
Wet Meadow Plant Communities of the Alliance Trifolion pallidi on the Southeastern Margin of the Pannonian Plain

 ,
,  , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
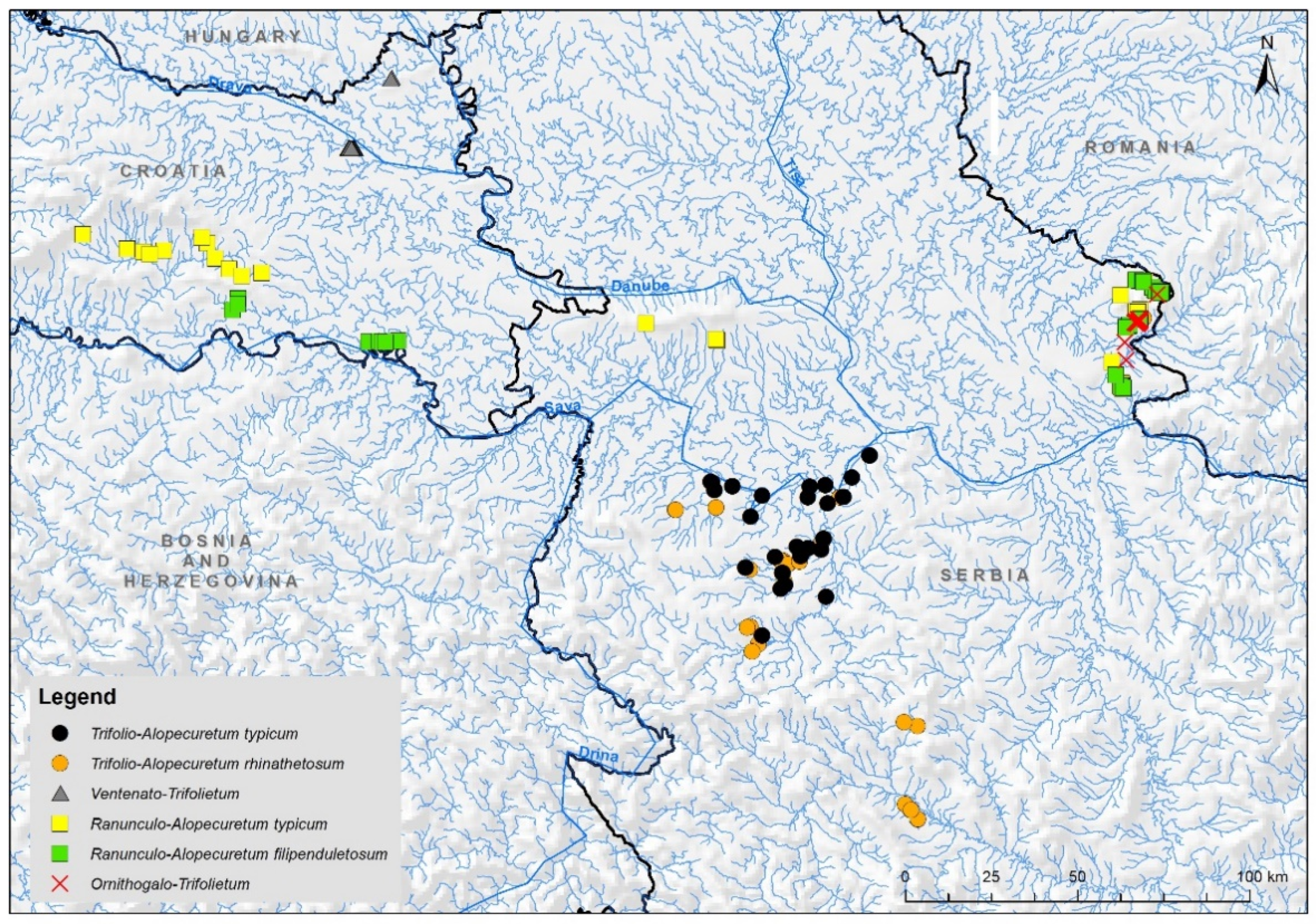
2.1. Study Area
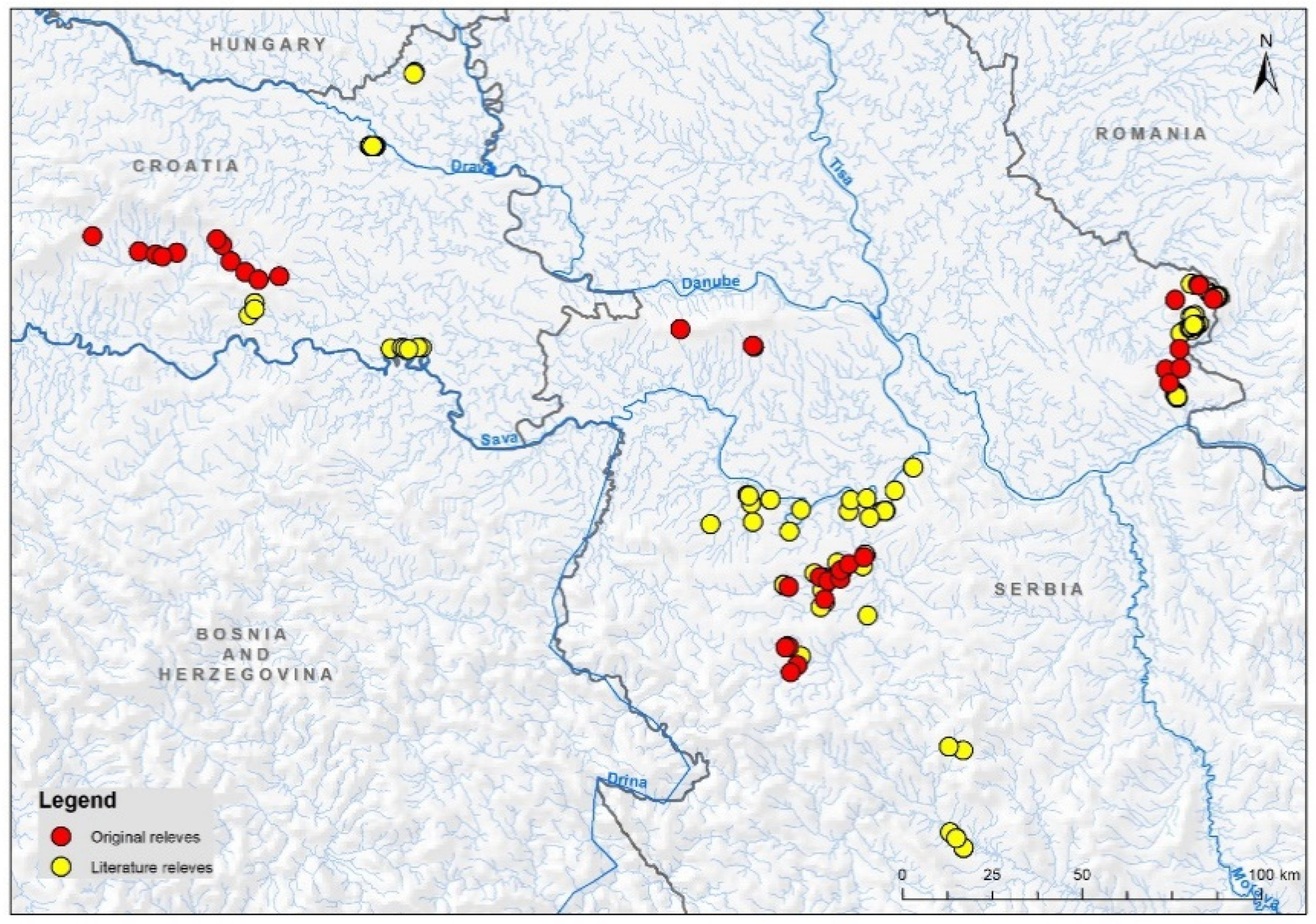
2.2. Data
2.3. Nomenclature
2.4. Numerical Analysis
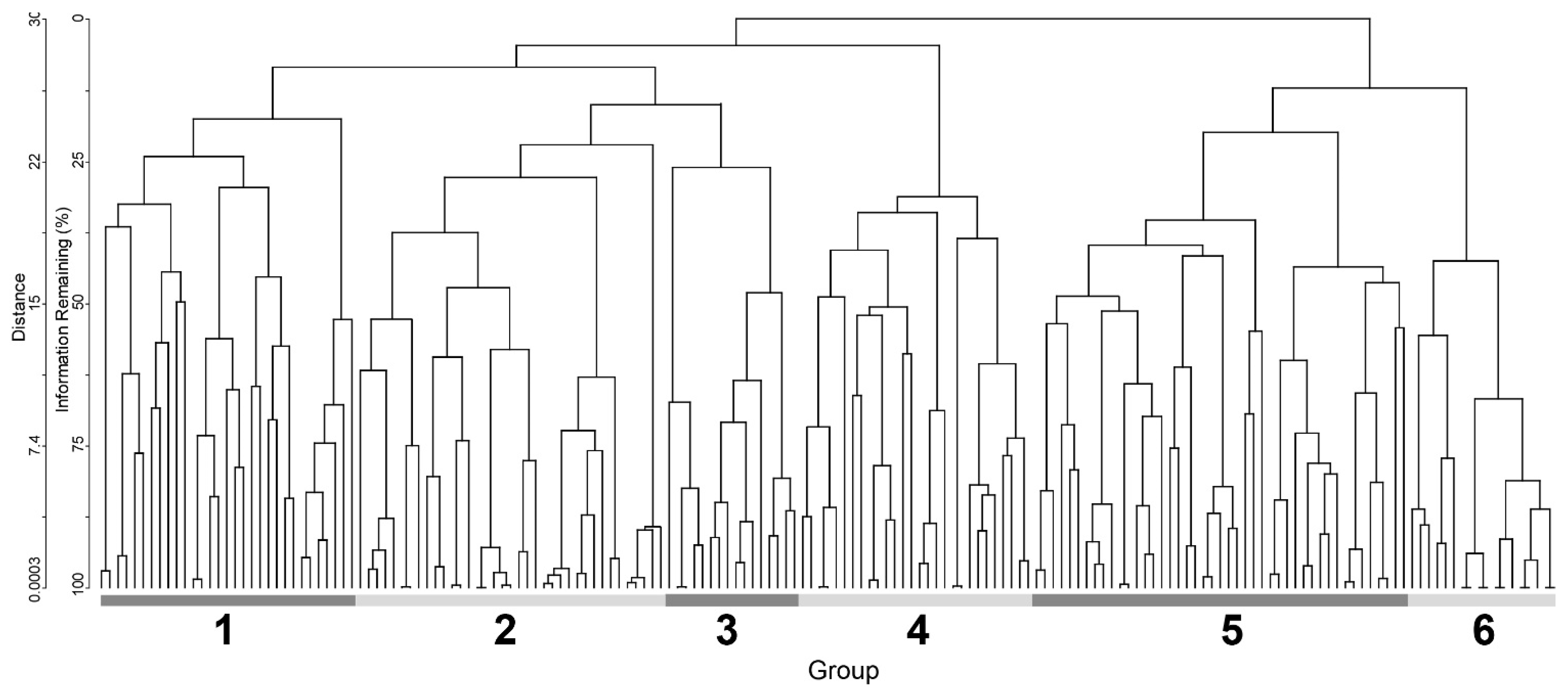
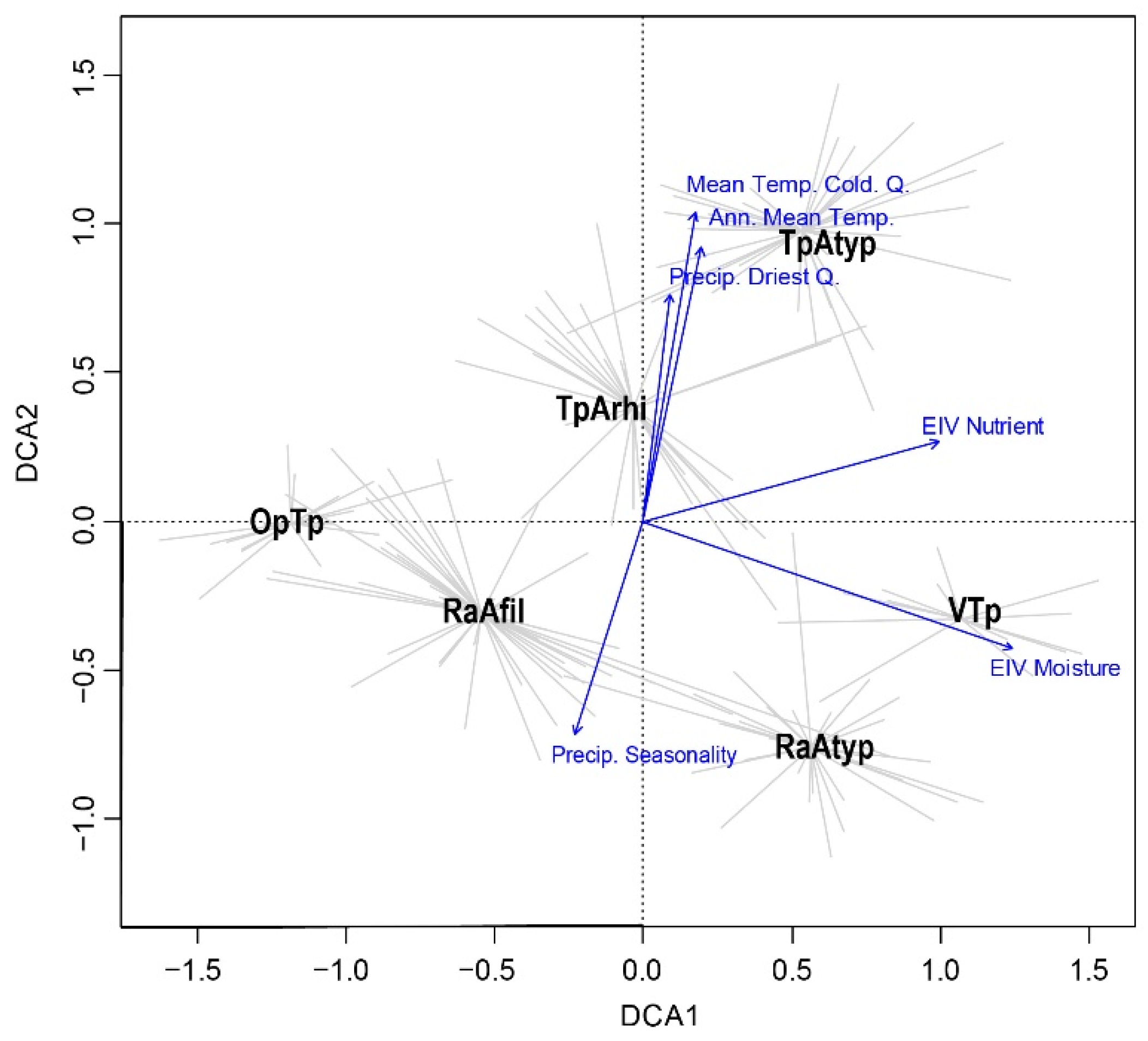
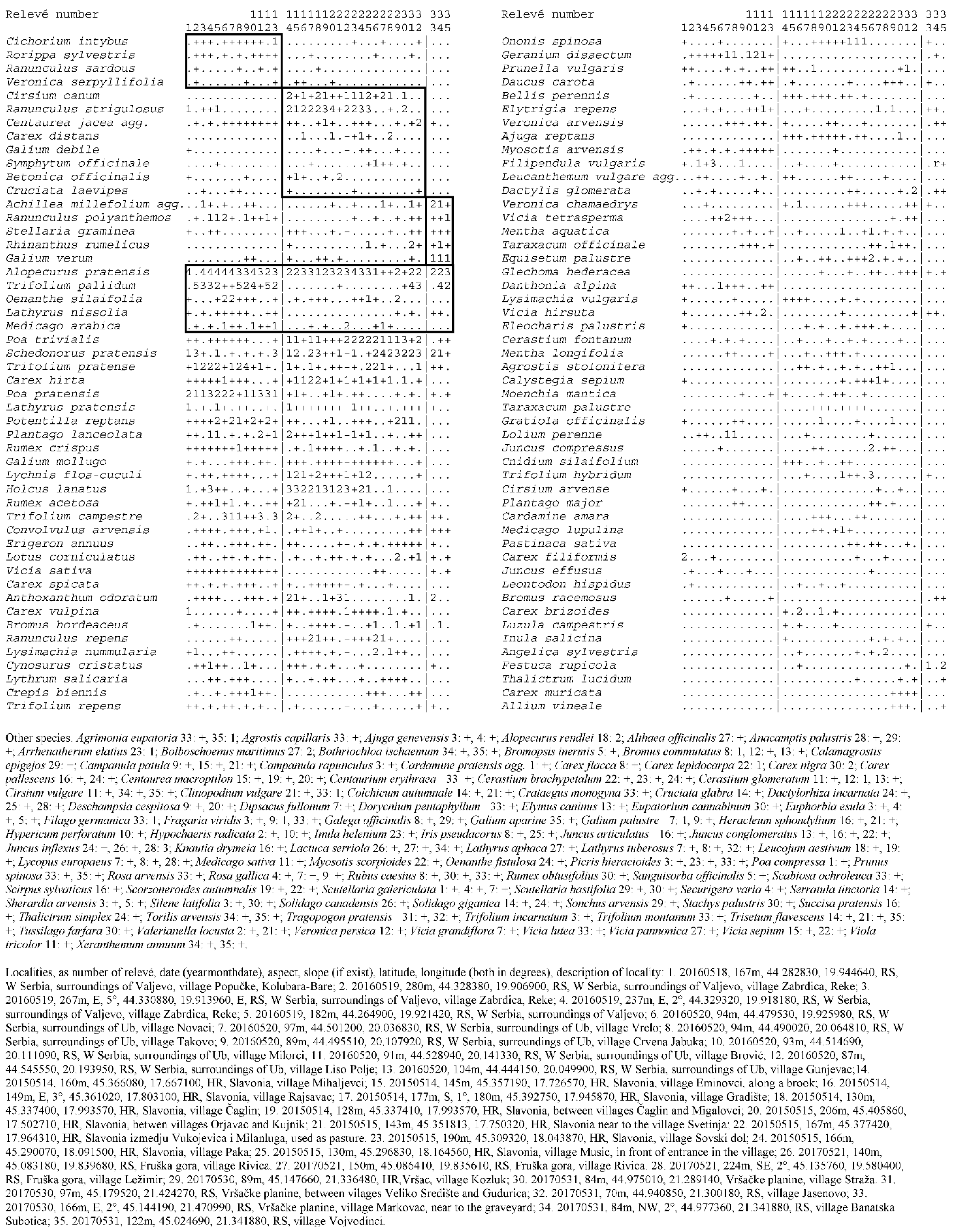
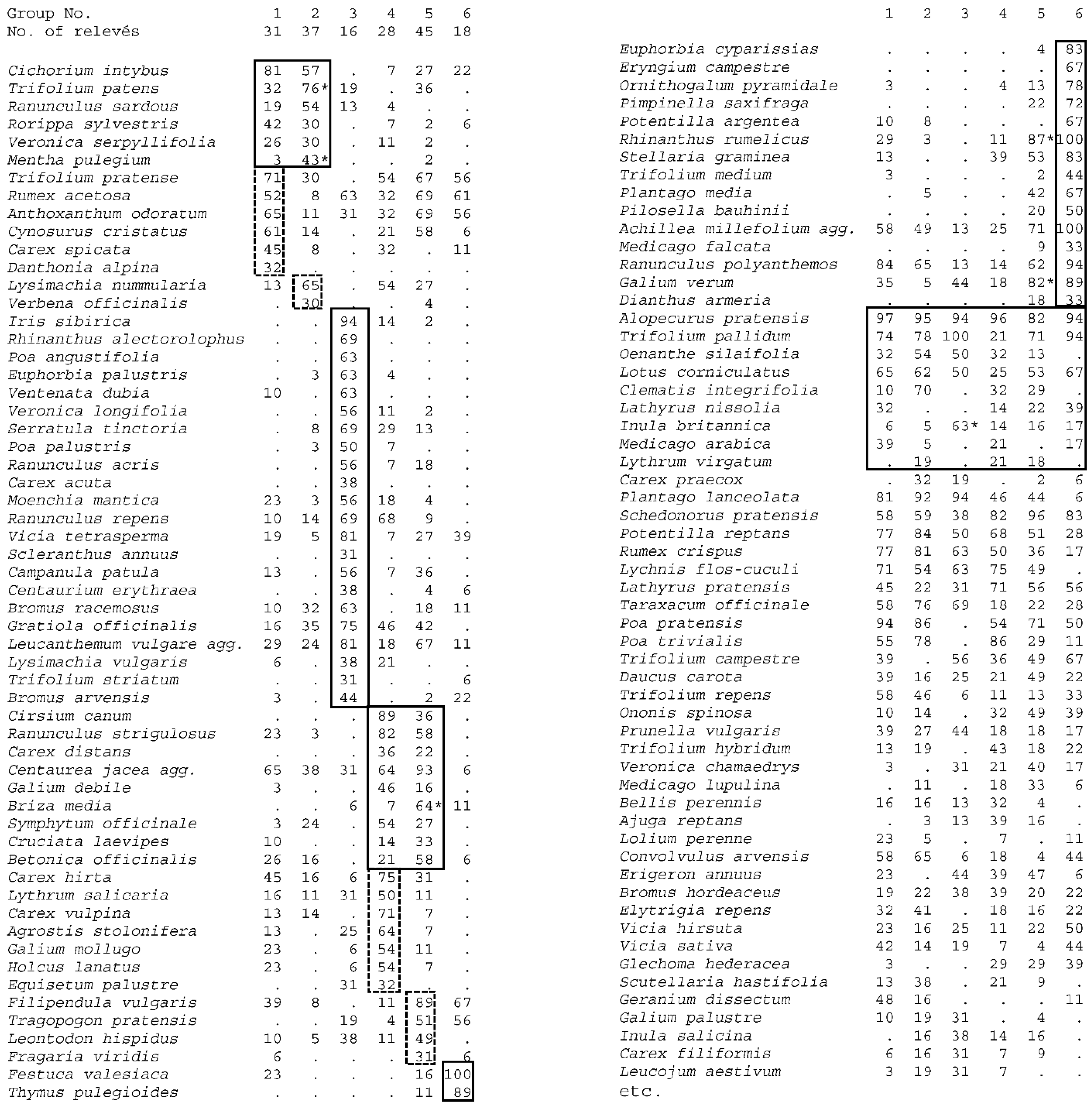
3. Results
4. Discussion
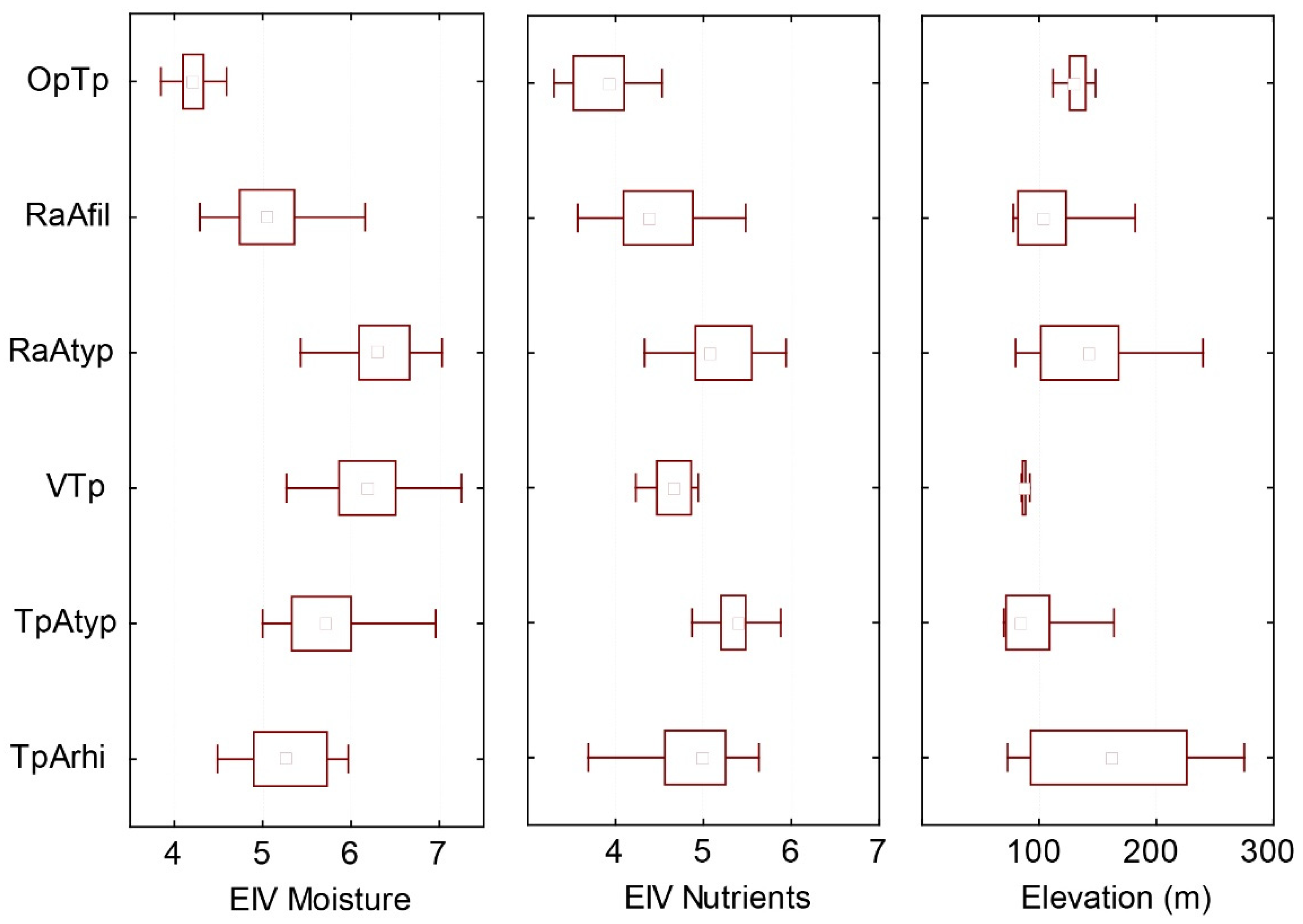
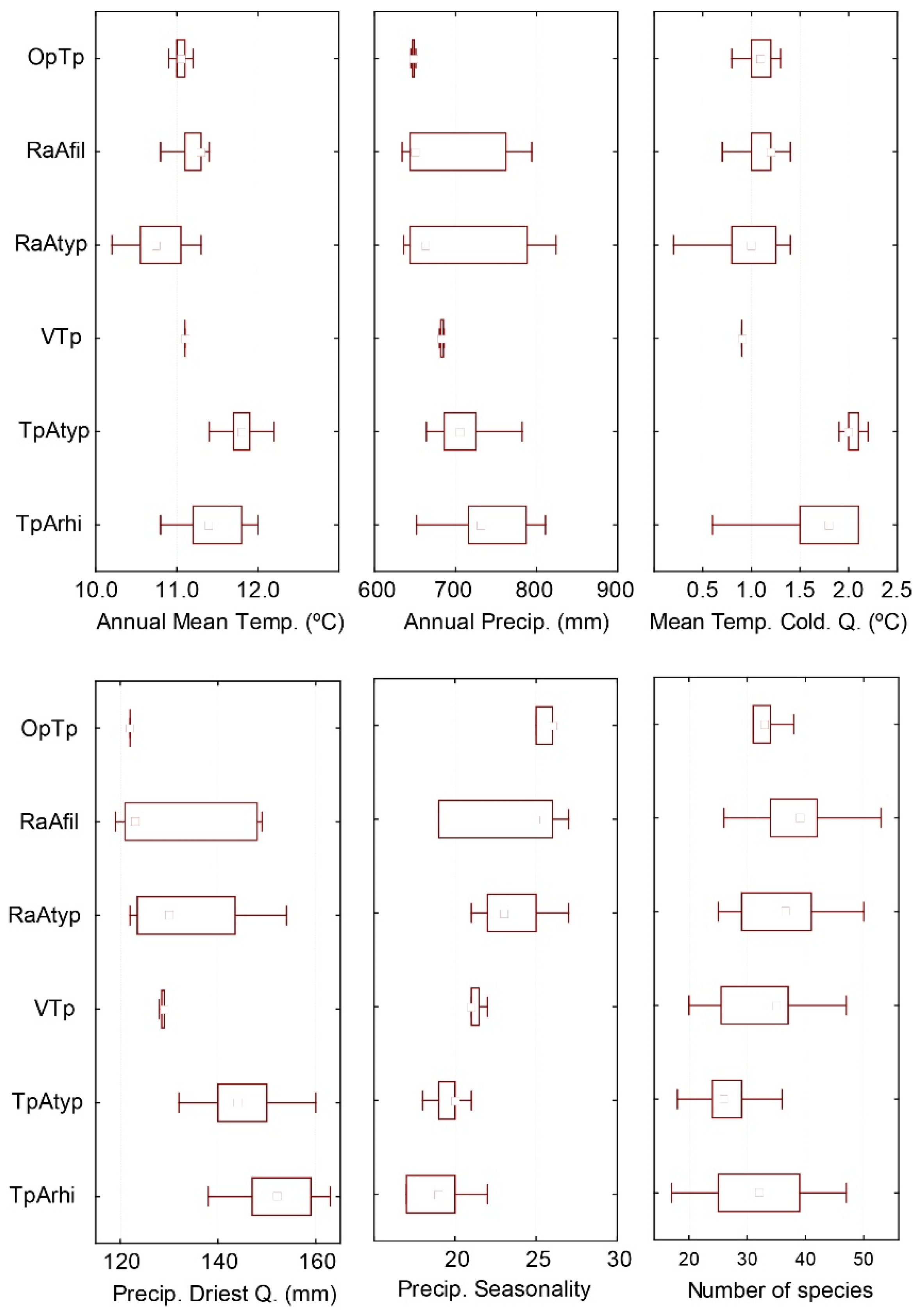
4.1. Description of Individual Plant Communities
4.1.1. Association Trifolio pallidi–Alopecuretum pratensis Cincović 1959
4.1.2. Association Ventenato dubii–Trifolietum pallidi Ilijanić 1968
4.1.3. Association Ranunuculo strigulosi–Alopecuretum pratensis Vučković ex Aćić et al. 2013 mut. Ćuk in Čarni et al. 2021
4.1.4. Association Ornithogalo pyramidale–Trifolietum pallidi Vučković ex Aćić et al. 2013
4.2. Syntaxomomic Scheme of the Studied Plant Communities
- Molinio–Arrhenatheretea Tx. 1937
- Trifolio–Hordeetalia Horvatić 1963
- Trifolion pallidi Ilijanić 1969
- Trifolio pallidi–Alopecuretum pratensis Cincović 1959
- typicum
- rhinathetosum rumelici Cincović 1959
- Ventenato–Trifolietum pallidi Ilijanić 1968
- Ranunculo strigulosi–Alopecuretum pratensis M. Vučković ex Aćić et al. 2013 mut. Ćuk in Čarni et al. 2021
- typicum
- filipenduletosum Škvorc et al. 2020
- Ornithogalo pyramidale–Trifolietum pallidi M. Vučković ex Aćić et al. 2013
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A



References
- Janssen, J.A.M.; Rodwell, J.S.; García Criado, M.; Gubbay, S.; Haynes, T.; Nieto, A.; Sanders, N.; Landucci, F.; Loidi, J.; Symank, A.; et al. European Red List of Habitats. Part 2. Terrestrial and Freshwater Habitats; European Union: Luxembourg, 2016; p. 44. [Google Scholar] [CrossRef]
