
Plasticity in Reproductive Traits, Condition and Energy Allocation of the Non-Native Pyrenean Gudgeon Gobio lozanoi in a Highly Regulated Mediterranean River Basin

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Information
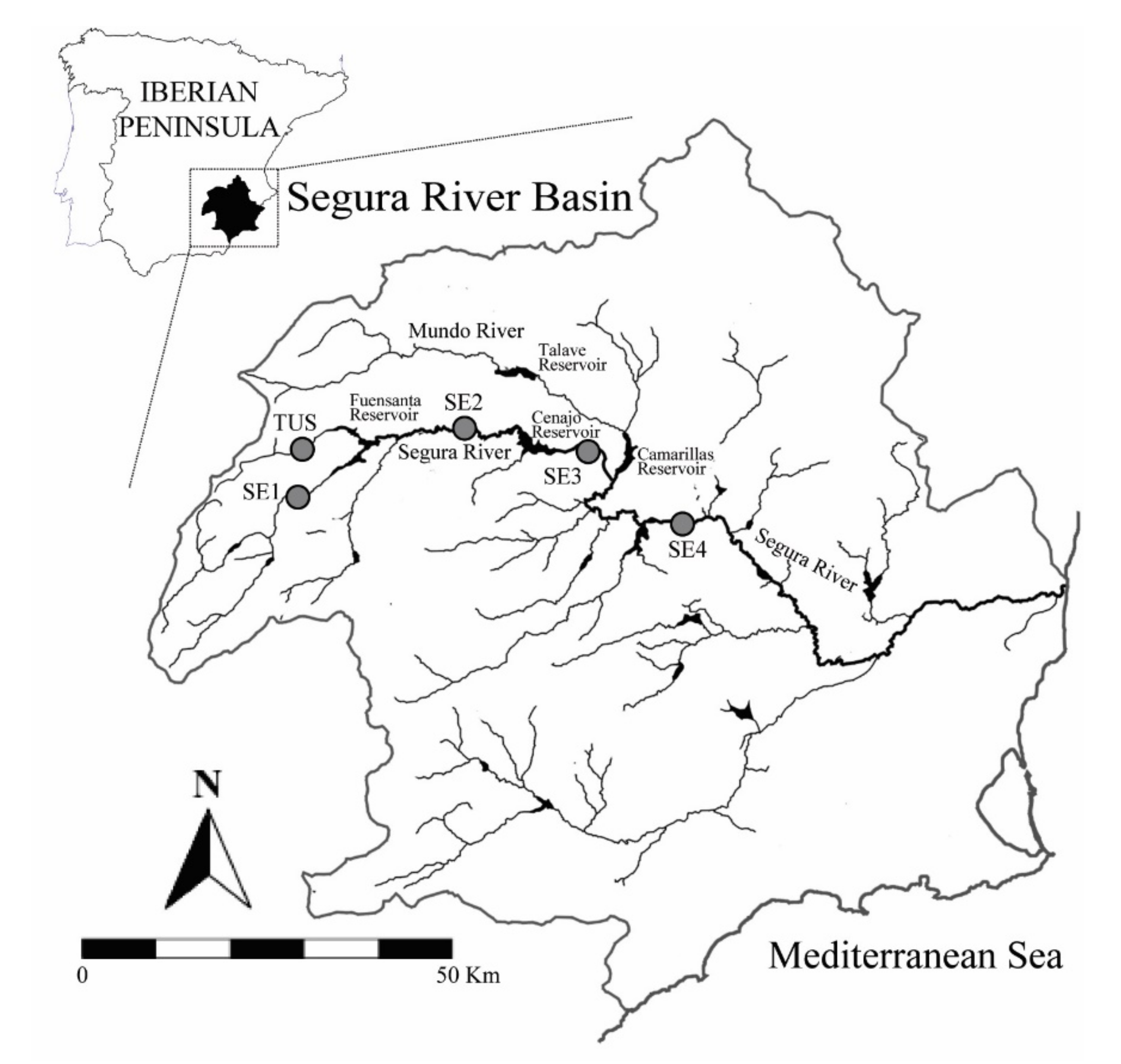
2.2. Study Area
2.3. Field Sampling and Laboratory Procedures
2.4. Statistical Analyses
3. Results
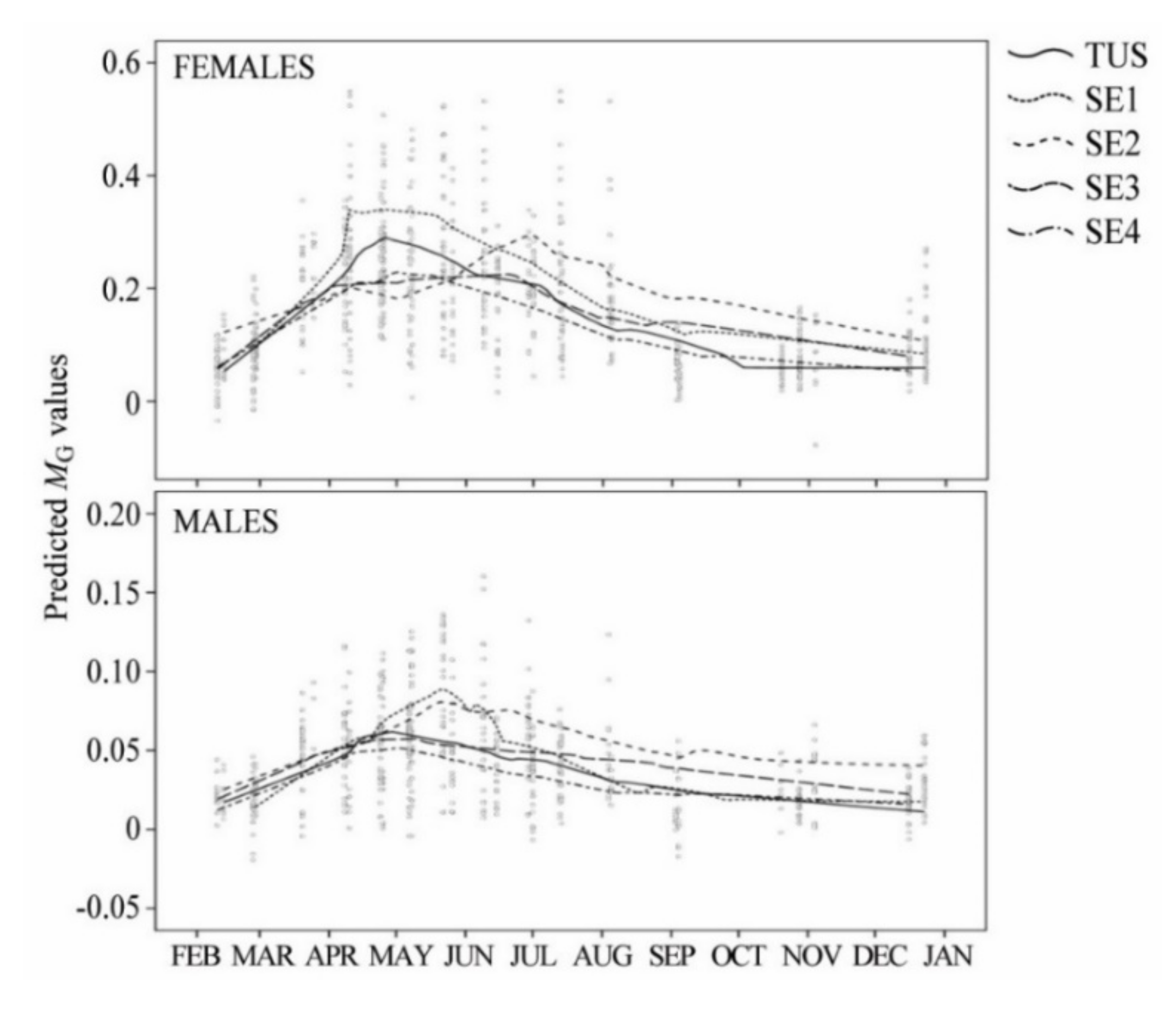
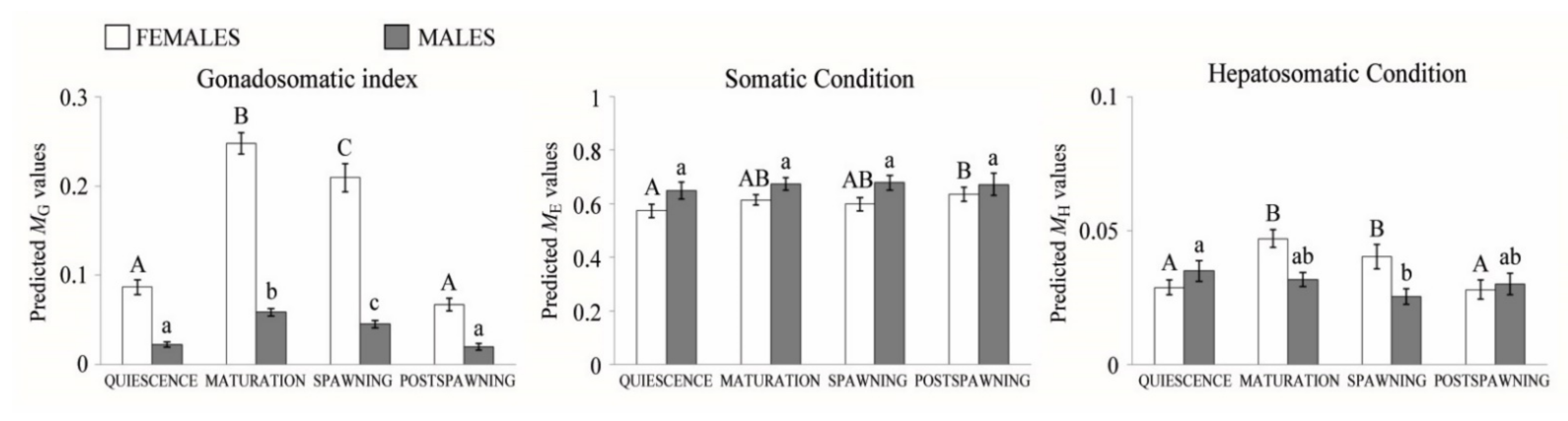
3.1. Reproductive Cycle and Temporal Variation in Fish Condition
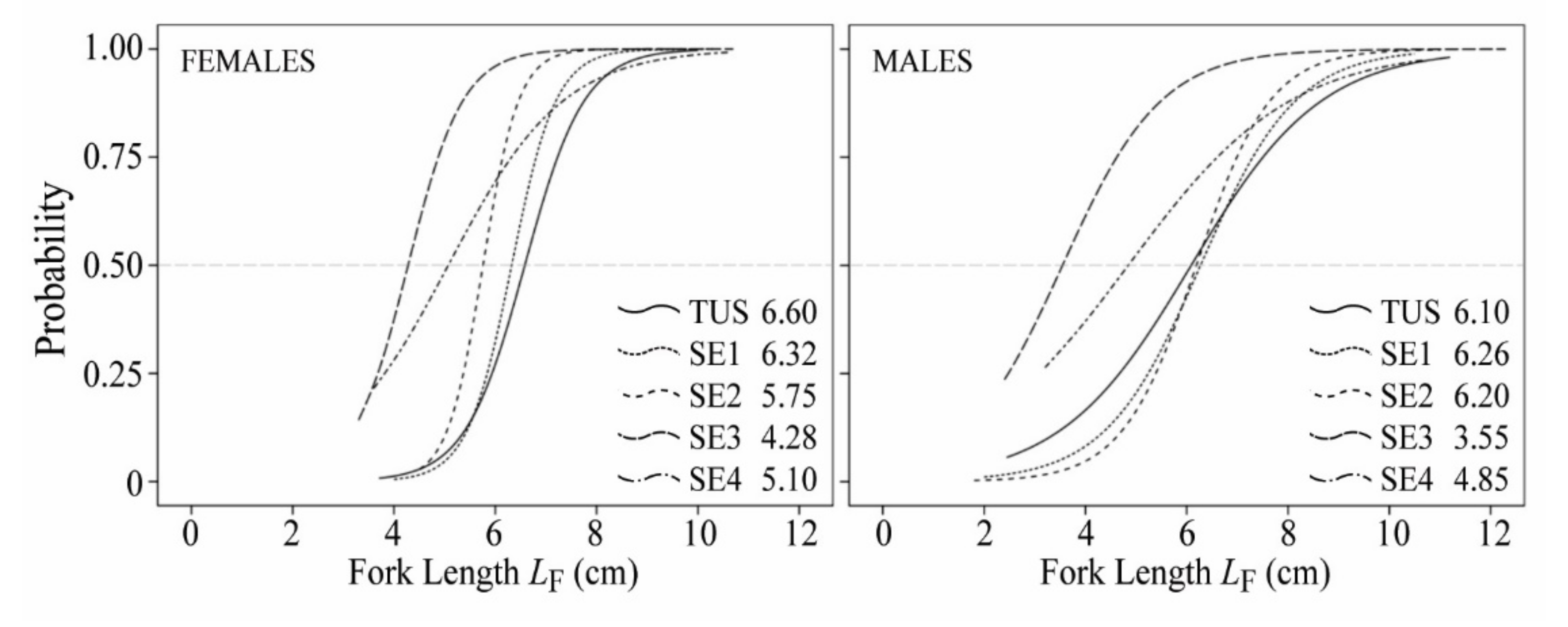
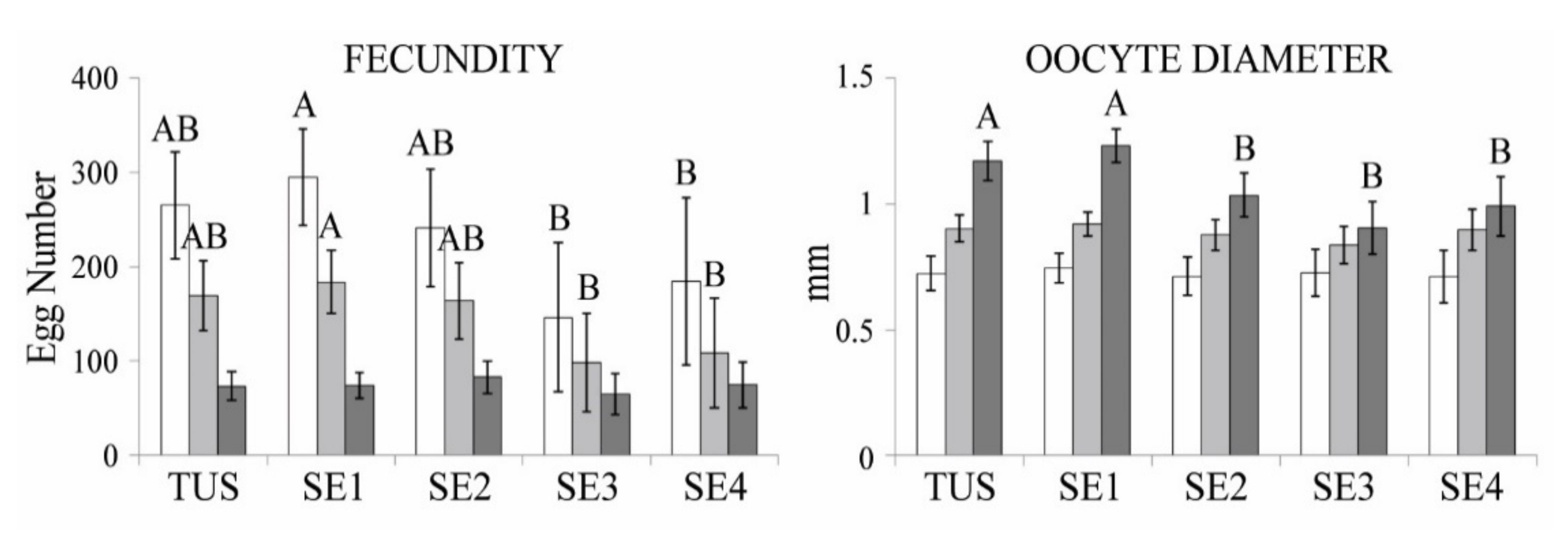
3.2. Population Structure and Reproduction Traits
3.3. Protein and Lipid Contents
3.4. Fish Metrics Relationships
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bunn, S.E.; Arthington, A.H. Basic principles and ecological consequences of altered flow regimes for aquatic biodiversity. Environ. Manag. 2002, 30, 492–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schinegger, R.; Palt, M.; Segurado, P.; Schmutz, S. Untangling the effects of multiple human stressors and their impacts on fish assemblages in European running waters. Sci. Total. Environ. 2016, 573, 1079–1088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mims, M.C.; Olden, J.D. Fish assemblages respond to altered flow regimes via ecological filtering of life history strategies. Freshw. Biol. 2013, 58, 50–62. [Google Scholar] [CrossRef]
- Oliveira, J.M.; Segurado, P.; Santos, J.M.; Teixeira, A.; Ferreira, M.T.; Cortes, R.V. Modelling stream-fish functional traits in reference conditions: Regional and local environmental correlates. PLoS ONE 2012, 7, 15–17. [Google Scholar] [CrossRef] [PubMed]
- Bergerot, B.; Hugueny, B.; Belliard, J. Relating life-history traits, environmental constraints and local extinctions in river fish. Freshw. Biol. 2015, 60, 1–13. [Google Scholar] [CrossRef]
- Mims, M.C.; Olden, J.D. Life history theory predicts fish assemblage response to hydrologic regimes. Ecol. Soc. Am. 2012, 93, 35–45. [Google Scholar] [CrossRef]
- Stearns, S.C. The Evolution of Life Histories; Oxford University Press: New York, NY, USA, 1992. [Google Scholar]
- Alexandre, C.M.; Ferreira, M.T.; Almeida, P.R. Life history of a cyprinid species in non-regulated and regulated rivers from permanent and temporary Mediterranean basins. Ecohydrology 2014, 8, 1137–1153. [Google Scholar] [CrossRef]
- Bailly, D.; Agostinho, A.A.; Suzuki, H.I. Influence of the flood regime on the reproduction of fish species with different reproductive strategies in the Cuiabá River, Upper Pantanal, Brazil. River Res. Appl. 2008, 24, 1218–1229. [Google Scholar] [CrossRef]
- Cambray, J.A.; King, J.M.; Bruwer, C. Spawning behaviour and early development of the Clanwilliam yellowfish (Barbus capensis; Cyprinidae), linked to experimental dam releases in the Olifants River, South Africa. Regul. Rivers Res. Manag. 1997, 13, 579–602. [Google Scholar] [CrossRef]
- Tan, X.; Li, X.; Lek, S.; Li, Y.-F.; Wang, C.; Li, J.; Luo, J. Annual dynamics of the abundance of fish larvae and its relationship with hydrological variation in the Pearl River. Environ. Biol. Fishes 2010, 88, 217–225. [Google Scholar] [CrossRef]
