probabilities are less than one. Ecology 2002, 83. [Google Scholar] [CrossRef]
- Budy, P.; Conner, M.M.; Salant, N.L.; Macfarlane, W.W. An occupancy-based quantification of the highly imperiled status of desert fishes of the southwestern United States. Conserv. Biol. 2015, 29. [Google Scholar] [CrossRef] [PubMed]
- Burnham, K.P.; Anderson, D.R. Model Selection and Inference: An Information-Theoretic Approach; Springer: New York, NY, USA, 1998. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach; Springer: New York, NY, USA, 2003. [Google Scholar]
- Arnold, T. Uninformative parameters and model selection using Akaike’s Information Criterion. J. Wildl. Manag. 2010, 74, 1175–1178. [Google Scholar] [CrossRef]
- Johansson, M.L.; Dufour, B.A.; Wellband, K.W.; Corkum, L.D.; MacIsaac, H.J.; Heath, D.D. Human-mediated and natural dispersal of an invasive fish in the eastern Great Lakes. Heredity 2018, 120, 533–546. [Google Scholar] [CrossRef] [PubMed]
- Britton, J.R.; Pegg, J. Ecology of European Barbel Barbus Barbus: Implications for River, Fishery, and Conservation Management. Rev. Fish Biol. Fish. 2011, 19, 321–330. [Google Scholar] [CrossRef]
- Gutmann Roberts, C.; Hindes, A.M.; Britton, J.R. Factors influencing individual movements and behaviours of invasive European barbel Barbus barbus in a regulated river. Hydrobiologia 2019, 830, 213–228. [Google Scholar] [CrossRef] [Green Version]
- Daufresne, M.; Boët, P. Climate change impacts on structure and diversity of fish communities in rivers. Glob. Chang. Biol. 2007, 13, 2467–2478. [Google Scholar] [CrossRef]
- Closs, G.P.; Angermeier, P.L.; Darwall, W.R.T.; Balcombe, S.R. Why are freshwater fish so threatened? In Conservation of Freshwater Fishes; Closs, G.P., Krkosek, M., Olden, J.D., Eds.; Cambridge University Press: Cambridge, UK, 2016; pp. 37–69. [Google Scholar]
- Gozlan, R.E.; Andreou, D.; Asaeda, T.; Beyer, K.; Bouhadad, R.; Burnard, D.; Caiola, N.; Cakic, P.; Djikanovic, V.; Esmaeili, H.R.; et al. Pan-continental invasion of Pseudorasbora parva: Towards a better understanding of freshwater fish invasions. Fish Fish. 2010, 11, 315–340. [Google Scholar] [CrossRef]
- Sunardi, A.T.; Manatunge, J. Physiological responses of topmouth gudgeon, Pseudorasbora Parva, to predator cues and variation of current velocity. Aquat. Ecol. 2007, 41, 111–118. [Google Scholar] [CrossRef]
- Sáez-Gómez, P.; Ramos-Merchante, A.; Prenda, J. Multiscale effects on freshwater fish distribution in a highly disturbed Mediterranean-type basin: Community-level and species-level responses. Aquat. Ecol. 2020, 54, 869–887. [Google Scholar] [CrossRef]
- Lorenzoni, M.; Mearelli, M.; Ghetti, L. Native and exotic fish species in the Tiber river watershed (Umbria—Italy) and their relationship to the longitudinal gradient. Bull. Fr. Pêche Piscic. 2006, 382, 19–44. [Google Scholar] [CrossRef] [Green Version]
- Huet, M. Aperçu des relations de la pente et des populations piscicoles des eaux courantes. Schweiz. Z. Fur Hydrol. Swiss J. Hydrol. 1949, 2, 332–351. [Google Scholar]
- Matulla, C.; Schmutz, S.; Melcher, A.; Gerersdorfer, T.; Haas, P. Assessing the impact of a downscaled climate change simulation on the fish fauna in an Inner-Alpine River. Int. J. Biometeorol. 2007, 52, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Broms, K.M.; Hooten, M.B.; Johnson, D.S.; Altwegg, R.; Conquest, L.L. Dynamic occupancy models for explicit colonization processes. Ecology 2016, 97, 194–204. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Study area and location of the sampling sites. Mediterranean map adapted from Isac-Cnr 2005.
Figure 1.
Study area and location of the sampling sites. Mediterranean map adapted from Isac-Cnr 2005.

