
Start-Up of Anammox SBR from Non-Specific Inoculum and Process Acceleration Methods by Hydrazine


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Setup
2.2. Inoculum and Operation Conditions in the Bioreactor
2.3. Batch Testing
2.4. Chemical Analyses and Calculations
3. Results
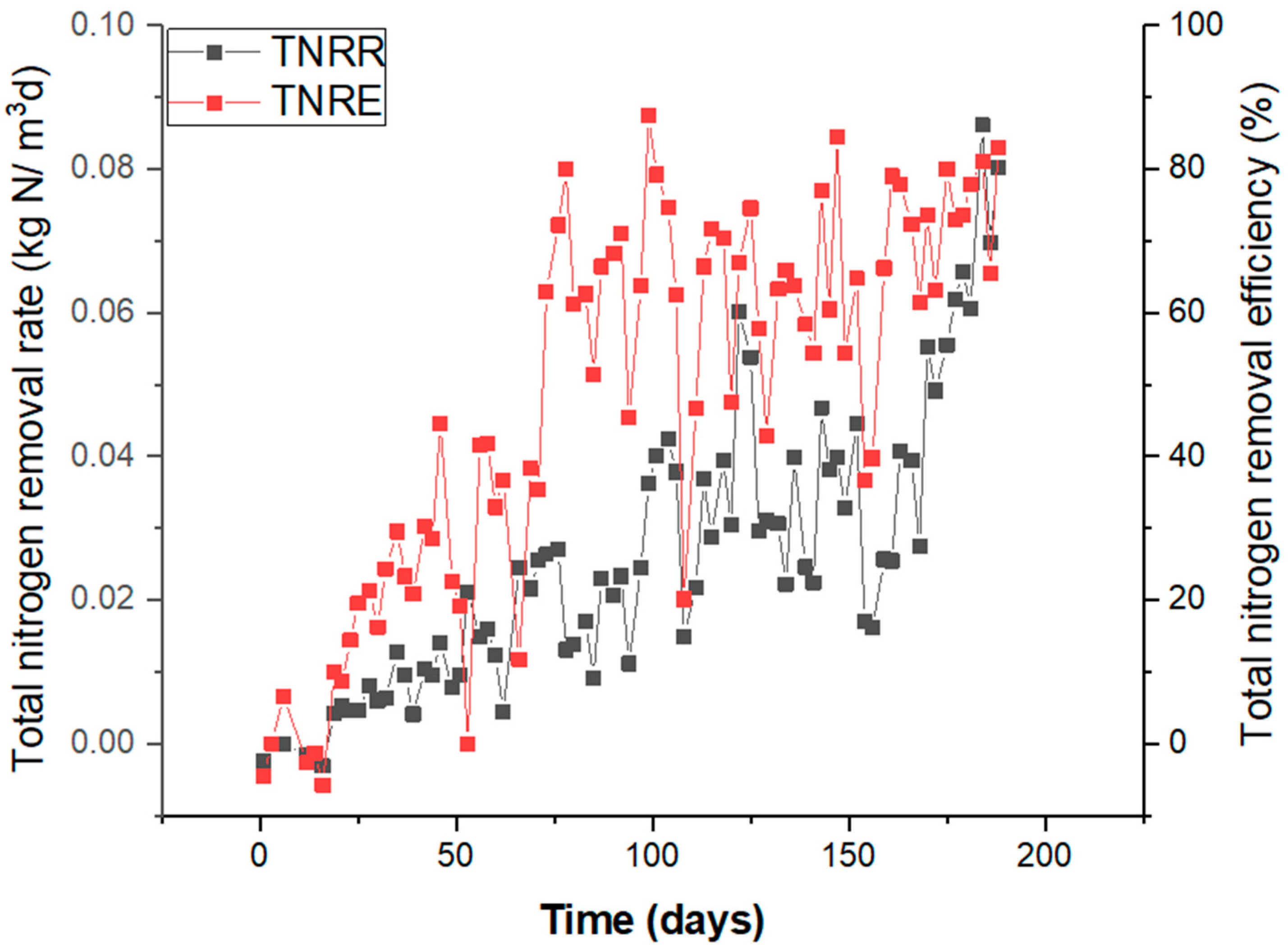
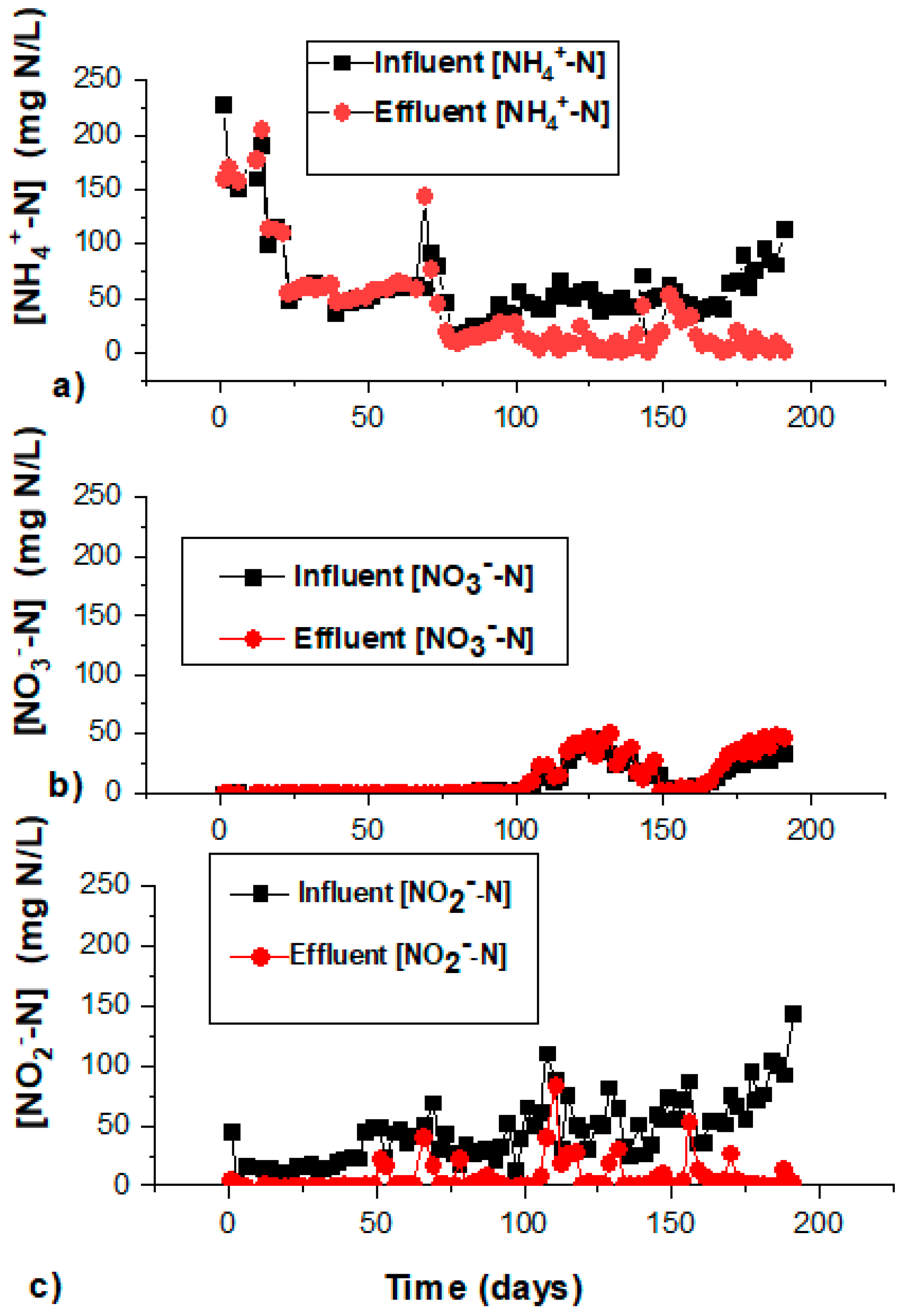
3.1. Bioreactor Performance and Division into Three Phases
3.2. Adaptation/Activation Phase
3.3. High Salinity Phase
3.4. Optimum Salinity Phase
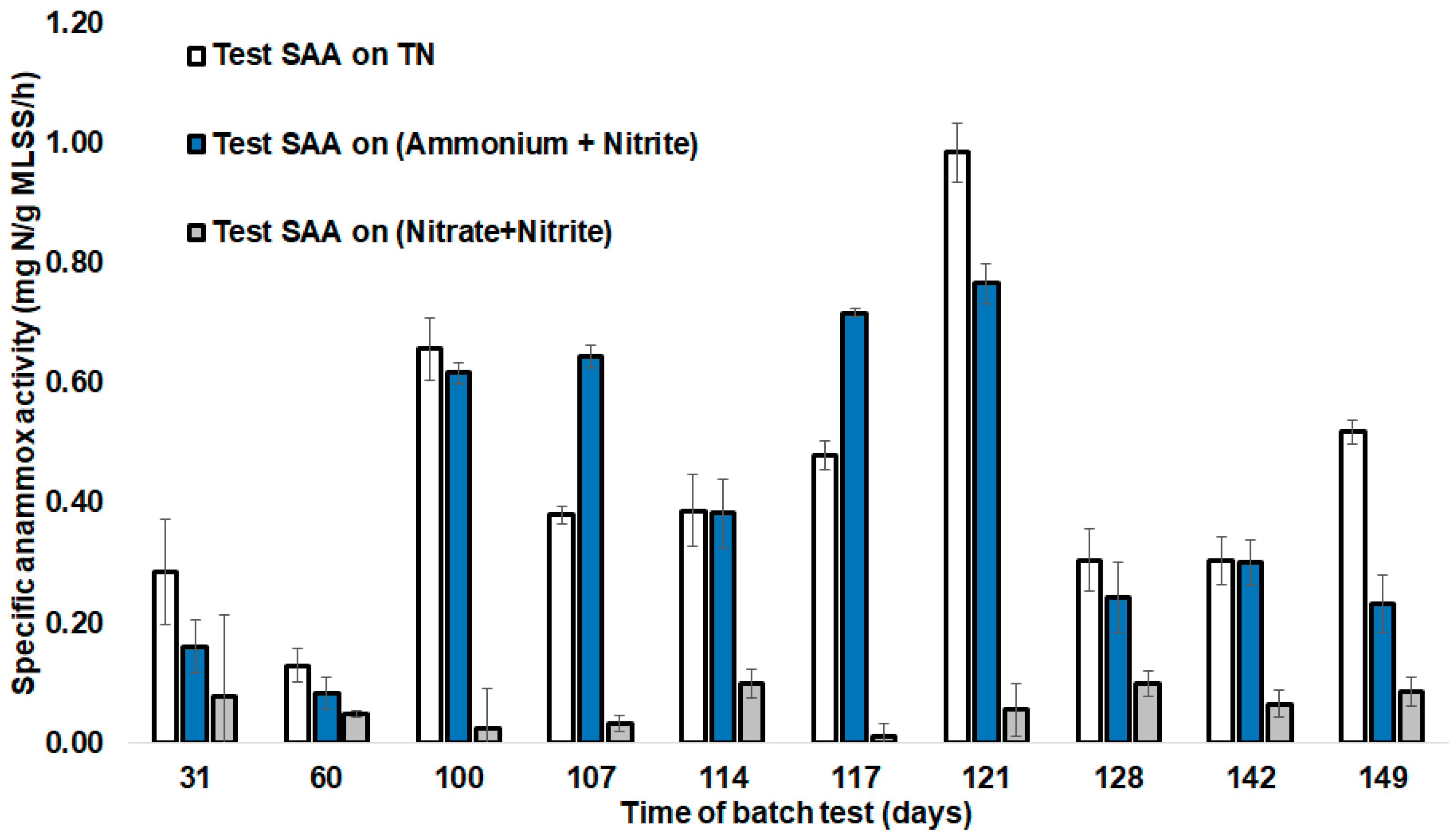
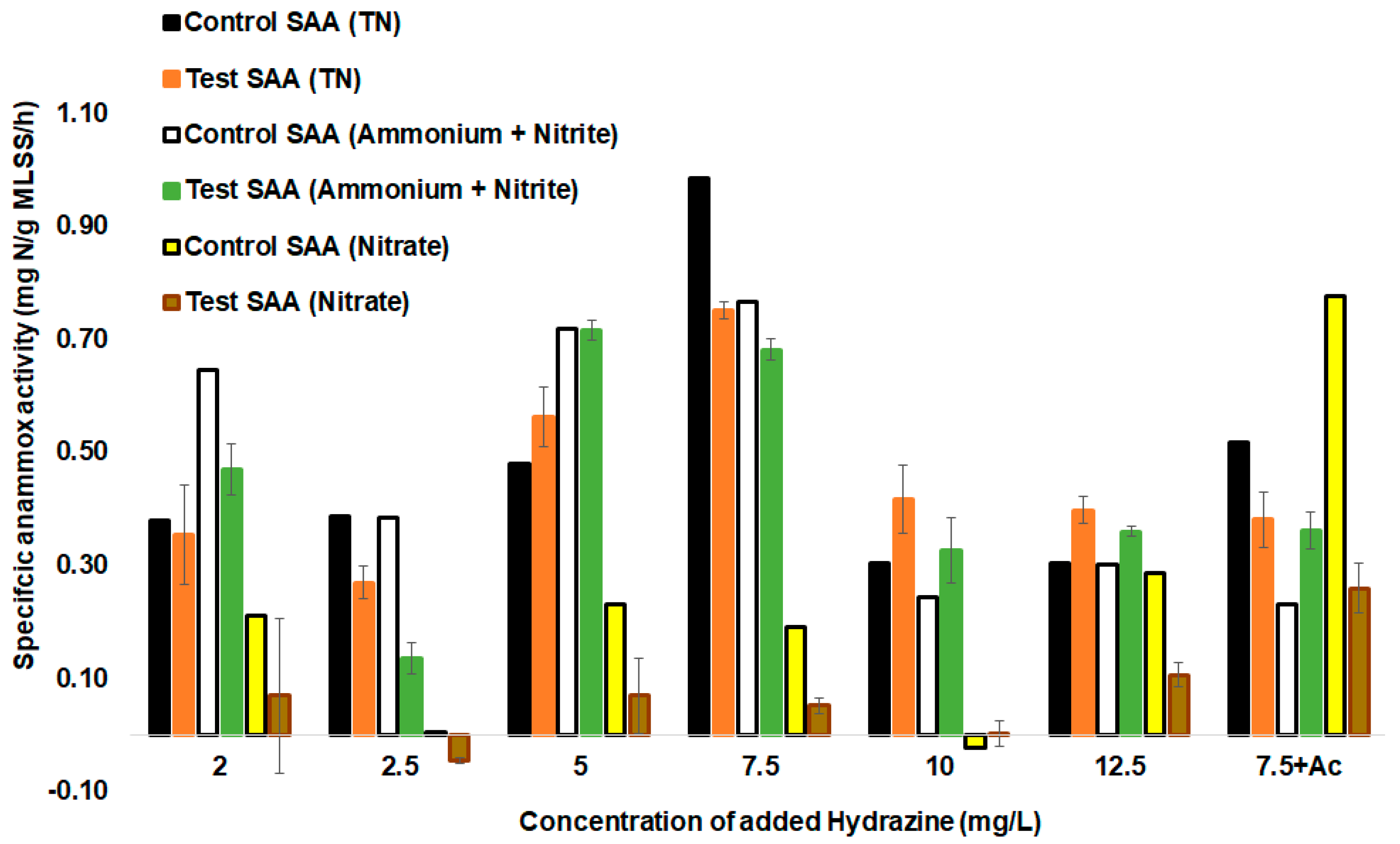
3.5. Batch Cycle Analysis
3.6. Main Challenges for the Treatment of Real Wastewater by Anammox Process
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AnAOB | Anaerobic ammonium oxidizing bacteria |
| Anammox | Anaerobic ammonium oxidation |
| AOB | Ammonium oxidizing bacteria |
| AMO | Ammonium monooxygenase |
| ATP | Adenosine triphosphate |
| CANON | Complete autotrophic nitrogen removal over nitrite |
| COD | Chemical oxygen demand |
| DEMON | Deammonification |
| DO | Dissolved oxygen |
| HAO | hydroxylamine oxidoreductase |
| HDH | Hydrazine dehydrogenase |
| HZO | Hydrazine oxidoreductase |
| HZS | Hydrazine synthase |
| MLSS | Mixed liquor suspended solid |
| NOB | Nitrite oxidizing bacteria |
| TNRR | Total nitrogen removal rate |
| TNLR | Total nitrogen loading rate |
| TNRE | Total nitrogen removal efficiency |
| NirS/NirK | Nitrite oxidoreductases (k- copper, S- Iron containing) |
| PCR | Polymerase chain reaction |
| RBC | Rotating biological contractor |
| rRNA | Ribosomal Ribonucleic acid |
| SAA | Specific anammox activity |
| SBR | Sequence Batch Reactor |
| SNAP | Single-stage nitrogen removal using anammox and partial nitrification |
