
Unravelling Microbial Communities Associated with Different Light Non-Aqueous Phase Liquid Types Undergoing Natural Source Zone Depletion Processes at a Legacy Petroleum Site

Abstract
:
1. Introduction
2. Materials and Methods
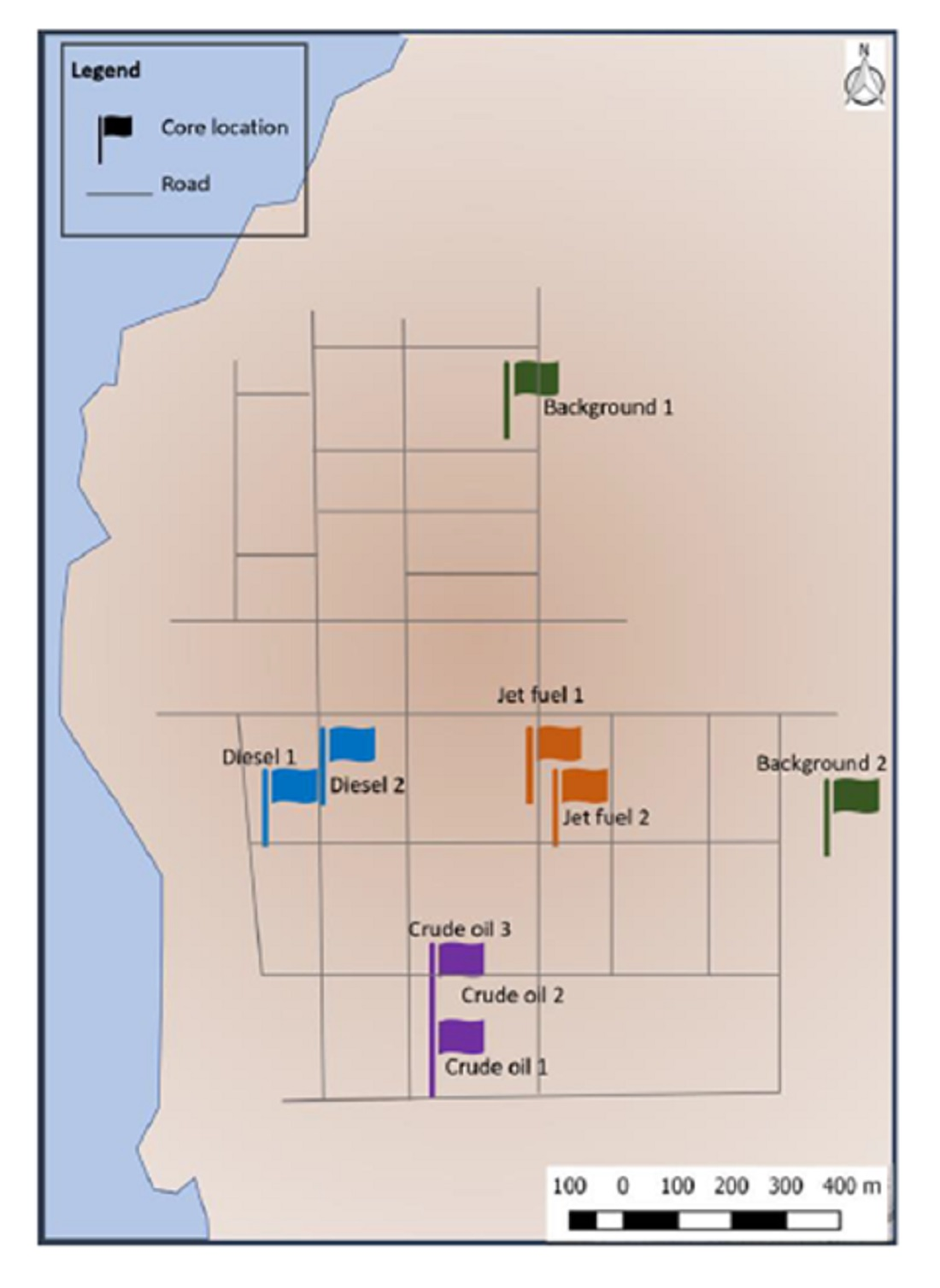
2.1. Site Description and Soil Coring
2.2. Chemical Analysis
2.2.1. Extraction of Soil Core Samples
2.2.2. GC–FID and GC–MS Analysis of Soil Core Samples
2.2.3. LNAPL Sampling
2.2.4. GC–FID and GC–MS Analysis of LNAPL Samples
2.3. Microbial Community Analysis
3. Results
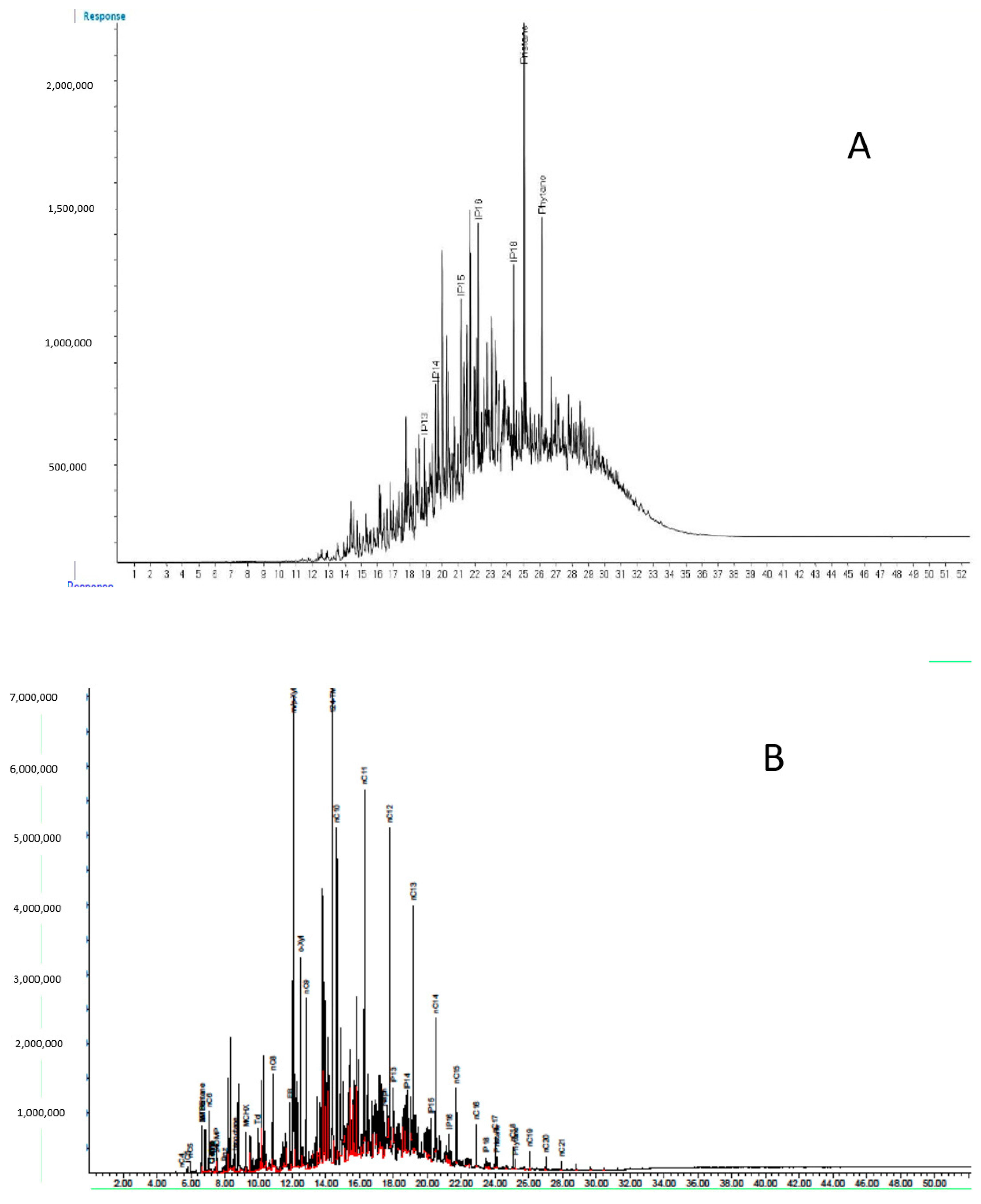
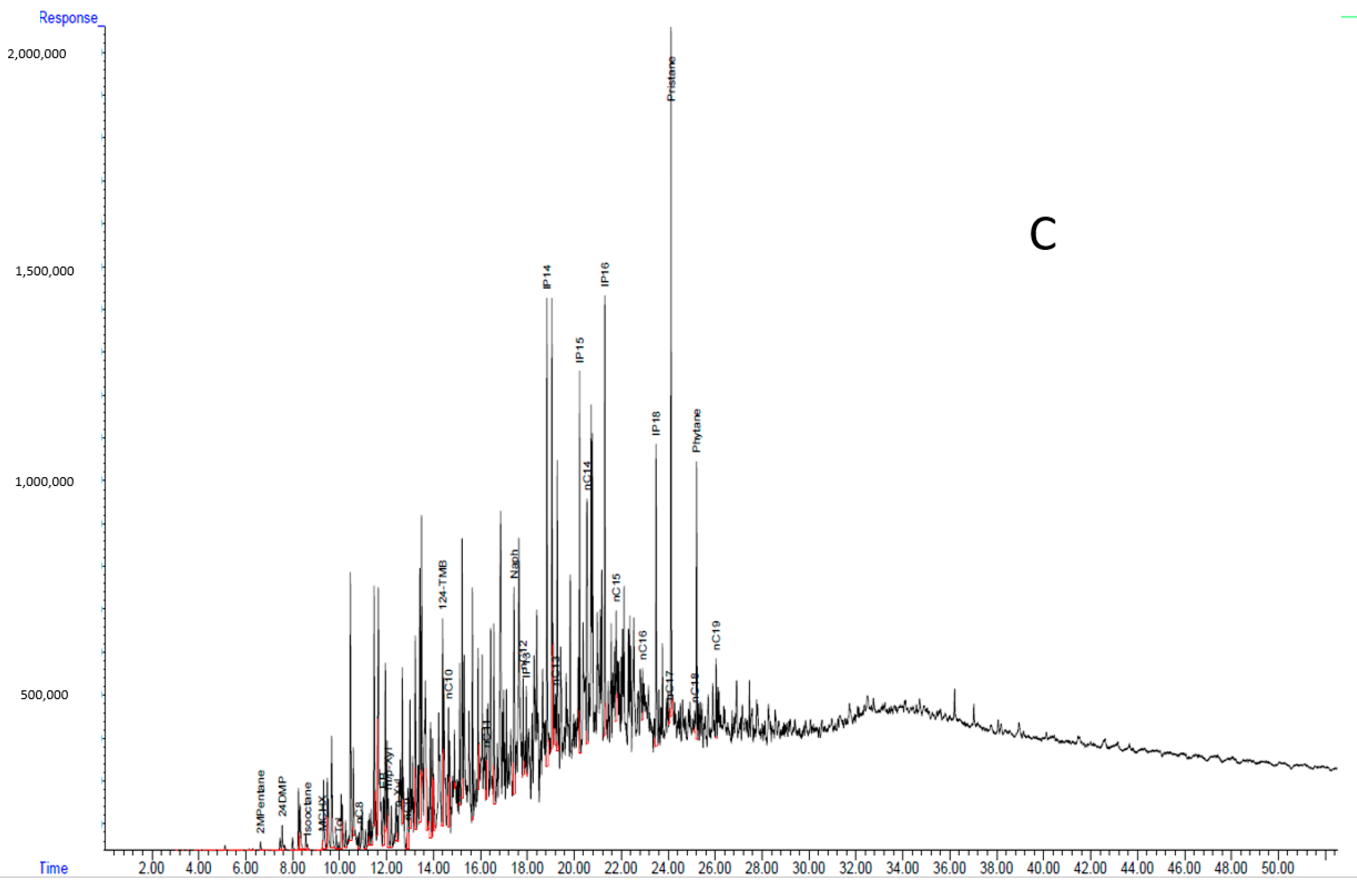
3.1. Chemical Analysis
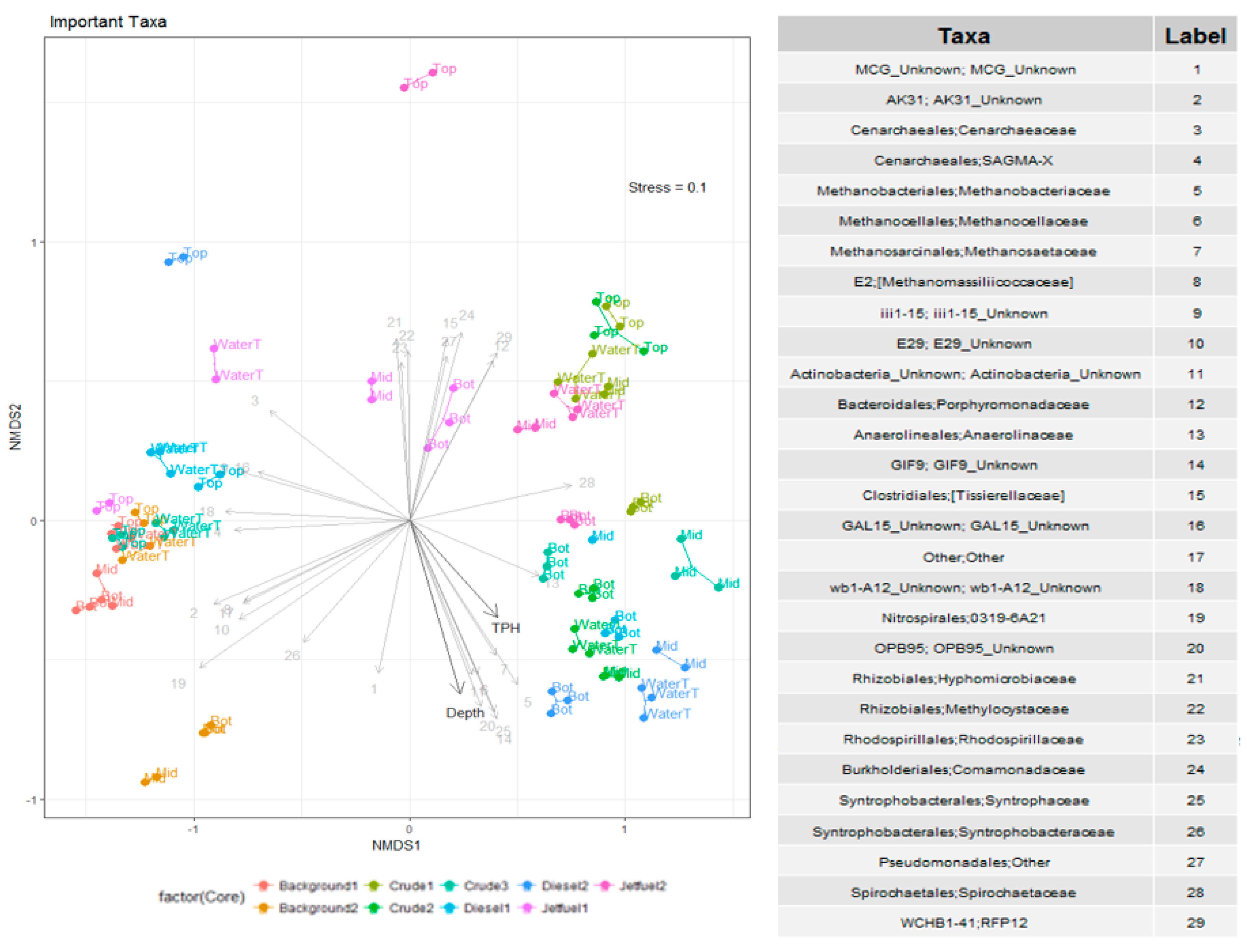
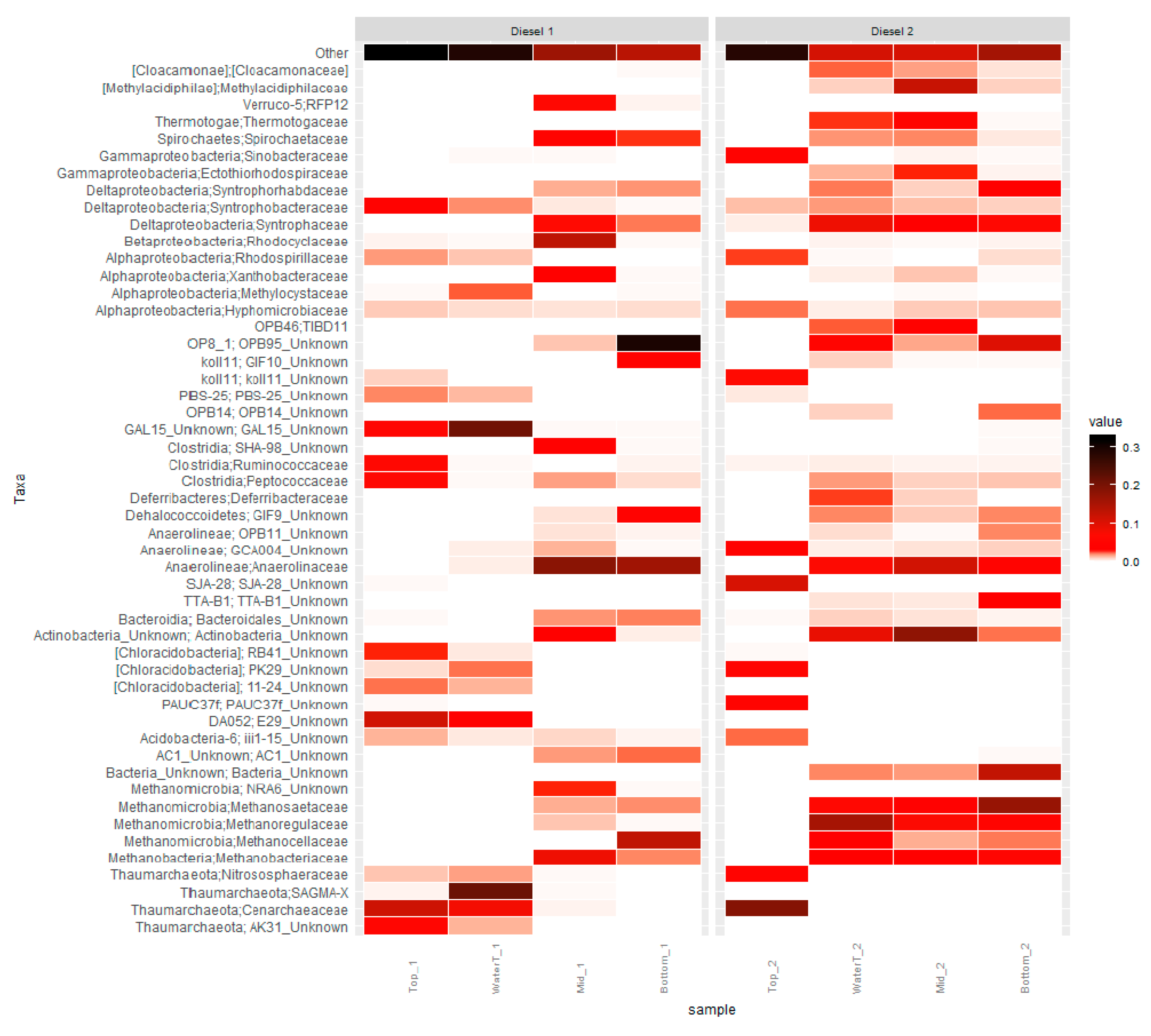
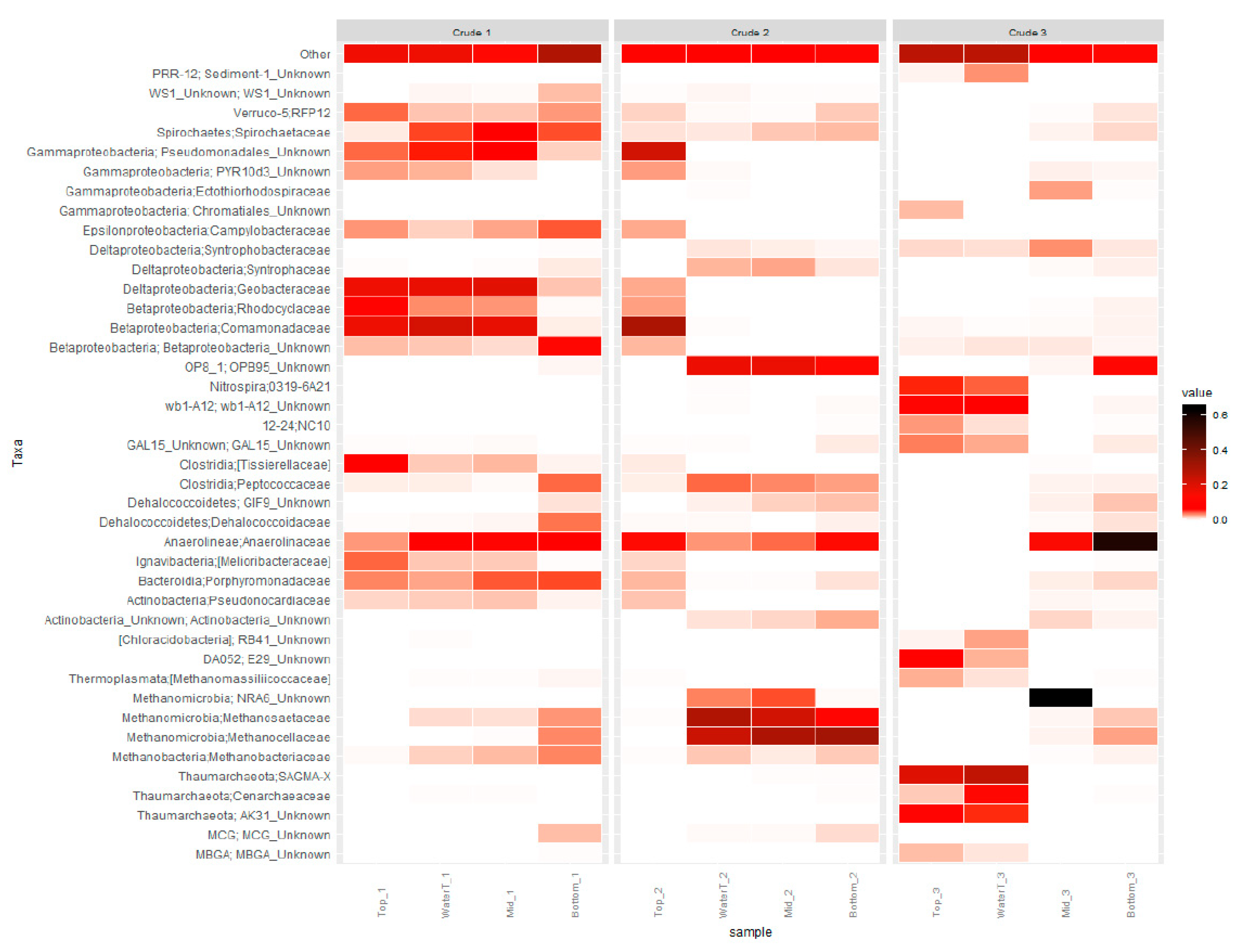
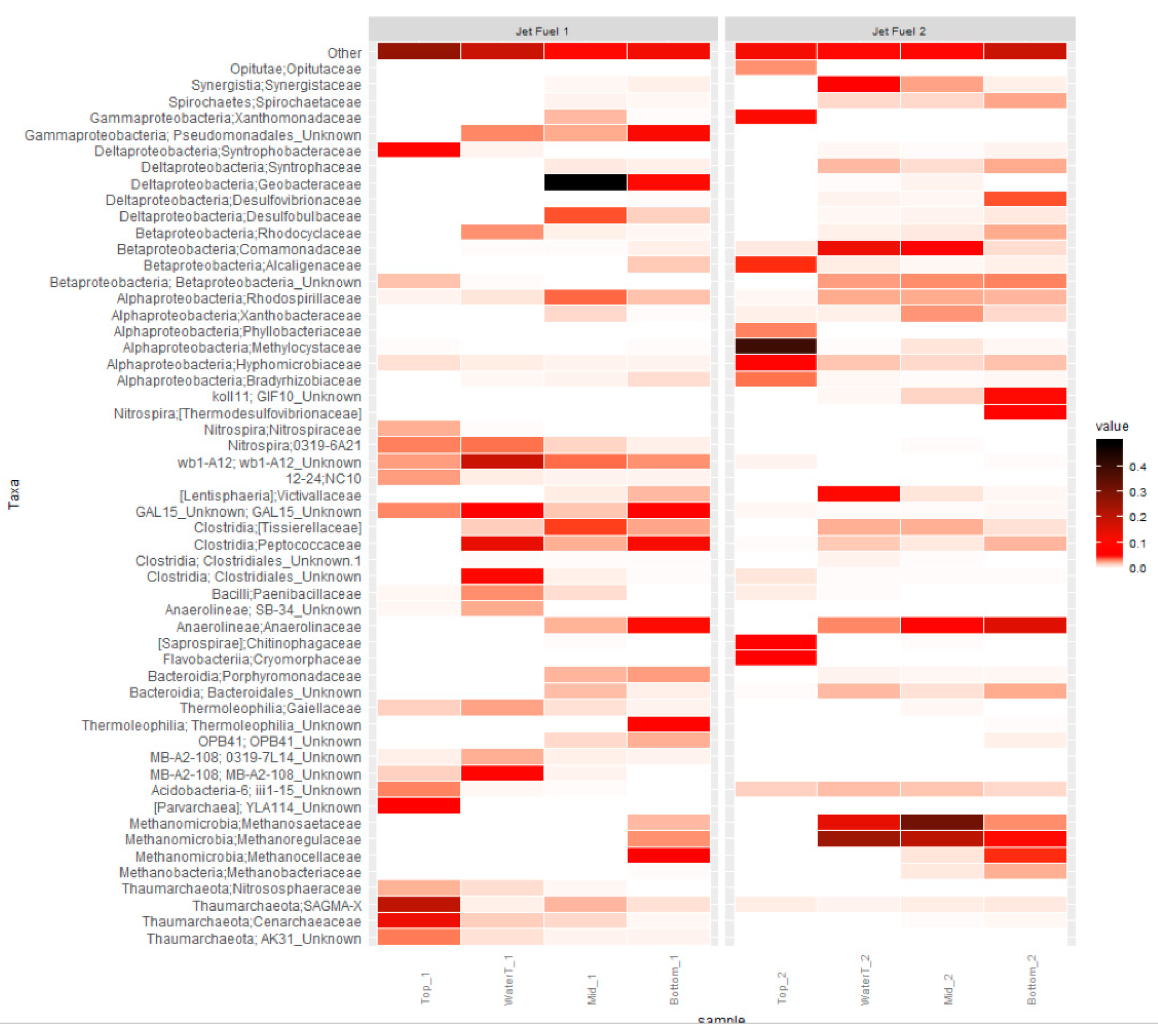
3.2. Microbial Community Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sutton, N.B.; Maphosa, F.; Morillo, J.A.; Abu Al-Soud, W.; Langenhoff, A.A.M.; Grotenhuis, T.; Rijnaarts, H.H.M.; Smidt, H. Impact of Long-Term Diesel Contamination on Soil Microbial Community Structure. Appl. Environ. Microbiol. 2012, 79, 619–630. [Google Scholar] [CrossRef]
- Aubé, J.; Senin, P.; Pringault, O.; Bonin, P.; Deflandre, B.; Bouchez, O.; Bru, N.; Biritxinaga-Etchart, E.; Klopp, C.; Guyoneaud, R.; et al. The impact of long-term hydrocarbon exposure on the structure, activity, and biogeochemical functioning of microbial mats. Mar. Pollut. Bull. 2016, 111, 115–125. [Google Scholar] [CrossRef]
