
Acoustic Telemetry Monitors Movements of Wild Adult Catfishes in the Mekong River, Thailand and Laos

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
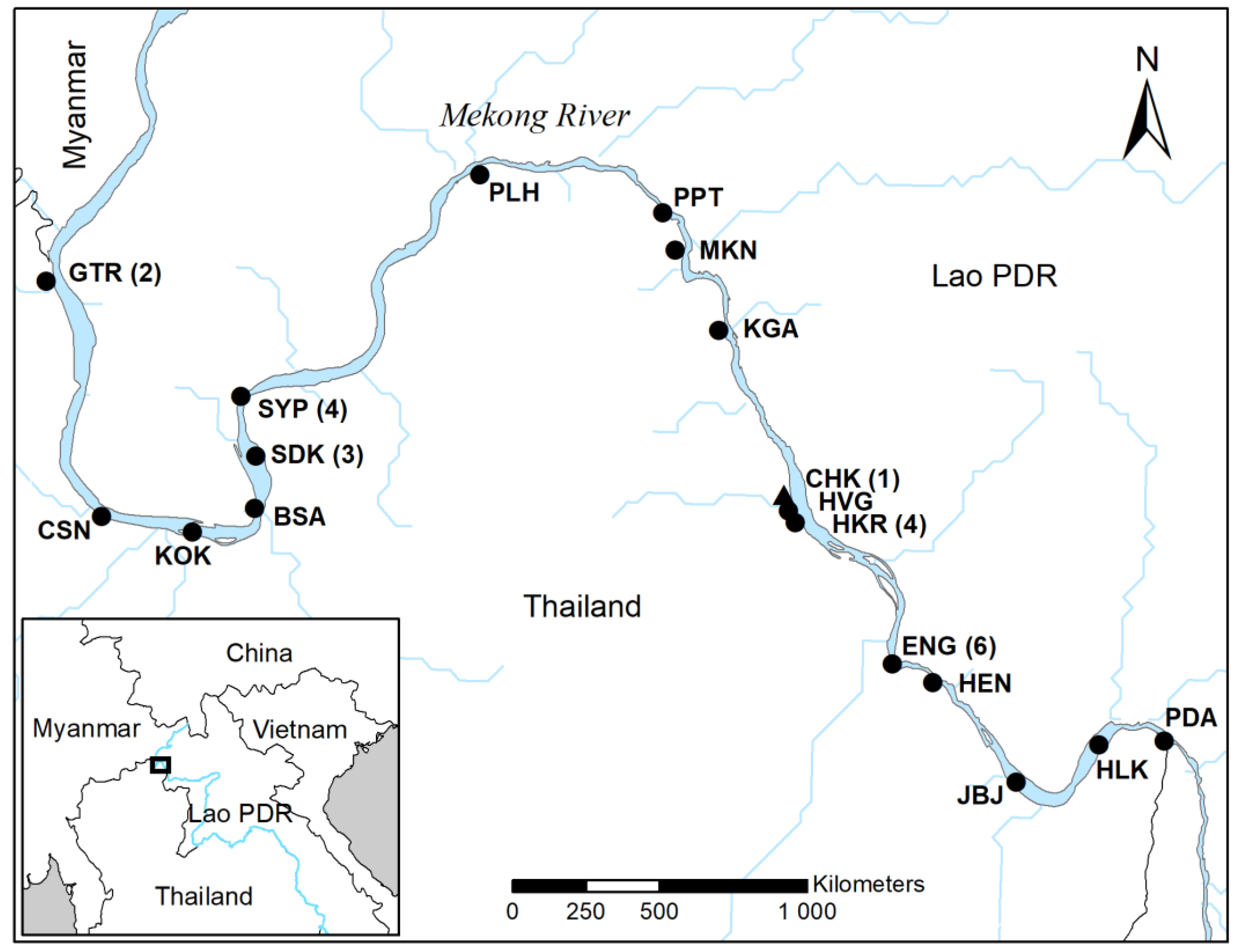
2.1. Study Area
2.2. Telemetry Monitoring
2.3. Fish Capture and Tagging
2.4. Data Processing and Analysis
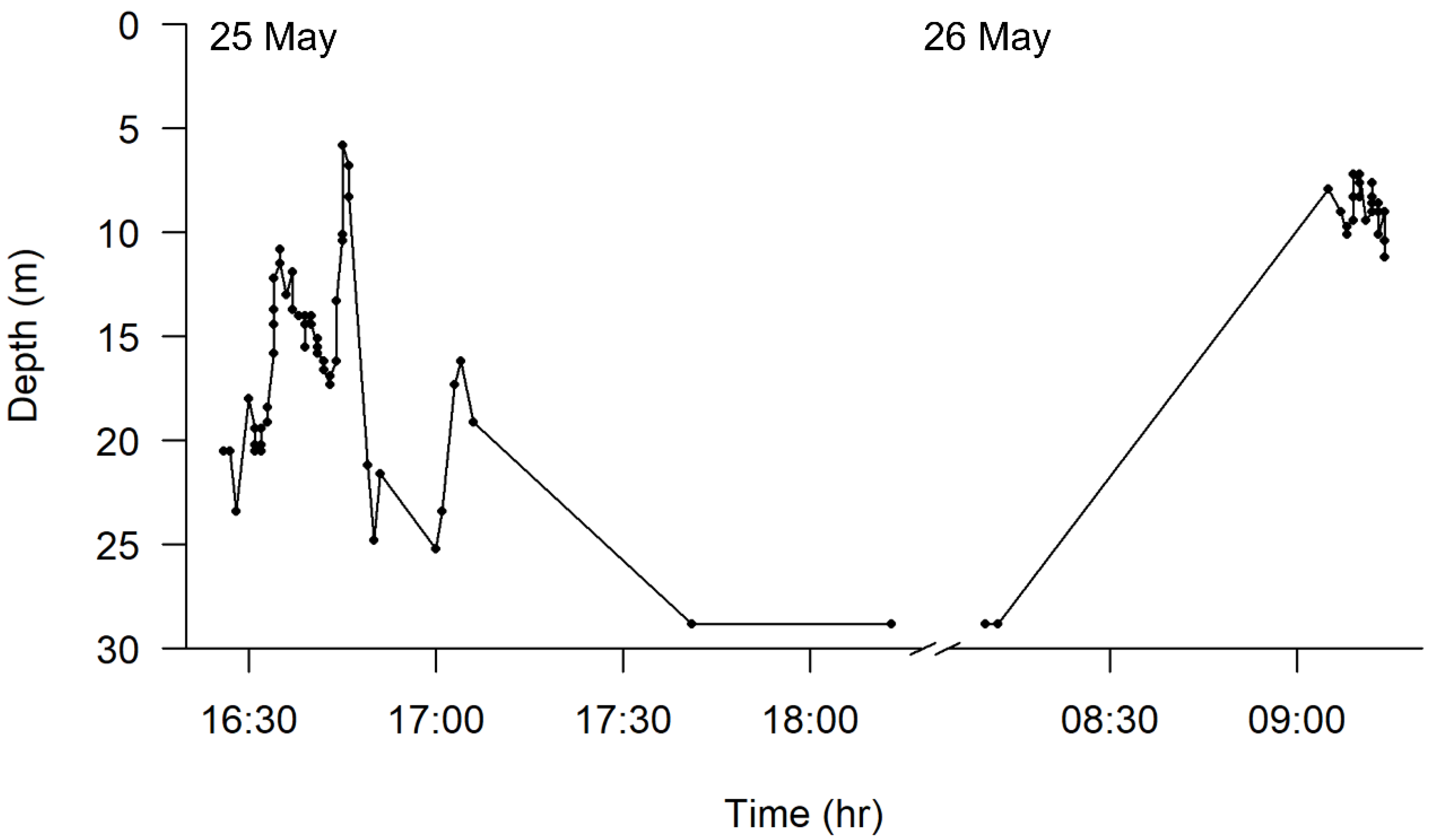
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dudgeon, D. Riverine biodiversity in Asia: A challenge for conservation biology. Hydrobiologia 2000, 418, 1–13. [Google Scholar] [CrossRef]
- Rainboth, W.J. Fishes of the Cambodian Mekong; Food & Agriculture Organization: Rome, Italy, 1996. [Google Scholar]
- Van Zalinge, N.; Nao, S.T.; Touch, S.T.; Deap, L. Where there is water, there is fish? Cambodian fisheries issues in a Mekong River basin perspective. Iclarm Stud. Rev. 2000, 26, 37–48. [Google Scholar]
- Baran, E. Mekong Fisheries and Mainstream Dams; Fisheries sections in ICEM: Hanoi, Vietnam, 2010; p. 145. [Google Scholar]
- Lucas, M.; Baras, E. Migration of Freshwater Fishes; Blackwell Science: Oxford, UK, 2001; p. 420. [Google Scholar]
- Barlow, C.; Baran, E.; Halls, A.S.; Kshatriya, M. How much of the Mekong fish catch is at risk from mainstream dam development? Catch Cult. 2008, 14, 16–21. [Google Scholar]
- Poulsen, A.F.; Poeu, O.; Viravong, S.; Suntornratana, U.; Thanh Tung, N. Fish migrations of the Lower Mekong River Basin: Implications for development, planning and environmental management. MRC Technical Paper No. 8; Mekong River Commission: Phnom Penh, Cambodia, 2002; p. 62. [Google Scholar]
- Baird, I.G.; Flaherty, M.S. Mekong river fish conservation zones in southern Laos: Assessing effectiveness using local ecological knowledge. Environ. Manag. 2005, 36, 439–454. [Google Scholar] [CrossRef]
- Baird, I.; Hogan, Z.; Phylaivanh, B.; Moyle, P. A communal fishery for the migratory catfish Pangasius macronema in the Mekong River. Asian Fish. Sci. 2001, 14, 25–41. [Google Scholar]
- Campbell, T.; Pin, K.; Ngor, P.B.; Hogan, Z. Conserving Mekong megafishes: Current status and critical threats in Cambodia. Water 2020, 12, 1820. [Google Scholar] [CrossRef]
