3.1. Sobol’s Sensitivity Analysis Method
The global sensitivity analysis with Sobol’s method was carried out to select the parameters that are the most sensitive to the uncertainty of 10% and 20% applied to all parameters around their nominal values; the data of the climatic variables of the spring–summer season were used to run the simulations. To estimate the sensitivity indices for the HORTSYST crop model, the simulation was run 10,000 times at the start of fructification (40 days after transplant (DAT)) and at the end of the crop cycle (119 DAT).
The most influential parameters in the model are listed in
Table 4. At 40 DAT, the sum of the first-order indices (main effects) for PTI was 0.95, and the sum of the total effect indices was 1.01; for the DMP variable, it was 1.00 for first-order indices, and the sum of total effect indices was 1.00; for Nup, it was 1.08 for first-order indices and 0.99 for the sum of total effect indices; for the LAI variable, 1.01 was obtained for first-order indices and the sum of total effect indices of 1.00; and finally, for ETc, 0.98 was for first-order indices and for the sum of total effect indices of 1.00. At 119 DAT, the sum of the first order for PTI was 0.96; for the DMP variable, the sum of this index was 0.92; for Nup, it was 0.99; for LAI, the value was 1.04; and for ETc, the sum was 0.93. The sum of the total effect indices for PTI was 1.00; for DMP, it was 1.02; for Nup, this sum was 1.01; for LAI, it was 0.99; and for ETc, the sum of these indices of 1.00 was found. In the fructification stage, the existence of interaction between parameters was not clear; however, for the second stage, such interaction was observed, since the values of the indexes
.
