
Changes in Pelagic Fish Community Composition, Abundance, and Biomass along a Productivity Gradient in Subtropical Lakes

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
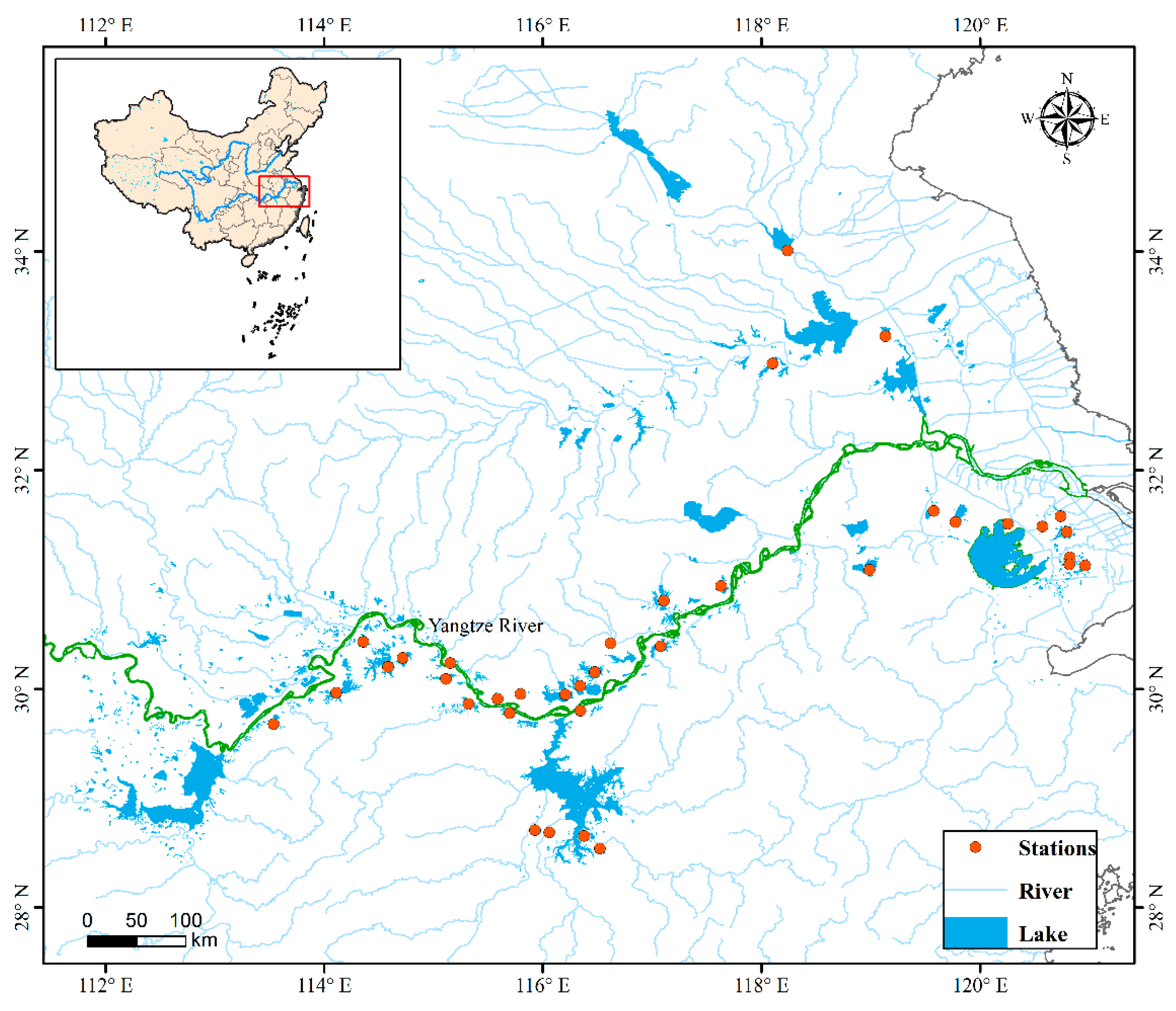
2.1. Sample Collection and Analysis
2.2. Data Analysis
3. Results
3.1. Physicochemical Parameters of the Studied Lakes
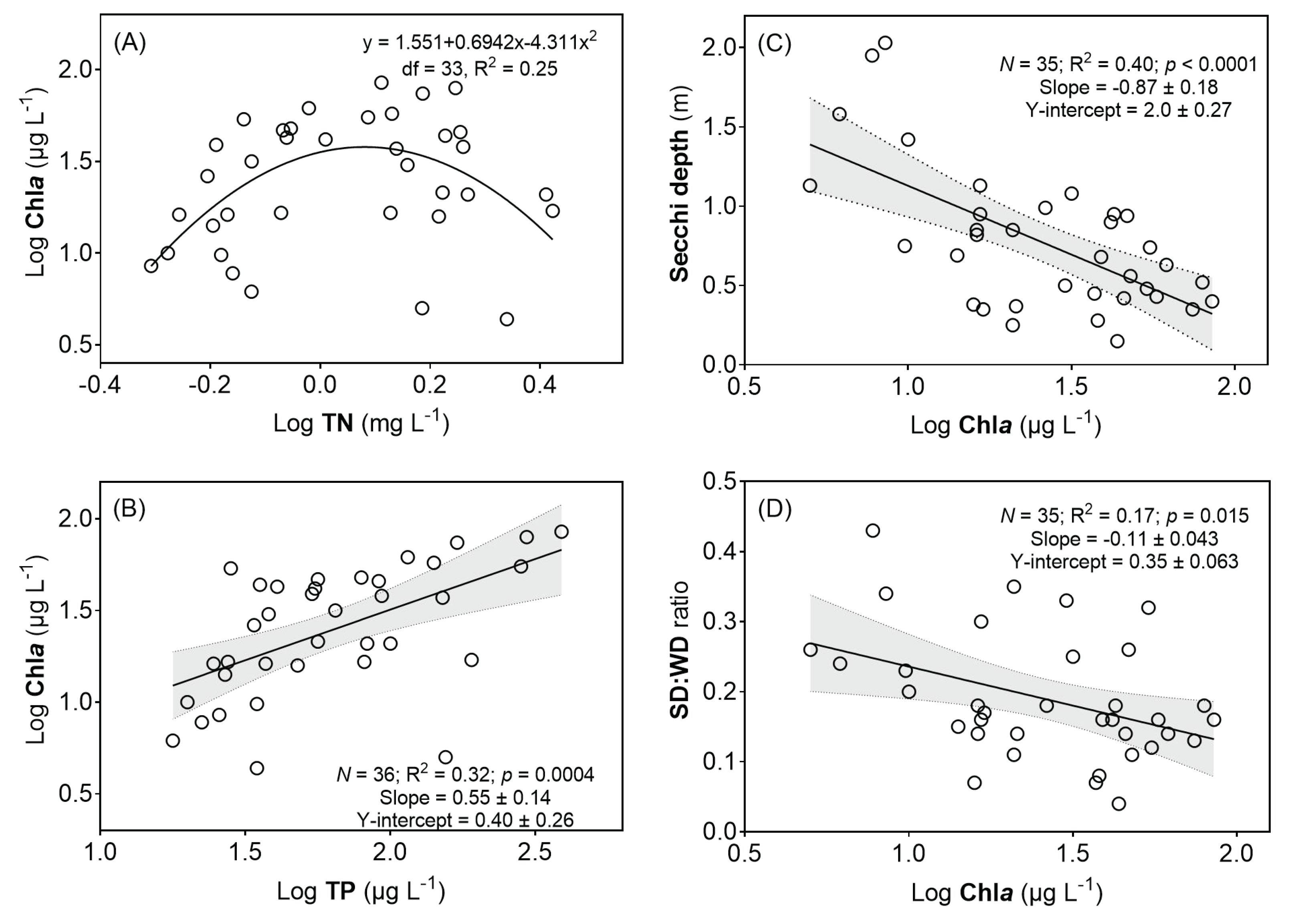
3.2. Relationships between Chla and Lake Nutrients and Transparency and Results of GLM Analysis
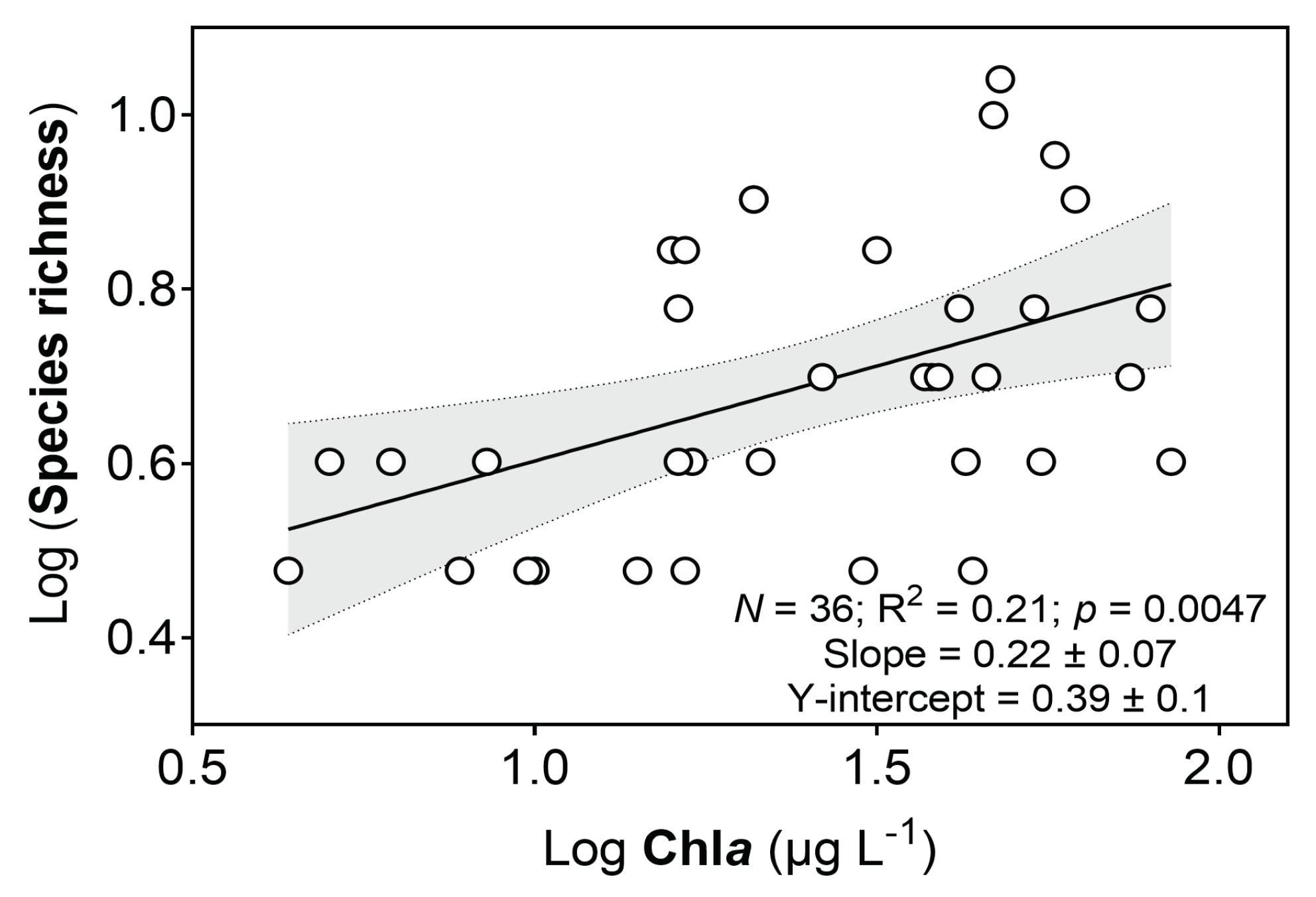
3.3. Fish Species Richness along the Chla Gradient
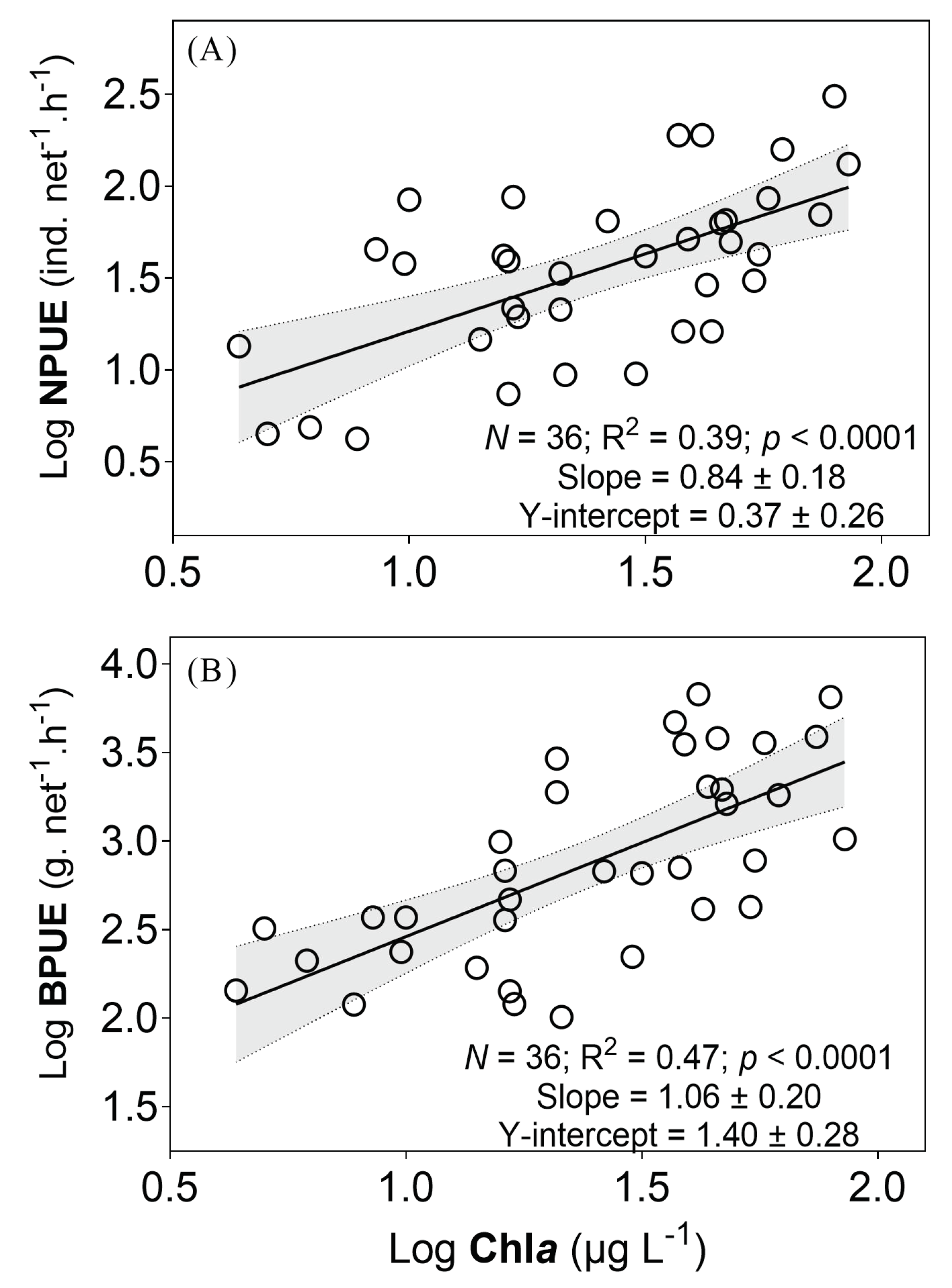
3.4. Relationships of Fish Catch with Lake Productivity
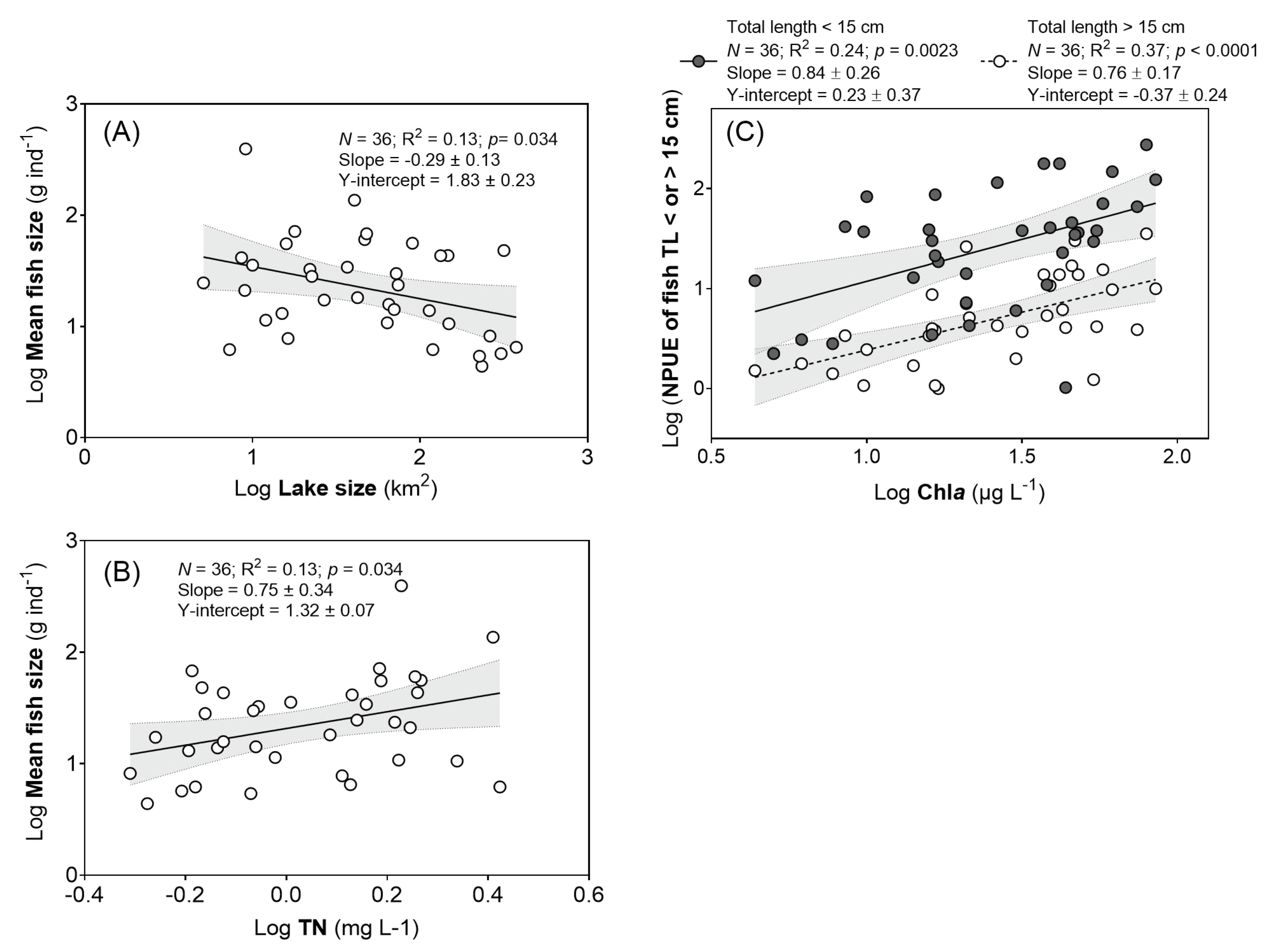
3.5. Fish Size and NPUE of Different Size Classes of Fish
3.6. Composition of Fish Community Structure and Trophic Groups
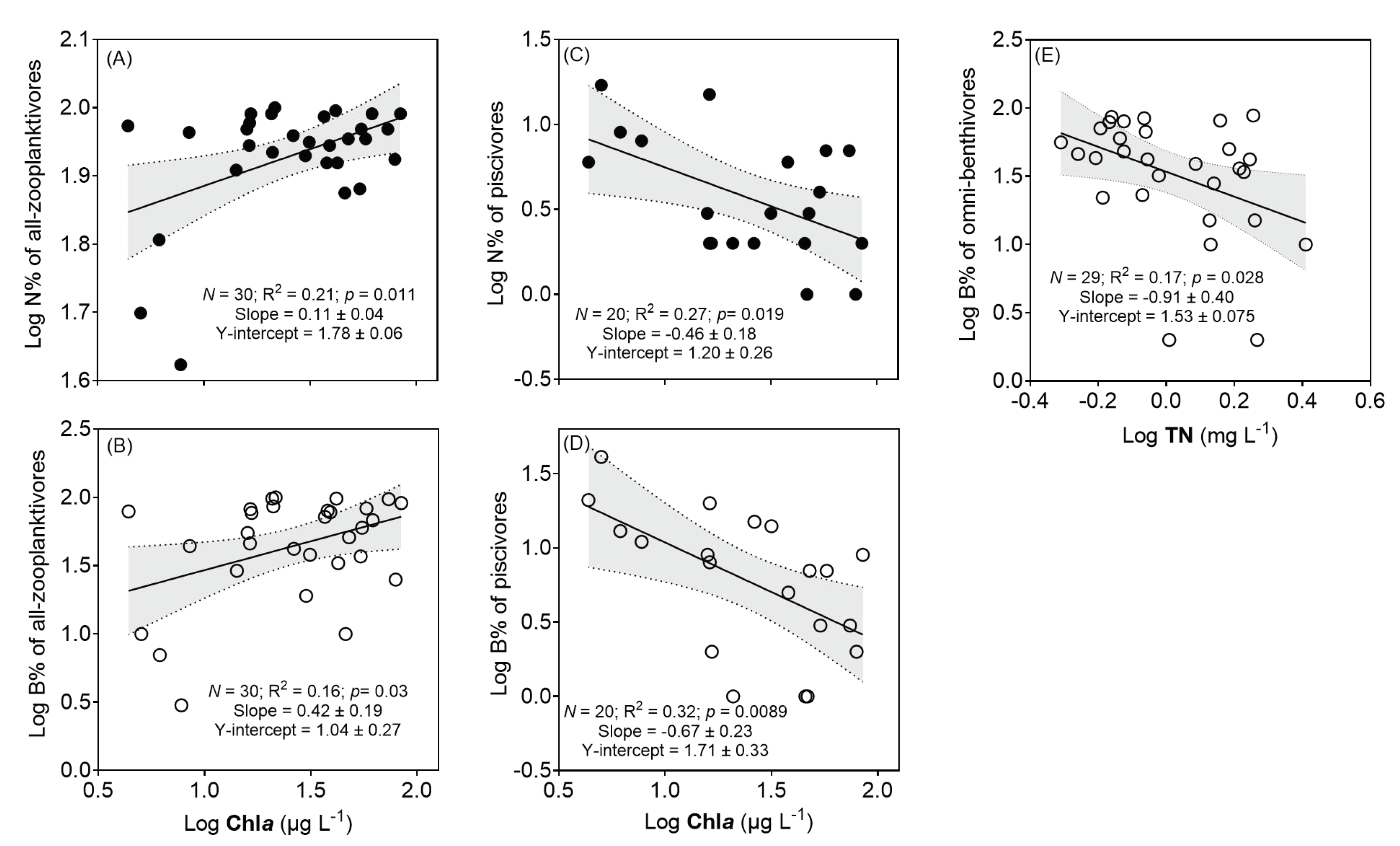
3.7. Correlations of Different Trophic Groups of Fish with Chla and TN
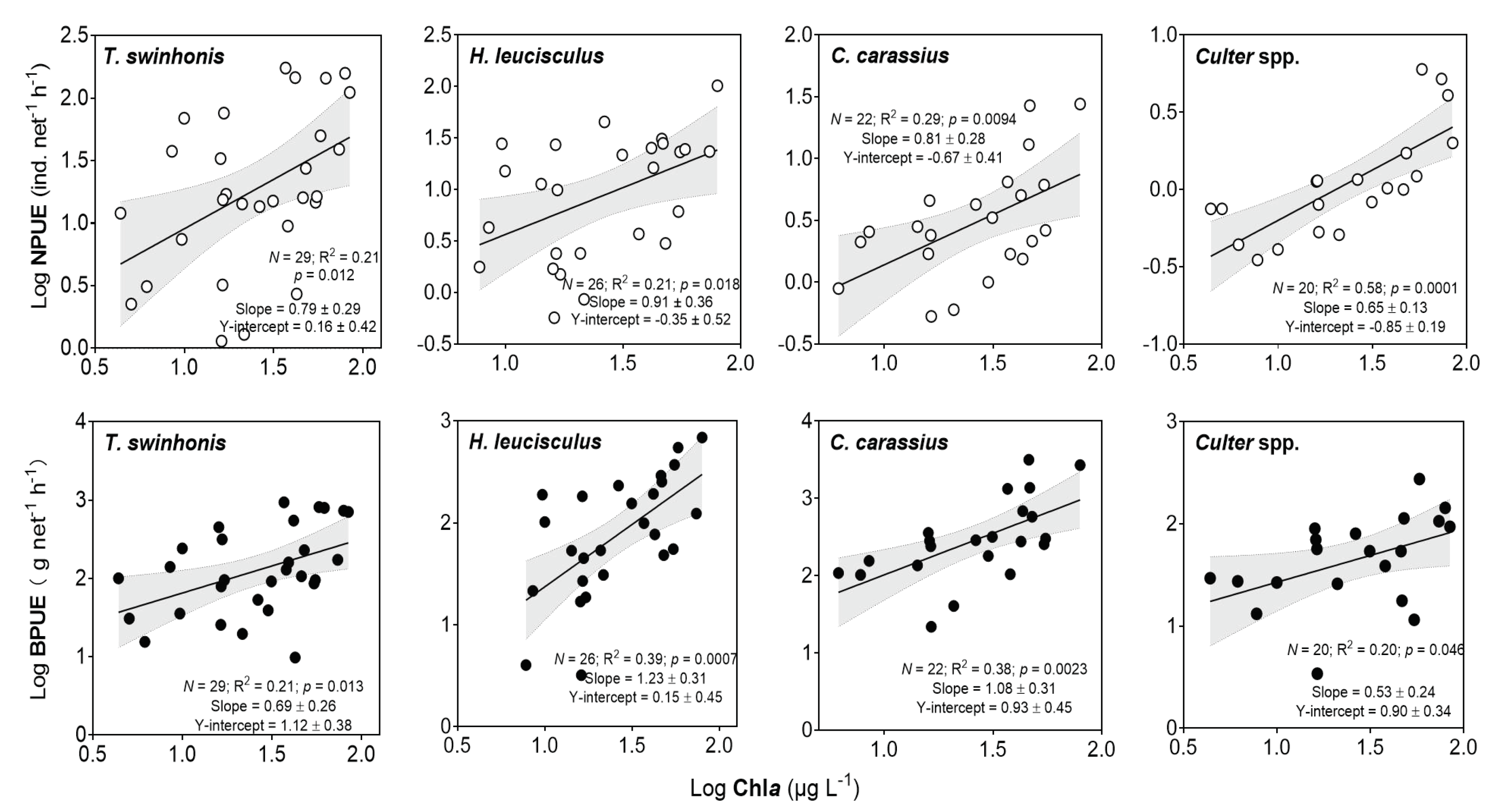
3.8. Relationships between the Catch of Three Dominant Species and Piscivorous Fish and Chla
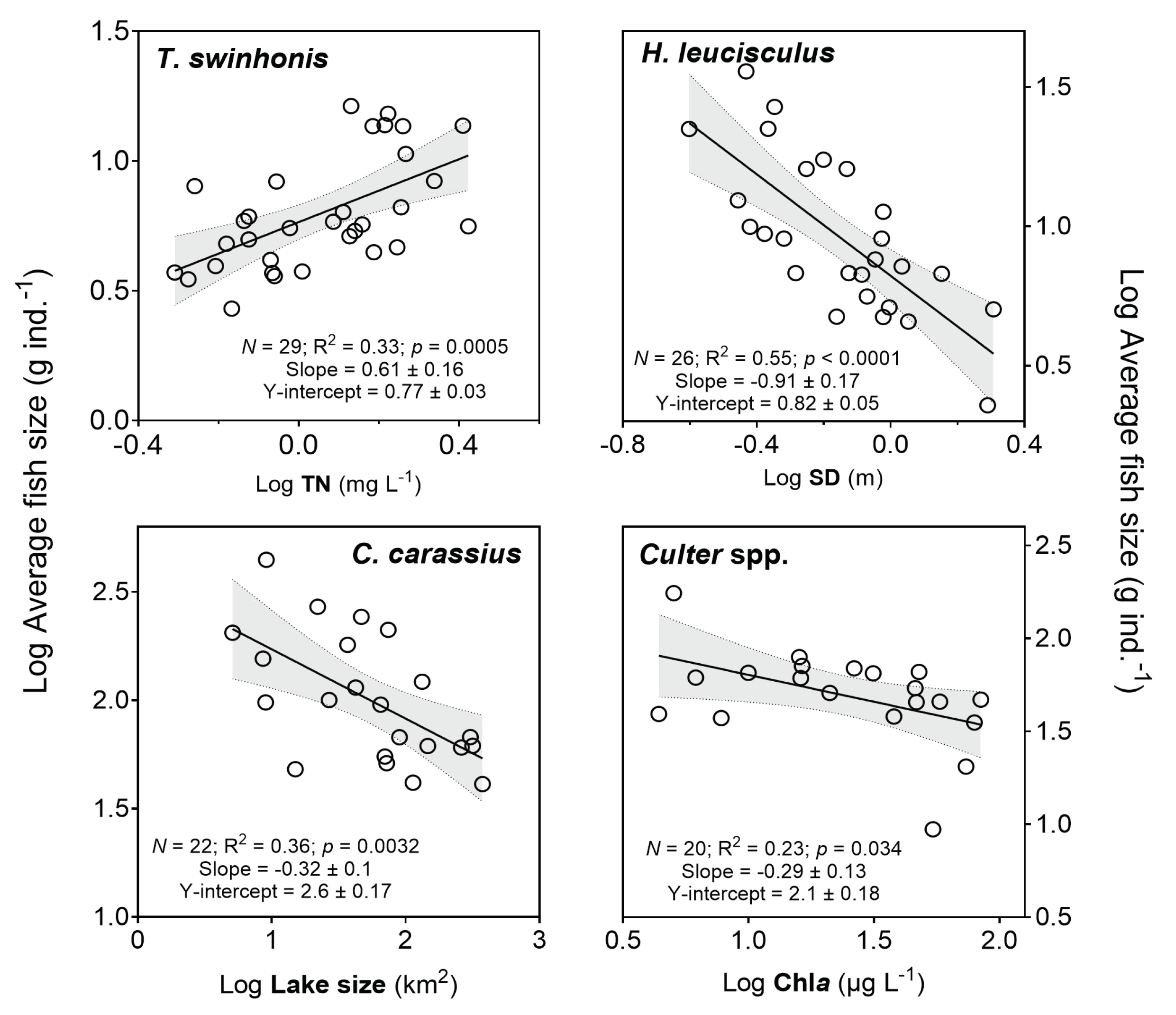
3.9. Relationships between Mean Body Weight of Three Dominant Species and Piscivorous Fish and Environmental Variables
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Brooks, J.L.; Dodson, S.I. Predation, body size, and composition of plankton. Science 1965, 150, 28–35. [Google Scholar] [CrossRef]
- Lazzaro, X. A review of planktivorous fishes: Their evolution, feeding behaviours, selectivities, and impacts. Hydrobiologia 1987, 146, 97–167. [Google Scholar] [CrossRef]
- Jeppesen, E.; Lauridsen, T.L.; Mitchell, S.F.; Christoffersen, K.; Burns, C.W. Trophic structure in the pelagial of 25 shallow New Zealand lakes: Changes along nutrient and fish gradients. J. Plankton Res. 2000, 22, 951–968. [Google Scholar] [CrossRef] [Green Version]
- Jeppesen, E.; Søndergaard, M.; Lauridsen, T.L.; Davidson, T.A.; Liu, Z.; Mazzeo, N.; Trochine, C.; Özkan, K.; Jensen, H.S.; Trolle, D.; et al. Biomanipulation as a restoration tool to combat eutrophication: Recent advances and future challenges. Adv. Ecol. Res. 2012, 47, 411–488. [Google Scholar] [CrossRef]
- Meijer, M.-L.; De Haan, M.W.; Breukelaar, A.W.; Buiteveld, H.; Donk, E. Is reduction of the benthivorous fish an important cause of high transparency following biomanipulation in shallow lakes? Hydrobiologia 1990, 200, 303–315. [Google Scholar] [CrossRef]
- Persson, L.; Andersson, G.; Hamrin, S.F.; Johansson, L. Predator regulation and primary production along the productivity gradient of temperate lake ecosystems. In Complex Interactions in Lake Communities; Carpenter, S.R., Ed.; Springer: New York, NY, USA, 1988; pp. 45–65. ISBN 978-1-4612-8372-0. [Google Scholar]