- Gray, T.N.E.; Phommachak, A.; Vannachomchan, K.; Guegan, F. Using local ecological knowledge to monitor threatened Mekong megafauna in Lao PDR. PLoS ONE 2017, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngor, P.B.; McCann, K.S.; Grenouillet, G.; So, N.; McMeans, B.C.; Fraser, E.; Lek, S. Evidence of indiscriminate fishing effects in one of the world’s largest inland fisheries. Sci. Rep. 2018, 8, 8947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- IUCN. The IUCN Red List of Threatened Species. Version 2020-2. International Union for the Conservation of Nature. 2020. Available online: http://www.iucnredlist.org (accessed on 23 July 2020).
- Roberts, T.R. Artisanal fisheries and fish ecology below the great waterfalls of the Mekong River in southern Laos. Nat. Hist. Bull. Siam Soc. 1993, 41, 31–62. [Google Scholar]
- Roberts, T.R.; Baird, I.G. Traditional fisheries and fish ecology on the Mekong River at Khone Waterfalls in southern Laos. Nat. Hist. Bull. Siam Soc. 1995, 43, 219–262. [Google Scholar]
- Ahmed, M.; Tana, T.S.; Thouk, N. Sustaining the gifts of the Mekong: The future of freshwater capture fisheries in Cambodia. Watershed 1996, 1, 33–38. [Google Scholar]
- Baird, I.G.; Flaherty, M.S.; Phylavanh, B. Mekong River Pangasiidae catfish migrations and the Khone Falls wing trap fishery in southern Laos. Nat. Hist. Bull. Siam Soc. 2004, 52, 81–109. [Google Scholar]
- Carrizo, S.F.; Jähnig, S.C.; Bremerich, V.; Freyhof, J.; Harrison, I.; He, F.; Langhans, S.D.; Tockner, K.; Zarfl, C.; Darwall, W. Freshwater megafauna: Flagships for freshwater biodiversity under threat. Bioscience 2017, 67, 919–927. [Google Scholar] [CrossRef]
- Hogan, Z.S.; Moyle, P.B.; May, B.; Vander Zanden, M.J.; Baird, I.G. The imperiled giants of the Mekong. Am. Sci. 2004, 92, 228–237. [Google Scholar]
- Meyer, C.G. Electronic tags reveal the hidden lives of fishes. Bull. Mar. Sci. 2017, 93, 301–318. [Google Scholar] [CrossRef]
- Thorstad, E.B.; Rikardsen, A.H.; Alp, A.; Okland, F. The use of electronic tags in fish research—an overview of fish telemetry methods. Turk. J. Fish. Aquat. Sci. 2013, 13, 881–896. [Google Scholar] [CrossRef]