| UASB | Upflow Anaerobic Sludge Blanket |
| WWTP | Wastewater treatment plant |
References
- Bulgariu, D.; Axinte, O.; Badescu, I.S.; Stroe, C.; Neacsu, V.; Bulgariu, L. Evolution of Trophic Parameters from Amara Lake. Environ. Eng. Manag. J. 2015, 14, 559–565. [Google Scholar] [CrossRef]
- Mandel, A.; Zekker, I.; Jaagura, M.; Tenno, T. Enhancement of anoxic phosphorus uptake of denitrifying phosphorus removal process by biomass adaption. Int. J. Environ. Sci. Technol. 2019, 16, 5965–5978. [Google Scholar] [CrossRef]
- Mulder, A.; Van De Graaf, A.; Robertson, L.; Kuenen, J. Anaerobic ammonium oxidation discovered in a denitrifying fluidized bed reactor. FEMS Microbiol. Ecol. 1995, 16, 177–183. [Google Scholar] [CrossRef]
- Hu, Z.; Lotti, T.; De Kreuk, M.; Kleerebezem, R.; Van Loosdrecht, M.; Kruit, J.; Jetten, M.S.M.; Kartal, B. Nitrogen Removal by a Nitritation-Anammox Bioreactor at Low Temperature. Appl. Environ. Microbiol. 2013, 79, 2807–2812. [Google Scholar] [CrossRef] [Green Version]
- Van Der Star, W.R.; Abma, W.R.; Blommers, D.; Mulder, J.-W.; Tokutomi, T.; Strous, M.; Picioreanu, C.; Van Loosdrecht, M.C. Startup of reactors for anoxic ammonium oxidation: Experiences from the first full-scale anammox reactor in Rotterdam. Water Res. 2007, 41, 4149–4163. [Google Scholar] [CrossRef]
- Shalini, S.S.; Joseph, K. Start-up of the SHARON and ANAMMOX process in landfill bioreactors using aerobic and anaerobic ammonium oxidising biomass. Bioresour. Technol. 2013, 149, 474–485. [Google Scholar] [CrossRef] [PubMed]
- Strous, M.; Heijnen, J.J.; Kuenen, J.G.; Jetten, M.S.M. The sequencing batch reactor as a powerful tool for the study of slowly growing anaerobic ammonium-oxidizing microorganisms. Appl. Microbiol. Biotechnol. 1998, 50, 589–596. [Google Scholar] [CrossRef]
