
Short-Term Response of Chlorophyll a Concentration Due to Intense Wind and Freshwater Peak Episodes in Estuaries: The Case of Fangar Bay (Ebro Delta)



Abstract
:1. Introduction
2. Materials and Methods
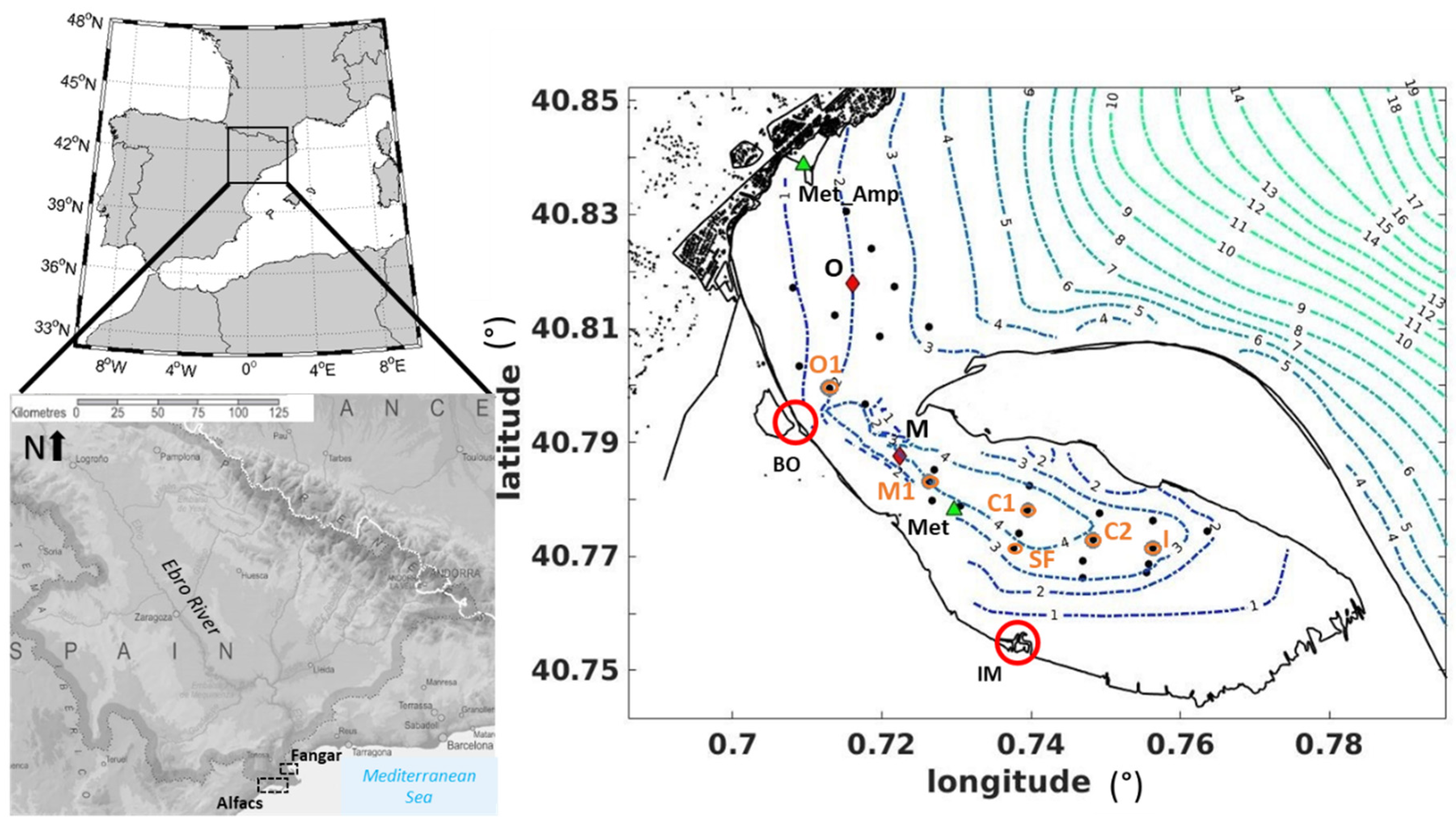
2.1. Study Area
2.2. Field Campaigns in Fangar Bay
2.3. Chlorophyll Field Data Collection
2.4. Chlorophyll Satellite Data Collection
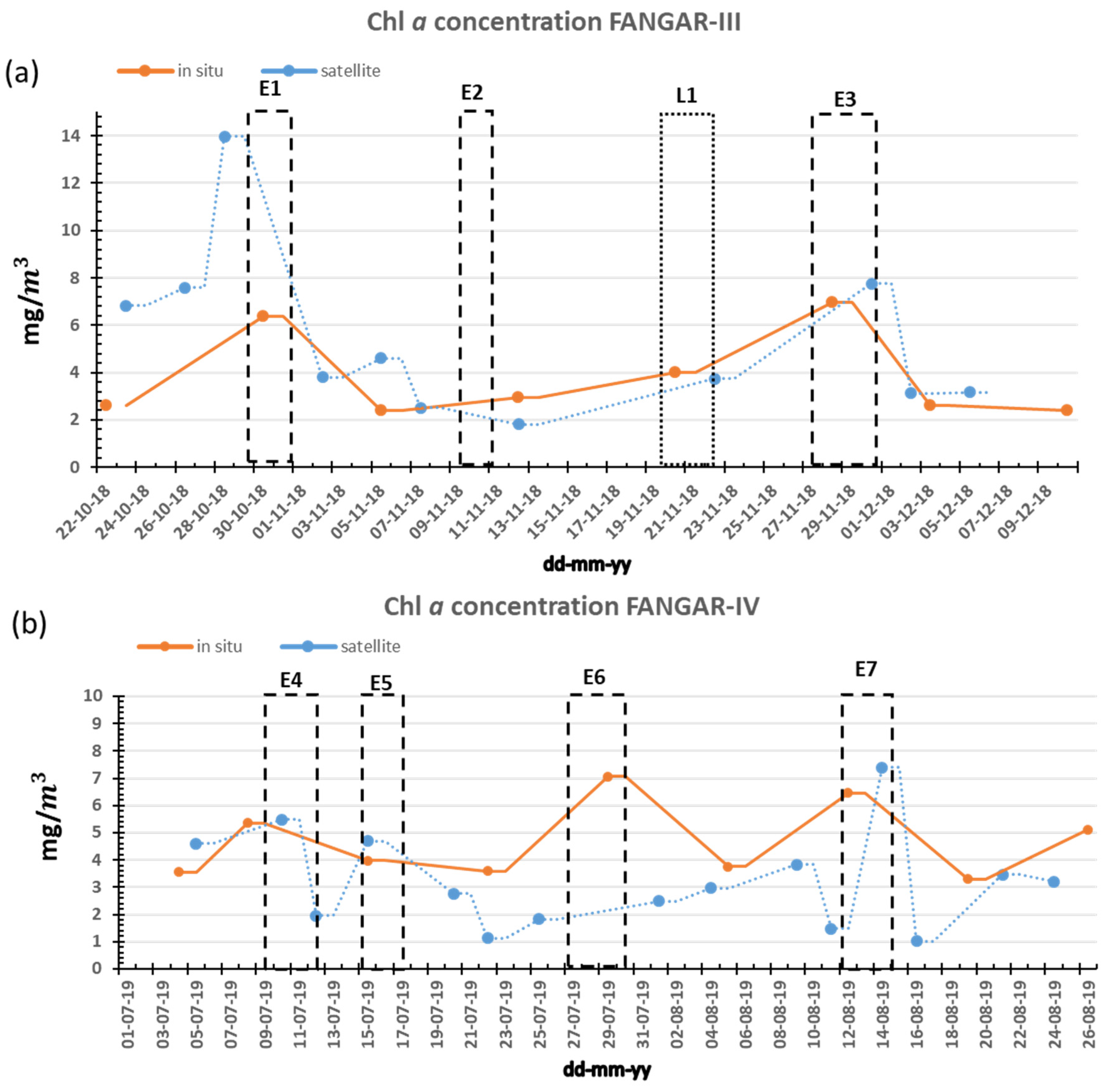
3. Results
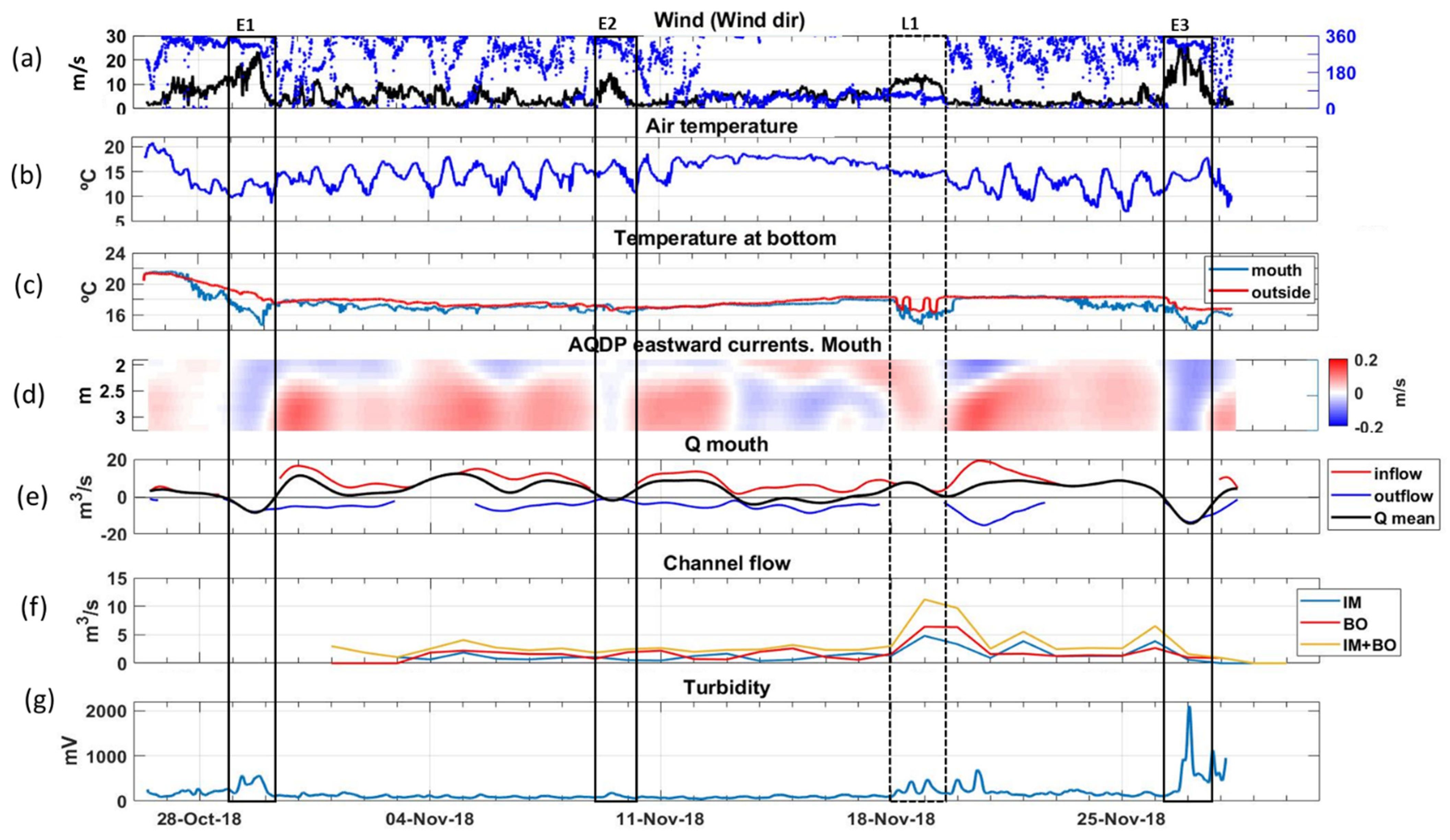
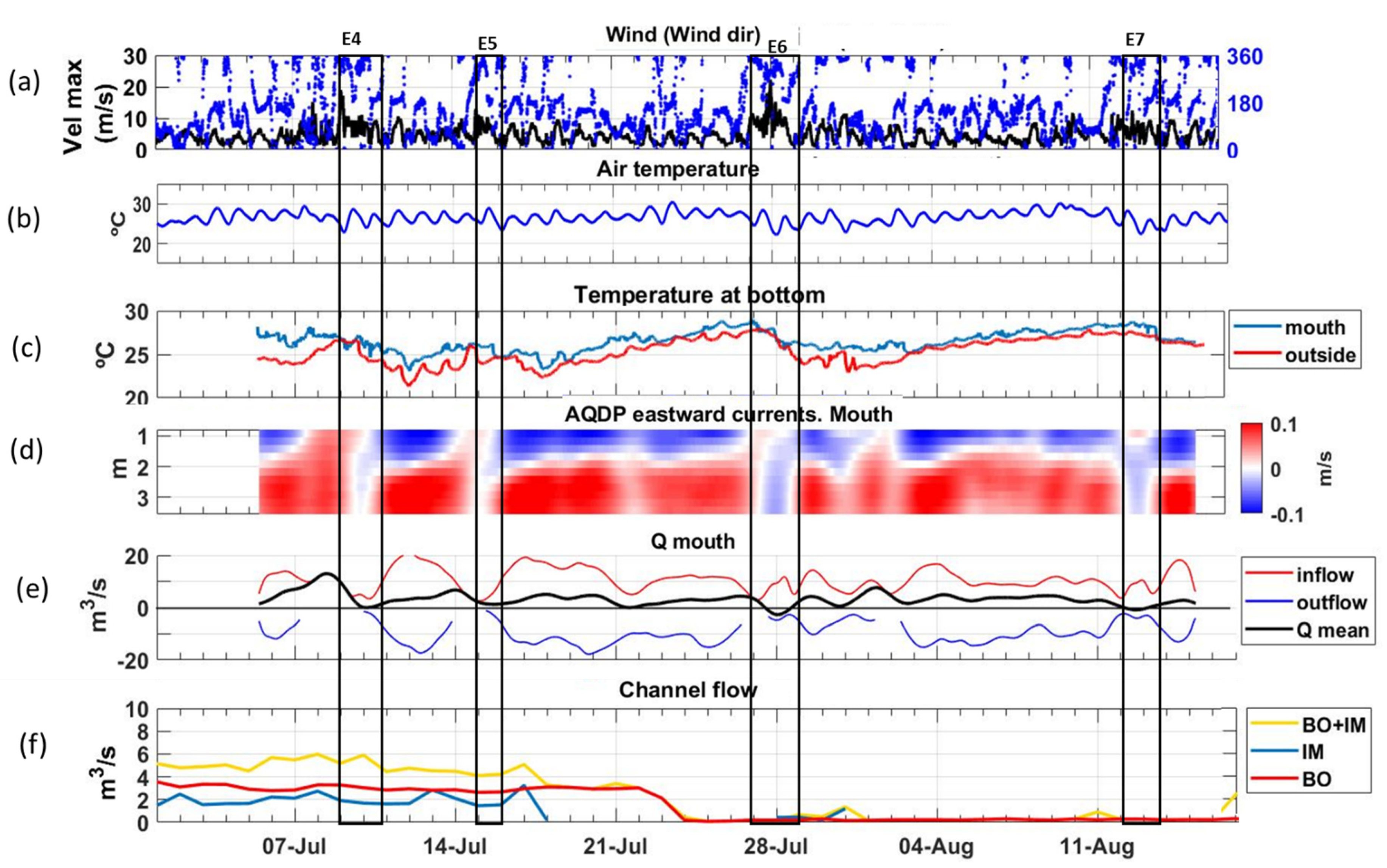
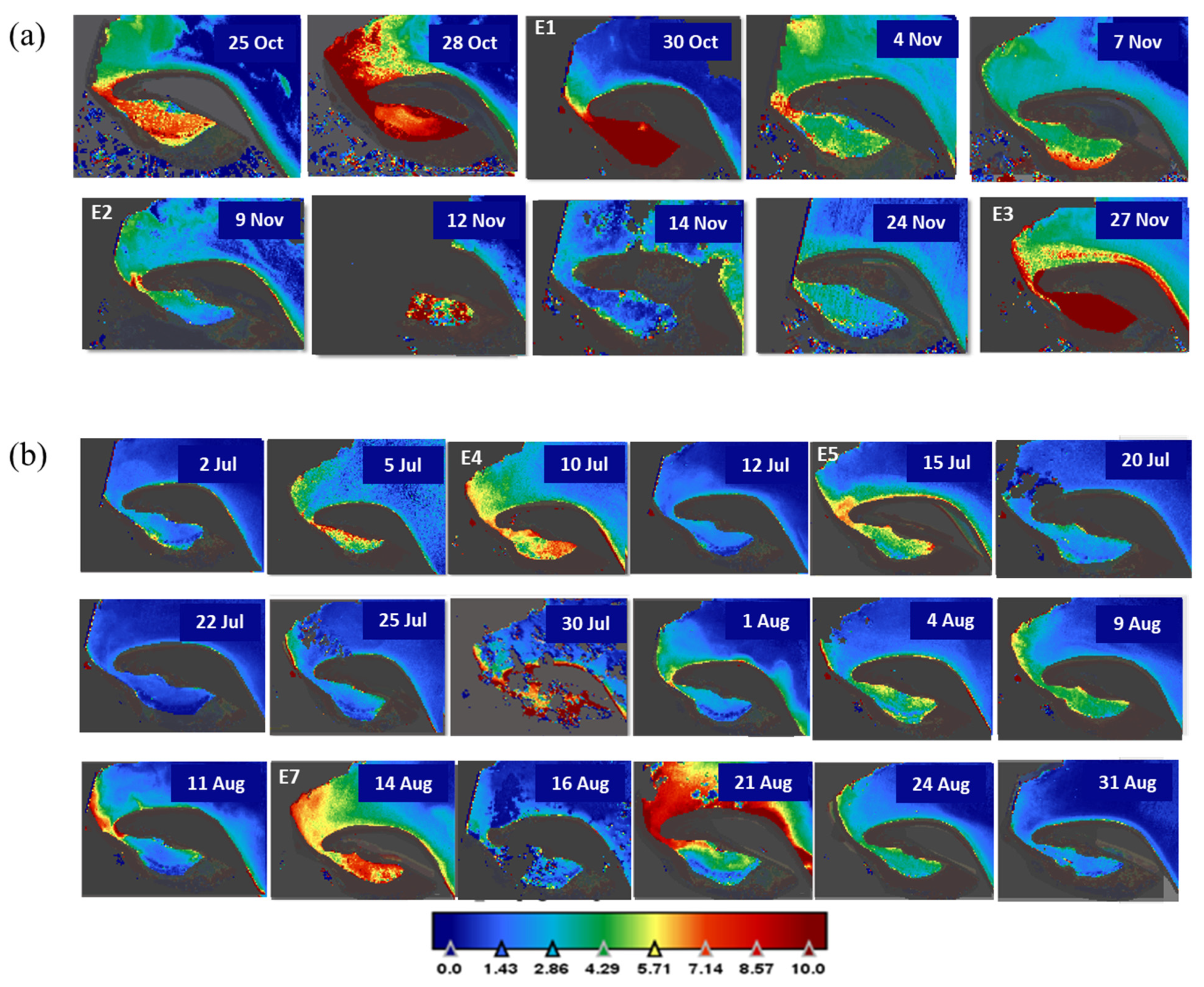
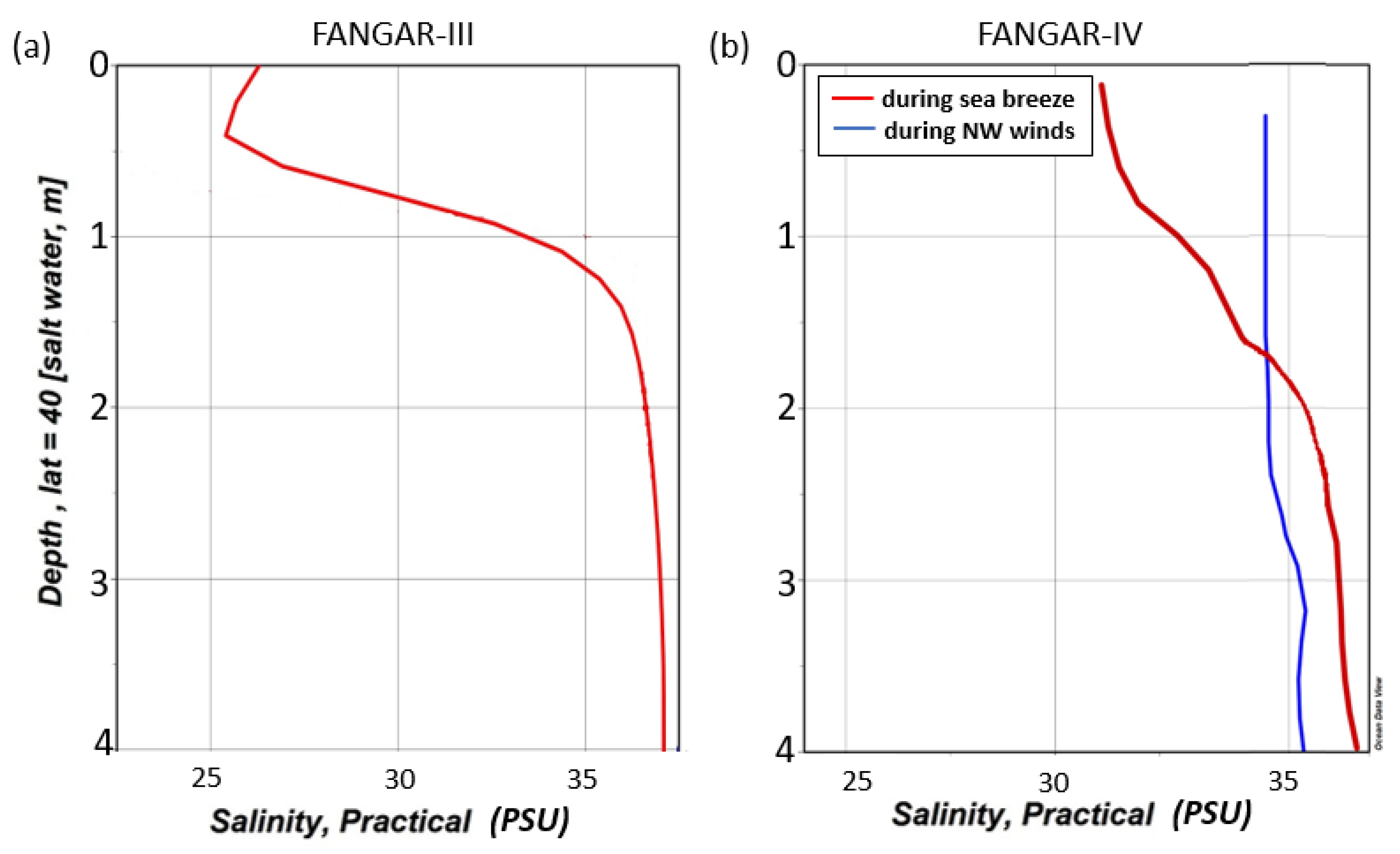
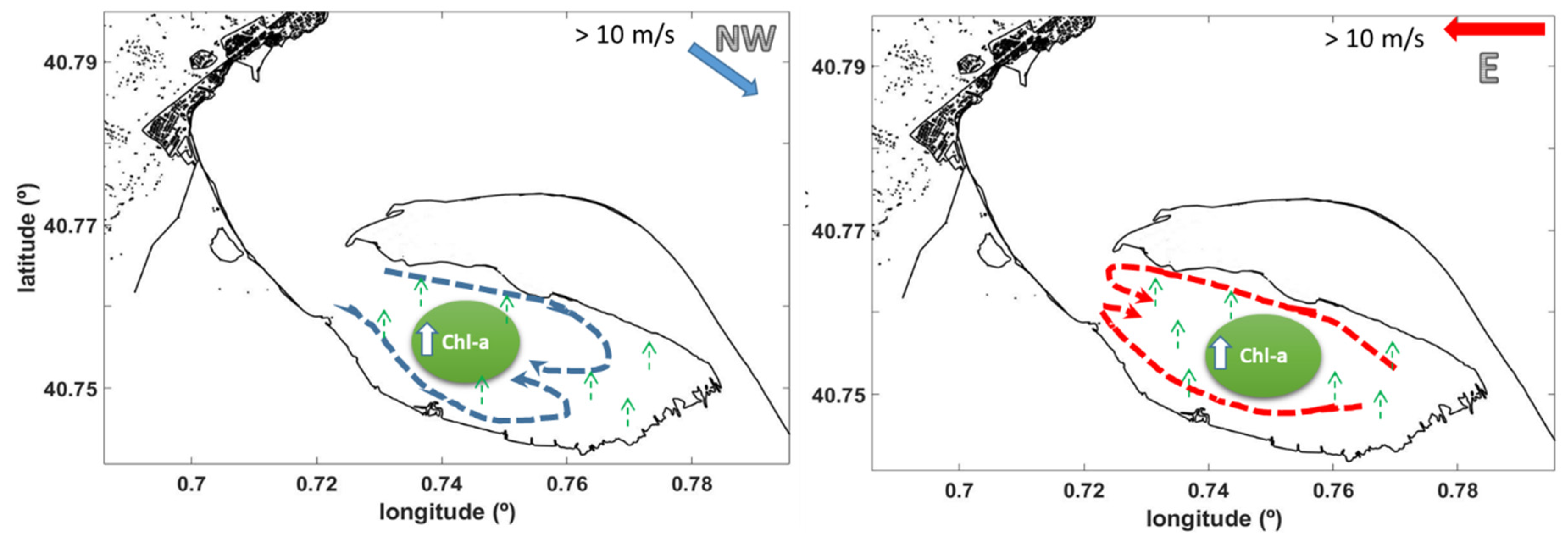
3.1. NW Wind Episode
3.2. E-NE Wind Episode
3.3. Breeze Episode
3.4. Freshwater Inputs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Geyer, W. Influence of Wind on Dynamics and Flushing of Shallow Estuaries. Estuar. Coast. Shelf Sci. 1997, 44, 713–722. [Google Scholar] [CrossRef]
- Cloern, F.H.; Nichols, J.E. Time scales and mechanisms of estuarine variability, a synthesis from studies of San Fran-cisco Bay. Hydrobiologia 1985, 29, 229–237. [Google Scholar] [CrossRef]
- Cerralbo, P.; Grifoll, M.; Valle-Levinson, A.; Espino, M. Tidal transformation and resonance in a short, microtidal Mediterranean estuary (Alfacs Bay in Ebre delta). Estuar. Coast. Shelf Sci. 2014, 145, 57–68. [Google Scholar] [CrossRef]