- Dwire, K.A.; Kauffman, J.B.; Brookshire, E.N.J.; Baham, J.E. Plant biomass and species composition along an environmental gradient in montane riparian meadows. Oecologia 2004, 139, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.B.; Song, F.B.; Xie, Y.H. Trade-off between tolerance to drought and tolerance to flooding in three wetland plants. Wetlands 2008, 28, 866–873. [Google Scholar] [CrossRef]
- Chen, X.; Li, X.; Xie, Y.; Li, F.; Hou, Z.; Zeng, J. Combined influence of hydrological gradient and edaphic factors on the distribution of macrophyte communities in Dongting Lake wetlands (China). Wetl. Ecol. Manag. 2015, 23, 481–490. [Google Scholar] [CrossRef]
- Gaberščik, A.; Krek, J.L.; Zelnik, I. Habitat diversity along a hydrological gradient in a complex wetland results in high plant species diversity. Ecol. Eng. 2018, 118, 84–92. [Google Scholar] [CrossRef]
- Moran, J.; Skeffington, M.S.; Gormally, M. The influence of hydrological regime and grazing management on the plant communities of a karst wetland (Skealoghan turlough) in Ireland. Appl. Veg. Sci. 2008, 11, 13–24. [Google Scholar] [CrossRef]
- Dolinar, N.; Rudolf, M.; Šraj, N.; Gaberščik, A. Environmental changes affect ecosystem services of the intermittent Lake Cerknica. Ecol. Complex. 2010, 7, 403–409. [Google Scholar] [CrossRef]
- Grime, J.P. Competitive exclusion in herbaceous vegetation. Nature 1973, 242, 344–347. [Google Scholar] [CrossRef]
- Grime, J.P. Plant Strategies Vegetation Processes and Ecosystem Properties, 2nd ed.; Wiley: Chichester, UK, 2001; p. 417. [Google Scholar]