- Olin, M.; Rask, M.; Ruuhljärvi, J.; Kurkilahti, M.; Ala-Opas, P.; Ylönen, O. Fish community structure in mesotrophic and eutrophic lakes of southern Finland: The relative abundances of percids and cyprinids along a trophic gradient. J. Fish Biol. 2002, 60, 593–612. [Google Scholar] [CrossRef]
- Mehner, T.; Diekmann, M.; Brämick, U.; Lemcke, R. Composition of fish communities in German lakes as related to lake morphology, trophic state, shore structure and human-use intensity. Freshw. Biol. 2005, 50, 70–85. [Google Scholar] [CrossRef]
- Jeppesen, E.; Søndergaard, M.; Jensen, J.P.; Havens, K.E.; Anneville, O.; Carvalho, L.; Coveney, M.F.; Deneke, R.; Dokulil, M.T.; Foy, B.; et al. Lake responses to reduced nutrient loading—an analysis of contemporary long-term data from 35 case studies. Freshw. Biol. 2005, 50, 1747–1771. [Google Scholar] [CrossRef]
- Brucet, S.; Pédron, S.; Mehner, T.; Lauridsen, T.L.; Argillier, C.; Winfield, I.J.; Volta, P.; Emmrich, M.; Hesthagen, T.; Holmgren, K.; et al. Fish diversity in European lakes: Geographical factors dominate over anthropogenic pressures. Freshw. Biol. 2013, 58, 1779–1793. [Google Scholar] [CrossRef] [Green Version]
- De Robertis, A.; Ryer, C.H.; Veloza, A.; Brodeur, R.D. Differential effects of turbidity on prey consumption of piscivorous and planktivorous fish. Can. J. Fish. Aquat. Sci. 2003, 60, 1517–1526. [Google Scholar] [CrossRef]
- Jia, J.; Shi, W.; Chen, Q.; Lauridsen, T.L. Spatial and temporal variations reveal the response of zooplankton to cyanobacteria. Harmful Algae 2017, 64, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Gong, Z.; Xie, P. Impact of eutrophication on biodiversity of the Macrozoobenthos community in a Chinese shallow lake. J. Freshw. Ecol. 2001, 16, 171–178. [Google Scholar] [CrossRef] [Green Version]
- Cai, Y.; Gong, Z.; Qin, B. Influences of habitat type and environmental variables on benthic macroinvertebrate communities in a large shallow subtropical lake (Lake Taihu, China). Ann. Limnol. Int. J. Limnol. 2011, 47, 85–95. [Google Scholar] [CrossRef] [Green Version]
- Cai, Y.; Gong, Z.; Qin, B. Benthic macroinvertebrate community structure in Lake Taihu, China: Effects of trophic status, wind-induced disturbance and habitat complexity. J. Great Lakes Res. 2012, 38, 39–48. [Google Scholar] [CrossRef]
- Mehner, T.; Diekmann, M.; Gonsiorczyk, T.; Kasprzak, P.; Koschel, R.; Krienitz, L.; Rumpf, M.; Schulz, M.; Wauer, G. Rapid recovery from eutrophication of a stratified lake by disruption of internal nutrient load. Ecosystems 2008, 11, 1142–1156. [Google Scholar] [CrossRef]
- Hartmann, J.; Nümann, W. Percids of lake constance, a lake undergoing eutrophication. J. Fish. Res. Board Can. 1977, 34, 1670–1677. [Google Scholar] [CrossRef]
- Persson, L. Competition, predation and environmental factors as structuring forces in freshwater fish communities: Sumari (1971) revisited. Can. J. Fish. Aquat. Sci. 1997, 54, 85–88. [Google Scholar] [CrossRef]