- Cerralbo, P.; Grifoll, M.; Espino, M. Hydrodynamic response in a microtidal and shallow bay under energetic wind and seiche episodes. J. Mar. Syst. 2015, 149, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Cerralbo, P.; Espino, M.; Grifoll, M. Modeling circulation patterns induced by spatial cross-shore wind variability in a small-size coastal embayment. Ocean Model. 2016, 104, 84–98. [Google Scholar] [CrossRef]
- Cerralbo, P.; Balsells, M.F.-P.; Mestres, M.; Fernandez, M.; Espino, M.; Grifoll, M.; Sanchez-Arcilla, A. Use of a hydrodynamic model for the management of water renovation in a coastal system. Ocean Sci. 2019, 15, 215–226. [Google Scholar] [CrossRef] [Green Version]
- Sanay, R.; Valle-Levinson, A. Wind-Induced Circulation in Semienclosed Homogeneous, Rotating Basins. J. Phys. Oceanogr. 2005, 35, 2520–2531. [Google Scholar] [CrossRef] [Green Version]
- Valle-Levinson, A.; Wong, K.-C.; Bosley, K.T. Observations of the wind-induced exchange at the entrance to Chesapeake Bay. J. Mar. Res. 2001, 59, 391–416. [Google Scholar] [CrossRef] [Green Version]
- Balsells, M.F.-P.; Grifoll, M.; Espino, M.; Cerralbo, P.; Sánchez-Arcilla, A. Wind-Driven Hydrodynamics in the Shallow, Micro-Tidal Estuary at the Fangar Bay (Ebro Delta, NW Mediterranean Sea). Appl. Sci. 2020, 10, 6952. [Google Scholar] [CrossRef]
- Llebot, C.; Sole, J.; Delgado, M.; Fernández-Tejedor, M.; Camp, J.; Estrada, M. Hydrographical forcing and phytoplankton variability in two semi-enclosed estuarine bays. J. Mar. Syst. 2011, 86, 69–86. [Google Scholar] [CrossRef]
- Soriano-González, J.; Angelats, E.; Fernández-Tejedor, M.; Diogene, J.; Alcaraz, C. First Results of Phytoplankton Spatial Dynamics in Two NW-Mediterranean Bays from Chlorophyll-a Estimates Using Sentinel 2: Potential Implications for Aquaculture. Remote Sens. 2019, 11, 1756. [Google Scholar] [CrossRef] [Green Version]