- Miranda, R.; Oscoz, J.; Leunda, P.M.; García-Fresca, C.; Escala, M.C. Effects of weir construction on fish population structure in the River Erro (North of Spain). Ann. Limnol. 2005, 41, 7–13. [Google Scholar] [CrossRef]
- Vasconcelos, L.P.; Alves, D.C.; Gomes, L.C. Fish reproductive guilds downstream of dams. J. Fish Biol. 2014, 85, 1489–1506. [Google Scholar] [CrossRef] [PubMed]
- Tedesco, P.A.; Hugueny, B.; Oberdorff, T.; Dürr, H.H.; Mérigoux, S.; De Mérona, B. River hydrological seasonality influences life history strategies of tropical riverine fishes. Oecologia 2008, 156, 691–702. [Google Scholar] [CrossRef] [PubMed]
- McBride, R.S.; Somarakis, S.; Fitzhugh, G.R.; Albert, A.; Yaragina, N.A.; Wuenschel, M.J.; Alonso-Fernández, A.; Basilone, G. Energy acquisition and allocation to egg production in relation to fish reproductive strategies. Fish Fish. 2015, 16, 23–57. [Google Scholar] [CrossRef] [Green Version]
- Henderson, B.A.; Wong, J.L.; Nepszy, S.J. Reproduction of walleye in Lake Erie: Allocation of energy. Can. J. Fish. Aquat. Sci. 1996, 53, 127–133. [Google Scholar] [CrossRef]
- Kennedy, J.; Skjæraasen, J.E.; Nash, R.D.M.; Thorsen, A.; Slotte, A.; Hansen, T.; Kjesbu, O.S. Do capital breeders like Atlantic herring (Clupea harengus) exhibit sensitive periods of nutritional control on ovary development and fecundity regulation? Can. J. Fish. Aquat. Sci. 2010, 67, 16–27. [Google Scholar] [CrossRef]
- Peebles, E.; Hall, J.; Tolley, S. Egg production by the bay anchovy Anchoa mitchilli in relation to adult and larval prey fields. Mar. Ecol. Prog. Ser. 1996, 131, 61–73. [Google Scholar] [CrossRef]
- Brown, M.L.; Murphy, B.R. Seasonal dynamics of direct and indirect condition indices in relation to energy allocation in largemouth bass Micropterus salmoides (Lacepede). Ecol. Freshw. Fish 2004, 13, 23–36. [Google Scholar] [CrossRef]
- Ganias, K. Determining the indeterminate: Evolving concepts and methods on the assessment of the fecundity pattern of fishes. Fish. Res. 2013, 138, 23–30. [Google Scholar] [CrossRef]
- Mollet, F.M.; Engelhard, G.H.; Vainikka, A.; Laugen, A.T.; Rijnsdorp, A.D.; Ernande, B. Spatial variation in growth, maturation schedules and reproductive investment of female sole Solea solea in the Northeast Atlantic. J. Sea Res. 2013, 84, 109–121. [Google Scholar] [CrossRef] [Green Version]
- Doadrio, I.; Madeira, M.J. A new species of the genus Gobio Cuvier, 1816 (Actynopterigii, Cyprinidae) from the Iberian Peninsula and southwestern France. Graellsia 2004, 60, 107–116. [Google Scholar] [CrossRef]
- Amat-Trigo, F. Gobio—Gobio Lozanoi. In Enciclopedia Virtual de los Vertebrados Españoles; Sanz, J.J., Oliva Paterna, F.J., Eds.; Museo Nacional de Ciencias Naturales: Madrid, Spain, 2017. [Google Scholar]
- Doadrio, I.; Perea, S.; Garzón-Heydt, P.; González, J.L. Ictiofauna Continental Española. Bases para su Seguimiento; DG Medio Natural y Política Forestal; MARM: Madrid, Spain, 2011. [Google Scholar]
- Muñoz-Mas, R.; Fukuda, S.; Vezza, P.; Martínez-Capel, F. Comparing four methods for decision-tree induction: A case study on the invasive Iberian gudgeon (Gobio lozanoi; Doadrio and Madeira, 2004). Ecol. Inform. 2016, 34, 22–34. [Google Scholar] [CrossRef]
