
The Utility of Dreissena polymorpha for Assessing the Viral Contamination of Rivers by Measuring the Accumulation of F-Specific RNA Bacteriophages


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
- an in situ experiment to confirm that D. polymorpha naturally accumulates FRNAPHs,
- a laboratory experiment to determine the accumulation and depuration kinetics of FRNAPHs in D. polymorpha tissue.
2. Materials and Methods
2.1. Mussel Collection and Maintenance
2.2. Experimental Design
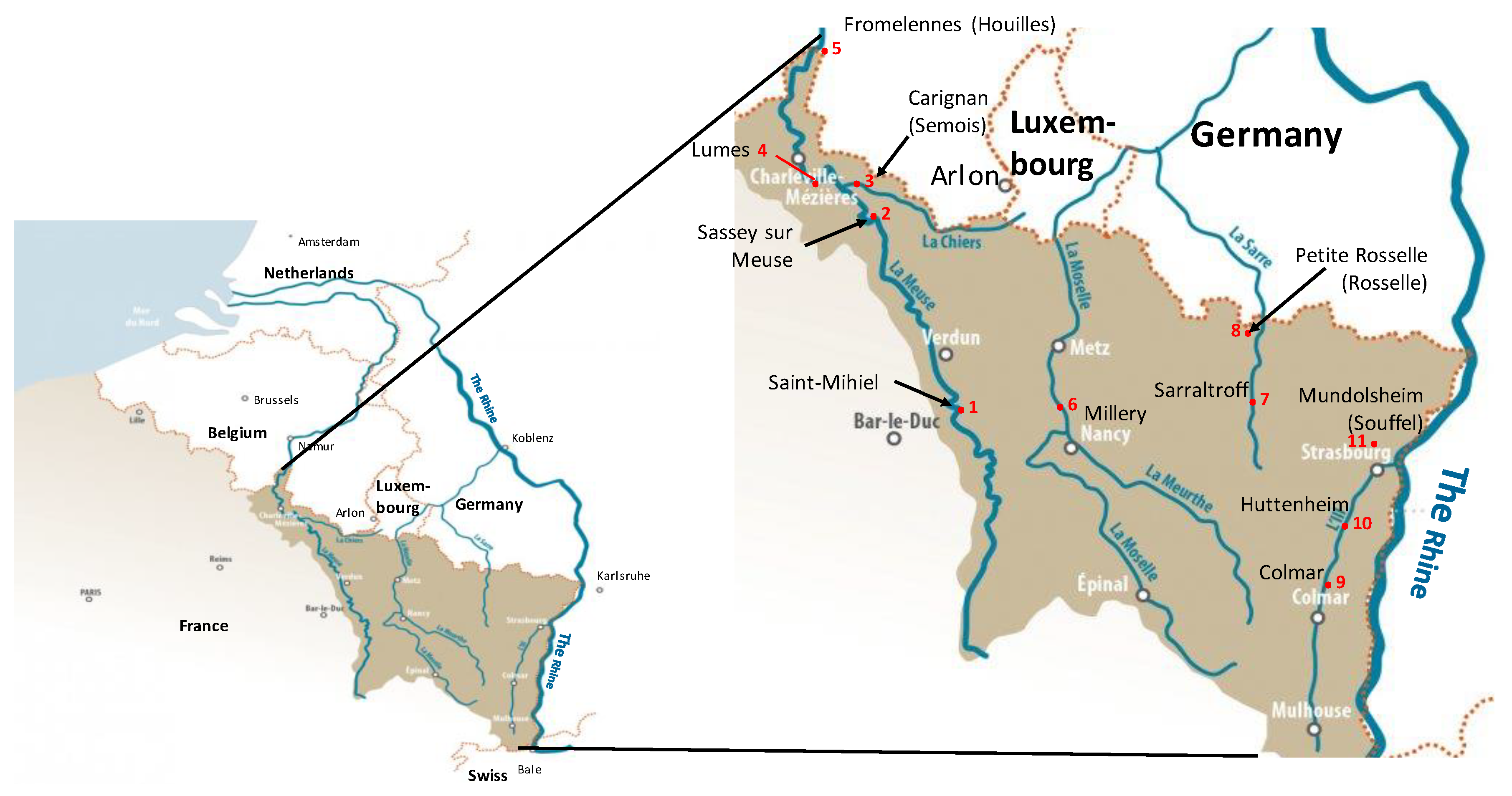
2.2.1. The In Situ Experiment
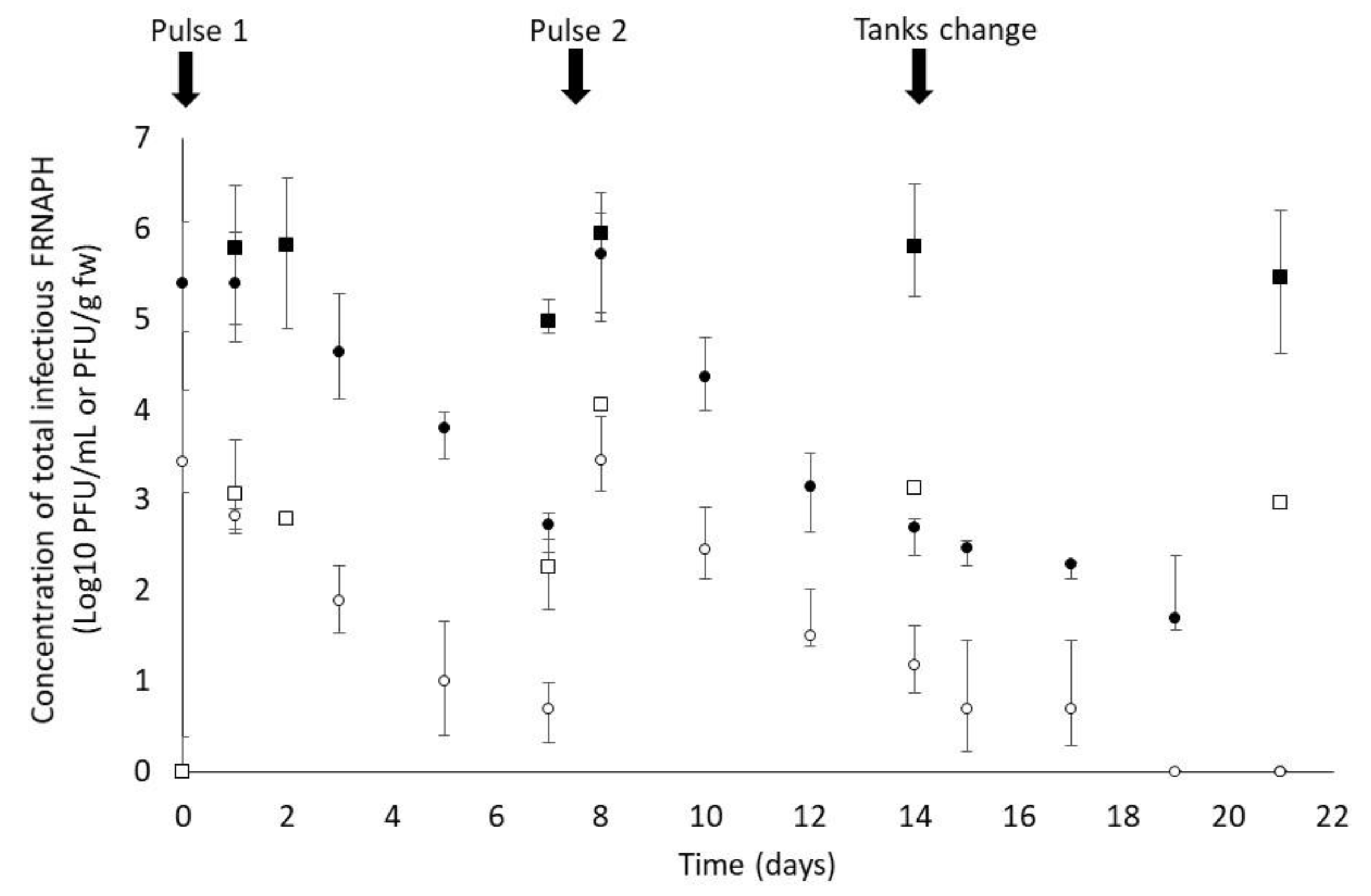
2.2.2. Laboratory Experiments
2.3. Quantification of Infectious FRNAPHs
2.4. Statistical Treatment
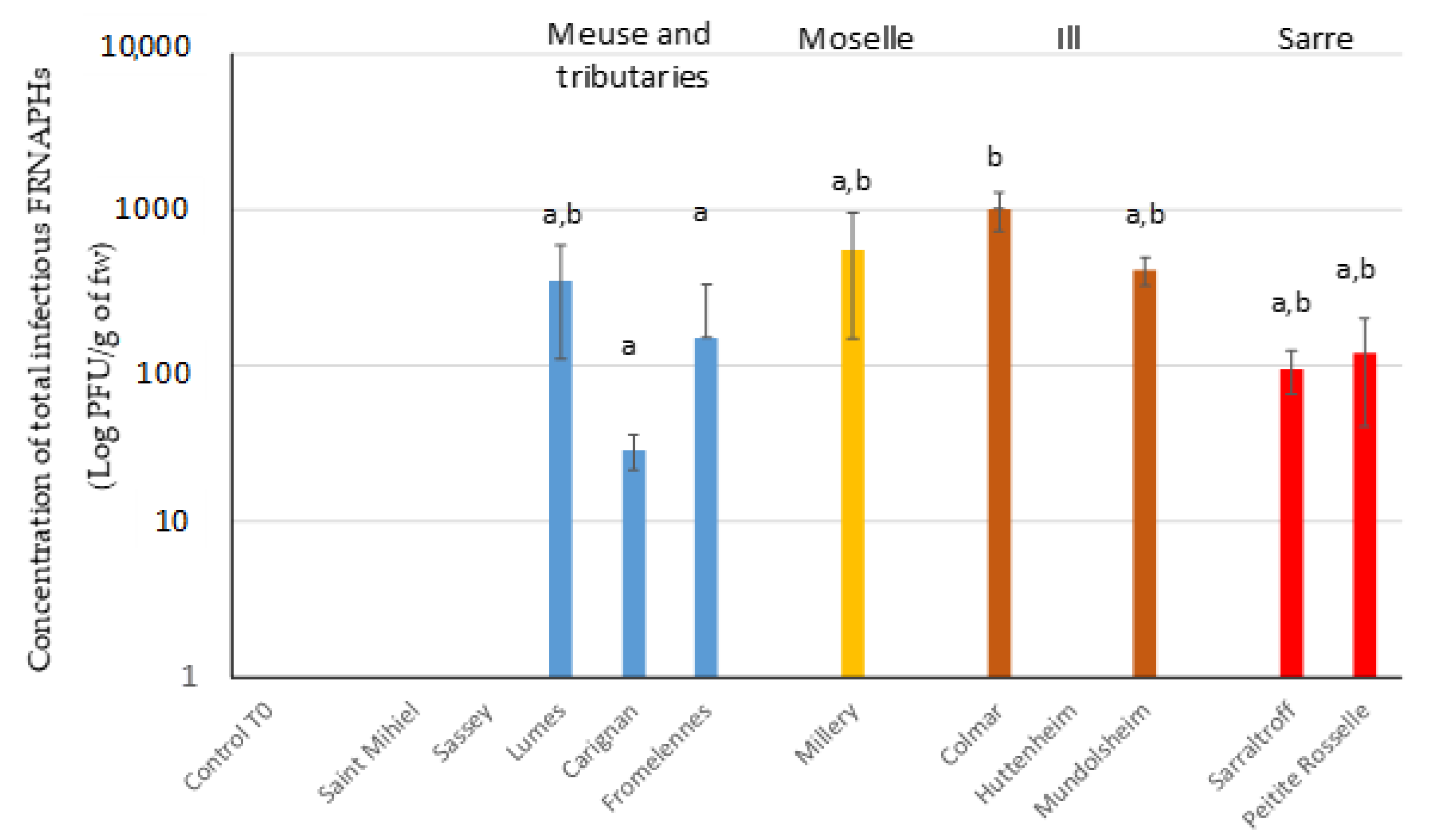
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bosch, A. Human Viruses in Water, 1st ed.; Elsevier: Amsterdam, The Netherlands, 2007; Volume 17. [Google Scholar]
- Fout, G.S.; Borchardt, M.A.; Kieke, B.A.; Karim, M.R. Human virus and microbial indicator occurrence in public-supply groundwater systems: Meta-analysis of 12 international studies. Hydrogeol. J. 2017, 25, 903–919. [Google Scholar] [CrossRef] [Green Version]
- Atmar, R.L.; Opekun, A.R.; Gilger, M.A.; Estes, M.K.; Crawford, S.E.; Neill, F.H.; Graham, D.Y. Norwalk virus shedding after experimental human infection. Emerg. Infect. Dis. 2008, 14, 1553–1557. [Google Scholar] [CrossRef] [PubMed]
- Teunis, P.F.M.; Moe, C.L.; Liu, P.; Miller, S.E.; Lindesmith, L.; Baric, R.S.; Le Pendu, J.; Calderon, R.L. Norwalk virus: How infectious is it? J. Med. Virol. 2008, 80, 1468–1476. [Google Scholar] [CrossRef] [PubMed]
- Atmar, R.L.; Opekun, A.R.; Gilger, M.A.; Estes, M.K.; Crawford, S.E.; Neill, F.H.; Ramani, S.; Hill, H.; Ferreira, J. Determination of the 50% human infectious dose for Norwalk virus. J. Infect. Dis. 2014, 209, 1016–1022. [Google Scholar] [CrossRef]