- Ramón, M.; Fernández, M.; Galimany, E. Development of mussel (Mytilus galloprovincialis) seed from two different origins in a semi-enclosed Mediterranean Bay (N.E. Spain). Aquaculture 2007, 264, 148–159. [Google Scholar] [CrossRef]
- D’Ortenzio, F.; D’Alcalà, M.R. On the trophic regimes of the Mediterranean Sea: A satellite analysis. Biogeosciences 2009, 6, 139–148. [Google Scholar] [CrossRef] [Green Version]
- Morales-Blake, A.R. Estudio Multitemporal de la Clorofila Superficial en el mar Mediterráneo Noroccidental, Evaluada a Partir de datos SeaWIFS: Septiembre de 1997 a Agosto del 2004. Ph.D. Thesis, University of Barcelona, Barcelona, Spain, 2006. [Google Scholar]
- Jou, S.; Folch, A.; Garcia-Orellana, J.; Carreño, F. Using freely available satellite thermal infrared data from Landsat 8 to identify groundwater discharge in coastal areas. Geophys. Res. Abstr. 2019, 21, 1. [Google Scholar]
- Artigas, M.; Llebot, C.; Ross, O.; Neszi, N.; Rodellas, V.; Garcia-Orellana, J.; Masque, P.; Piera, J.; Estrada, M.; Berdalet, E. Understanding the spatio-temporal variability of phytoplankton biomass distribution in a microtidal Mediterranean estuary. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2014, 101, 180–192. [Google Scholar] [CrossRef]
- Demers, S.; LaFleur, P.E.; Legendre, L.; Trump, C.L. Short-Term Covariability of Chlorophyll and Temperature in the St. Lawrence Estuary. J. Fish. Res. Board Can. 1979, 36, 568–573. [Google Scholar] [CrossRef]