- Kartal, B.; De Almeida, N.M.; Maalcke, W.J.; Camp, H.J.O.D.; Jetten, M.S.; Keltjens, J.T. How to make a living from anaerobic ammonium oxidation. FEMS Microbiol. Rev. 2013, 37, 428–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chamchoi, N.; Nitisoravut, S. Anammox enrichment from different conventional sludges. Chemosphere 2007, 66, 2225–2232. [Google Scholar] [CrossRef]
- Quan, Z.-X.; Rhee, S.-K.; Zuo, J.-E.; Yang, Y.; Bae, J.-W.; Park, J.R.; Lee, S.-T.; Park, Y.-H. Diversity of ammonium-oxidizing bacteria in a granular sludge anaerobic ammonium-oxidizing (anammox) reactor. Environ. Microbiol. 2008, 10, 3130–3139. [Google Scholar] [CrossRef]
- Zekker, I.; Rikmann, E.; Tenno, T.; Kroon, K.; Seiman, A.; Loorits, L.; Fritze, H.; Tuomivirta, T.; Vabamäe, P.; Raudkivi, M.; et al. Start-up of low-temperature anammox in UASB from mesophilic yeast factory anaerobic tank inoculum. Environ. Technol. 2014, 36, 214–225. [Google Scholar] [CrossRef] [PubMed]
- Macé, S.; Mata-Alvarez, J. Utilization of SBR Technology for Wastewater Treatment: An Overview. Ind. Eng. Chem. Res. 2002, 41, 5539–5553. [Google Scholar] [CrossRef]
- Laureni, M.; Weissbrodt, D.G.; Villez, K.; Robin, O.; De Jonge, N.; Rosenthal, A.; Wells, G.; Nielsen, J.L.; Morgenroth, E.; Joss, A. Biomass segregation between biofilm and flocs improves the control of nitrite-oxidizing bacteria in mainstream partial nitritation and anammox processes. Water Res. 2019, 154, 104–116. [Google Scholar] [CrossRef] [PubMed]