- Baek, K.H.; Kim, H.S.; Oh, H.M.; Yoon, B.D.; Kim, J.; Lee, I.S. Effects of crude oil, oil components, and bioremediation on plant growth. J. Environ. Sci. Health Part A 2004, 39, 2465–2472. [Google Scholar] [CrossRef]
- Maletić, S.P.; Dalmacija, B.D.; Rončević, S.D.; Agbaba, J.R.; Perović, S.D.U. Impact of hydrocarbon type, concentration and weathering on its biodegradability in soil. J. Environ. Sci. Health Part A 2011, 46, 1042–1049. [Google Scholar] [CrossRef]
- Atlas, R.M. Microbial degradation of petroleum hydrocarbons: An environmental perspective. Microbiol. Rev. 1981, 45, 180–209. [Google Scholar] [CrossRef]
- Greene, E.A.; Kay, J.G.; Jaber, K.; Stehmeier, L.G.; Voordouw, G. Composition of Soil Microbial Communities Enriched on a Mixture of Aromatic Hydrocarbons. Appl. Environ. Microbiol. 2000, 66, 5282–5289. [Google Scholar] [CrossRef] [PubMed]
- Bruckberger, M.C.; Bastow, T.P.; Morgan, M.J.; Gleeson, D.; Banning, N.; Davis, G.; Puzon, G.J. Biodegradability of polar compounds formed from weathered diesel. Biodegradation 2018, 29, 443–461. [Google Scholar] [CrossRef] [PubMed]
- Bruckberger, M.C.; Morgan, M.J.; Walsh, T.; Bastow, T.P.; Prommer, H.; Mukhopadhyay, A.; Kaksonen, A.H.; Davis, G.; Puzon, G.J. Biodegradability of legacy crude oil contamination in Gulf War damaged groundwater wells in Northern Kuwait. Biodegradation 2019, 30, 71–85. [Google Scholar] [CrossRef] [PubMed]
- Atlas, R.M.; Bartha, R. Microbial Ecology: Fundamentals and Applications; Addison-Wesley Publishing Company: Boston, MA, USA, 1981. [Google Scholar]