- Geyer, N.L.; Huettel, M.; Wetz, M.S. Phytoplankton Spatial Variability in the River-Dominated Estuary, Apalachicola Bay, Florida. Chesap. Sci. 2018, 41, 2024–2038. [Google Scholar] [CrossRef]
- Masson, D.; Peña, A. Chlorophyll distribution in a temperate estuary: The Strait of Georgia and Juan de Fuca Strait. Estuar. Coast. Shelf Sci. 2009, 82, 19–28. [Google Scholar] [CrossRef]
- Delgado, M.; Camp, J. Abundancia y distribución de nutrientes inorgánicos disueltos en las bahías del delta del Ero. Inv. Pesq. 1987, 51, 427–441. [Google Scholar]
- Garcia, M.A.; Ballester, A. Notas acerca de la meteorología y la circulación local en la región del delta del Ebro. Inv. Pesq. 1984, 48, 469–493. [Google Scholar]
- Archetti, G.; Bernia, S.; Salvà-Catarineu, M. Análisis de los vectores ambientales que afectan la calidad del medio en la bahía del Fangar (Delta del Ebro) mediante herramientas SIG. Rev. Int. Cienc. Tecnol. Inf. Geogr. 2010, 10, 252–279. [Google Scholar]
- Bolaños, R.; Jorda, G.; Cateura, J.; Lopez, J.; Puigdefàbregas, J.; Gómez, J.; Espino, M. The XIOM: 20 years of a regional coastal observatory in the Spanish Catalan coast. J. Mar. Syst. 2009, 77, 237–260. [Google Scholar] [CrossRef] [Green Version]
- Grifoll, M.; Navarro, J.; Pallares, E.; Ràfols, L.; Espino, M.; Palomares, A. Ocean–atmosphere–wave characterisation of a wind jet (Ebro shelf, NW Mediterranean Sea). Nonlinear Process. Geophys. 2016, 23, 143–158. [Google Scholar] [CrossRef] [Green Version]