- Brownscombe, J.W.; Ledee, E.J.I.; Raby, G.D.; Struthers, D.P.; Gutowsky, L.F.G.; Nguyen, V.M.; Young, N.; Stokesbury, M.J.W.; Holbrook, C.M.; Brenden, T.O.; et al. Conducting and interpreting fish telemetry studies: Considerations for researchers and resource managers. Rev. Fish Biol. Fish. 2019, 29, 369–400. [Google Scholar] [CrossRef]
- Crossin, G.T.; Heupel, M.R.; Holbrook, C.M.; Hussey, N.E.; Lowerre-Barbieri, S.K.; Nguyen, V.M.; Raby, G.D.; Cooke, S.J. Acoustic telemetry and fisheries management. Ecol. Appl. 2017, 27, 1031–1049. [Google Scholar] [CrossRef]
- Dewar, H.; Mous, P.; Domeier, M.; Muljadi, A.; Pet, J.; Whitty, J. Movements and site fidelity of the giant manta ray, Manta birostris, in the Komodo Marine Park, Indonesia. Mar. Biol. 2008, 155, 121–133. [Google Scholar] [CrossRef]
- Moore, A.; Ives, S.; Mead, T.A.; Talks, L. The migratory behaviour of wild Atlantic salmon (Salmo salar L.) smelts in the River Test and Southampton Water, southern England. Hydrobiologia 1998, 372, 295–304. [Google Scholar] [CrossRef]
- Hahn, L.; Martins, E.G.; Nunes, L.D.; da Câmara, L.F.; Machado, L.S.; Garrone-Neto, D. Biotelemetry reveals migratory behaviour of large catfish in the Xingu River, Eastern Amazon. Sci. Rep. 2019, 9, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Ngamsiri, T.; Nakajima, M.; Sukmanomon, S.; Sukumasavin, N.; Kamonrat, W.; Na-Nakorn, U.; Taniguchi, N. Genetic diversity of wild Mekong giant catfish Pangasianodon gigas collected from Thailand and Cambodia. Fish. Sci. 2007, 73, 792–799. [Google Scholar] [CrossRef]
- Chan, B.; Ngor, P.B.; Hogan, Z.S.; So, N.; Brosse, S.; Lek, S. Temporal dynamics of fish assemblages as a reflection of policy shift from fishing concession to co-management in one of the world’s largest tropical flood pulse fisheries. Water 2020, 12, 2974. [Google Scholar] [CrossRef]
- Allan, J.D.; Abell, R.; Hogan, Z.E.B.; Revenga, C.; Taylor, B.W.; Welcomme, R.L.; Winemiller, K. Overfishing of inland waters. BioScience 2005, 55, 1041–1051. [Google Scholar] [CrossRef] [Green Version]
- Poulsen, A.; Poeu, O.; Viravong, S.; Suntornratana, U.; Thanh Tung, N. Deep Pools as Dry Season Fish Habitats in the Mekong Basin. MRC Technical Paper No. 4; Mekong River Commission: Phnom Penh, Cambodia, 2002; p. 22. [Google Scholar]