- Leunda, P. Impacts of non-native fishes on Iberian freshwater ichthyofauna: Current knowledge and gaps. Aquat. Invasions 2010, 5, 239–262. [Google Scholar] [CrossRef]
- Oliva-Paterna, F.J.; Verdiell-Cubedo, D.; Ruiz-Navarro, A.; Torralva, M. La ictiofauna continental de la Cuenca del río Segura (S.E. Península Ibérica): Décadas después de Mas (1986). An. Biol. 2014, 37–45. [Google Scholar] [CrossRef]
- Valladolid, M.; Przybylski, M. Feeding relations among cyprinids in the Lozoya River (Madrid, central Spain). Pol. Arch. Hydrobiol. 1996, 43, 213–223. [Google Scholar]
- Oscoz, J.; Leunda, P.M.; Miranda, R.; Escala, M.C. Summer feeding relationships of the cooccurring Phoxinus phoxinus and Gobio lozanoi (Cyprinidae) in an Iberian river. Folia Zool. 2006, 55, 418–432. [Google Scholar]
- Saraiva, A.; Hermida, M.; Costa, M.J.; Maia, C.; Reis, A.R.; Cruz, C.; Valente, A. First record ofPhilometra ovata (Nematoda) infection in Gobio lozanoiin Portugal. J. Fish Biol. 2008, 73, 2288–2292. [Google Scholar] [CrossRef]
- Hermoso, V.; Clavero, M. Threatening processes and conservation management of endemic freshwater fish in the Mediterranean basin: A review. Mar. Freshw. Res. 2011, 62, 244–254. [Google Scholar] [CrossRef]
- Muñoz-Mas, R.; García-Berthou, E. Alien animal introductions in Iberian inland waters: An update and analysis. Sci. Total. Environ. 2020, 703, 134505. [Google Scholar] [CrossRef]
- Winemiller, K.O.; Rose, K.A. Patterns of life-history diversification in North American fishes: Implications for population regulation. Can. J. Fish. Aquat. Sci. 1992, 49, 2196–2218. [Google Scholar] [CrossRef]
- Vila-Gispert, A.; Moreno-Amich, R. Life-history patterns of 25 species from European freshwater fish communities. Environ. Biol. Fishes 2002, 65, 387–400. [Google Scholar] [CrossRef]
- Lobón-Cerviá, J.; Torres, S. On the growth and reproduction of two populations of gudgeon (Gobio gobio L.) in Central Spain. Acta Hydrobiol. 1984, 1, 101–115. [Google Scholar]
- Lobon-Cervia, J.; Montańges, C.; Sostoa, A. Influence of environment upon the life history of gudgeon, Gobio gobio (L.): A recent and successful colonizer of the Iberian Peninsula. J. Fish Biol. 1991, 39, 285–300. [Google Scholar] [CrossRef]
- Miñano, P.A.; García-Mellado, A.; Oliva-Paterna, F.J.; Torralva, M. Edad, crecimiento y reproducción de Gobio gobio L. (Pisces, Cyprinidae) en un tramo regulado del río Segura (SE España). Anim. Biodivers. Conserv. 2003, 26, 67–77. [Google Scholar]
- Britton, J.R.; Gozlan, R.E.; Copp, G.H. Managing non-native fish in the environment. Fish Fish. 2011, 12, 256–274. [Google Scholar] [CrossRef]
- Ribeiro, F.; Leunda, P.M. Non-native fish impacts on Mediterranean freshwater ecosystems: Current knowledge and research needs. Fish. Manag. Ecol. 2012, 19, 142–156. [Google Scholar] [CrossRef]
- Almeida, D.; Stefanoudis, P.V.; Fletcher, D.H.; Rangel, C.; da Silva, E. Population traits of invasive bleak Alburnus alburnus between different habitats in Iberian fresh waters. Limnologica 2014, 46, 70–76. [Google Scholar] [CrossRef]
- García-Berthou, E. The characteristics of invasive fishes: What has been learned so far? J. Fish Biol. 2007, 71, 33–55. [Google Scholar] [CrossRef]