- Muñoz, I. Limnología de la part baixa del riu ebre i els canals de reg: Els factors fisico-quimics, el fitoplancton i els macroinvetebrats bentonics. Tesis para la postulación al grado de doctor. Ph.D. Thesis, Departamento de Ecología, Facultad de Biología, Universidad de Barcelona, Barcelona, Spain, 1990. [Google Scholar]
- Automatic Water Quality Information System, DEL EBRO, CHE-Hydrographic Confederation, Quality Alert Network, SAICA Project. 2013. Available online: https://www.saica.co.za/ (accessed on 30 January 2020).
- Perez, J.; Marta, C. Distribución espacial y biomasa de las fanerógamas marinas de las bahías del delta del Ebro. Inv. Pesq. 1986, 50, 519–530. [Google Scholar]
- Camp, J.; Delgado, M. Hidrogafia de las bahías del delta del Ebro. Inv. Pesq. 1987, 51, 351–369. [Google Scholar]
- Welschmeyer, N.A. Fluorometric analysis of chlorophyll a in the presence of chlorophyll b and pheopigments. Limnol. Oceanogr. 1994, 39, 1985–1992. [Google Scholar] [CrossRef]
- ESA. SENTINEL-2 User Handbook; ESA: Pairs, France, 2015. [Google Scholar]
- Mishra, D.R.; Mishra, S. Normalized Difference Chlorophyll Index: A Novel Model for Remote Estimation of Chloro-phyll-a Concentration in Turbid Productive Waters. Remote Sens. Environ. 2012, 117, 394–406. [Google Scholar] [CrossRef]
- Brockmann, C.; Doerffer, R.; Peters, M.; Stelzer, K.; Embacher, S.; Ruescas, A. Evolution of the c2rcc neural network for sentinel 2 and 3 for the retrieval of ocean colour products in normal and extreme optically complex waters. Living Planet Symp. 2016, 740, 54. [Google Scholar]