- Ilijanić, L. Typologisch-geographische Gliederung der Niederungswiesen Nordkroatiens im klimatischen Zusammenhang. Acta Bot. Croat. 1963, 22, 119–132. [Google Scholar]
- Cincović, T. Tipovi livada u Posavini. Zb. Rad. Poljopr. Fak. Beograd. 1956, 4, 31–56. [Google Scholar]
- Cincović, T. Livadska vegetacija u rečnim dolinama zapadne Srbije. Zb. Rad. Poljopr. Fak. Beograd. 1959, 7, 1–62. [Google Scholar]
- Ilijanić, L. Das Trifolion pallidi, ein neuer Verband der Ordnung Trifolio-Hordeetalia H-ić. Acta Bot. Croat. 1969, 28, 151–160. [Google Scholar]
- Mucina, L.; Bültmann, H.; Dierßen, K.; Theurillat, J.P.; Raus, T.; Čarni, A.; Šumberová, K.; Willner, W.; Dengler, J.; Gavilán, R.G.; et al. Vegetation of Europe: Hierarchical floristic classification system of vascular plant, bryophyte, lichen, and algal communities. Appl. Veg. Sci. 2016, 19 (Suppl. 1), 3–264. [Google Scholar] [CrossRef]
- Purger, D. Subterranean clover (Trifolium subterraneum L.) in communties in the Great Hungraian Plain: Questions of coenology, nomenclature and syntaxonomy. Acta Bot. Hung. 2006, 46, 399–413. [Google Scholar] [CrossRef]
- Zeidler, H. Das Alopecurion utriculati, ein neuer Verband Balkanischer Wiesengesellschaften. Vegetatio 1954, 5–6, 292–301. [Google Scholar] [CrossRef]
- Jovanović, R. Tipovi dolinskih livada Jasenice. Arch. Biol. Sci. 1957, 9, 1–14. [Google Scholar]
- Rodríguez-Rojo, M.P.; Jiménez-Alfaro, B.; Jandt, U.; Bruelheide, H.; Rodwell, J.S.; Schaminée, J.H.J.; Perrin, P.M.; Kącki, Z.; Willner, W.; Fernández-González, F.; et al. Diversity of lowland hay meadows and pastures in Western and Central Europe. Appl. Veg. Sci. 2017, 20, 702–719. [Google Scholar] [CrossRef]
- Goncharenko, I.; Kozyr, M.; Senchylo, O. Classification of the floodplain meadows of the Seym and the Dnieper river valleys in the north-eastern part of Ukraine. Biologia 2020, 75, 53–70. [Google Scholar] [CrossRef]