- Fahy, A.; McGenity, T.J.; Timmis, K.N.; Ball, A.S. Heterogeneous aerobic benzene-degrading communities in oxygen-depleted groundwaters. FEMS Microbiol. Ecol. 2006, 58, 260–270. [Google Scholar] [CrossRef] [PubMed]
- Davis, G.; Barber, C.; Power, T.; Thierrin, J.; Patterson, B.; Rayner, J.; Wu, Q. The variability and intrinsic remediation of a BTEX plume in anaerobic sulphate-rich groundwater. J. Contam. Hydrol. 1999, 36, 265–290. [Google Scholar] [CrossRef]
- Ghattas, A.-K.; Fischer, F.; Wick, A.; Ternes, T.A. Anaerobic biodegradation of (emerging) organic contaminants in the aquatic environment. Water Res. 2017, 116, 268–295. [Google Scholar] [CrossRef]
- Lundegard, P.D.; Knott, J.R. Polar organics in crude oil and their potential impacts on water quality. In Proceedings of the Petroleum Hydrocarbons and Organic Chemicals in the Ground Water NGWA/API Conference, Houston, TX, USA, 14–16 November 2001. [Google Scholar]
- Lundegard, P.D.; Sweeney, R.E. Total Petroleum Hydrocarbons in Groundwater—Evaluation of Nondissolved and Nonhydrocarbon Fractions. Environ. Forensics 2004, 5, 85–95. [Google Scholar] [CrossRef]
- Lang, D.A.; Bastow, T.P.; Van Aarssen, B.G.; Warton, B.; Davis, G.B.; Johnston, C.D. Polar Compounds from the Dissolution of Weathered Diesel. Ground Water Monit. Remediat. 2009, 29, 85–93. [Google Scholar] [CrossRef]
- Lekmine, G.; Bastow, T.P.; Johnston, C.D.; Davis, G.B. Dissolution of multi-component LNAPL gasolines: The effects of weathering and composition. J. Contam. Hydrol. 2014, 160, 1–11. [Google Scholar] [CrossRef]