- Zekker, I.; Raudkivi, M.; Artemchuk, O.; Rikmann, E.; Priks, H.; Jaagura, M.; Tenno, T. Mainstream-sidestream wastewater switching promotes anammox nitrogen removal rate in organic-rich, low-temperature streams. Environ. Technol. 2020, 1–10. [Google Scholar] [CrossRef]
- Windey, K.; De Bo, I.; Verstraete, W. Oxygen-limited autotrophic nitrification–denitrification (OLAND) in a rotating biological contactor treating high-salinity wastewater. Water Res. 2005, 39, 4512–4520. [Google Scholar] [CrossRef]
- Jin, R.-C.; Yang, G.-F.; Yu, J.-J.; Zheng, P. The inhibition of the Anammox process: A review. Chem. Eng. J. 2012, 197, 67–79. [Google Scholar] [CrossRef]
- Cho, S.; Kambey, C.; Nguyen, V.K. Performance of Anammox Processes for Wastewater Treatment: A Critical Review on Effects of Operational Conditions and Environmental Stresses. Water 2019, 12, 20. [Google Scholar] [CrossRef] [Green Version]
- Lotti, T.; Van Der Star, W.; Kleerebezem, R.; Lubello, C.; Van Loosdrecht, M. The effect of nitrite inhibition on the anammox process. Water Res. 2012, 46, 2559–2569. [Google Scholar] [CrossRef]
- Zekker, I.; Kroon, K.; Rikmann, E.; Tenno, T.; Tomingas, M.; Vabamäe, P.; Vlaeminck, S.E.; Tenno, T. Accelerating effect of hydroxylamine and hydrazine on nitrogen removal rate in moving bed biofilm reactor. Biodegradation 2012, 23, 739–749. [Google Scholar] [CrossRef]
- Schalk, J.; Oustad, H.; Kuenen, J.; Jetten, M.S. The Anaerobic Oxidation of Hydrazine: A Novel Reaction in Microbial Nitrogen Metabolism. FEMS Microbiol. Lett. 1998, 158, 61–67. [Google Scholar] [CrossRef]
- Ganesan, S.; Vadivelu, V. Effect of external hydrazine addition on anammox reactor start-up time. Chemosphere 2019, 223, 668–674. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.-B.; Cai, Q.; Zhang, D.; Xiao, P.-Y.; Lu, P.