- Perrow, M.R.; Jowitt, A.J.D.; Leigh, S.A.C.; Hindes, A.M.; Rhodes, J.D. The stability of fish communities in shallow lakes undergoing restoration: Expectations and experiences from the Norfolk Broads (U.K.). Hydrobiologia 1999, 408, 85–100. [Google Scholar] [CrossRef]
- Teixeira-de Mello, F.; Meerhoff, M.; Jeppesen, E.; Pekcan-Hekim, Z. Substantial differences in littoral fish community structure and dynamics in subtropical and temperate shallow lakes. Freshw. Biol. 2009, 54, 1202–1215. [Google Scholar] [CrossRef]
- Mao, Z.; Gu, X.; Zeng, Q.; Zhou, L.; Sun, M. Status and changes of fishery resources (2009–2010) in Lake Taihu and their responses to water eutrophication. J. Lake Sci. 2011, 23, 967–973. [Google Scholar]
- Hoyer, M.V.; Bennett, J.P.; Canfield, D.E. Monitoring freshwater fish in Florida lakes using electrofishing: Lessons learned. Lake Reserv. Manag. 2011, 27, 329–342. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Liu, Z.; Jeppesen, E. Fish community assemblages changed but biomass remained similar after lake restoration by biomanipulation in a Chinese tropical eutrophic lake. Hydrobiologia 2014, 724, 127–140. [Google Scholar] [CrossRef]
- Boll, T.; Levi, E.E.; Bezirci, G.; Özuluğ, M.; Tavşanoğlu, Ü.N.; Çakıroğlu, A.I.; Özcan, S.; Brucet, S.; Jeppesen, E.; Beklioğlu, M. Fish assemblage and diversity in lakes of western and central Turkey: Role of geo-climatic and other environmental variables. Hydrobiologia 2016, 771, 31–44. [Google Scholar] [CrossRef]
- Jin, B.-S.; Winemiller, K.O.; Shao, B.; Si, J.-K.; Jin, J.-F.; Ge, G. Fish assemblage structure in relation to seasonal environmental variation in sub-lakes of the Poyang Lake floodplain, China. Fish. Manag. Ecol. 2019, 26, 131–140. [Google Scholar] [CrossRef]
- González-Bergonzoni, I.; Meerhoff, M.; Davidson, T.A.; Teixeira-de Mello, F.; Baattrup-Pedersen, A.; Jeppesen, E. Meta-analysis shows a consistent and strong latitudinal pattern in fish omnivory across ecosystems. Ecosystems 2012, 15, 492–503. [Google Scholar] [CrossRef]
- Jeppesen, E.; Jensen, J.P.; Søndergaard, M.; Lauridsen, T.L.; Landkildehus, F. Trophic structure, species richness and biodiversity in Danish lakes: Changes along a phosphorus gradient. Freshw. Biol. 2000, 45, 201–218. [Google Scholar] [CrossRef]
- Meerhoff, M.; Clemente, J.M.; Teixeira De Mello, F.; Iglesias, C.; Pedersen, A.R.; Jeppesen, E. Can warm climate-related structure of littoral predator assemblies weaken the clear water state in shallow lakes? Glob. Chang. Biol. 2007, 13, 1888–1897. [Google Scholar] [CrossRef]
- Ye, S.; Li, Z.; Zhang, T.; Liu, J.; Xie, S. Assessing fish distribution and threats to fish biodiversity in the Yangtze River Basin, China. Ichthyol. Res. 2014, 61, 183–188. [Google Scholar] [CrossRef]
- Ma, R.; Yang, G.; Duan, H.; Jiang, J.; Wang, S.; Feng, X.; Li, A.; Kong, F.; Xue, B.; Wu, J.; et al. China’s lakes at present: Number, area and spatial distribution. Sci. China Earth Sci. 2011, 54, 283–289. [Google Scholar] [CrossRef]