- Johnston, C.D.; Bastow, T.P.; Innes, N.L. The use of biodegradation signatures and biomarkers to differentiate spills of petroleum hydrocarbon liquids in the subsurface and estimate natural mass loss. Eur. J. Soil Biol. 2007, 43, 328–334. [Google Scholar] [CrossRef]
- Garg, S.; Newell, C.J.; Kulkarni, P.R.; King, D.C.; Adamson, D.T.; Renno, M.I.; Sale, T. Overview of Natural Source Zone Depletion: Processes, Controlling Factors, and Composition Change. Ground Water Monit. Remediat. 2017, 37, 62–81. [Google Scholar] [CrossRef]
- Lari, K.S.; Davis, G.B.; Rayner, J.L.; Bastow, T.P.; Puzon, G.J. Natural source zone depletion of LNAPL: A critical review supporting modelling approaches. Water Res. 2019, 157, 630–646. [Google Scholar] [CrossRef] [PubMed]
- Lari, K.S.; Rayner, J.L.; Davis, G.B. Towards characterizing LNAPL remediation endpoints. J. Environ. Manag. 2018, 224, 97–105. [Google Scholar] [CrossRef]
- Johnston, C.; Rayner, J.; Patterson, B.; Davis, G. Volatilisation and biodegradation during air sparging of dissolved BTEX-contaminated groundwater. J. Contam. Hydrol. 1998, 33, 377–404. [Google Scholar] [CrossRef]
- Johnston, C.; Rayner, J.; Briegel, D. Effectiveness of in situ air sparging for removing NAPL gasoline from a sandy aquifer near Perth, Western Australia. J. Contam. Hydrol. 2002, 59, 87–111. [Google Scholar] [CrossRef]
- Johnston, C.; Fisher, S.; Rayner, J. Removal of Petroleum Hydrocarbons from the Vadose Zone during Multi-Phase Extraction at a Contaminated Industrial Site; IAHS PUBLICATION: Philadelphia, PA, USA, 2002; pp. 287–294. [Google Scholar]