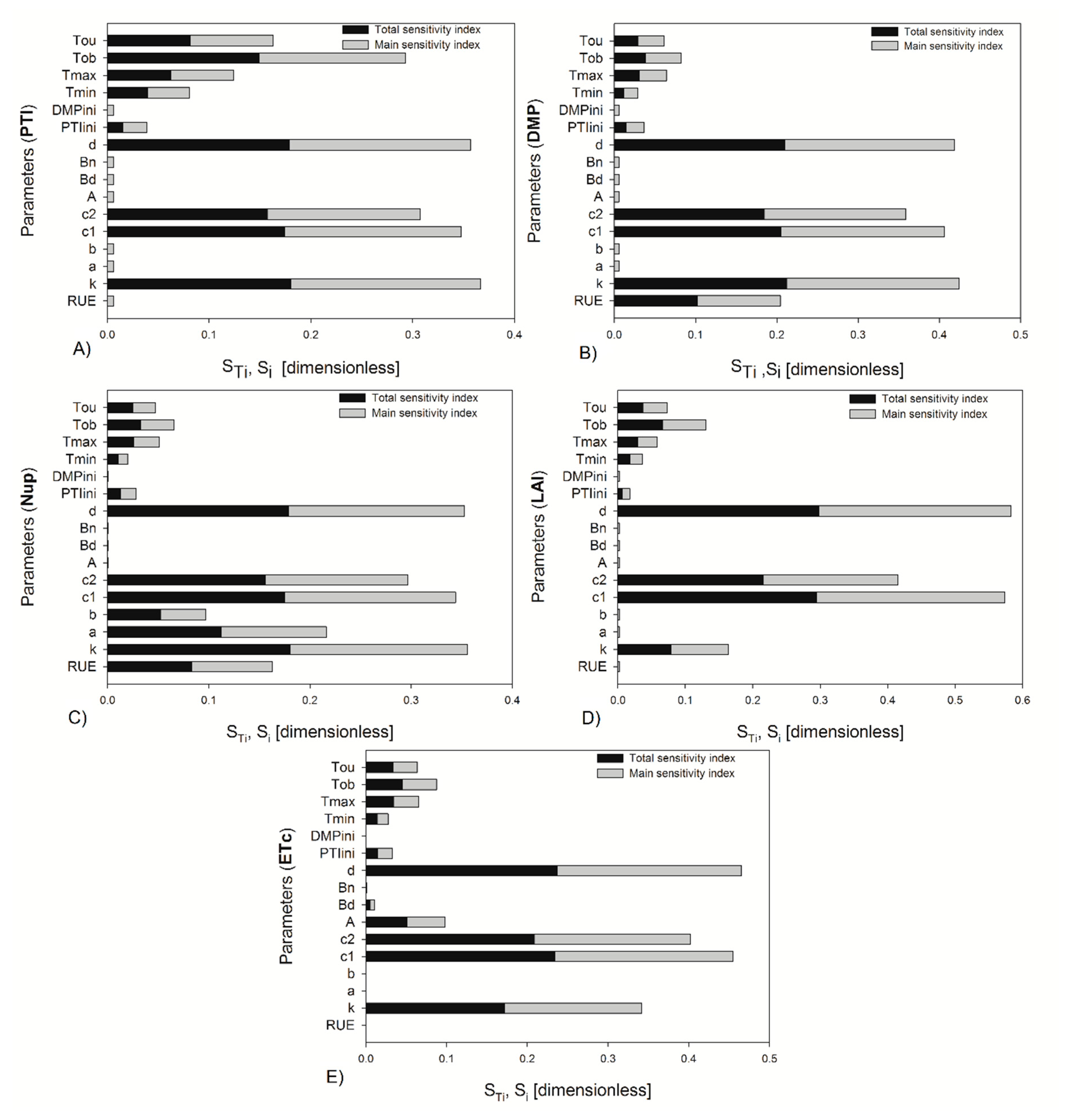
Figure 2 and
Figure 3 show the indices for 20% uncertainty on the parameters at the start of fructification and at the end of growth; in both cases the most important parameters were the same, with 10% uncertainty on the parameters. They only varied in order of importance, with these changes being the most evident for 40 DAT. The most important parameters in descending order are shown in
Table 4. The sum of the first-order effects and the total indices were for PTI (1.08, 1.04), DMP (1.08, 1.04), Nup (0.99, 1.05), LAI (1.02, 1.05), and ETc (1.00, 1.06) at fructification stage. In the analysis performed with 20% uncertainty at 119 DAT, the parameter d (crop density) became more important than c
1 for ETc output, whereas the other parameters kept their order of importance found with 10% uncertainty. In the case of 119 DAT, the sum of the first-order effects and the total indices were for PTI (0.90, 1.00), DMP (0.91, 1.01), Nup (0.95, 1.02), LAI (0.99, 1.00), and ETc (0.96, 1.02).
The sum of total sensitivity indices for the most important parameters (
) was slightly higher than 1 when the sensitivity analysis was carried out with 10% uncertainty at 40 DAT and 119 DAT, but it is not conclusive to say that the model is nonadditive. Nevertheless, with the 20% uncertainty, the sums of total indices for all output responses for both crop stages were different from 1, so the model was nonadditive; this also was checked with the sum of the first-order effects
< 1, according to Saltelli et al. [
2]. The interactions between parameters were clearer when the uncertainty on the parameters was increased to 20% at the beginning of fructification and at the end of the crop cycle
for all output variables.