- Shintani, T.; De La Fuente, A.; Niño, Y.; Imberger, J. Generalizations of the Wedderburn number: Parameterizing upwelling in stratified lakes. Limnol. Oceanogr. 2010, 55, 1377–1389. [Google Scholar] [CrossRef]
- De Madariaga, I. Photosynthetic Characteristics of Phytoplankton during the Development of a Summer Bloom in the Urdaibai Estuary, Bay of Biscay. Estuar. Coast. Shelf Sci. 1995, 40, 559–575. [Google Scholar] [CrossRef]
- Pinckney, J.L.; Paerl, H.W.; Harrington, M.B.; Howe, K.E. Annual cycles of phytoplankton community-structure and bloom dynamics in the Neuse River Estuary, North Carolina. Mar. Biol. 1998, 131, 371–381. [Google Scholar] [CrossRef]
- Delgado, M. Abundance and distribution of microphytobenthos in the bays of Ebro Delta (Spain). Estuar. Coast. Shelf Sci. 1989, 29, 183–194. [Google Scholar] [CrossRef]
- Díez-Minguito, M.; De Swart, H.E. Relationships between Chlorophyll-a and Suspended Sediment Concentration in a High-Nutrient Load Estuary: An Observational and Idealized Modeling Approach. J. Geophys. Res. Oceans 2020, 125. [Google Scholar] [CrossRef]
- Loureiro, S.; Garcés, E.; Fernández-Tejedor, M.; Vaqué, D.; Camp, J. Pseudo-nitzschia spp. (Bacillariophyceae) and dissolved organic matter (DOM) dynamics in the Ebro Delta (Alfacs Bay, NW Mediterranean Sea). Estuar. Coast. Shelf Sci. 2009, 83, 539–549. [Google Scholar] [CrossRef] [Green Version]