- Johnston, C.; Robertson, B.; Bastow, T. Evidence for the success of biosparging LNAPL diesel in the water table zone of a shallow sand aquifer. In GQ07: Securing Groundwater Quality in Urban and Industrial Environments, Proceedings of the 6th International Groundwater Quality Conference, Fremantle, Australia, 2–7 December 2007; International Association of Hydrological Sciences: Wallingford, UK, 2007. [Google Scholar]
- Davis, G.B.; Rayner, J.L.; Trefry, M.G.; Fisher, S.J.; Patterson, B.M. Measurement and Modeling of Temporal Variations in Hydrocarbon Vapor Behavior in a Layered Soil Profile. Vadose Zone J. 2005, 4, 225–239. [Google Scholar] [CrossRef]
- Franzmann, P.; Patterson, B.; Power, T.; Nichols, P.; Davis, G. Microbial biomass in a shallow, urban aquifer contaminated with aromatic hydrocarbons: Analysis by phospholipid fatty acid content and composition. J. Appl. Bacteriol. 1996, 80, 617–625. [Google Scholar] [CrossRef]
- Franzmann, P.D.; Zappia, L.R.; Power, T.R.; Davis, G.B.; Patterson, B.M. Microbial mineralisation of benzene and characterisation of microbial biomass in soil above hydro-carbon-contaminated groundwater. FEMS Microbiol. Ecol. 1999, 30, 67–76. [Google Scholar] [CrossRef]
- Ko, D.; Yoo, G.; Yun, S.-T.; Jun, S.-C.; Chung, H. Bacterial and fungal community composition across the soil depth profiles in a fallow field. J. Ecol. Environ. 2017, 41, 34. [Google Scholar] [CrossRef]
- Fierer, N.; Schimel, J.P.; A Holden, P. Variations in microbial community composition through two soil depth profiles. Soil Biol. Biochem. 2003, 35, 167–176. [Google Scholar] [CrossRef]