- Poulsen, A.F.; Valbo-Jørgensen, J. Fish Migrations and Spawning Habits in the Mekong Mainstream—A Survey Using Local Knowledge (Basin-Wide). Assessment of Mekong Fisheries: Fish Migrations and Spawning and the Impact of Water Management Component, Vientiane, Lao PDR; Internal Report; Mekong River Commission: Vientiane, Laos, 2000. [Google Scholar]
- Hogan, Z. Imperiled Giant Fish and Mainstream Dams in the Lower Mekong Basin: Assessment of Current Status, Threats, and Mitigation; University of Nevada: Reno, NV, USA, 2011. [Google Scholar]
- Clements, S.; Jepsen, D.; Karnowski, M.; Schreck, C.B. Optimization of an acoustic telemetry array for detecting transmitter-implanted fish. N. Am. J. Fish. Manag. 2005, 25, 429–436. [Google Scholar] [CrossRef]
- Hogan, Z.; Heng, K.; Zalinge, N. Underwater biotelemetry to study fish migrations in the Mekong River. Mekong Fish Catch Cult. 2002, 7, 9–13. [Google Scholar]
- Mulcahy, D.M. Surgical implantation of transmitters into fish. Ilar J. 2003, 44, 295–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baird, I.G.; Flaherty, M.S.; Phylavanh, B. Rhythms of the river: Lunar phases and migrations of small carps (Cyprinidae) in the Mekong River. Nat. Hist. Bull. Siam Soc. 2003, 51, 5–36. [Google Scholar]
- Mitamura, H.; Mitsunaga, Y.; Arai, N.; Viputhanumas, T. Movements of immature hatchery-reared Mekong giant catfish Pangasianodon gigas released in the Mekong River, measured using acoustic telemetry. Fish. Sci. 2008, 74, 1034–1039. [Google Scholar] [CrossRef]
- Heupel, M.R.; Semmens, J.M.; Hobday, A.J. Automated acoustic tracking of aquatic animals: Scales, design and deployment of listening station arrays. Mar. Freshw. Res. 2006, 57, 1–13. [Google Scholar] [CrossRef]
- Jepsen, N.; Thorstad, E.B.; Havn, T.; Lucas, M.C. The use of external electronic tags on fish: An evaluation of tag retention and tagging effects. Anim. Biotelemetry 2015, 3, 49. [Google Scholar] [CrossRef] [Green Version]
- Vemco. How Much Detection Range Can I Expect from My Tags and Receivers? Available online: https://support.vemco.com/s/article/How-much-detection-range-can-I-expect-from-my-tags-and-receivers (accessed on 4 February 2021).