- Yang, G.; Ma, R.; Zhang, L.; Jiang, J.; Yao, S.; Zhang, M.; Zeng, H. Lake status, major problems and protection strategy in China. J. Lake Sci. 2010, 22, 799–810. [Google Scholar]
- Jin, X.; Tu, Q. The Standard Methods for Observation and Analysis in Lake Eutrophication, 2nd ed.; Environmental Science: Beijing, China, 1990. [Google Scholar]
- SEPA. Analytical Methods for Water and Wastewater Monitor, 4th ed.; Chinese Environmental Science Press: Beijing, China, 2002. [Google Scholar]
- Pinkas, L.; Oliphant, M.S.; Iverson, I.L.K. Food habitats of Albacore, Blue-Fintuna, and Bonito in California waters. Fish. Bull. 1971, 152, 1–105. [Google Scholar]
- Selleslagh, J.; Amara, R.; Laffargue, P.; LeSourd, S.; Lepage, M.; Girardin, M. Fish composition and assemblage structure in three Eastern English Channel macrotidal estuaries: A comparison with other French estuaries. Estuar. Coast. Shelf Sci. 2009, 81, 149–159. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2020; Available online: https://www.r-project.org (accessed on 23 January 2021).
- Hijmans, R.J.; van Etten, J.; Sumner, M.; Cheng, J.; Baston, D.; Bevan, A.; Bivand, R.; Busetto, L.; Canty, M.; Fasoli, B.; et al. Raster: Geographic Data Analysis and Modeling. 2020. Available online: https://cran.r-project.org/web/packages/raster/index.html (accessed on 14 November 2020).
- Naimi, B.; Hamm, N.A.S.; Groen, T.A.; Skidmore, A.K.; Toxopeus, A.G. Where is positional uncertainty a problem for species distribution modelling? Ecography 2013, 37, 191–203. [Google Scholar] [CrossRef]
- Dodson, S.I.; Arnott, S.E.; Cottingham, K.L. The relationship in lake communities between primary productivity and species richness. Ecology 2000, 81, 2662–2679. [Google Scholar] [CrossRef]
- Amarasinghe, U.S.; Welcomme, R.L. An analysis of fish species richness in natural lakes. Environ. Boil. Fishes 2002, 65, 327–339. [Google Scholar] [CrossRef]
- Jeppesen, E.; Canfield, D.E.; Bachmann, R.W.; Søndergaard, M.; Havens, K.E.; Johansson, L.S.; Lauridsen, T.L.; Sh, T.; Rutter, R.P.; Warren, G.; et al. Toward predicting climate change effects on lakes: A comparison of 1656 shallow lakes from Florida and Denmark reveals substantial differences in nutrient dynamics, metabolism, trophic structure, and top-down control. Inland Waters 2020, 10, 197–211. [Google Scholar] [CrossRef]
- Menezes, R.F.; Borchsenius, F.; Svenning, J.-C.; Søndergaard, M.; Lauridsen, T.L.; Landkildehus, F.; Jeppesen, E. Variation in fish community structure, richness, and diversity in 56 Danish lakes with contrasting depth, size, and trophic state: Does the method matter? Hydrobiologia 2012, 710, 47–59. [Google Scholar] [CrossRef]
- Helminen, H.; Karjalainen, J.; Kurkilahti, M.; Rask, M.; Sarvala, J. Eutrophication and fish biodiversity in Finnish lakes. SIL Proc. 2000, 27, 194–199. [Google Scholar] [CrossRef]