- Ding, A.; Sun, Y.; Dou, J.; Cheng, L.; Jiang, L.; Zhang, D.; Zhao, X. Characterizing Microbial Activity and Diversity of Hydrocarbon-Contaminated Sites. Hydrocarbon 2013, 18. [Google Scholar] [CrossRef]
- Zhou, J.; Xia, B.; Treves, D.S.; Wu, L.-Y.; Marsh, T.L.; O’Neill, R.V.; Palumbo, A.V.; Tiedje, J.M. Spatial and Resource Factors Influencing High Microbial Diversity in Soil. Appl. Environ. Microbiol. 2002, 68, 326–334. [Google Scholar] [CrossRef]
- Das, N.; Chandran, P. Microbial Degradation of Petroleum Hydrocarbon Contaminants: An Overview. Biotechnol. Res. Int. 2011, 2011, 941810. [Google Scholar] [CrossRef]
- Playford, P.E.; Low, G.; Cockbain, A.E. Geology of the Perth Basin, Western Australia; Geological Survey of Western Australia: Perth, Australia, 1976. [Google Scholar]
- McArthur, W.M.; Bettenay, E. The Development and Distribution of the Soils of the Swan Coastal Plain, Western Australia; Soil Publication; CSIRO: Melbourne, Australia, 1960; p. 16. [Google Scholar]
- Playford, P.E.; Low, G.H. Definitions of some new and revised rock units in the Perth Basin. West. Aust. Geol. Surv. Annu. Rep. 1971, 44–46. [Google Scholar]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME Allows Analysis of High-Throughput Community Sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Comeau, A.M.; Douglas, G.M.; Langille, M.G.I. Microbiome Helper: A Custom and Streamlined Workflow for Microbiome Research. mSystems 2017, 2, e00127-16. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Kobert, K.; Flouri, T.; Stamatakis, A. PEAR: A fast and accurate Illumina Paired-End reAd mergeR. Bioinformatics 2014, 30, 614–620. [Google Scholar] [CrossRef] [PubMed]