- Kauppinen, A.; Miettinen, I.T. Persistence of Norovirus GII Genome in Drinking Water and Wastewater at Different Temperatures. Pathogens 2017, 6, 48. [Google Scholar] [CrossRef]
- Skraber, S.; Gassilloud, B.; Gantzer, C. Comparison of coliforms and coliphages as tools for assessment of viral contamination in river water. Appl. Environ. Microbiol. 2004, 70, 3644–3649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jofre, J.; Lucena, F.; Blanch, A.R.; Muniesa, M. Coliphages as Model Organisms in the Characterization and Management of Water Resources. Water 2016, 8, 199. [Google Scholar] [CrossRef] [Green Version]
- Lucena, F.; Méndez, X.; Morón, A.; Calderón, E.; Campos, C.; Guerrero, A.; Cárdenas, M.; Gantzer, C.; Shwartzbrood, L.; Skraber, S.; et al. Occurrence and densities of bacteriophages proposed as indicators and bacterial indicators in river waters from Europe and South America. J. Appl. Microbiol. 2003, 94, 808–815. [Google Scholar] [CrossRef]
- Hartard, C.; Rivet, R.; Banas, S.; Gantzer, C. Occurrence of and Sequence Variation among F-Specific RNA Bacteriophage Subgroups in Feces and Wastewater of Urban and Animal Origins. Appl. Environ. Microbiol. 2015, 8, 6505–6515. [Google Scholar] [CrossRef] [Green Version]
- Flannery, J.; Keaveney, S.; Doré, W. Use of FRNA Bacteriophages to Indicate the Risk of Norovirus Contamination in Irish Oysters. J. Food Prot. 2009, 72, 2358–2362. [Google Scholar] [CrossRef]
- Hartard, C.; Banas, S.; Rivet, R.; Boudaud, N.; Gantzer, C. Rapid and sensitive method to assess human viral pollution in shellfish using infectious F-specific RNA bacteriophages: Application to marketed products. Food Microbio. 2017, 63, 48–254. [Google Scholar] [CrossRef]