-L. The enhancement of completely autotrophic nitrogen removal over nitrite (CANON) by N 2 H 4 addition. Bioresour. Technol. 2013, 146, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Xiang, T.; Gao, D. Comparing two hydrazine addition strategies to stabilize mainstream deammonification: Performance and microbial community analysis. Bioresour. Technol. 2019, 289, 121710. [Google Scholar] [CrossRef] [PubMed]
- Tenno, T.; Rikmann, E.; Uiga, K.; Zekker, I.; Mashirin, A.; Tenno, T. A novel proton transfer model of the closed equilibrium system H2O–CO2–CaCO3–NHX. Proc. Est. Acad. Sci. 2018, 4017, 2. [Google Scholar] [CrossRef]
- Tenno, T.; Rikmann, E.; Zekker, I. Modelling the solubility of sparingly soluble compounds depending on their particles size. Proc. Est. Acad. Sci. 2018, 67, 300. [Google Scholar] [CrossRef]
- Zekker, I.; Bhowmick, G.D.; Priks, H.; Nath, D.; Rikmann, E.; Jaagura, M.; Tenno, T.; Tämm, K.; Ghangrekar, M.M. ANAMMOX-denitrification biomass in microbial fuel cell to enhance the electricity generation and nitrogen removal efficiency. Biodegradation 2020, 31, 249–264. [Google Scholar] [CrossRef]
- Greenberg, A.; Clesceri, L.; Eaton, A. Standard Methods for the Examination of Water and Wastewater, 18th ed.; American Public Health Association Publications: Washington, DC, USA, 1992. [Google Scholar]
- Barros, C.M.C.; Jia, M.; Van Loosdrecht, M.C.; Volcke, E.I.; Winkler, M.K. Evaluating the potential for dissimilatory nitrate reduction by anammox bacteria for municipal wastewater treatment. Bioresour. Technol. 2017, 233, 363–372. [Google Scholar] [CrossRef]
- Shu, D.; He, Y.; Yue, H.; Gao, J.; Wang, Q.; Yang, S. Enhanced long-term nitrogen removal by organotrophic anammox bacteria under different C/N ratio constraints: Quantitative molecular mechanism and microbial community dynamics. RSC Adv. 2016, 6, 87593–87606. [Google Scholar] [CrossRef]