- Bachmann, R.W.; Jones, B.L.; Fox, D.D.; Hoyer, M.; Bull, L.A.; Canfield, D.E. Relations between trophic state indicators and fish in Florida (U.S.A.) lakes. Can. J. Fish. Aquat. Sci. 1996, 53, 842–855. [Google Scholar] [CrossRef]
- Drakou, E.G.; Bobori, D.C.; Kallimanis, A.S.; Mazaris, A.D.; Sgardelis, S.P.; Pantis, J.D. Freshwater fish community structured more by dispersal limitation than by environmental heterogeneity. Ecol. Freshw. Fish 2009, 18, 369–379. [Google Scholar] [CrossRef]
- Zhao, S.; Fang, J.; Peng, C.; Tang, Z.; Piao, S. Patterns of fish species richness in China’s lakes. Glob. Ecol. Biogeogr. 2006, 15, 386–394. [Google Scholar] [CrossRef]
- Allan, J.D.; Abell, R.; Hogan, Z.; Revenga, C.; Taylor, B.W.; Welcomme, R.L.; Winemiller, K.O. Overfishing of inland waters. Bioscience 2005, 55, 1041–1051. [Google Scholar] [CrossRef] [Green Version]
- Vander Zanden, M.J.; Vadeboncoeur, Y. Fishes as integrators of benthic and pelagic food webs in lakes. Ecology 2002, 83, 2152–2161. [Google Scholar] [CrossRef]
- Persson, L.; Diehl, S.; Johansson, L.; Andersson, G.; Hamrin, S.F. Shifts in fish communities along the productivity gradient of temperate lakes-patterns and the importance of size-structured interactions. J. Fish Biol. 1991, 38, 281–293. [Google Scholar] [CrossRef]
- Quiros, R. Empirical relationships between nutrients, phyto- and zooplankton and relative fish biomass in lakes and reservoirs of Argentina. SIL Proc. 1991, 24, 1198–1206. [Google Scholar] [CrossRef]
- Hayden, B.; Myllykangas, J.-P.; Rolls, R.J.; Kahilainen, K.K. Climate and productivity shape fish and invertebrate community structure in subarctic lakes. Freshw. Biol. 2017, 62, 990–1003. [Google Scholar] [CrossRef]
- Persson, L.; Diehl, S.; Johansson, L.; Andersson, G.; Hamrin, S.F. Trophic interactions in temperate lake ecosystems: A test of food chain theory. Am. Nat. 1992, 140, 59–84. [Google Scholar] [CrossRef]
- Yang, G.; Zhong, C.; Pan, H. Comparative studies on seasonal variations of metazooplankton in waters with different eutrophic states in Lake Taihu. Environ. Monit. Assess. 2009, 150, 445–453. [Google Scholar] [CrossRef]
- Chen, F.; Dai, X.; Shu, T.; Gulati, R.D.; Liu, Z. Microcystins derived from lysing Microcystis cells do not cause negative effects on crustacean zooplankton in Lake Taihu, China. Aquat. Ecol. 2013, 47, 379–387. [Google Scholar] [CrossRef]
- Yu, J.; Liu, Z.; He, H.; Zhen, W.; Guan, B.; Chen, F.; Li, K.; Zhong, P.; Teixeira-de Mello, F.T.; Jeppesen, E. Submerged macrophytes facilitate dominance of omnivorous fish in a subtropical shallow lake: Implications for lake restoration. Hydrobiologia 2016, 775, 97–107. [Google Scholar] [CrossRef]
- Li, Y.; Ye, J.; Chen, F.; Lliu, Z.; Wang, W.; Liu, X. Feeding ecology of Hemiculter Leucisculus in the Meiliang Bay of Taihu Lake. J. Hohai Univ. 2008, 36, 82–85. (In Chinese) [Google Scholar]