- Ogorzaly, L.; Gantzer, C. Development of real-time RT-PCR methods for specific detection of F-specific RNA bacteriophage genogroups: Application to urban raw wastewater. J. Virol Methods 2006, 138, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Wolf, S.; Hewitt, J.; Greening, G.E. Viral multiplex quantitative PCR assays for tracking sources of fecal contamination. Appl. Environ. Microbiol. 2010, 76, 1388–1394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cole, D.; Long, S.C.; Sobsey, M.D. Evaluation of F+ RNA and DNA Coliphages as Source-Specific Indicators of Fecal Con-tamination in Surface Waters. Appl. Environ. Microbiol. 2003, 69, 6507–6514. [Google Scholar] [CrossRef] [Green Version]
- Fauvel, B.; Cauchie, H.-M.; Gantzer, C.; Ogorzaly, L. Contribution of hydrological data to the understanding of the spatio-temporal dynamics of F-specific RNA bacteriophages in river water during rainfull-runoff events. Water Res. 2016, 94, 328–340. [Google Scholar] [CrossRef] [PubMed]
- Fauvel, B.; Ogorzaly, L.; Cauchie, H.M.; Gantzer, C. Interactions of infectious F-specific RNA bacteriophages with suspended matter and sediment: Towards an understanding of FRNAPH distribution in a river water system. Sci. Total Environ. 2017, 574, 960–968. [Google Scholar] [CrossRef] [Green Version]
- Carlton, E.J.; Eisenberg, J.N.S.; Goldstick, J.; Cevallos, W.; Trostle, J.; Levy, K. Heavy rainfall events and diarrhea incidence: The role of social and environmental factors. Am. J. Epidemiol. 2014, 179, 344–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stumpf, P.; Failing, K.; Papp, T.; Nazir, J.; Böhm, R.; Marschang, R.E. Accumulation of a low pathogenic avian influenza virus in zebra mussels (Dreissena polymorpha). Avian Dis. 2010, 54, 1183–1190. [Google Scholar] [CrossRef] [PubMed]