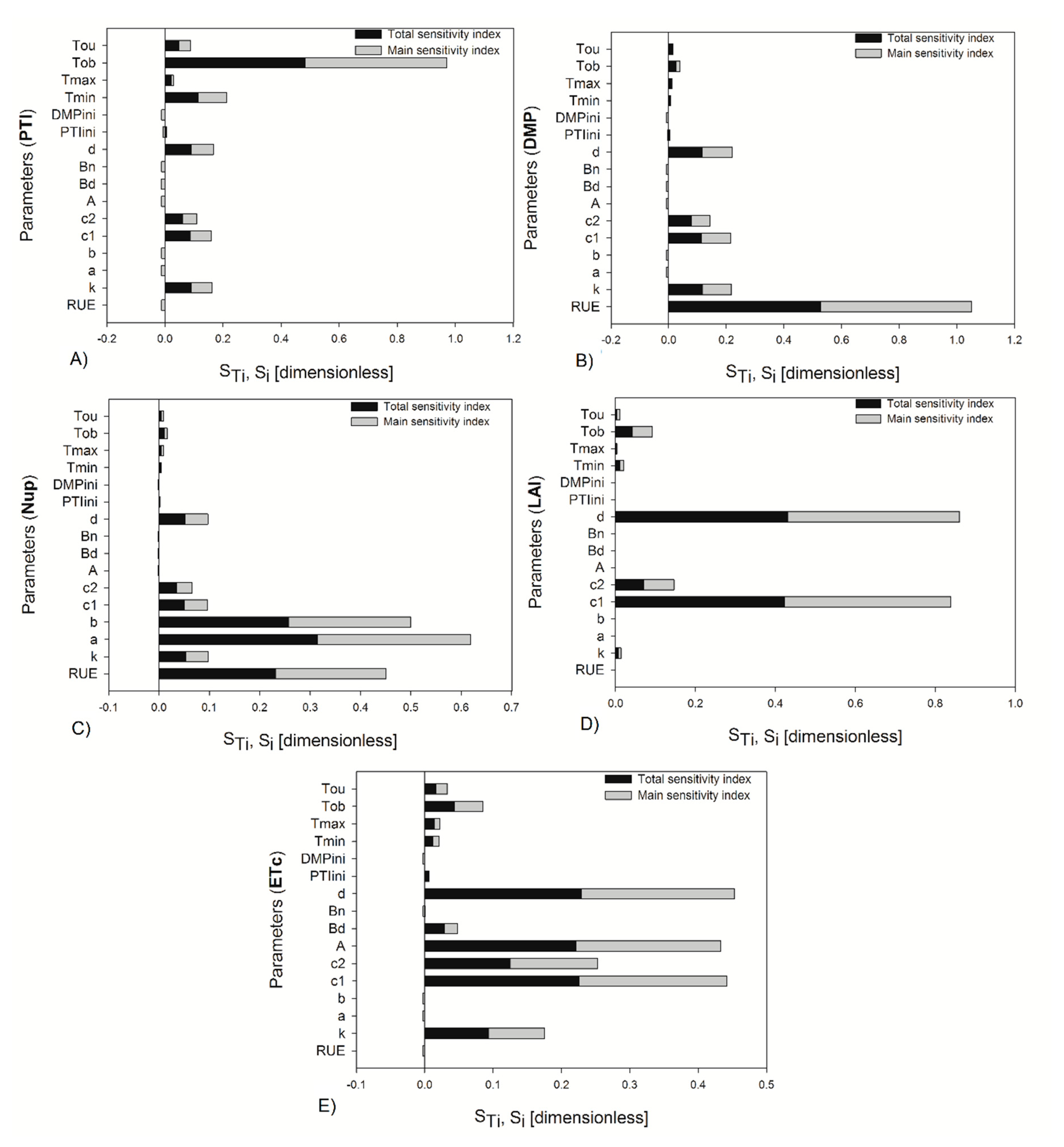
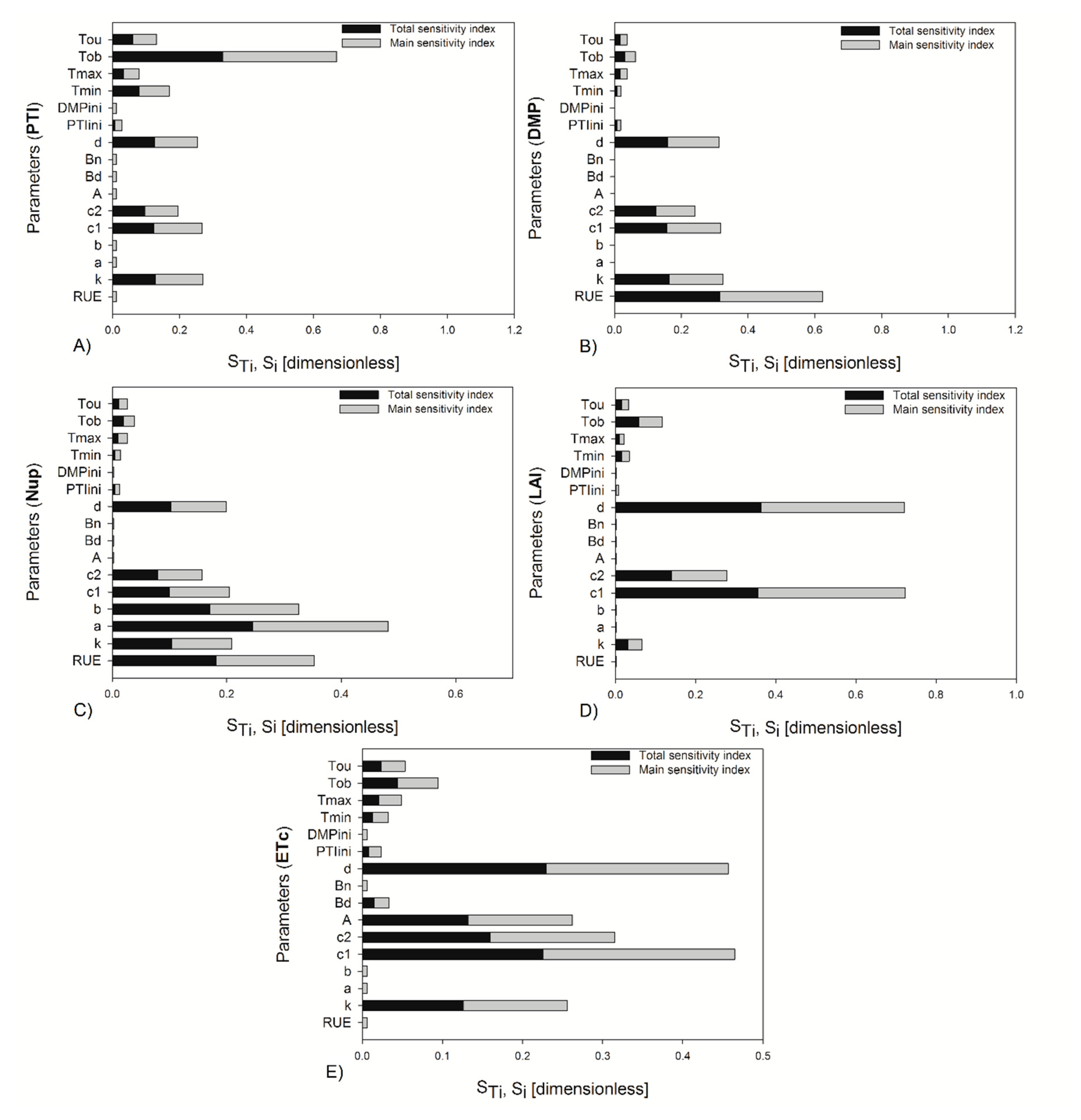
The sensitivity indices were also estimated, taking into account the daily integration of output variables at the end of the crop cycle with 20% uncertainty, and the indices found were different for some parameters than when two specific stages were considered in the analysis (
Figure 4). For the daily integration of outputs, some parameters changed in their magnitude of importance; for example, for PTI and DMP,
and
decreased their index values, and the rest of the parameters increased. The parameter
increased the magnitude of its indices for Nup, LAI, and ETc. In the sensitivity analysis run in the two growth stages, it was observed that a greater number of parameters were more important at the beginning of fructification than at the end of crop growth for 10% and 20% of the variation of the parameters (
Table 4).
These results indicate that the parameters changed over time, with some of them increasing in importance and other decreasing; for example, the parameter
increased its importance in PTI,
in DMP,
and
in Nup,
in LAI, and
and
in ETc. Two of the parameters that decreased their importance with the growth and development of the crop were
(LAI, ETc, and DMP) and the parameter
. This temporal variation was also reported by López-Cruz et al. [
52] with the NICOLET model for lettuce and the SUCROS model applied to husk tomato [
53]. In addition, Wang et al. [
54] found this variation throughout crop growth and the temporal dynamic variation as a result of the increase in uncertainty in the parameters of the WOFOST model used in corn cultivation.
The cardinal temperatures
,
, and
had an influence on the model’s performance (
Figure 2,
Figure 3 and
Figure 4), particularly at the beginning of fructification. However, these parameters were not selected for the parameter estimation technique, because they were taken from the literature [
35,
40,
41]. All of these parameters were obtained by experimentation; other parameters, such as
(extinction coefficient) and
(crop density), were also not considered, though they showed high sensitivity in the analysis, because the
parameter can be estimated with a ceptometer, and crop density (
) is defined before setting the experiment.