- Belmar, O.; Velasco, J.; Martinez-Capel, F. Hydrological classification of natural flow regimes to support environmental flow assessments in intensively regulated Mediterranean rivers, Segura River basin (Spain). Environ. Manag. 2011, 47, 992–1004. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Pérez, A.; Oliva-Paterna, F.J.; Colin, N.; Torralva, M.; Górski, K. Functional response of fish assemblage to multiple stressors in a highly regulated Mediterranean river system. Sci. Total. Environ. 2020, 730, 1–10. [Google Scholar] [CrossRef]
- Pardo, I.; Álvarez, M.; Casas, J.; Moreno, J.L.; Vivas, S.; Bonada, N.; Alba-Tercedor, J.; Jáimez-Cuéllar, P.; Moyà, G.; Prat, N.; et al. El hábitat de los ríos mediterráneos. Diseño de un índice de diversidad de hábitat. Limnetica 2002, 21, 115–133. [Google Scholar]
- González del Tánago, M.; García De Jalón, D.; Lara, F.; Garilleti, R. Riparian quality index (RQI) for assessing riparian conditions in the context of the Water Framework Directive. Ing. Civ. 2006, 143, 97–108. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G.S. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Bagenal, T.B.; Braum, E. Eggs and early life history. In Methods for Assessment of Fish Production in Fresh Waters; Bagenal, T.B., Ed.; Blackwell Scientific Publications: Oxford, UK, 1978; pp. 165–201. [Google Scholar]
- Murua, H.; Kraus, G.; Saborido-Rey, F.; Witthames, P.R.; Thorsen, A.; Junquera, S. Procedures to estimate fecundity of marine fish species in relation to their reproductive strategy. J. Northwest Atl. Fish. Sci. 2003, 33, 33–54. [Google Scholar] [CrossRef]
- Humphries, P.; King, A.J.; Koehn, J.D. Fish, flows and flood plains: Links between freshwater fishes and their environment in the Murray-Darling River System, Australia. Environ. Biol. Fishes 1999, 56, 129–151. [Google Scholar] [CrossRef]
- Munz, J.T.; Higgins, C.L. The influence of discharge, photoperiod, and temperature on the reproductive ecology of cyprinids in the Paluxy River, Texas. Aquat. Ecol. 2013, 47, 67–74. [Google Scholar] [CrossRef]
- Olden, J.D.; Naiman, R.J. Incorporating thermal regimes into environmental flows assessments: Modifying dam operations to restore freshwater ecosystem integrity. Freshw. Biol. 2010, 55, 86–107. [Google Scholar] [CrossRef]
- Rinchard, J.; Kestemont, P. Liver changes related to oocyte growth in roach, a single spawner fish, and in bleak and white bream, two multiple spawner fish. Int. Rev. Hydrobiol. 2003, 88, 68–76. [Google Scholar] [CrossRef]
- Dahle, R.; Taranger, G.L.; Karlsen, Ø.; Kjesbu, O.S.; Norberg, B. Gonadal development and associated changes in liver size and sexual steroids during the reproductive cycle of captive male and female Atlantic cod (Gadus morhua L.). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2003, 136, 641–653. [Google Scholar] [CrossRef]
- Moyle, P.B.; Cech, J.J. Fishes: An Introduction to Ichthyology; Pearson Prentice Hall: Upper Saddle River, NJ, USA, 2004. [Google Scholar]
- Vila-Gispert, A.; Zamora, L.; Moreno-Amich, R. Use of the condition of Mediterranean barbel (Barbus meridionalis) to assess habitat quality in stream ecosystems. Arch. Hydrobiol. 2000, 148, 135–145. [Google Scholar] [CrossRef]