- Deák, B.; Valkó, O.; Török, P.; Tóthmérész, B. Solonetz meadow vegetation (Beckmannion eruciformis) in East-Hungary—An alliance driven by moisture and salinity. Tuexenia 2014, 34, 187–203. [Google Scholar] [CrossRef]
- Zaliberová, M.; Škodová, I. Potentillion anserinae. In Rastlinné Spoločenstvá Slovenska. 5. Travinno-Bylinná Vegetácia; Hegedüšová Vantarová, K., Škodová, I., Eds.; Veda: Bratislava, Slovakia, 2014; pp. 331–350. [Google Scholar]
- Botta-Dukát, Z.; Chytrý, M.; Hájková, P.; Havlová, M. Vegetation of lowland wet meadows along a climatic continentality gradient in Central Europe. Preslia 2005, 77, 89–111. [Google Scholar]
- Škvorc, Ž.; Ćuk, M.; Zelnik, I.; Franjić, J.; Igić, R.; Ilić, M.; Krstonošić, D.; Vukov, D.; Čarni, A. Diversity of wet and mesic grasslands along a climatic gradient on the southern margin of the Pannonian Basin. Appl. Veg. Sci. 2020, 23, 676–697. [Google Scholar] [CrossRef]
- Borhidi, A.; Kevey, B.; Lendvai, G. Plant Communities of Hungary; Akadémiai Kiadó: Budapest, Hungary, 2012; p. 544. [Google Scholar]
- Chytrý, M.; Tichý, L.; Hennekens, S.M.; Knollová, I.; Janssen, J.A.; Rodwell, J.S.; Peterka, T.; Marcenò, C.; Landucci, F.; Danielka, J.; et al. EUNIS Habitat Classification: Expert system, characteristic species combinations and distribution maps of European habitats. Appl. Veg. Sci. 2020, 23, 648–675. [Google Scholar] [CrossRef]
- European Council. Council Directive 92/43/EEC of 21 May 1992 on the Conservation of natural habitats and of wild fauna and flora. Off. J. Eur. Union 1992, 206, 7–50. [Google Scholar]
- European Commission. Interpretation Manual of European Union Habitats; European Commission, DG Environment: Brussels, Belgium, 2013; p. 144. [Google Scholar]