- Vemco. V16 Continuous Transmitters Datasheet. 2018. Available online: https://www.oceans-research.com/wp-content/uploads/2016/09/v16-cont-1.pdf (accessed on 4 February 2021).
- Burger, C.V.; Wilmot, R.L.; Wangaard, D.B. Comparison of spawning areas and times for two runs of chinook salmon (Oncorhynchus tshawytscha) in the Kenai River, Alaska. Can. J. Fish. Aquat. Sci. 1985, 42, 693–700. [Google Scholar] [CrossRef]
- Bendock, T.; Alexandersdottir, M. Hooking mortality of chinook salmon released in the Kenai River, Alaska. N. Am. J. Fish. Manag. 1993, 13, 540–549. [Google Scholar] [CrossRef]
- Bernard, D.R.; Hasbrouck, J.J.; Fleischman, S.J. Handling-induced delay and downstream movement of adult chinook salmon in rivers. Fish. Res. 1999, 44, 37–46. [Google Scholar] [CrossRef]
- Manns, R.E.; Whiteside, B.G. Behavioral variations associated with ultrasonic tagging of Guadalupe bass in Lake Travis, Texas. Underw. Telem. Newsl. 1979, 9, 4–9. [Google Scholar]
- Diana, J.S. Diel activity pattern and swimming speeds of northern pike (Esox lucius) in Lac Ste. Anne, Alberta. Can. J. Fish. Aquat. Sci. 1980, 37, 1454–1458. [Google Scholar] [CrossRef]
- Mesing, C.L.; Wicker, A.M. Home range, spawning migrations, and homing of radio-tagged Florida largemouth bass in two central Florida lakes. Trans. Am. Fish. Soc. 1986, 115, 286–295. [Google Scholar] [CrossRef]
- Thoreau, X.; Baras, E. Evaluation of surgery procedures for implanting telemetry transmitters into the body cavity of tilapia Oreochromis aureus. Aquat. Living Resour. 1997, 10, 207–211. [Google Scholar] [CrossRef]
- Arnold, G.P.; Holford, B.H. The physical effects of an acoustic tag on the swimming performance of plaice and cod. ICES J. Mar. Sci. 1978, 38, 189–200. [Google Scholar] [CrossRef]
- McCleave, J.D.; Stred, K.A. Effect of dummy telemetry transmitters on stamina of Atlantic salmon (Salmo salar) smolts. J. Fish. Board Can. 1975, 32, 559–563. [Google Scholar] [CrossRef]
- Mellas, E.J.; Haynes, J.M. Swimming performance and behavior of rainbow trout (Salmo gairdneri) and white perch (Morone americana): Effects of attaching telemetry transmitters. Can. J. Fish. Aquat. Sci. 1985, 42, 488–493. [Google Scholar] [CrossRef]
- Moore, A.; Russell, I.C.; Potter, E.C.E. The effects of intraperitoneally implanted dummy acoustic transmitters on the behaviour and physiology of juvenile Atlantic salmon, Salmo salar L. J. Fish Biol. 1990, 37, 713–721. [Google Scholar] [CrossRef]
- Pahlke, K.A.; Bernard, D.R. Abundance of the chinook salmon escapement in the Taku River, 1989 to 1990. Alsk. Fish. Res. Bull. 1996, 3, 9–20. [Google Scholar]
- Heupel, M.R.; Simpfendorfer, C.A. Estimation of mortality of juvenile blacktip sharks, Carcharhinus limbatus, within a nursery area using telemetry data. Can. J. Fish. Aquat. Sci. 2002, 59, 624–632. [Google Scholar] [CrossRef] [Green Version]
- Keefer, M.L.; Peery, C.A.; Daigle, W.R.; Jepson, M.A.; Lee, S.R.; Boggs, C.T.; Tolotti, K.R.; Burke, B.J. Escapement, harvest, and unknown loss of radio-tagged adult salmonids in the Columbia River Snake River hydrosystem. Can. J. Fish. Aquat. Sci. 2005, 62, 930–949. [Google Scholar] [CrossRef]