- La Rosa, G.; Mancini, P.; Bonanno Ferraro, G.; Iaconelli, M.; Veneri, C.; Paradiso, R.; De Medici, J.D.; Vicenza, T.; Proroga, Y.T.R.; Di Maro, O.; et al. Hepatitis A Virus Strains Circulating in the Campania Region (2015–2018) Assessed through Bivalve Biomonitoring and Environmental Surveillance. Viruses 2020, 13, 16. [Google Scholar] [CrossRef]
- Bourgeault, A.; Gourlay-Francé, C.; Vincent-Hubert, F.; Palais, F.; Geffard, A.; Biagianti-Risbourg, S.; Pain-Devin, S.; Tusseau-Vuillemin, M.-H. Lessons from a transplantation of zebra mussels into a small urban river: An integrated ecotoxicological assessment. Environ. Toxicol. 2010, 25, 468–478. [Google Scholar] [CrossRef]
- Besse, J.P.; Coquery, M.; Lopes, C.; Chaumot, A.; Budzinski, H.; Labadie, P.; Geffard, O. Caged Gammarus fossarum (Crustacea) as a robust tool for the characterization of bioavailable contamination levels in continental waters: Towards the determination of threshold values. Water Res. 2013, 47, 650–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palos Ladeiro, M.; Barjhoux, I.; Bigot-Clivot, A.; Bonnard, M.; David, E.; Dedourge-Geffard, O.; Geba, E.; Lance, E.; Lepretre, M.; Magniez, G.; et al. Mussel as a tool to define continental watershed quality. Org. Mol. Malacol. 2007. [Google Scholar] [CrossRef] [Green Version]
- Muniain-Mujika, I.; Calvo, M.; Lucena, F.; Girones, R. Comparative analysis of viral pathogens and potential indicators in shellfish. Int. J. Food Microbiol. 2003, 83, 75–85. [Google Scholar] [CrossRef]
- Hartard, C.; Leclerc, M.; Rivet, R.; Maul, A.; Loutreul, J.; Banas, S.; Boudaud, N.; Gantzer, C. F-specific RNA bacteriophages, especially members of subgroup II, should be reconsidered as good indicators of viral pollution of oysters. Appl. Environ. Microbiol. 