- Moss, B. Climate change, nutrient pollution and the bargain of Dr Faustus. Freshw. Biol. 2010, 55, 175–187. [Google Scholar] [CrossRef]
- Rao, W.; Ning, J.; Zhong, P.; Jeppesen, E.; Liu, Z. Size-dependent feeding of omnivorous Nile tilapia in a macrophyte-dominated lake: Implications for lake management. Hydrobiologia 2015, 749, 125–134. [Google Scholar] [CrossRef]
- Gao, J.; Zhong, P.; Ning, J.; Liu, Z.; Jeppesen, E. Herbivory of omnivorous fish shapes the food web Structure of a Chinese tropical eutrophic lake: Evidence from stable isotope and fish gut content analyses. Water 2017, 9, 69. [Google Scholar] [CrossRef] [Green Version]
- Penttinen, O.-P.; Holopainen, I.J. Seasonal feeding activity and ontogenetic dietary shifts in crucian carp, Carassius carassius. Environ. Biol. Fish. 1992, 33, 215–222. [Google Scholar] [CrossRef]
- Richardson, M.J.; Whoriskey, F.G.; Roy, L.H. Turbidity generation and biological impacts of an exotic fish Carassius auratus, introduced into shallow seasonally anoxic ponds. J. Fish Biol. 1995, 47, 576–585. [Google Scholar] [CrossRef]
- Chen, K.-N.; Bao, C.-H.; Zhou, W.-P. Ecological restoration in eutrophic Lake Wuli: A large enclosure experiment. Ecol. Eng. 2009, 35, 1646–1655. [Google Scholar] [CrossRef]
- Li, B.; Wang, Z.; Jin, L.; Ye, F.; Chen, D.; Zhang, Y. Study on diet shift of Erythroulter Mongolicus with stable isotope technology. J. Fish. Chin. 2011, 35, 1419–1425. (In Chinese) [Google Scholar]
- Zhou, D.; Ye, J.; Wang, W.; Liu, Z. Feeding habits and the relation to the size of Cultrichthys Erythropterus in Meiliang Bay, Lake Taihu: Implication for biomanipulation and management. J. Lake Sci. 2011, 23, 796–800. (In Chinese) [Google Scholar]
- Hossain, M.; Arhonditsis, G.B.; Hoyle, J.A.; Randall, R.G.; Koops, M.A. Nutrient management and structural shifts in fish assemblages: Lessons learned from an Area of Concern in Lake Ontario. Freshw. Biol. 2019, 64, 967–983. [Google Scholar] [CrossRef]
- Diehl, S. Foraging Efficiency of Three Freshwater Fishes: Effects of Structural Complexity and Light. Oikos 1988, 53, 207–214. [Google Scholar] [CrossRef]
- Jönsson, M.; Ranåker, L.; Nilsson, P.A.; Brönmark, C. Foraging efficiency and prey selectivity in a visual predator: Differential effects of turbid and humic water. Can. J. Fish. Aquat. Sci. 2013, 70, 1685–1690. [Google Scholar] [CrossRef]
- Liang, X. On the feeding behavior of Mandarin Fish (Siniperca Chuatsi). Oceanol. Limnol. Sin. 1995, 26, 119–125. (In Chinese) [Google Scholar]
- Zhou, X.; Hu, Z.; Liu, Q.; Yang, L.; Wang, Y. Feeding ecology of the non-indigenous fish Hypomesus nipponensis in Lake Ulungur, China: Insight into the relationship between its introduction and the collapse of the native Eurasian perch population. Mar. Freshw. Res. 2013, 64, 549–557. [Google Scholar] [CrossRef]