- Harvey, B.C.; Nakamoto, R.J.; White, J.L. Reduced Streamflow Lowers Dry-Season Growth of Rainbow Trout in a Small Stream. Trans. Am. Fish. Soc. 2006, 135, 998–1005. [Google Scholar] [CrossRef]
- Wootton, R.J. Ecology of Teleost Fishes; Fish and Fisheries Series 1; Chapman and Hall: London, UK, 1990. [Google Scholar]
- Saborido-Rey, F. Ecología de la Reproducción y Potencial Reproductivo en las Poblaciones de Peces Marinos; Instituto de Investigaciones Marinas (CSIC): Pontevedra, Spain; Universidad de Vigo: Pontevedra, Spain, 2004. [Google Scholar]
- Aparicio, E.; De Sostoa, A. Reproduction and growth of Barbus haasi in a small stream in the N. E. of the Iberian Peninsula. Arch. Hydrobiol. 1998, 142, 95–110. [Google Scholar] [CrossRef]
- Vila-Gispert, A.; Alcaraz, C.; García-Berthou, E. Life-history traits of invasive fish in small Mediterranean streams. Biol. Invasions 2005, 7, 107–116. [Google Scholar] [CrossRef]
- Gasith, A.; Resh, V.H. Streams in Mediterranean Climate Regions: Abiotic Influences and Biotic Responses to Predictable Seasonal Events. Annu. Rev. Ecol. Syst. 1999, 30, 51–81. [Google Scholar] [CrossRef] [Green Version]
- Arukwe, A.; Goksøyr, A. Eggshell and egg yolk proteins in fish: Hepatic proteins for the next generation: Oogenetic, population, and evolutionary implications of endocrine disruption. Comp. Hepatol. 2003, 2, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Grande, M.; Murua, H.; Zudaire, I.; Arsenault-Pernet, E.J.; Pernet, F.; Bodin, N. Energy allocation strategy of skipjack tuna Katsuwonus pelamis during their reproductive cycle. J. Fish Biol. 2016, 89, 2434–2448. [Google Scholar] [CrossRef]
- Nunes, C.; Silva, A.; Soares, E.; Ganias, K. The Use of Hepatic and Somatic Indices and Histological Information to Characterize the Reproductive Dynamics of Atlantic SardineSardina pilchardusfrom the Portuguese Coast. Mar. Coast. Fish. 2011, 3, 127–144. [Google Scholar] [CrossRef]
- Beuchel, J.S.; Marschall, E.A.; Aday, D.D. Energy allocation patterns in a multiple spawning sunfish: Evidence for an income-based reproductive strategy. Fish. Manag. Ecol. 2013, 20, 508–517. [Google Scholar] [CrossRef]
- Johnson, R.B. Lipid Deposition in Oocytes of Teleost Fish During Secondary Oocyte Growth. Rev. Fish. Sci. 2009, 17, 78–89. [Google Scholar] [CrossRef]
- Saborido-Rey, F.; Murua, H.; Tomkiewicz, J.; Lowerre-Barbieri, S. Female Reproductive Strategies: An Energetic Balance Between Maturation, Growth and Egg Production. In 4th Workshop on Gonadal Histology of Fishes, El Puerto de Santa Maria, Spain, 16–19 June 2010; Wyanski, D.M., Brown-Peterson, N.J., Eds.; Digital CSIC: El Puerto de Santa Maria, Spain, 2010; pp. 15–18. Available online: http://hdl.handle.net/10261/24937 (accessed on 17 January 2018).
- Stephens, P.A.; Boyd, I.L.; McNamara, J.M.; Houston, A.I. Capital breeding and income breeding: Their meaning, measurement, and worth. Ecology 2009, 90, 2057–2067. [Google Scholar] [CrossRef] [PubMed]
- Wiegand, M.D. Composition, accumulation and utilization of yolk lipids in teleost fish. Rev. Fish Biol. Fish. 1996, 6, 259–286. [Google Scholar] [CrossRef]