Figure 2.
Comparison of the distribution and population abundances between the periods 1998–2004 (black circles), 2005–2011 (dark grey circles), and 2012–2018 (light grey circles) for: (a) Barbus barbus; (b) Gobio gobio; (c) Padogobius bonelli; (d) Pseudorasbora parva.
Figure 2.
Comparison of the distribution and population abundances between the periods 1998–2004 (black circles), 2005–2011 (dark grey circles), and 2012–2018 (light grey circles) for: (a) Barbus barbus; (b) Gobio gobio; (c) Padogobius bonelli; (d) Pseudorasbora parva.

Figure 3.
Number of invaded sites over the three census periods for: (a) B. barbus, (b) G. gobio, (c) P. bonelli, (d) P. parva.
Figure 3.
Number of invaded sites over the three census periods for: (a) B. barbus, (b) G. gobio, (c) P. bonelli, (d) P. parva.

Figure 4.
Occupancy elevation plots for: (a) B. barbus, (b) G. gobio, (c) P. bonelli, (d) P. parva.

Figure 5.
Trend over time of the mean population density values for: (a) Barbus barbus, (b) Gobio gobio, (c) Padogobius bonelli, (d) Pseudorasbora parva. Vertical bars denote 0.95 confidence intervals.
Figure 5.
Trend over time of the mean population density values for: (a) Barbus barbus, (b) Gobio gobio, (c) Padogobius bonelli, (d) Pseudorasbora parva. Vertical bars denote 0.95 confidence intervals.

Figure 6.
Trend over time of the mean condition factor (Kn) values for: (a) Barbus barbus, (b) Gobio gobio, (c) Padogobius bonelli, (d) Pseudorasbora parva. Vertical bars denote 0.95 confidence intervals.
Figure 6.
Trend over time of the mean condition factor (Kn) values for: (a) Barbus barbus, (b) Gobio gobio, (c) Padogobius bonelli, (d) Pseudorasbora parva. Vertical bars denote 0.95 confidence intervals.

Figure 7.
Ecological requirements: trend over time of the mean values of: dissolved oxygen for Barbus barbus (a); conductivity and dissolved oxygen for Padogobius bonelli (b); average current speed, chlorides, and sulphates for Pseudorasbora parva (c). For each species, only the occurring sites in the census periods 1998–2004, 2005–2011, 2012–2018 were considered. Only statistically significant results at ANOVA are shown.
Figure 7.
Ecological requirements: trend over time of the mean values of: dissolved oxygen for Barbus barbus (a); conductivity and dissolved oxygen for Padogobius bonelli (b); average current speed, chlorides, and sulphates for Pseudorasbora parva (c). For each species, only the occurring sites in the census periods 1998–2004, 2005–2011, 2012–2018 were considered. Only statistically significant results at ANOVA are shown.

Figure 8.
Canonical Correspondence Analysis results: biplot of fish and environmental variables for the census periods 1998–2004, 2005–2011, 2012–2018.
Figure 8.
Canonical Correspondence Analysis results: biplot of fish and environmental variables for the census periods 1998–2004, 2005–2011, 2012–2018.

Figure 9.
The effects of elevation on the density of (a) B. barbus, (b) G. gobio, (c) P. bonelli and (d) P. parva over the census periods 1998–2004, 2005–2011, 2012–2018. Solid lines indicate significant results from the general linear model (GLM).
Figure 9.
The effects of elevation on the density of (a) B. barbus, (b) G. gobio, (c) P. bonelli and (d) P. parva over the census periods 1998–2004, 2005–2011, 2012–2018. Solid lines indicate significant results from the general linear model (GLM).

Figure 10.
Barbus barbus: effects of fragmentation degree (a), and water temperature (b) on colonization probabilities from the best informative models. Dashed gray lines indicate upper and lower 95% confidence intervals.
Figure 10.
Barbus barbus: effects of fragmentation degree (a), and water temperature (b) on colonization probabilities from the best informative models. Dashed gray lines indicate upper and lower 95% confidence intervals.

Figure 11.
Gobio gobio: effects of average current speed on colonization probabilities from the best informative models. Dashed gray lines indicate upper and lower 95% confidence intervals.
Figure 11.
Gobio gobio: effects of average current speed on colonization probabilities from the best informative models. Dashed gray lines indicate upper and lower 95% confidence intervals.

Figure 12.
Padogobius bonelli: effects of elevation on colonization probabilities, from the best informative models. Dashed gray lines indicate upper and lower 95% confidence intervals.
Figure 12.
Padogobius bonelli: effects of elevation on colonization probabilities, from the best informative models. Dashed gray lines indicate upper and lower 95% confidence intervals.