2018, 84, e01866-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binelli, A.; Della Torre, C.; Magni, S.; Parolini, M. Does zebra mussel (Dreissena polymorpha) represent the freshwater counterpart of Mytilus in ecotoxicological studies? A critical review. Environ. Pollut. 2015, 196, 386–403. [Google Scholar] [CrossRef] [PubMed]
- Kerambrun, E.; Palos Ladeiro, M.; Bigot-Clivot, A.; Dedourge-Geffard, O.; Dupuis, E.; Villena, I.; Aubert, D.; Geffard, A. Zebra mussel as a new tool to show evidence of freshwater contamination by waterborne Toxoplasma gondii. J. Appl. Microbiol. 2016, 120, 498–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bervoets, L.; Voets, J.; Covaci, A.; Chu, S.; Qadah, D.; Smolders, R.; Schepens, P.; Blust, R. Use of transplanted zebra mussels (Dreissena polymorpha) to assess the bioavailability of microcontaminants in flemish surface waters. Environ. Sci. Technol. 2005, 39, 1492–1505. [Google Scholar] [CrossRef]
- Palos Ladeiro, M.; Aubert, D.; Villena, I.; Geffard, A.; Bigot, A. Bioaccumulation of human waterborne protozoa by zebra mussel (Dreissena polymorpha): Interest for water biomonitoring. Water Res. 2014, 48, 148–155. [Google Scholar] [CrossRef]
- Géba, E.; Aubert, D.; Durand, L.; Escotte, S.; La Carbona, S.; Cazeaux, C.; Bonnard, I.; Bastien, F.; Palos Ladeiro, M.; Dubey, J.P.; et al. Use of the bivalve Dreissena polymorpha as a biomonitoring tool to reflect the protozoan load in freshwater bodies. Water Res. 2020, 170, 115297. [Google Scholar] [CrossRef]
- Mezzanotte, V.; Marazzi, F.; Bissa, M.; Pacchioni, S.; Binelli, A.; Parolini, M.; Magni, S.; Franco, M.; Ruggeri, F.; De Giuli Morghen, C.; et al. Removal of enteric viruses and Escherichia coli from municipal treated effluent by zebra mussels. Sci. Total Environ. 2016, 539, 395–400. [Google Scholar] [CrossRef]