- Dapena-Mora, A.; Fernández, I.; Campos, J.; Mosquera-Corral, A.; Méndez, R.; Jetten, M. Evaluation of activity and inhibition effects on Anammox process by batch tests based on the nitrogen gas production. Enzym. Microb. Technol. 2007, 40, 859–865. [Google Scholar] [CrossRef]
- Oshiki, M.; Shimokawa, M.; Fujii, N.; Satoh, H.; Okabe, S. Physiological characteristics of the anaerobic ammonium-oxidizing bacterium ‘Candidatus Brocadia sinica’. Microbiology 2011, 157, 1706–1713. [Google Scholar] [CrossRef] [Green Version]
- Kartal, B.; Koleva, M.; Arsov, R.; Van Der Star, W.; Jetten, M.S.; Strous, M. Adaptation of a freshwater anammox population to high salinity wastewater. J. Biotechnol. 2006, 126, 546–553. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.; Zeng, M.; Zhu, B.; Zhang, W.; Liu, H.; Yang, L.; Wang, L. Impacts of different morphologies of anammox bacteria on nitrogen removal performance of a hybrid bioreactor: Suspended sludge, biofilm and gel beads. Chemosphere 2018, 208, 460–468. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Wang, T.; Gao, D.; Yang, F.; Zhang, G. Comparison between MBR and SBR on Anammox start-up process from the conventional activated sludge. Bioresour. Technol. 2012, 122, 78–82. [Google Scholar] [CrossRef] [PubMed]
- Hendrickx, T.L.; Wang, Y.; Kampman, C.; Zeeman, G.; Temmink, H.; Buisman, C.J.N. Autotrophic nitrogen removal from low strength waste water at low temperature. Water Res. 2012, 46, 2187–2193. [Google Scholar] [CrossRef] [PubMed]