- Lauridsen, T.L.; Landkildehus, F.; Jeppesen, E.; Jørgensen, T.B.; Søndergaard, M. A comparison of methods for calculating Catch Per Unit Effort (CPUE) of gill net catches in lakes. Fish. Res. 2008, 93, 204–211. [Google Scholar] [CrossRef]
- Mehner, T.; Emmrich, M.; Hartwig, S. Spatial predictors of fish species composition in European lowland lakes. Ecography 2014, 37, 73–79. [Google Scholar] [CrossRef]
- Arranz, I.; Mehner, T.; Benejam, L.; Argillier, C.; Holmgren, K.; Jeppesen, E.; Lauridsen, T.L.; Volta, P.; Winfield, I.J.; Brucet, S. Density-dependent effects as key drivers of intraspecific size structure of six abundant fish species in lakes across Europe. Can. J. Fish. Aquat. Sci. 2016, 73, 519–534. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Chang, J.; Shu, G. Applications of factor-criteria system reconstruction analysis in the reproduction research on Grass Carp, Black Carp, Silver Carp and Bighead in the Yangtze River. Int. J. Gen. Syst. 2000, 29, 419–428. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Minimum | Maximum | Mean | ±SD |
|---|---|---|---|---|
| Lake area (km2) | 7.3 | 375 | 94 | 102.6 |
| Secchi depth (m) | 0.1 | 2 | 0.8 | 0.5 |
| Depth (m) | 1.5 | 7.1 | 4.2 | 1.6 |
| Secchi depth: water depth ratio | 0.02 | 0.4 | 0.2 | 0.09 |
| Total suspended solids (mg L−1) | 3.2 | 59 | 16.5 | 10.5 |
| Total nitrogen (mg L−1) | 0.5 | 2.7 | 1.2 | 0.6 |
| Total phosphorus (µg L−1) | 17.6 | 393 | 89 | 87.2 |
| Chla (µg L−1) | 4.4 | 84 | 32 | 21.7 |
| Fish Compositional Parameters | Environmental Variables | AIC | Estimate | t-Value | Pr (>|t|) |
|---|---|---|---|---|---|
| Log Fish species richness | Log Chla | 160.5 | 2.7 | 2.6 | 0.012 |
| Log NPUE | Log Chla | 43.2 | 0.8 | 4.2 | 0.00017 |
| Log BPUE | Log Chla | 40.7 | 0.9 | 4.5 | <0.0001 |
| Log Mean fish size | Log TN | 39.6 | 1.0 | 2.6 | 0.014 |
| Log Lake size | 42.5 | −0.3 | −2.5 | 0.019 | |
| Log NPUE of fish TL < 15 cm | Log Chla | 60.0 | 0.9 | 3.6 | 0.0012 |
| Log NPUE of fish TL > 15 cm | Log Chla | 48.0 | 0.7 | 3.2 | 0.0028 |
| Log N% of zooplanktivores | Log Chla | −64.9 | 0.1 | 3.0 | 0.006 |
| Log SD | −64.0 | −0.1 | −2.2 | 0.04 | |
| Log B% of zooplanktivores | Log SD | 25.0 | −0.6 | −2.5 | 0.02 |
| Log Chla | 27.5 | 0.4 | 2.3 | 0.033 | |
| Log B% of benthivores | Log TN | 33.2 | −0.9 | −2.3 | 0.028 |
| Log N% of piscivores | Log Chla | 14.2 | −0.5 | −2.6 | 0.019 |
| Log B% of piscivores | Log Chla | 24.1 | −1.4 | −6.5 | <0.0001 |
| Log TN | 29.7 | −2.8 | −5.4 | <0.0001 | |
| Log TP | 30.5 | 1.2 | 4.3 | 0.00063 | |
| Log SD | 31.9 | −1.0 | −3.8 | 0.0019 | |
| Log NPUE of Sijiao | Log Chla | 53.4 | 0.8 | 2.7 | 0.012 |
| Log BPUE of Sijiao | Log Chla | 53.5 | 0.8 | 2.6 | 0.014 |
| Log NPUE of sharpbelly | Log Chla | 45.6 | 1.3 | 3.5 | 0.0022 |
| Log BPUE of sharpbelly | Log Chla | 38.6 | 1.1 | 3.2 | 0.0041 |