- Bushnell, B. BBMap: A Fast, Accurate, Splice-Aware Aligner; Lawrence Berkeley National Lab (LBNL): Berkeley, CA, USA, 2014. [Google Scholar]
- Gordon, A.; Hannon, G. Fastx-Toolkit. FASTQ/A Short-Reads Pre-Processing Tools. 2010. Available online: http://hannonlab.cshl.edu/fastx_toolkit (accessed on 20 January 2021).
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed]
- Cole, J.R.; Wang, Q.; Fish, J.A.; Chai, B.; McGarrell, D.M.; Sun, Y.; Brown, C.T.; Porras-Alfaro, A.; Kuske, C.R.; Tiedje, J.M. Ribosomal Database Project: Data and tools for high throughput rRNA analysis. Nucleic Acids Res. 2014, 42, D633–D642. [Google Scholar] [CrossRef]
- Bruckberger, M.C.; Morgan, M.J.; Bastow, T.P.; Walsh, T.; Prommer, H.; Mukhopadhyay, A.; Kaksonen, A.H.; Davis, G.B.; Puzon, G.J. Investigation into the microbial communities and associated crude oil-contamination along a Gulf War impacted groundwater system in Kuwait. Water Res. 2020, 170, 115314. [Google Scholar] [CrossRef] [PubMed]
- Team R Core. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Oksanen, J. Vegan: Community Ecology Package. R Package Version 2.4–2. 2017. Available online: https://mran.microsoft.com/snapshot/2017-02-04/web/packages/vegan/vegan.pdf (accessed on 19 January 2021).
- Taylor, M. sinkr: A Collection of Functions Featured on the Blog ‘me nugget’. R Package Version 1.0. 2014. Available online: https://github.com/menugget/sinkr (accessed on 20 January 2021).