- Zudaire, I.; Murua, H.; Grande, M.; Pernet, F.F.; Bodin, N.N. Accumulation and mobilization of lipids in relation to reproduction of yellowfin tuna (Thunnus albacares) in the Western Indian Ocean. Fish. Res. 2014, 160, 50–59. [Google Scholar] [CrossRef] [Green Version]
- Henderson, B.A.; Morgan, G.E. Maturation of walleye by age, size and surplus energy. J. Fish Biol. 2002, 61, 999–1011. [Google Scholar] [CrossRef]
- Henderson, B.A.; Trivedi, T.; Collins, N.C. Annual cycle of energy allocation to growth and reproduction of yellow perch. J. Fish Biol. 2000, 57, 122–133. [Google Scholar] [CrossRef]
- Henderson, B.A.; Wong, J.L. Control of lake trout reproduction: Role of lipids. J. Fish Biol. 1998, 52, 1078–1082. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Site | Code | Latitude | Longitude | Altitude (m.a.s.l.) | Status | Conductivity (µs cm−1) | IHF | RQI | Water Tempeture (°C, ± 95% Cl) |
|---|---|---|---|---|---|---|---|---|---|
| Tus | TUS | 38°24′40.5″ N | 2°19′01.3″ W | 809 | 1 | 399.44 ± 9.89 | 84 | 65 | 15.63 ± 2.77 |
| Camping | SE1 | 38°17′48.0″ N | 2°24′42.3″ W | 685 | 1 | 374.33 ± 8.06 | 61 | 98 | 14.71 ± 2.15 |
| Letur | SE2 | 38°24′31.9″ N | 2°06′33.5″ W | 460 | 1 | 362.62 ± 4.21 | 68 | 80 | 13.04 ± 1.59 |
| Bajo Cenajo | SE3 | 38°21′50.9″ N | 1°46′17.2″ W | 363 | 2 | 383.28 ± 3.73 | 67 | 77 | 12.49 ± 1.28 |
| Hoya García | SE4 | 38°14′30.6″ N | 1°32′35.7″ W | 200 | 2 | 686.19 ± 55.20 | 66 | 70 | 16.25 ± 2.01 |
| Sampling Site | Reproductive Stages | ANOVA | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Total | Quiescence | Maturation | Spawning | Postspawning | df | F | p | ||
| FEMALES | |||||||||
| SC | TUS | 0.651 | 0.569 | 0.679 | 0.683 | 0.629 | 3 | 4.851 | 0.004 |
| SE1 | 0.696 | 0.635 | 0.736 | 0.686 | 0.689 | 3 | 5.124 | 0.002 | |
| SE2 | 0.634 | 0.664 | 0.625 | 0.605 | 0.665 | 3 | 0.908 | 0.443 | |
| SE3 | 0.488 | 0.503 | 0.480 | 0.466 | 0.511 | 3 | 0.348 | 0.791 | |
| SE4 | 0.583 | 0.543 | 0.598 | 0.567 | 0.638 | 3 | 4.656 | 0.004 | |
| HS | TUS | 0.043 | 0.024 | 0.059 | 0.051 | 0.021 | 3 | 20.996 | <0.001 |
| SE1 | 0.051 | 0.036 | 0.074 | 0.040 | 0.025 | 3 | 43.062 | <0.001 | |
| SE2 | 0.044 | 0.037 | 0.041 | 0.057 | 0.043 | 3 | 1.603 | 0.199 | |
| SE3 | 0.025 | 0.020 | 0.027 | 0.029 | 0.023 | 3 | 1.888 | 0.135 | |
| SE4 | 0.034 | 0.028 | 0.044 | 0.032 | 0.026 | 3 | 6.309 | 0.001 | |
| GSI | TUS | 0.179 | 0.057 | 0.293 | 0.205 | 0.058 | 3 | 61.079 † | <0.001 |
| SE1 | 0.225 | 0.099 | 0.352 | 0.212 | 0.074 | 3 | 107.369 † | <0.001 | |
| SE2 | 0.196 | 0.142 | 0.188 | 0.323 | 0.098 | 3 | 33.643 | <0.001 | |
| SE3 | 0.161 | 0.084 | 0.206 | 0.229 | 0.058 | 3 | 49.019 | <0.001 | |
| SE4 | 0.140 | 0.063 | 0.234 | 0.153 | 0.054 | 3 | 146.528 † | <0.001 | |
| MALES | |||||||||
| SC | TUS | 0.697 | 0.641 | 0.747 | 0.725 | 0.586 | 3 | 3.409 | 0.022 |
| SE1 | 0.745 | 0.671 | 0.821 | 0.756 | 0.620 | 3 | 6.767 | <0.001 | |
| SE2 | 0.741 | 0.779 | 0.686 | 0.775 | 0.776 | 3 | 1.920 | 0.133 | |
| SE3 | 0.610 | 0.578 | 0.621 | 0.585 | 0.690 | 3 | 1.407 | 0.249 | |
| SE4 | 0.638 | 0.631 | 0.628 | 0.636 | 0.700 | 3 | 2.171 | 0.101 | |
| HS | TUS | 0.031 | 0.034 | 0.036 | 0.030 | 0.021 | 3 | 1.561 | 0.206 |
| SE1 | 0.030 | 0.044 | 0.036 | 0.025 | 0.018 | 3 | 11.160 | <0.001 | |