- De Jorge, V.N.; Van Beusekom, J.E.E. Wind- and tide-induced resuspension of sediment and microphytobenthos from tidal flats in the Ems estuary. Limnol. Oceanogr. 1995, 40, 776–778. [Google Scholar] [CrossRef] [Green Version]
- Sondergaard, M.; Kristensen, P.; Jeppesen, E. Phosphorus release from ressuspended sediment in the shallow and wind-exposed Lake Arreso, Denmark. Hydrobiologia 1992, 228, 91–99. [Google Scholar] [CrossRef]
- Grifoll, M.; Cerralbo, P.; Guillén, J.; Espino, M.; Hansen, L.B.; Sánchez-Arcilla, A. Characterization of bottom sediment resuspension events observed in a micro-tidal bay. Ocean Sci. 2019, 15, 307–319. [Google Scholar] [CrossRef] [Green Version]
- Roque, A.; Lopez-Joven, C.; Lacuesta, B.; Elandaloussi, L.; Wagley, S.; Furones, M.D.; Ruiz-Zarzuela, I.; De Blas, I.; Rangdale, R.; Gomez-Gil, B. Detection and identification of tdh- And trh-positive Vibrio parahaemolyticus strains from four species of cultured bivalve molluscs on the Spanish Mediterranean coast. Appl. Environ. Microbiol. 2009, 75, 7574–7577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarangi, R.K.; Nayak, S.; Panigraphy, R.C. Monthly variability of chlorophyll and associated physical parameters in the southwest Bay of Bengal water using remote sensing data. Indian J. Mar. Sci. 2008, 37, 256–266. [Google Scholar]
- Llebot, C.; Rueda, F.J.; Sole, J.; Artigas, M.L.; Estrada, M. Hydrodynamic states in a wind-driven microtidal estuary (Alfacs Bay). J. Sea Res. 2014, 85, 263–276. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name (ID) | Observations | Period | Data Interval (min) |