- Hendrickx, T.L.; Kampman, C.; Zeeman, G.; Temmink, H.; Hu, Z.; Kartal, B.; Buisman, C.J.N. High specific activity for anammox bacteria enriched from activated sludge at 10 °C. Bioresour. Technol. 2014, 163, 214–221. [Google Scholar] [CrossRef] [PubMed]
- De Clippeleir, H.; Vlaeminck, S.E.; De Wilde, F.; Daeninck, K.; Mosquera, M.; Boeckx, P.; Verstraete, W.; Boon, N. One-stage partial nitritation/anammox at 15 A degrees C on pretreated sewage: Feasibility demonstration at lab-scale. Appl. Microbiol. Biot. 2013, 97, 10199–10210. [Google Scholar] [CrossRef]
- Arroyo, J.M.C.; Sun, W.; Sierra-Alvarez, R.; A Field, J. Inhibition of anaerobic ammonium oxidizing (anammox) enrichment cultures by substrates, metabolites and common wastewater constituents. Chemosphere 2013, 91, 22–27. [Google Scholar] [CrossRef]
- Bettazzi, E.; Caffaz, S.; Vannini, C.; Lubello, C. Nitrite inhibition and intermediates effects on Anammox bacteria: A batch-scale experimental study. Process. Biochem. 2010, 45, 573–580. [Google Scholar] [CrossRef]
- Strous, M.; Fuerst, J.A.; Kramer, E.H.M.; Logemann, S.; Muyzer, G.; Van De Pas-Schoonen, K.T.; Webb, R.M.T.; Kuenen, J.G.; Jetten, M.S.M. Missing lithotroph identified as new planctomycete. Nature 1999, 400, 446–449. [Google Scholar] [CrossRef]
- Xiao, P.; Lu, P.; Zhang, D.; Han, X.; Yang, Q. Effect of trace hydrazine addition on the functional bacterial community of a sequencing batch reactor performing completely autotrophic nitrogen removal over nitrite. Bioresour. Technol. 2015, 175, 216–223. [Google Scholar] [CrossRef]
- Zekker, I.; Kivirüüt, A.; Rikmann, E.; Mandel, A.; Jaagura, M.; Tenno, T.; Artemchuk, O.; Rubin, S.D.; Tenno, T. Enhanced Efficiency of Nitritating-Anammox Sequencing Batch Reactor Achieved at Low Decrease Rates of Oxidation–Reduction Potential. Environ. Eng. Sci. 2019, 36, 350–360. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Start-Up Period (Days) | NO2−-N/NH4+-N |