During the analysis, it was found that these two parameters were the most sensitive in all output variables of the model, since these output variables are strongly influenced by the interception of the light, which is dependent on the simulation of the dry matter produced, leaf area index area, and crop transpiration and indirectly dependent on the nutritional uptake, since its determination depends on the biomass produced by the crop.
The effects of these parameters were discussed by De Reffye et al. [
33], who argued that limitations occur for light interception when density is low, because the expression of light interception assumes a homogeneous distribution of leaves. Therefore, the parameters that finally were considered for calibration were:
,
,
,
,
,
,
,
, and
. The
parameter explains the quantity of carbon assimilated and converted to total dry biomass; thus, it was important for DMP and Nup, because the two variables are correlated. For models with the light-use efficiency approach, this parameter and
become more significant, as was found for the CERES–maize model [
55] and WOFOST model studied by Dzitsi et al. [
56], the SALUS model for maize, peanut and cotton reported by Wang et al. [
54], and AZODYN for wheat [
57]; all of them found higher values of
and
for
, and
.
Parameters and are important for quantifying nitrogen uptake. The increase in the indices of these two parameters and from 40 DAT to the end of crop growth is explained by the increased slope of the exponential growth curve of total dry matter production due to the fruit’s filling, and this, in turn, increased crop nitrogen demand. and explain the expansion of leaf area. The indices for decreased over time because the maximum LAI value of the crop was reached, meaning the plateau of this variable’s curve was attained; however, increased its importance over time.
On the other hand,
,
, and
influence the radiation and vapor pressure deficit (VPD) in the estimation of crop transpiration. The second and third parameters were not significant in this analysis. Similar results were found by Sánchez [
58]. However, these authors found that these parameters become important for the autumn–winter season; for this reason, they were considered as significant parameters. The parameter
(one of the two initial conditions) did not have high values for
and
, but we realized that it improved the performance of the calibration of the other selected parameters. The differences between two indices were for
(0.004),
(0.010),
(0.015),
(0.004),
(0.010),
(0.01). The only parameters that did not have interaction were
,
, and
.
3.2. Calibration of HORTSYST Model by Differential Evolution Algorithm
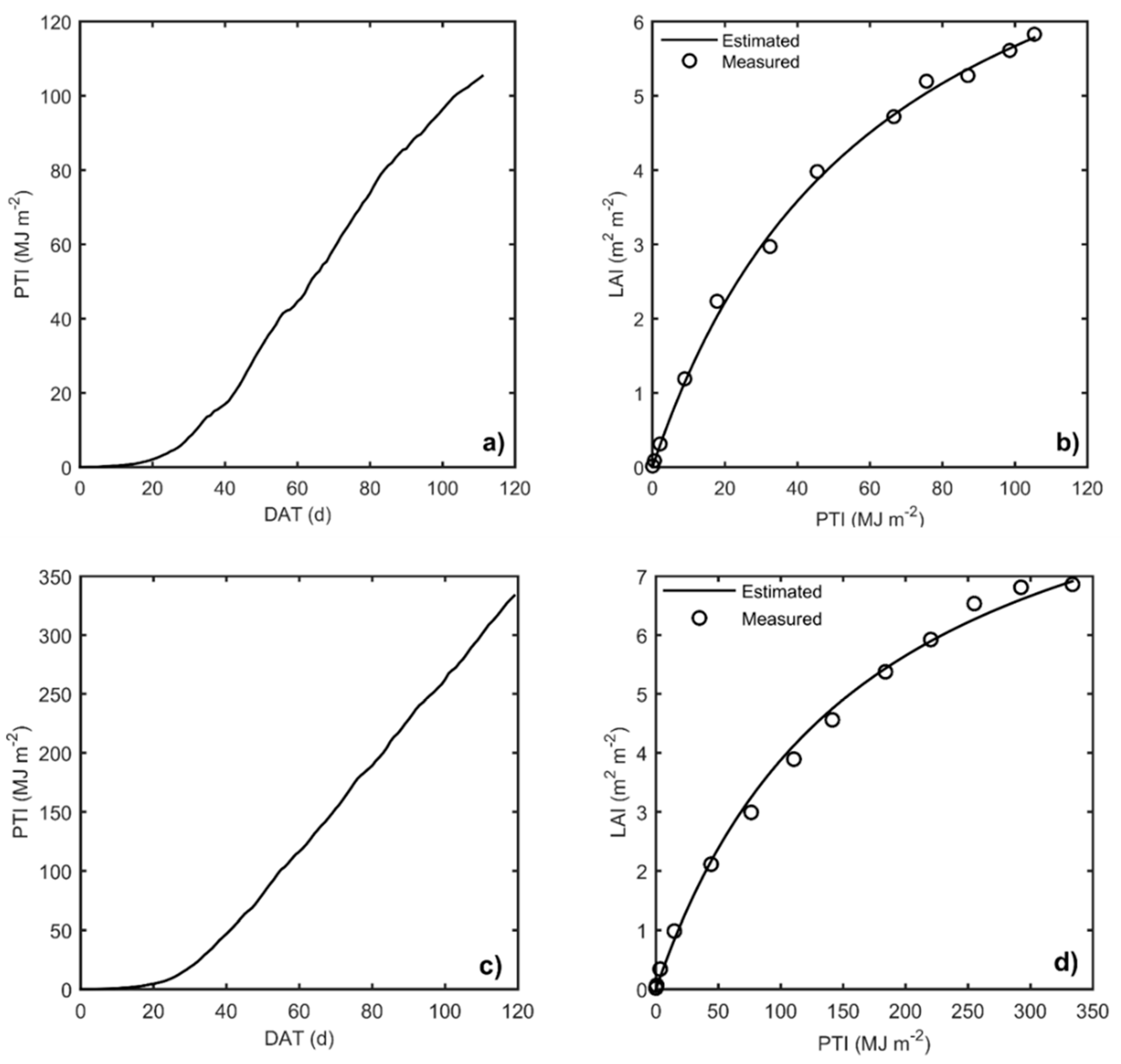