- Zaninović, K.; Gajić-Čapka, M.; Perčec Tadić, M.; Vučetić, N.; Milković, J.; Bajić, A.; Cindrić, L.; Cvitan, L.; Katušin, Z.; Kaučić, D.; et al. Climate Atlas of Croatia 1961–1990, 1971–2000; Državni Hidrometeorološki Zavod: Zagreb, Croatia, 2008; p. 200. [Google Scholar]
- Milovanović, B. Climate regionalization of Serbia according to Köppen climate classification. J. Geogr. Inst. Jovan Cvijic SASA 2017, 67, 103–114. [Google Scholar] [CrossRef] [Green Version]
- Stupar, V. Mesophilous Quercus frainetto dominated forests from Western Balkans. South-East Eur. For. 2020, 11, 97–110. [Google Scholar] [CrossRef]
- Vukelić, J. Šumska Vegetacija Hrvatske; Sveučilište u Zagrebu: Zagreb, Croatia, 2012; p. 403. [Google Scholar]
- Pantović, J.; Grdović, S.; Veljić, M.; Sabovljević, M.S. Biogeographic traits of the bryophyte flora of Serbia. Bot. Serbica 2019, 43, 143–150. [Google Scholar] [CrossRef] [Green Version]
- Ćalić, J.; Gaudenyi, T.; Milošević, M.V.; Štrbac, D.; Milivojević, M. Geomorphometrical method for delineation of plains-case study of the south-eastern (Serbian) segment of the Pannonian Plain. Carpathian J. Earth Environ. Sci. 2012, 7, 239–248. [Google Scholar]
- Adamović, L. Die Vegetationsverhältnisse der Balkanländer (Mösische Länder): Umfassend Serbien, Altserbien, Bulgarien, Ostrumelien, Nordthrakien und Nordmazedonien; Vegetation der Erde 11; W. Engelmann: Leipzig, Germany, 1909; p. 567. [Google Scholar]
- Oberdorfer, E. Gliederung und Umgrenzung der Mittelmeervegetation auf der Balkanhalbinsel. Ber. Geobot. Forsch. Rübel 1948, 1974, 84–111. [Google Scholar]
- Braun-Blanquet, J. Pflanzensoziologie, Grundzüge der Vegetationskunde, 3rd ed.; Springer Verlag: Wien, Austria, 1964; p. 631. [Google Scholar]