Figure 13.
Pseudorasbora parva: effects of elevation on occupancy from the best informative models. Dashed gray lines indicate upper and lower 95% confidence intervals.
Figure 13.
Pseudorasbora parva: effects of elevation on occupancy from the best informative models. Dashed gray lines indicate upper and lower 95% confidence intervals.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Canonical and correlation coefficients of environmental variables with axis. p < 0.05 is in bold.
Table 1.
Canonical and correlation coefficients of environmental variables with axis. p < 0.05 is in bold.
| AX1 | p | AX2 | p | |
|---|---|---|---|---|
| Altitude (m a.s.l.) | 0.855 | 0.001 | 0.110 | 0.343 |
| Average current speed (m s−1) | −0.101 | 0.387 | 0.436 | 0.001 |
| BOD5 (mg L−1) | −0.510 | 0.001 | 0.148 | 0.203 |
| Cl (mg L−1) | −0.247 | 0.032 | −0.432 | 0.001 |
| Dissolved oxygen (mg L−1) | −0.323 | 0.004 | −0.798 | 0.001 |
| Distance from the source (km) | −0.208 | 0.071 | 0.339 | 0.003 |
| EBI Quality Class (units) | −0.037 | 0.748 | 0.207 | 0.073 |
| Flow rate (m3 s−1) | −0.068 | 0.558 | 0.532 | 0.001 |
| NNH3 (mg L−1) | −0.280 | 0.014 | 0.125 | 0.283 |
| NNO3 (mg L−1) | −0.242 | 0.035 | 0.010 | 0.935 |
| River fragmentation degree (units) | 0.246 | 0.032 | 0.179 | 0.122 |
| Water temperature (°C) | −0.004 | 0.976 | 0.092 | 0.430 |
| Watershed area (km−2) | −0.316 | 0.005 | 0.438 | 0.001 |
Table 2.
Overall occupancy (ψ), colonization (ɣ), extirpation (ε) and detection (p) probabilities (±SE) for Barbus barbus, Gobio gobio, Padogobius bonelli and Pseudorasbora parva.
Table 2.
Overall occupancy (ψ), colonization (ɣ), extirpation (ε) and detection (p) probabilities (±SE) for Barbus barbus, Gobio gobio, Padogobius bonelli and Pseudorasbora parva.
| Species | Occupancy (ψ) | Colonization (ɣ) | Extirpation (ε) | Detection (p) |
|---|---|---|---|---|
| Barbus barbus | 0.14 ± 0.04 | 0.20 ± 0.05 | 0.05 ± 0.01 | 0.79 ± 0.07 |
| Gobio gobio | 0.21 ± 0.04 | 0.12 ± 0.04 | 0.00 ± 0.00 | 0.86 ± 0.06 |
| Padogobius bonelli | 0.15 ± 0.04 | 0.10 ± 0.03 | 0.00 ± 0.00 | 0.85 ± 0.06 |
| Pseudorasbora parva | 0.31 ± 0.07 | 0.26 ± 0.07 | 0.18 ± 0.10 | 0.74 ± 0.06 |
Table 3.
Probability of detection (±standard error (SE)) across the four time periods for Barbus barbus, Gobio gobio, Padogobius bonelli and Pseudorasbora parva.
Table 3.
Probability of detection (±standard error (SE)) across the four time periods for Barbus barbus, Gobio gobio, Padogobius bonelli and Pseudorasbora parva.
| Species | Detection Probability | ||
|---|---|---|---|
| Time Period 1 1998–2004 | Time Period 2 2005–2011 | Time Period 3 2012–2018 | |
| Barbus barbus | 0.53 ± 0.06 | 0.79 ± 0.08 | 0.80 ± 0.08 |
| Gobio gobio | 0.42 ± 0.01 | 0.83 ± 0.09 | 0.88 ± 0.08 |
| Padogobius bonelli | 0.67 ± 0.11 | 0.72 ± 0.11 | 0.93 ± 0.07 |
| Pseudorasbora parva | 0.53 ± 0.06 | 0.79 ± 0.08 | 0.71 ± 0.08 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Carosi, A.; Ghetti, L.; Lorenzoni, M. The Role of Climate Changes in the Spread of Freshwater Fishes: Implications for Alien Cool and Warm-Water Species in a Mediterranean Basin. Water 2021, 13, 347. https://doi.org/10.3390/w13030347
AMA Style
Carosi A, Ghetti L, Lorenzoni M. The Role of Climate Changes in the Spread of Freshwater Fishes: Implications for Alien Cool and Warm-Water Species in a Mediterranean Basin. Water. 2021; 13(3):347. https://doi.org/10.3390/w13030347
Chicago/Turabian StyleCarosi, Antonella, Lucia Ghetti, and Massimo Lorenzoni. 2021. "The Role of Climate Changes in the Spread of Freshwater Fishes: Implications for Alien Cool and Warm-Water Species in a Mediterranean Basin" Water 13, no. 3: 347. https://doi.org/10.3390/w13030347
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.