- Franzmann, P.; Robertson, W.; Zappia, L.; Davis, G. The role of microbial populations in the containment of aromatichydrocarbons in the subsurface. Biodegradation 2002, 13, 65–78. [Google Scholar] [CrossRef]
- Lee, H.J.; Jeong, S.E.; Kim, P.J.; Madsen, E.L.; Jeon, C.O. High resolution depth distribution of Bacteria, Archaea, methanotrophs, and methanogens in the bulk and rhi-zosphere soils of a flooded rice paddy. Front. Microbiol. 2015, 6, 639. [Google Scholar] [CrossRef]
- Sharp, C.E.; Smirnova, A.V.; Graham, J.M.; Stott, M.B.; Khadka, R.; Moore, T.R.; Grasby, S.E.; Strack, M.; Dunfield, P.F. Distribution and diversity of V errucomicrobia methanotrophs in geothermal and acidic environments. Environ. Microbiol. 2014, 16, 1867–1878. [Google Scholar] [CrossRef]
- Campeão, M.E.; Reis, L.; Leomil, L.; De Oliveira, L.; Otsuki, K.; Gardinali, P.; Pelz, O.; Valle, R.; Thompson, F.L.; Thompson, C.C. The Deep-Sea Microbial Community from the Amazonian Basin Associated with Oil Degradation. Front. Microbiol. 2017, 8, 1019. [Google Scholar] [CrossRef]
- Zhang, T.; Ke, S.Z.; Liu, Y.; Fang, H. Microbial characteristics of a methanogenic phenol-degrading sludge. Water Sci. Technol. 2005, 52, 73–78. [Google Scholar] [CrossRef]
- Toth, C.R.A.; Gieg, L.M. Time Course-Dependent Methanogenic Crude Oil Biodegradation: Dynamics of Fumarate Addition Metabolites, Biodegradative Genes, and Microbial Community Composition. Front. Microbiol. 2017, 8, 2610. [Google Scholar] [CrossRef] [PubMed]
- Perminova, I.V.; Grechishcheva, N.Y.; Kovalevskii, D.V.; Kudryavtsev, A.V.; Petrosyan, V.S.; Matorin, D.N. Quantification and prediction of the detoxifying properties of humic substances related to their chemical binding to polycyclic aromatic hydrocarbons. Environ. Sci. Technol. 2001, 35, 3841–3848. [Google Scholar] [CrossRef]
- Qiu, Y.L.; Hanada, S.; Ohashi, A.; Harada, H.; Kamagata, Y.; Sekiguchi, Y. Syntrophorhabdus aromaticivorans gen. nov., sp. nov., the first cultured anaerobe capable of degrading phe-nol to acetate in obligate syntrophic associations with a hydrogenotrophic methanogen. Appl. Environ. Microbiol. 2008, 74, 2051–2058. [Google Scholar] [CrossRef] [PubMed]
- Kuever, J. The Family Syntrophorhabdaceae. In The Prokaryotes; Springer: Berlin/Heidelberg, Germany, 2014; pp. 301–303. [Google Scholar]
- Kleinsteuber, S.; Schleinitz, K.M.; Breitfeld, J.; Harms, H.; Richnow, H.H.; Vogt, C. Molecular characterization of bacterial communities mineralizing benzene under sulfate-reducing conditions. FEMS Microbiol. Ecol. 2008, 66, 143–157. [Google Scholar] [CrossRef] [PubMed]
- Kunapuli, U.; Lueders, T.; Meckenstock, R.U. The use of stable isotope probing to identify key iron-reducing microorganisms involved in anaerobic benzene degradation. ISME J. 2007, 1, 643. [Google Scholar] [CrossRef]
- Herrmann, S.; Kleinsteuber, S.; Chatzinotas, A.; Kuppardt, S.; Lueders, T.; Richnow, H.-H.; Vogt, C. Functional characterization of an anaerobic benzene-degrading enrichment culture by DNA stable isotope probing. Environ. Microbiol. 2010, 12, 401–411. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, I.R.; Galperin, Y.; Alimi, H.; Lee, R.-P.; Lu, S.-T. Patterns of Chemical Changes During Environmental Alteration of Hydrocarbon Fuels. Ground Water Monit. Remediat. 1996, 16, 113–124. [Google Scholar] [CrossRef]
- Wentzel, A.; Ellingsen, T.E.; Kotlar, H.-K.; Zotchev, S.B.; Throne-Holst, M. Bacterial metabolism of long-chain n-alkanes. Appl. Microbiol. Biotechnol. 2007, 76, 1209–1221. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Section * | Core 1 | Core 2 | Core 3 | ||||
|---|---|---|---|---|---|---|---|
| Depth [m] | TPH [g/kg] | Depth [m] | TPH [g/kg] | Depth [m] | TPH [g/kg] | ||
| Background | T | 2.00–2.05 | 0.0 | 2.00–2.05 | 0.0 | ||
| WT | 2.65–2.70 | 0.0 | 2.60–2.65 | 0.0 | |||
| M | 3.35–3.40 | 0.0 | 3.90–3.95 | 0.0 | |||
| B | 4.00–4.05 | 0.0 | 5.00–5.05 | 0.0 | |||
| Jet fuel | T | 1.50–1.55 | 0.0 | 2.60–2.65 | 0.0 | ||
| WT | 2.20–2.25 | 0.9 | 3.35–3.40 | 16.4 | |||
| M | 3.35–3.40 | 25.0 | 3.80–3.85 | 22 | |||
| B | 4.00–4.05 | 4.9 | 4.15–4.20 | 1.3 | |||
| Diesel | T | 3.00–3.05 | 0.0 | 2.95–3.00 | 0.0 | ||
| WT | 3.75–3.80 | 0.0 | 3.85–3.90 | 50.9 | |||
| M | 4.15–4.2. | 92.7 | 3.90–3.95 | 155.7 | |||
| B | 5.05–5.10 | 0.0 | 4.60–4.67 | 9.2 | |||
| Crude oil | T | 2.65–2.70 | 13.2 | 3.00–3.05 | 0.0 | 0.0 | 2.95–3.00 |
| WT | 3.10–3.15 | 25.3 | 3.70–3.75 | 21.1 | 0.0 | 3.10–3.15 | |
| M | 3.30–3.35 | 45.6 | 3.80–3.85 | 56.4 | 55.9 | 3.35–3.40 | |
| B | 3.90–3.95 | 1.6 | 4.05–4.10 | 2.3 | 0.0 | 3.90–3.95 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bruckberger, M.C.; Gleeson, D.B.; Bastow, T.P.; Morgan, M.J.; Walsh, T.; Rayner, J.L.; Davis, G.B.; Puzon, G.J. Unravelling Microbial Communities Associated with Different Light Non-Aqueous Phase Liquid Types Undergoing Natural Source Zone Depletion Processes at a Legacy Petroleum Site. Water 2021, 13, 898. https://doi.org/10.3390/w13070898
Bruckberger MC, Gleeson DB, Bastow TP, Morgan MJ, Walsh T, Rayner JL, Davis GB, Puzon GJ. Unravelling Microbial Communities Associated with Different Light Non-Aqueous Phase Liquid Types Undergoing Natural Source Zone Depletion Processes at a Legacy Petroleum Site. Water. 2021; 13(7):898. https://doi.org/10.3390/w13070898
Chicago/Turabian StyleBruckberger, Melanie C., Deirdre B. Gleeson, Trevor P. Bastow, Matthew J. Morgan, Tom Walsh, John L. Rayner, Greg B. Davis, and Geoffrey J. Puzon. 2021. "Unravelling Microbial Communities Associated with Different Light Non-Aqueous Phase Liquid Types Undergoing Natural Source Zone Depletion Processes at a Legacy Petroleum Site" Water 13, no. 7: 898. https://doi.org/10.3390/w13070898