|---|---|---|---|
| Meteo station | Wind and atmospheric pressure | 19 October–28 November 2018 | 10 |
| 25 June–6 September 2019 | |||
| ADCP and OBS mouth (M) | Currents, sea level, waves, bottom temperature, and turbidity | 26 October–28 November 2018 | 10 |
| 5 July–6 September 2019 | |||
| ADCP and OBS outside bay (O) | Currents, sea level, waves, bottom temperature, and turbidity | 26 October–28 November 2018 | 10 |
| 5 July–6 September 2019 | |||
| CTD | Temperature and salinity | November 2018 | - |
| 4 July–4 September 2019 | |||
| Bottle samples | Chl a | October–November 2018 | - |
| July–August 2019 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
F-Pedrera Balsells, M.; Grifoll, M.; Fernández-Tejedor, M.; Espino, M. Short-Term Response of Chlorophyll a Concentration Due to Intense Wind and Freshwater Peak Episodes in Estuaries: The Case of Fangar Bay (Ebro Delta). Water 2021, 13, 701. https://doi.org/10.3390/w13050701
F-Pedrera Balsells M, Grifoll M, Fernández-Tejedor M, Espino M. Short-Term Response of Chlorophyll a Concentration Due to Intense Wind and Freshwater Peak Episodes in Estuaries: The Case of Fangar Bay (Ebro Delta). Water. 2021; 13(5):701. https://doi.org/10.3390/w13050701
Chicago/Turabian StyleF-Pedrera Balsells, Marta, Manel Grifoll, Margarita Fernández-Tejedor, and Manuel Espino. 2021. "Short-Term Response of Chlorophyll a Concentration Due to Intense Wind and Freshwater Peak Episodes in Estuaries: The Case of Fangar Bay (Ebro Delta)" Water 13, no. 5: 701. https://doi.org/10.3390/w13050701