| SE2 | 0.048 | 0.053 | 0.048 | 0.044 | 0.044 | 3 | 0.437 | 0.727 | |
| SE3 | 0.026 | 0.023 | 0.028 | 0.021 | 0.037 | 3 | 4.472 | 0.007 | |
| SE4 | 0.025 | 0.033 | 0.025 | 0.020 | 0.032 | 3 | 6.724 | 0.001 | |
| GSI | TUS | 0.040 | 0.017 | 0.063 | 0.041 | 0.012 | 3 | 37.531 | <0.001 |
| SE1 | 0.052 | 0.020 | 0.085 | 0.050 | 0.014 | 3 | 52.044 | <0.001 | |
| SE2 | 0.054 | 0.038 | 0.057 | 0.087 | 0.026 | 3 | 11.223 | <0.001 | |
| SE3 | 0.040 | 0.019 | 0.055 | 0.042 | 0.031 | 3 | 52.816 | <0.001 | |
| SE4 | 0.037 | 0.016 | 0.050 | 0.032 | 0.017 | 3 | 46.862 † | <0.001 | |
| Site | Potential Fecundity | Absolute Fecundity | Batch Fecundity | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | Min | Max | Mean | Min | Max | Mean | Min | Max | ||||
| TUS | 239.3 | 89 | 401 | 152.4 | 71 | 250 | 65.8 | 37 | 101 | |||
| LogFecPOT = −3.64+ 6.30 LogLF | LogFecVIT = −1.61 + 3.987 LogLF | LogFecBAT = −1.942 + 3.988 LogLF | ||||||||||
| r2 = 0.82 | p = 0.001 | r2 = 0.54 | p = 0.015 | r2 = 0.72 | p = 0.004 | |||||||
| SE1 | 311.25 | 111 | 461 | 194.58 | 78 | 324 | 79 | 33 | 152 | |||
| LogFecPOT = −4.337 + 7.045 LogLF | LogFecVIT = −3.783 + 6.266 LogLF | LogFecBAT = −4.511 + 6.628 LogLF | ||||||||||
| r2 = 0.62 | p = 0.007 | r2 = 0.56 | p = 0.013 | r2 = 0.70 | p = 0.003 | |||||||
| SE2 | 248.13 | 136 | 389 | 168.5 | 83 | 317 | 85 | 42 | 140 | |||
| LogFecPOT = −2.023 + 4.634 LogLF | LogFecVIT = −3.482 + 5.974 LogLF | LogFecBAT = −3.444 + 5.575 LogLF | ||||||||||
| r2 = 0.75 | p = 0.012 | r2 = 0.77 | p = 0.010 | r2 = 0.63 | p = 0.018 | |||||||
| SE3 | 117.4 | 41 | 181 | 79 | 36 | 103 | 56 | 35 | 74 | |||
| LogFecPOT = −3.085 + 5.509 LogLF | LogFecVIT = −0.861 + 1.154 LogLF | LogFecBAT = −4.143 + 6.165 LogLF | ||||||||||
| r2 = 0.94 | p = 0.033 | r2 = 0.34 | p = 0.417 | r2 = 0.86 | p = 0.073 | |||||||
| SE4 | 220.25 | 70 | 427 | 132.5 | 41 | 252 | 85.5 | 37 | 124 | |||
| LogFecPOT = −3.671 + 6.115 LogLF | LogFecVIT = −3.942 + 6.172 LogLF | LogFecBAT = −2.105 + 4.119 LogLF | ||||||||||
| r2 = 0.91 | p = 0.044 | r2 = 0.95 | p = 0.023 | r2 = 0.77 | p = 0.121 | |||||||
| Muscle % | Liver % | Gonad % | Kruskal–Wallis | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| n | Range | Mean | n | Range | Mean | n | Range | Mean | χ2 | ||
| Female | Protein | 110 | 4.18–11.10 | 6.95 | 108 | 4.10–15.14 | 9.69 | 107 | 0.28–25.59 | 9.99 | 86.016 |
| Lipid | 108 | 0.40–1.35 | 0.73 | 105 | 0.52–18.79 | 6.94 | 109 | 1.05–9.09 | 3.00 | 265.237 | |
| Male | Protein | 23 | 3.49–8.18 | 5.67 | 23 | 9.26–18.13 | 12.51 | 23 | 0.16–8.99 | 2.68 | 54.916 |
| Lipid | 23 | 0.39–0.97 | 0.58 | 23 | 2.45–22.42 | 7.18 | 21 | 1.84–8.73 | 4.65 | 46.484 | |
| SC | HC | IGS | FecPOT | FecABS | FecBAT | ØPOT | ØABS | |
|---|---|---|---|---|---|---|---|---|
| SC | - | 0.713 ** | 0.426 ** | 0.491 ** | 0.386 ** | 0.749 ** | 0.032 | 0.155 |
| HC | <0.001 | - | 0.816 ** | 0.415 ** | 0.635 ** | 0.755 ** | 0.465 ** | 0.472 ** |
| IGS | <0.001 | <0.001 | - | 0.267** | 0.611 ** | 0.748 ** | 0.570 ** | 0.470 ** |
| FecPOT | <0.001 | <0.001 | 0.005 | - | 0.695 ** | 0.744 ** | 0.079 | 0.136 |
| FecABS | <0.001 | <0.001 | <0.001 | <0.001 | - | 0.768 ** | 0.732 ** | 0.680 ** |