- Westhoff, V.; van der Maarel, E. The Braun-Blanquet approach. In Handbook of Vegetation Science 5, Classification and Ordination of Communities; Whittaker, R.H., Ed.; Dr. W. Junk: The Hague, The Netherlands, 1973; pp. 287–399. [Google Scholar]
- Ilijanić, L. Die Ordnung Molinetalia in der Vegetation Nordostkroatiens. Acta Bot. Croat. 1968, 26–27, 161–179. [Google Scholar]
- Cindrić, M. Biljni Pokrov Podnožja Dilj-Gore Sjeveroistočno od Slavonskog Broda. Master’s Thesis, University of Zagreb, Zagreb, Croatia, 1974. [Google Scholar]
- Rauš, Đ.; Šegulja, N.; Topić, J. Vegetacija sjeveroistočne Hrvatske. Glas. šumske pokuse 1985, 23, 223–355. [Google Scholar]
- Šegulja, N.; Topić, J. Prilog poznavanju pašnjaka istočne Slavonije. Poljopr. Znan. Smotra. 1978, 46, 91–94. [Google Scholar]
- Parabućski, S.; Stojanović, S. Oenanthe (banatica)-Alopecuretum pratensis ass. nova. Zb. Matice Srp. Prir. Nauke. 1988, 74, 71–78. [Google Scholar]
- Vučković, M. Livadska i Livadsko-Stepska Vegetacija Vršačkih Planina; Matica Srpska, Odeljenje za Prirodne Nauke: Novi Sad, Serbia, 1991; p. 95. [Google Scholar]
- Butorac, B. Vegetacija Fruškogorskog Lesnog Platoa, Monografija Fruške Gore; Matica Srpska, Odeljenje za Prirodne Nauke: Novi Sad, Serbia, 1992; p. 163. [Google Scholar]
- Hennekens, S.M.; Schaminée, J.H.J. TURBOVEG, a comprehensive data base management system for vegetation data. J. Veg. Sci. 2001, 12, 589–591. [Google Scholar] [CrossRef]
- Tichý, L. JUICE, software for vegetation classification. J. Veg. Sci. 2002, 13, 451–453. [Google Scholar] [CrossRef]
- Trinajstić, I. Biljne Zajednice Republike Hrvatske; Akademija Šumarskih Znanosti: Zagreb, Croatia, 2008; p. 179. [Google Scholar]