- Francis, C.M.; Saurola, P. Estimating age-specific survival rates of tawny owls—Recaptures versus recoveries. J. Appl. Stat. 2002, 29, 637–647. [Google Scholar] [CrossRef]
- Bjorndal, K.A.; Bolten, A.B.; Chaloupka, M.Y. Survival probability estimates for immature green turtles Chelonia mydas in the Bahamas. Mar. Ecol. Prog. Ser. 2003, 252, 273–281. [Google Scholar] [CrossRef] [Green Version]
- Gardali, T.; Barton, D.C.; White, J.D.; Geupel, G.R. Juvenile and adult survival of Swainson’s Thrush (Catharus ustulatus) in coastal California: Annual estimates using capture-recapture analyses. Auk 2003, 120, 1188–1194. [Google Scholar] [CrossRef]
- Moser, M.L.; Ocker, P.A.; Stuehrenberg, L.C.; Bjornn, T.C. Passage efficiency of adult Pacific lampreys at hydropower dams on the lower Columbia River, USA. Trans. Am. Fish. Soc. 2002, 131, 956–965. [Google Scholar] [CrossRef]
- Wagner, G.N.; Cooke, S.J. Methodological approaches and opinions of researchers involved in the surgical implantation of telemetry transmitters in fish. J. Aquat. Anim. Health 2005, 17, 160–169. [Google Scholar] [CrossRef]
- Keefer, M.L.; Moser, M.L.; Boggs, C.T.; Daigle, W.R.; Peery, C.A. Effects of body size and river environment on the upstream migration of adult Pacific lampreys. N. Am. J. Fish. Manag. 2009, 29, 1214–1224. [Google Scholar] [CrossRef]
- Hogan, Z.; Baird, I.G.; Radtke, R.; Vander Zanden, M.J. Long distance migration and marine habitation in the tropical Asian catfish, Pangasius krempfi. J. Fish Biol. 2007, 71, 818–832. [Google Scholar] [CrossRef]
- Hogan, Z.S.; Em, S.; Tach, P.; Hortle, K.G. Tagging Fish: A Case Study from the Tonle Sap, Cambodia. MRC Technical Paper No. 12; Mekong River Commission: Vientiane, Laos, 2006; p. 34. [Google Scholar]
- Pflieger, W.L. The Fishes of Missouri; Missouri Department of Conservation: Jefferson City, MO, USA, 1997. [Google Scholar]
- Vokoun, J.C.; Rabeni, C.F. Summer diel activity and movement paths of flathead catfish (Pylodictis olivaris) in two Missouri streams. Am. Midl. Nat. 2006, 155, 113–122. [Google Scholar] [CrossRef]
- Schilt, C.R. Developing fish passage and protection at hydropower dams. Appl. Anim. Behav. Sci. 2007, 104, 295–325. [Google Scholar] [CrossRef]
- Valbo-Jørgensen, J.; Poulsen, A.F. Using local knowledge as a research tool in the study of river fish biology: Experiences from the Mekong. Environ. Dev. Sustain. 2000, 2, 253–376. [Google Scholar] [CrossRef]
- Baird, I.G.; Phylavanh, B.; Vongsenesouk, B.; Xaiyamanivong, K. The ecology and conservation of the smallscale croaker Boesemania microlepis (Bleeker 1858–59) in the mainstream Mekong River, southern Laos. Nat. Hist. Bull. Siam Soc. 2001, 49, 161–176. [Google Scholar]
- Keefer, M.L.; Peery, C.A.; Bjornn, T.C.; Jepson, M.A.; Stuehrenberg, L.C. Hydrosystem, dam, and reservoir passage rates of adult Chinook salmon and steelhead in the Columbia and Snake rivers. Trans. Am. Fish. Soc. 2004, 133, 1413–1439. [Google Scholar] [CrossRef]