- Bighiu, M.A.; Haldén, A.N.; Goedkoop, W.; Ottoson, J. Assessing microbial contamination and antibiotic resistant bacteria using zebra mussels (Dreissena polymorpha). Sci. Total Environ. 2019, 650, 2141–2149. [Google Scholar] [CrossRef]
- International Organization for Standardization. Water Quality—Detection and Enumeration of Bacteriophages—Part 1: Enumeration of F-Specific RNA Bacteriophages; ISO 10705-1; International Organization for Standardization: Geneva, Switzerland, 2001. [Google Scholar]
- Havelaar, A.H.; Hogeboom, W.M. A method for the enumeration of male-specific bacteriophages in sewage. J. Appl. Bacteriol. 1984, 56, 439–447. [Google Scholar] [CrossRef]
- Schaper, M.; Jofre, J.; Uys, M.; Grabow, W.O.K. Distribution of genotypes of F-specific RNA bacteriophages in human and non-human sources of faecal pollution in South Africa and Spain. J. Appl. Microbiol. 2002, 92, 657–667. [Google Scholar] [CrossRef] [PubMed]
- Leduc, A.; Leclerc, M.; Challant, J.; Loutreul, J.; Robin, M.; Maul, A.; Majou, D.; Boudaud, N.; Gantzer, C. F-Specific RNA bacteriophages model the behavior of human noroviruses during purification of oysters: The main mechanism is probably inactivation rather than release. Appl. Environ. Microbiol. 2020, 86, e00526-20. [Google Scholar] [CrossRef] [PubMed]
- Le Guyader, F.; Loisy, F.; Atmar, R.L.; Hutson, A.M.; Estes, M.K.; Ruvoën-Clouet, N.; Pommepuy, M.; Le Pendu, J. Norwalk virus-specific binding to oyster digestive tissues. Emerg. Infect. Dis. 2006, 12, 931–936. [Google Scholar] [CrossRef] [PubMed]
- Tian, P.; Engelbrektson, A.L.; Jiang, X.; Zhong, W.; Mandrell, R.E. Norovirus recognizes histo-blood group antigens on gastrointestinal cells of clams, mussels, and oysters: A possible mechanism of bioaccumulation. J. Food Protect. 2007, 70, 2140–2147. [Google Scholar] [CrossRef] [Green Version]