- Ellenberg, H.; Weber, H.E.; Düll, R.; Wirth, V.; Werner, W.; Paulißen, D. Zeigerwerte von Pflanzen in Mitteleuropa; Erich Goltze: Göttingen, Germany, 1992; p. 248. [Google Scholar]
- Fick, S.E.; Hijmans, R.J. Worldclim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Euro + Med Plantbase. Available online: https://www.emplantbase.org (accessed on 20 November 2020).
- Theurillat, J.P.; Willner, W.; Fernández-González, F.; Bültmann, H.; Čarni, A.; Gigante, D.; Mucina, L.; Weber, H.E. International Code of Phytosociological Nomenclature, 4th edition. Appl. Veg. Sci. 2020, 24. [Google Scholar] [CrossRef] [Green Version]
- Tichý, L.; Chytrý, M.; Hájek, M.; Talbot, S.S.; Botta-Dukát, Z. OptimClass: Using species-to-cluster fidelity to determine the optimal partition in classification of ecological communities. J. Veg. Sci. 2010, 21, 287–299. [Google Scholar] [CrossRef]
- Chytrý, M.; Tichý, L.; Holt, J.; Botta-Dukát, Z. Determination of diagnostic species with statistical fidelity measures. J. Veg. Sci. 2002, 13, 79–90. [Google Scholar] [CrossRef]
- Tichý, L.; Chytrý, M. Statistical determination of diagnostic species for site groups of unequal size. J. Veg. Sci. 2006, 17, 809–818. [Google Scholar] [CrossRef]
- Horvatić, S. Vegetacijska karta otoka Paga s općim pregledom vegetacijskih jedinica Hrvatskog primorja. Prir. Istraž. Jugosl. Akad. 1963, 33, 1–187. [Google Scholar]
- Šilc, U.; Aćić, S.; Škvorc, Ž.; Krstonošić, D.; Franjić, J.; Dajić Stevanović, Z. Grassland vegetation of the Molinio-Arrhenatheretea class in the NW Balkan Peninsula. Appl. Veg. Sci. 2014, 17, 591–603. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Package “vegan”. Community Ecology Package. Available online: https://cran.rproject.org/web/packages/vegan (accessed on 20 January 2020).