The HORTSYST crop growth model was calibrated by solving a minimization optimization problem. Nine parameters were estimated during the calibration for the autumn–winter and spring–summer seasons. PTI vs. LAI Michaelis–Menten function behavior after calibration are shown in
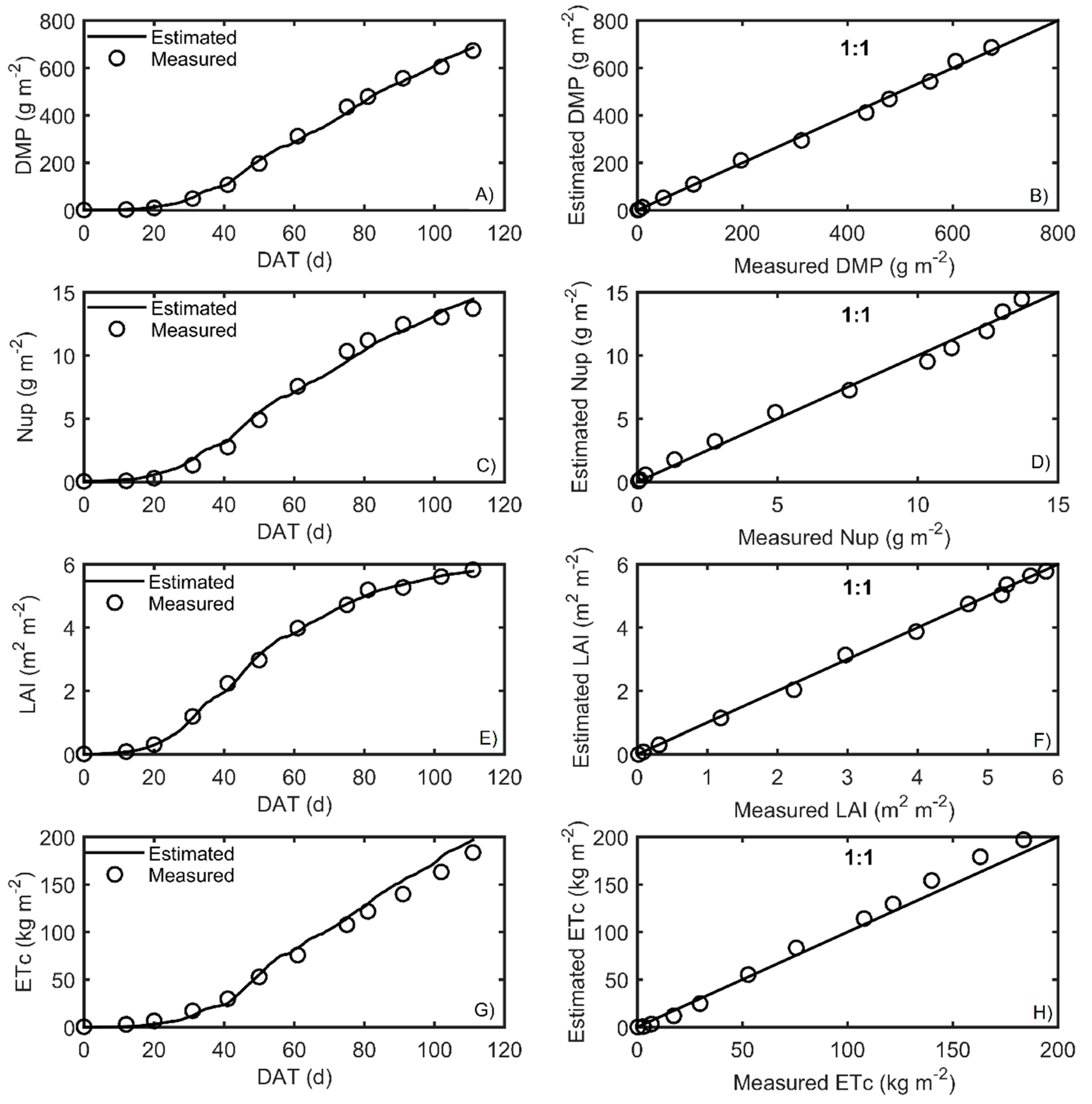
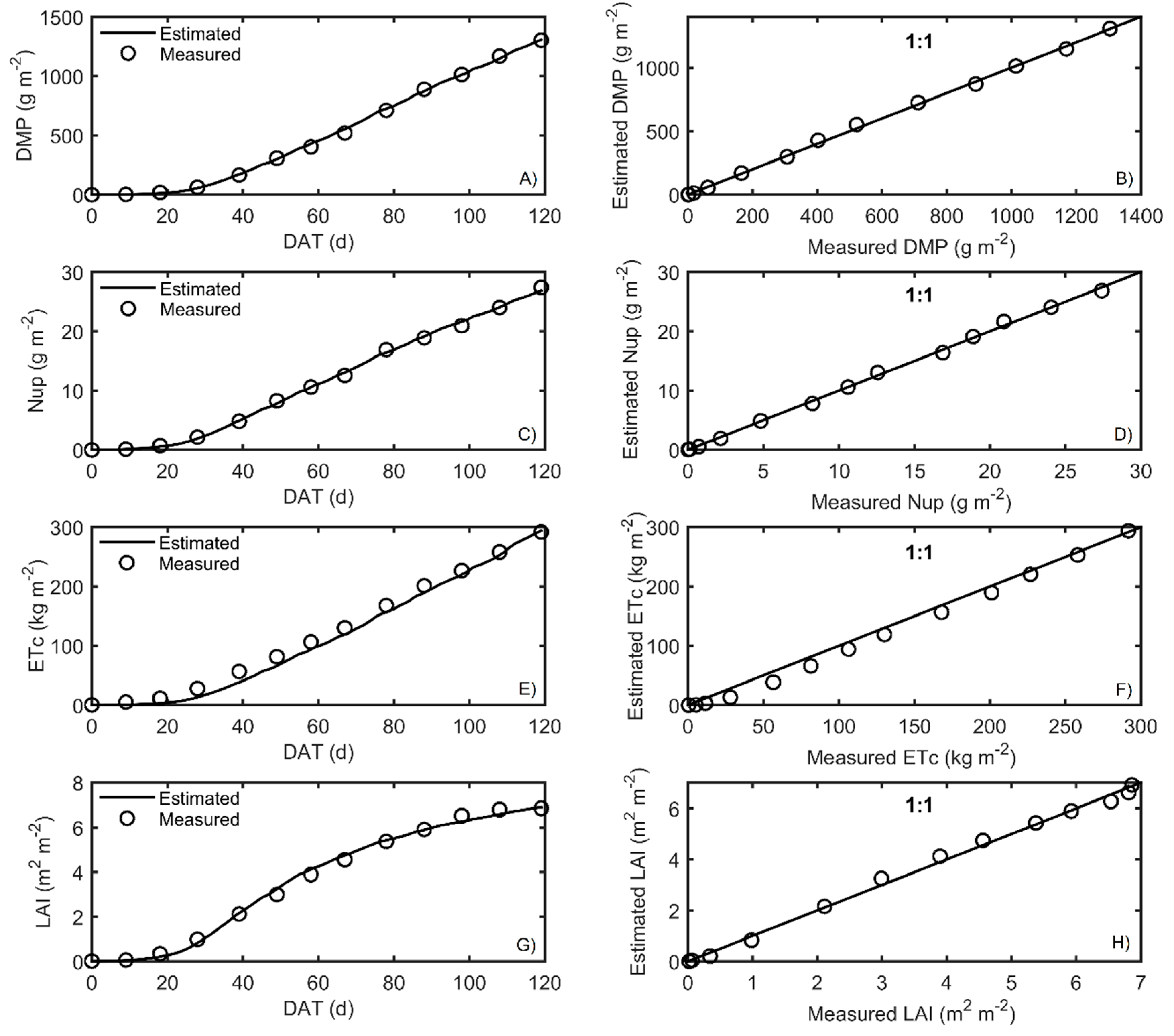
Figure 5. Measured and simulated data after calibration are shown in
Figure 6 and
Figure 7. The values of the calibrated parameters are listed in
Table 4, and the model’s goodness of fit statistics are presented in
Table 5.
The best fits, according to RMSE, were for LAI, followed by Nup, DMP, and ETc for both crop seasons. The RMSE was close to zero, indicating an excellent prediction of model performance.
Another goodness fit index used to evaluate the model was the efficiency of the model (EF), which resulted in values close to one, which means that the model presented good adjustments when correlating the variables of the model, according to Xuan et al. [
47] and Wallach et al. [
51]. As for the bias values found in the autumn–winter season, the nitrogen uptake was slightly underestimated, and DMP, LAI, and ETc were overestimated; in the case of the spring–summer season, LAI and DMP were underestimated, and Nup and ETc were overestimated. Furthermore, the 1:1 plots are presented in
Figure 6 and
Figure 7 to visualize the quality of the prediction of the output responses in the HORTSYST model.
All parameters were accurately calibrated; only parameter
in the transpiration rate turned out to have high standard deviation during the autumn–winter season (
Table 6). This means that it was very uncertain for autumn–winter, but this problem was not found for spring–summer. The calibrated
value for autumn–winter was higher than that found by Gallardo et al. [
30], and, for spring–summer, it was closer to what was reported by Challa and Bakker [
42]; the values obtained from RUE were different for each crop cycle evaluated. The parameter
was lower for the two crop periods, and
was higher than reported by Gallardo et al. [
30]; these calibrated values were quite similar for both seasons. For the LAI variable, the parameter
was closer for two seasons, but
during the spring–summer was more than twice that found in autumn–winter. The parameters
,
concerning ETc were higher than those found by Sánchez et al. [
43] for both the autumn–winter and spring–summer seasons. These parameters were different between each crop cycle.
The HORTSYST model’s parameters calibrated using the DE algorithm were closer than those found by Martinez-Ruiz et al. [
21], who used a nonlinear least square method to find the correct values of the parameters for spring–summer, except for parameter
.


 ,
, 








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}