| Log NPUE of crucian carp | Log Chla | 25.4 | 1.3 | 4.2 | 0.00048 |
| Log SD | 32.9 | 0.9 | 2.7 | 0.016 | |
| Log BPUE of crucian carp | Log Chla | 29.3 | 0.9 | 2.8 | 0.011 |
| Log NPUE of Culter | Log Chla | 2.5 | 0.5 | 3.7 | 0.002 |
| Log Lake size | 12.0 | −0.2 | −2.2 | 0.042 | |
| Log BPUE of Culter | Log Lake size | 18.8 | −0.5 | −3.0 | 0.008 |
| Log Chla | 22.1 | 0.5 | 2.2 | 0.04 | |
| Log Mean size of Sijiao | Log TN | −16.5 | 0.6 | 3.8 | 0.00059 |
| Log Mean size of sharpbelly | Log SD | −2.9 | −0.8 | −4.4 | 0.00016 |
| Log Mean size of crucian carp | Log Lake size | 5.9 | −0.2 | −2.2 | 0.042 |
| Log mean size of Culter | Log Chla | 13.2 | −0.4 | −2.1 | 0.05 |
| Trophic Group | Species | FO% | N% | B% | IRI | IRI% |
|---|---|---|---|---|---|---|
| Zooplanktivores | Toxabramis swinhonisa | 89 | 62 | 14 | 6719 | 52 |
| Coilia nasusb | 42 | 3 | 1 | 187 | 1.5 | |
| Hyporhamphus intermediusc | 28 | 0.5 | 0.1 | 17 | 0.1 | |
| Neosalanx taihuensisd | 6 | 0.05 | 0.01 | 0.3 | 0.0 | |
| Omniplanktivores | Hemiculter leucisculusa | 78 | 21 | 6 | 2164 | 17 |
| Hypophthalmichthys nobilisa | 28 | 1 | 36 | 1027 | 8 | |
| Hypophthalmichthys molitrixa | 11 | 0.6 | 5 | 57 | 0.4 | |
| Pseudobrama simonia | 11 | 0.8 | 1 | 21 | 0.2 | |
| Acheilognathus macropterusa | 17 | 0.3 | 0.2 | 9 | 0.07 | |
| Pseudorasbora parvaa | 6 | 0.3 | 0.05 | 2 | 0.01 | |
| Omnibenthivores | Carassius carassiusa | 69 | 7 | 26 | 2253 | 18 |
| Tachysurus fulvidracoe | 28 | 0.7 | 2 | 75 | 0.6 | |
| Cyprinus carpioa | 6 | 0.05 | 1 | 7 | 0.06 | |
| Xenocypris microlepisa | 8 | 0.1 | 0.6 | 6 | 0.05 | |
| Xenocypris hupeinensisa | 6 | 0.04 | 0.06 | 0.5 | 0.0 | |
| Xenocypris davidia | 3 | 0.02 | 0.05 | 0.2 | 0.0 | |
| Hemibarbus labeoa | 3 | 0.02 | 0.08 | 0.3 | 0.0 | |
| Sarcocheilichthys sinensisa | 3 | 0.04 | 0.1 | 0.4 | 0.0 | |
| Omniherbivore | Megalobrama amblycephalaa | 11 | 0.9 | 4 | 54 | 0.4 |
| Piscivores | Culter spp. a | 56 | 1.2 | 3 | 234 | 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, J.; Zhen, W.; Kong, L.; He, H.; Zhang, Y.; Yang, X.; Chen, F.; Zhang, M.; Liu, Z.; Jeppesen, E. Changes in Pelagic Fish Community Composition, Abundance, and Biomass along a Productivity Gradient in Subtropical Lakes. Water 2021, 13, 858. https://doi.org/10.3390/w13060858
Yu J, Zhen W, Kong L, He H, Zhang Y, Yang X, Chen F, Zhang M, Liu Z, Jeppesen E. Changes in Pelagic Fish Community Composition, Abundance, and Biomass along a Productivity Gradient in Subtropical Lakes. Water. 2021; 13(6):858. https://doi.org/10.3390/w13060858
Chicago/Turabian StyleYu, Jinlei, Wei Zhen, Lingyang Kong, Hu He, Yongdong Zhang, Xiangdong Yang, Feizhou Chen, Min Zhang, Zhengwen Liu, and Erik Jeppesen. 2021. "Changes in Pelagic Fish Community Composition, Abundance, and Biomass along a Productivity Gradient in Subtropical Lakes" Water 13, no. 6: 858. https://doi.org/10.3390/w13060858