- Sanda, V.; Öllerer, K.; Burescu, P. Fitocenozele din Romania-Sintaxonomie, Structura, Dinamica si Evolutie; Ars Docendi: Bucureşti, Romania, 2008; p. 570. [Google Scholar]
- Tzonev, R.T.; Dimitrov, M.A.; Roussakova, V.H. Syntaxa according to the Braun-Blanquet approach in Bulgaria. Phytol. Balc. 2009, 15, 209–233. [Google Scholar]
- Karácsonyi, C. Two new meso-hygrophytic plant communities in the northern part of Dealurile de Vest (Western Hills) area (Romania). Contrib. Bot. 2011, 46, 7–15. [Google Scholar]
- Čarni, A.; Košir, P.; Karadžić, B.; Matevski, V.; Redžić, S.; Škvorc, Ž. Thermophilous deciduous forests in Southeastern Europe. Plant Biosyst. 2009, 143, 1–13. [Google Scholar] [CrossRef]
- Purger, D.; Lengyel, A.; Kevey, B.; Lendvai, G.; Horváth, A.; Tomić, Z.; Csiky, J. Numerical classification of oak forests on loess in Hungary, Croatia and Serbia. Preslia 2014, 87, 47–66. [Google Scholar]
- Zelnik, I.; Čarni, A. Plant species diversity and composition of wet grasslands in relation to environmental factors. Biodivers. Conserv. 2013, 22, 2179–2192. [Google Scholar] [CrossRef]
- Stromberg, J.C. Restoration of riparian vegetation in the south-western United States: Importance of flow regimes and fluvial dynamism. J. Arid. Environ. 2002, 49, 17–34. [Google Scholar] [CrossRef] [Green Version]
- Aćić, S.; Šilc, U.; Lakušić, D.; Vukojičić, S.; Dajić Stevanović, Z. Typification and correction of syntaxa from the class Molinio-Arrhenatheretea Tx. 1937 in Serbia. Hacquetia 2013, 12, 39–54. [Google Scholar] [CrossRef]
- Martinčič, A. Mala Flora Slovenije. Ključ za Določanje Praprotnic in Semenk, 4th ed.; Tehniška Založba Slovenije: Ljubljana, Slovenia, 2007; p. 967. [Google Scholar]
- Nikolić, T. Flora Croatica–Vaskularna flora Republike Hrvatske; Alfa d.d.: Zagreb, Croatia, 2019; Volume 4, p. 664. [Google Scholar]
- Hayek, A. Prodromus Florae Peninsulae Balcanicae. 1. Band: Pteridophyta, Gymnospermae, Dicotyledoneae (Apetalae et Choripetalae). Repertorium Speciorum Novarum Regni Vegetabilis; Verlag des Repertoriums: Dahlem bei Berlin, Germany, 1927; p. 1193, Beihefte 30. [Google Scholar]
- Josifović, M. Flora SR Srbije; Srpska Akademija Nauka i Umetnosti: Beograd, Serbia, 1970; Volume 1, p. 326. [Google Scholar]
- Hörandl, E.; von Raab-Straube, E. Ranunculeae. In Euro + Med Plantbase. Available online: http://www.europlusmed.org/cdm_dataportal/taxon/562ee98a-4851-4af1-acef-126d2a5e450f (accessed on 20 November 2020).
- Szépligeti, M.; Kőrösi, Á.; Szentirmai, I.; Házi, J.; Bartha, D.; Bartha, S. Evaluating alternative mowing regimes for conservation management of Central European mesic hay meadows: A field experiment. Plant Biosyst. 2018, 152, 90–97. [Google Scholar] [CrossRef]
- Török, P.; Janišová, M.; Kuzemko, A.; Rūsiņa, S.; Dajić Stevanović, Z. Grasslands, their threats and management in Eastern Europe. In Grasslands of the World: Diversity, Management and Conservation; Squires, V.R., Dengler, J., Hua, L., Feng, H., Eds.; CRC Press: Boca Raton, FL, USA, 2018; pp. 64–88. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environmental Variable | DCA1 | DCA2 |
|---|---|---|
| Elevation | 0.25 | |
| Latitude | 0.66 | |
| Longitude | 0.31 | |
| Annual Mean Temperature | 0.62 | |
| Temperature Seasonality | 0.38 | |
| Max Temperature of Warmest Month | 0.33 | |
| Min Temperature of Coldest Month | 0.39 | |
| Mean Temperature of Wettest Quarter | 0.30 | 0.50 |
| Mean Temperature of Warmest Quarter | 0.31 | 0.55 |
| Mean Temperature of Coldest Quarter | 0.70 | |
| Annual Precipitation | 0.30 | |
| Precipitation of Wettest Month | 0.41 | 0.44 |
| Precipitation of Driest Month | 0.34 | |
| Precipitation Seasonality | 0.54 | |
| Precipitation of Wettest Quarter | 0.40 | |
| Precipitation of Driest Quarter | 0.45 | |
| Precipitation of Warmest Quarter | 0.34 | 0.20 |
| Precipitation of Coldest Quarter | 0.55 | |
| Light EIV | 0.36 | |
| Moisture EIV | 0.89 | 0.28 |
| Nutrients EIV | 0.66 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Čarni, A.; Ćuk, M.; Zelnik, I.; Franjić, J.; Igić, R.; Ilić, M.; Krstonošić, D.; Vukov, D.; Škvorc, Ž. Wet Meadow Plant Communities of the Alliance Trifolion pallidi on the Southeastern Margin of the Pannonian Plain. Water 2021, 13, 381. https://doi.org/10.3390/w13030381
Čarni A, Ćuk M, Zelnik I, Franjić J, Igić R, Ilić M, Krstonošić D, Vukov D, Škvorc Ž. Wet Meadow Plant Communities of the Alliance Trifolion pallidi on the Southeastern Margin of the Pannonian Plain. Water. 2021; 13(3):381. https://doi.org/10.3390/w13030381
Chicago/Turabian StyleČarni, Andraž, Mirjana Ćuk, Igor Zelnik, Jozo Franjić, Ružica Igić, Miloš Ilić, Daniel Krstonošić, Dragana Vukov, and Željko Škvorc. 2021. "Wet Meadow Plant Communities of the Alliance Trifolion pallidi on the Southeastern Margin of the Pannonian Plain" Water 13, no. 3: 381. https://doi.org/10.3390/w13030381