|---|---|
| 66–99 | 2.52 (±0.51)/1 |
| 101–125 | 1.03 (±0.16)/1 |
| 126–145 | 1.12 (±0.12)/1 |
| 146–194 | 1.33 (±0.06)/1 |
| Bioreactor Type | Temp. (°C) | Start-Up Time (Days) | Volume of Reactor (L) | TNRR (kg N/m3/d) | Include Nitration | Sludge Type | Author |
|---|---|---|---|---|---|---|---|
| SBR/MBR | 10 | 727 | 4.2 | 0.027 | - | Mixed activated sludge | [36] |
| SBR | 30 | 345 | 5 | 0.028 | + | Start-up | [4] |
| RBC | 29 | 300 | 2.5 | 0.60 | + | Start-up | [37] |
| Gas-lift reactor | 20 | 253 | 4.5 | 0.29 | - | Anammox sludge | [35] |
| MBR | 35 | 110 | 4.8 | 0.159 | + | Conventional activated sludge | [34] |
| SBR | 30 | 193 | 20 | 0.127 | - | Start-up with activated and anaerobic sludge | Current study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zekker, I.; Artemchuk, O.; Rikmann, E.; Ohimai, K.; Dhar Bhowmick, G.; Madhao Ghangrekar, M.; Burlakovs, J.; Tenno, T. Start-Up of Anammox SBR from Non-Specific Inoculum and Process Acceleration Methods by Hydrazine. Water 2021, 13, 350. https://doi.org/10.3390/w13030350
Zekker I, Artemchuk O, Rikmann E, Ohimai K, Dhar Bhowmick G, Madhao Ghangrekar M, Burlakovs J, Tenno T. Start-Up of Anammox SBR from Non-Specific Inoculum and Process Acceleration Methods by Hydrazine. Water. 2021; 13(3):350. https://doi.org/10.3390/w13030350
Chicago/Turabian StyleZekker, Ivar, Oleg Artemchuk, Ergo Rikmann, Kelvin Ohimai, Gourav Dhar Bhowmick, Makarand Madhao Ghangrekar, Juris Burlakovs, and Taavo Tenno. 2021. "Start-Up of Anammox SBR from Non-Specific Inoculum and Process Acceleration Methods by Hydrazine" Water 13, no. 3: 350. https://doi.org/10.3390/w13030350