- Welch, D.W.; Turo, S.; Batten, S.D. Large-scale marine and freshwater movements of white sturgeon. Trans. Am. Fish. Soc. 2006, 135, 386–389. [Google Scholar] [CrossRef]
- Grumbine, R.E.; Xu, J. Mekong hydropower development. Science 2011, 332, 178–179. [Google Scholar] [CrossRef] [PubMed]
- Grumbine, R.E.; Dore, J.; Xu, J. Mekong hydropower: Drivers of change and governance challenges. Front. Ecol. Environ. 2012, 10, 91–98. [Google Scholar] [CrossRef] [Green Version]
- Soukhaphon, A.; Baird, I.G.; Hogan, Z.S. The impacts of hydropower dams in the Mekong River Basin: A review. Water 2021, 13, 265. [Google Scholar] [CrossRef]
- Yoshida, Y.; Lee, H.S.; Trung, B.H.; Tran, H.-D.; Lall, M.K.; Kakar, K.; Xuan, T.D. Impacts of mainstream hydropower dams on fisheries and agriculture in lower Mekong Basin. Sustainability 2020, 12, 2408. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Receiver Site Code | Receiver Site Name | Location (rkm) | Channel Width (km) | Number of Fish Detected | Number of Fish Missed | Detection Probability |
|---|---|---|---|---|---|---|
| GTR | Golden Triangle | 0 | 0.51 | 3 | - | - |
| CSN | Chiang Saen village | 11.5 | 0.57 | 1 | 2 | 0.33 |
| KOK | Sop Kok River | 15.7 | 0.77 | 4 | 2 | 0.67 |
| BSA | Ban Saow village | 19.7 | 0.32 | 4 | 2 | 0.67 |
| SDK | Suan Dok village | 22.7 | 0.46 | 4 | 1 | 0.80 |
| SYP | Sop Yap village | 25.1 | 0.38 | 4 | 1 | 0.80 |
| PLH | Pah Leh | 42.5 | 0.62 | 4 | 1 | 0.80 |
| PPT | Pha Pra Thai | 52.0 | 0.35 | 4 | 1 | 0.80 |
| MKN | Meuang Gan village | 54.2 | 0.32 | 0 | 3 | 0.00 |
| KGA | Kaeng Gai village | 59.3 | 0.36 | 1 | 2 | 0.33 |
| HVG | Huay Vieng village | 68.3 | 0.64 | 4 | 0 | 1.00 |
| HKR | Hat Khrai | 68.9 | 0.59 | 2 | 1 | 0.67 |
| ENG | Pak Ing River | 77.7 | 0.44 | 3 | 0 | 1.00 |
| HEN | Huay Eian village | 79.9 | 0.31 | 6 | 0 | 1.00 |
| JBJ | Jam Bong village | 85.8 | 0.21 | 0 | 4 | 0.00 |
| HLK | Huay Leuk village | 90.9 | 0.60 | 2 | 2 | 0.50 |
| PDA | Pa Dai village | 94.6 | 0.26 | 2 | - | - |
| Tag Type | Diameter (mm) | Length (mm) | Weight in Water (g) | Power Output (dB re 1 µPa @ 1 m) | Transmission Interval (s) | Rated Operating Life (d) |
|---|---|---|---|---|---|---|
| V13-1L | 13 | 36 | 6.3 | 147 | 20–60 | 360 |
| V16-4H | 16 | 68 | 10.3 | 158 | 5–30 | 309 |
| V16-6H | 16 | 95 | 14.9 | 158 | 5–30 | 618 |
| V16P-4H | 16 | 71 | 11.7 | 158 | 5–30 | 279 |
| V16P-6H | 16 | 98 | 16.3 | 158 | 5–30 | 557 |
| Species | Length (cm) | Weight (kg) | Date Tagged | Tag Type | Total Days Monitored (d) | Total Distance Traveled (rkm) | Receivers with Records (count) | Moved Upstream? (yes/no) | Recorded During Day (d) or Night (n) |
|---|---|---|---|---|---|---|---|---|---|
| B. yarrelli | 92.5 | 4.5 | 5-May | V16-4H | 17.8 | 43.8 | 6 | yes | dn |
| 72.0 | n/a | 9-May | V16-4H | 106.4 | 39.4 | 4 | yes | dn | |
| 82.0 | 4.2 | 9-May | V16-4H | 4.2 | 0.0 | 1 | no | d | |
| 77.0 | 3.9 | 11-May | V16-4H | 3.4 | 26.9 | 2 | no | dn | |
| 147.0 | 30.0 | 13-May | V16-4H | 0.0 | - | 0 | - | - | |
| 84.0 | 5.6 | 16-May | V16-6H | 140.7 | 35.8 | 5 | yes | dn | |