| FecBAT | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | - | 0.015 | 0.058 |
| ØPOT | 0.744 | <0.001 | <0.001 | 0.413 | <0.001 | 0.924 | - | 0.925 ** |
| ØABS | 0.138 | <0.001 | <0.001 | 0.192 | <0.001 | 0.720 | <0.001 | - |
| ØBAT | 0.023 | 0.246 | 0.520 | 0.009 | 0.002 | 0.279 | 0.013 | <0.001 |
| ØMAX | 0.141 | <0.001 | <0.001 | 0.016 | <0.001 | 0.184 | <0.001 | <0.001 |
| % PMUS | 0.581 | 0.870 | 0.860 | 0.368 | 0.387 | 0.470 | 0.760 | 0.959 |
| % PLIV | 0.888 | 0.695 | 0.675 | 0.006 | 0.216 | 0.050 | 0.460 | 0.036 |
| % POVA | 0.015 | <0.001 | 0.003 | 0.072 | <0.001 | 0.739 | <0.001 | <0.001 |
| % LMUS | 0.005 | 0.025 | 0.178 | 0.669 | 0.365 | 0.852 | 0.250 | 0.001 |
| % LLIV | 0.139 | 0.754 | 0.303 | 0.099 | 0.104 | −0.910 | 0.576 | 0.380 |
| % LOVA | 0.707 | 0.847 | 0.533 | 0.450 | 0.020 | 0.813 | 0.003 | <0.001 |
| ØBAT | ØMAX | % PMUS | % PLIV | % POVA | % LMUS | % LLIV | % LOVA | |
| SC | 0.347 * | 0.142 | −0.054 | 0.014 | 0.234 * | −0.267 ** | −0.146 | −0.037 |
| HC | 0.186 | 0.556 ** | 0.016 | 0.038 | 0.435 ** | −0.215 * | −0.031 | −0.019 |
| IGS | 0.103 | 0.627 ** | −0.017 | 0.041 | 0.282 ** | −0.130 | −0.101 | −0.060 |
| FecPOT | 0.401 ** | 0.228 * | 0.087 | −0.259 ** | 0.173 | 0.041 | 0.161 | 0.073 |
| FecABS | 0.466 ** | 0.780 ** | 0.088 | −0.126 | 0.467 ** | −0.092 | 0.167 | 0.233 * |
| FecBAT | 0.173 | 0.212 | 0.116 | −0.308 * | −0.054 | −0.030 | −0.018 | −0.038 |
| ØPOT | 0.383 * | 0.874 ** | −0.029 | 0.071 | 0.587 ** | −0.111 | 0.055 | 0.565 ** |
| ØABS | 0.570 ** | 0.922 ** | −0.005 | 0.216 * | 0.682 ** | −0.326 ** | 0.093 | 0.661 ** |
| ØBAT | - | 0.896 ** | −0.190 | 0.107 | 0.384 ** | −0.234 | −0.030 | 0.284 |
| ØMAX | <0.001 | - | −0.034 | 0.054 | 0.605 ** | −0.174 | 0.132 | 0.583 ** |
| % PMUS | 0.234 | 0.724 | - | −0.070 | −0.054 | 0.096 | 0.175 | 0.134 |
| % PLIV | 0.507 | 0.574 | 0.466 | - | 0.119 | −0.255 ** | −0.279 ** | 0.030 |
| % POVA | 0.013 | <0.001 | 0.574 | 0.217 | - | −0.213 * | 0.143 | 0.368 ** |
| % LMUS | 0.141 | 0.060 | 0.317 | 0.007 | 0.026 | - | 0.240* | −0.083 |
| % LLIV | 0.855 | 0.179 | 0.073 | 0.004 | 0.146 | 0.013 | - | 0.141 |
| % LOVA | 0.025 | 0.001 | 0.162 | 0.753 | <0.001 | 0.389 | 0.148 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amat-Trigo, F.; Torralva, M.; González-Silvera, D.; Martínez-López, F.J.; Oliva-Paterna, F.J. Plasticity in Reproductive Traits, Condition and Energy Allocation of the Non-Native Pyrenean Gudgeon Gobio lozanoi in a Highly Regulated Mediterranean River Basin. Water 2021, 13, 387. https://doi.org/10.3390/w13030387
Amat-Trigo F, Torralva M, González-Silvera D, Martínez-López FJ, Oliva-Paterna FJ. Plasticity in Reproductive Traits, Condition and Energy Allocation of the Non-Native Pyrenean Gudgeon Gobio lozanoi in a Highly Regulated Mediterranean River Basin. Water. 2021; 13(3):387. https://doi.org/10.3390/w13030387
Chicago/Turabian StyleAmat-Trigo, Fátima, Mar Torralva, Daniel González-Silvera, Francisco Javier Martínez-López, and Francisco José Oliva-Paterna. 2021. "Plasticity in Reproductive Traits, Condition and Energy Allocation of the Non-Native Pyrenean Gudgeon Gobio lozanoi in a Highly Regulated Mediterranean River Basin" Water 13, no. 3: 387. https://doi.org/10.3390/w13030387