- Maalouf, H.; Schaeffer, J.; Parnaudeau, S.; Le Pendu, J.; Atmar, R.L.; Crawford, S.E.; Le Guyader, F.S. Strain-dependent norovirus bioaccumulation in oysters. Appl. Environ. Microbiol. 2011, 77, 3189–3196. [Google Scholar] [CrossRef] [Green Version]
- Maalouf, H.; Zakhour, M.; Pendu, J.L.; Le Saux, J.-C.; Atmar, R.L.; Le Guyader, F.S. Distribution in tissue and seasonal variation of norovirus genogroup I and II ligands in oysters. App. Environ. Microbiol. 2010, 76, 5621–5630. [Google Scholar] [CrossRef] [Green Version]
- Lowther, J.A.; Cross, L.; Stapleton, T.; Gustar, N.E.; Walker, D.I.; Sills, M.; Treagus, S.; Pollington, V.; Lees, D.N. Use of F-Specific RNA Bacteriophage to Estimate Infectious Norovirus Levels in Oysters. Food. Environ. Virol. 2019, 11, 247–258. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Capizzi-Banas, S.; Palos Ladeiro, M.; Bastien, F.; Bonnard, I.; Boudaud, N.; Gantzer, C.; Geffard, A. The Utility of Dreissena polymorpha for Assessing the Viral Contamination of Rivers by Measuring the Accumulation of F-Specific RNA Bacteriophages. Water 2021, 13, 904. https://doi.org/10.3390/w13070904
Capizzi-Banas S, Palos Ladeiro M, Bastien F, Bonnard I, Boudaud N, Gantzer C, Geffard A. The Utility of Dreissena polymorpha for Assessing the Viral Contamination of Rivers by Measuring the Accumulation of F-Specific RNA Bacteriophages. Water. 2021; 13(7):904. https://doi.org/10.3390/w13070904
Chicago/Turabian StyleCapizzi-Banas, Sandrine, Mélissa Palos Ladeiro, Fanny Bastien, Isabelle Bonnard, Nicolas Boudaud, Christophe Gantzer, and Alain Geffard. 2021. "The Utility of Dreissena polymorpha for Assessing the Viral Contamination of Rivers by Measuring the Accumulation of F-Specific RNA Bacteriophages" Water 13, no. 7: 904. https://doi.org/10.3390/w13070904