| 96.5 | 6.3 | 16-May | V16-6H | 354.4 | 37.2 | 2 | yes | dn | |
| 82.0 | 4.7 | 16-May | V16-6H | 94.8 | 0.0 | 1 | no | dn | |
| 121.0 | 17.5 | 19-May | V16P-6H | 6.9 | 26.9 | 1 | no | dn | |
| 106.0 | 8.0 | 20-May | V16-6H | 19.9 | 53.2 | 5 | yes | dn | |
| H. wyckioides | 81.0 | 3.5 | 8-May | V16-4H | 8.4 | 2.2 | 2 | no | n |
| 71.0 | 3.0 | 14-May | V13-1L | 0.0 | - | 0 | - | - | |
| 90.0 | 5.0 | 16-May | V16-6H | 0.5 | 2.2 | 1 | no | n | |
| 89.5 | 6.5 | 17-May | V16P-6H | 336.1 | 69.0 | 6 | yes | dn | |
| 79.0 | 4.6 | 17-May | V16-6H | 0.1 | 0.0 | 1 | no | n | |
| P. bocourti | 93.0 | 9.5 | 14-May | V16-4H | 0.8 | 11.0 | 2 | no | n |
| 77.0 | 6.4 | 19-May | V16P-4H | 0.8 | 25.7 | 2 | no | dn | |
| W. attu | 98.0 | n/a | 9-May | V16-4H | 5.3 | 88.3 | 3 | yes | dn |
| 82.5 | 3.3 | 15-May | V16-4H | 9.7 | 0.0 | 1 | no | dn | |
| P. conchophilus | 75.0 | 5.5 | 20-May | V16-4H | 0.1 | 0.6 | 1 | yes | dn |
| Species | Length (cm) | Weight (kg) | Total Days Monitored (d) | Total Distance Traveled (rkm) | Downstream Distance (rkm) | Downstream Movement Rate (rkm/d) | Upstream Distance (rkm) | Upstream Movement Rate (rkm/d) |
|---|---|---|---|---|---|---|---|---|
| B. yarrelli | 92.5 | 4.5 | 17.8 | 43.8 | - | - | 43.8 | 2.5 |
| 72.0 | n/a | 106.4 | 39.4 | 19.7 | 0.4 | 19.7 | 0.6 | |
| 82.0 | 4.2 | 4.2 | 0.0 | - | - | - | - | |
| 77.0 | 3.9 | 3.4 | 26.9 | 26.9 | 7.8 | - | - | |
| 147.0 | 30.0 | 0.0 | - | - | - | - | - | |
| 84.0 | 5.6 | 140.7 | 35.8 | 15.2 | 0.2 | 20.6 | 0.3 | |
| 96.5 | 6.3 | 354.4 | 37.2 | 19.8 | 0.1 | 17.4 | 0.3 | |
| 82.0 | 4.7 | 94.8 | 0.0 | - | - | - | - | |
| 121.0 | 17.5 | 6.9 | 26.9 | 26.9 | 4.3 | - | - | |
| 106.0 | 8.0 | 19.9 | 53.2 | - | - | 53.2 | 2.7 | |
| H. wyckioides | 81.0 | 3.5 | 8.4 | 2.2 | 2.2 | 0.3 | - | - |
| 71.0 | 3.0 | 0.0 | - | - | - | - | - | |
| 90.0 | 5.0 | 0.5 | 2.2 | 2.2 | 4.1 | - | - | |
| 89.5 | 6.5 | 336.1 | 69.0 | 16.9 | 0.1 | 52.1 | 15.3 | |
| 79.0 | 4.6 | 0.1 | 0.0 | - | - | - | - | |
| P. bocourti | 93.0 | 9.5 | 0.8 | 11.0 | 11.0 | 14.2 | - | - |
| 77.0 | 6.4 | 0.8 | 25.7 | 25.7 | 32.8 | - | - | |
| W. attu | 98.0 | n/a | 5.3 | 88.3 | 52.0 | 156.7 | 36.3 | 25.3 |
| 82.5 | 3.3 | 9.7 | 0.0 | - | - | - | - | |
| P. conchophilus | 75.0 | 5.5 | 0.1 | 0.6 | - | - | 0.6 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naughton, G.P.; Hogan, Z.S.; Campbell, T.; Graf, P.J.; Farwell, C.; Sukumasavin, N. Acoustic Telemetry Monitors Movements of Wild Adult Catfishes in the Mekong River, Thailand and Laos. Water 2021, 13, 641. https://doi.org/10.3390/w13050641
Naughton GP, Hogan ZS, Campbell T, Graf PJ, Farwell C, Sukumasavin N. Acoustic Telemetry Monitors Movements of Wild Adult Catfishes in the Mekong River, Thailand and Laos. Water. 2021; 13(5):641. https://doi.org/10.3390/w13050641
Chicago/Turabian StyleNaughton, George P., Zeb S. Hogan, Teresa Campbell, Peter J. Graf, Charles Farwell, and Naruepon Sukumasavin. 2021. "Acoustic Telemetry Monitors Movements of Wild Adult Catfishes in the Mekong River, Thailand and Laos" Water 13, no. 5: 641. https://doi.org/10.3390/w13050641