
Distribution and Public Health Significance of Vibrio Pathogens Recovered from Selected Treated Effluents in the Eastern Cape Province, South Africa


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Water Collections
2.2. Determination of Physicochemical Parameters
2.3. Bacterial Strains
2.4. Bacterial Isolation and Enrichments of Samples
2.5. Confirmation of Probable Vibrio Isolates
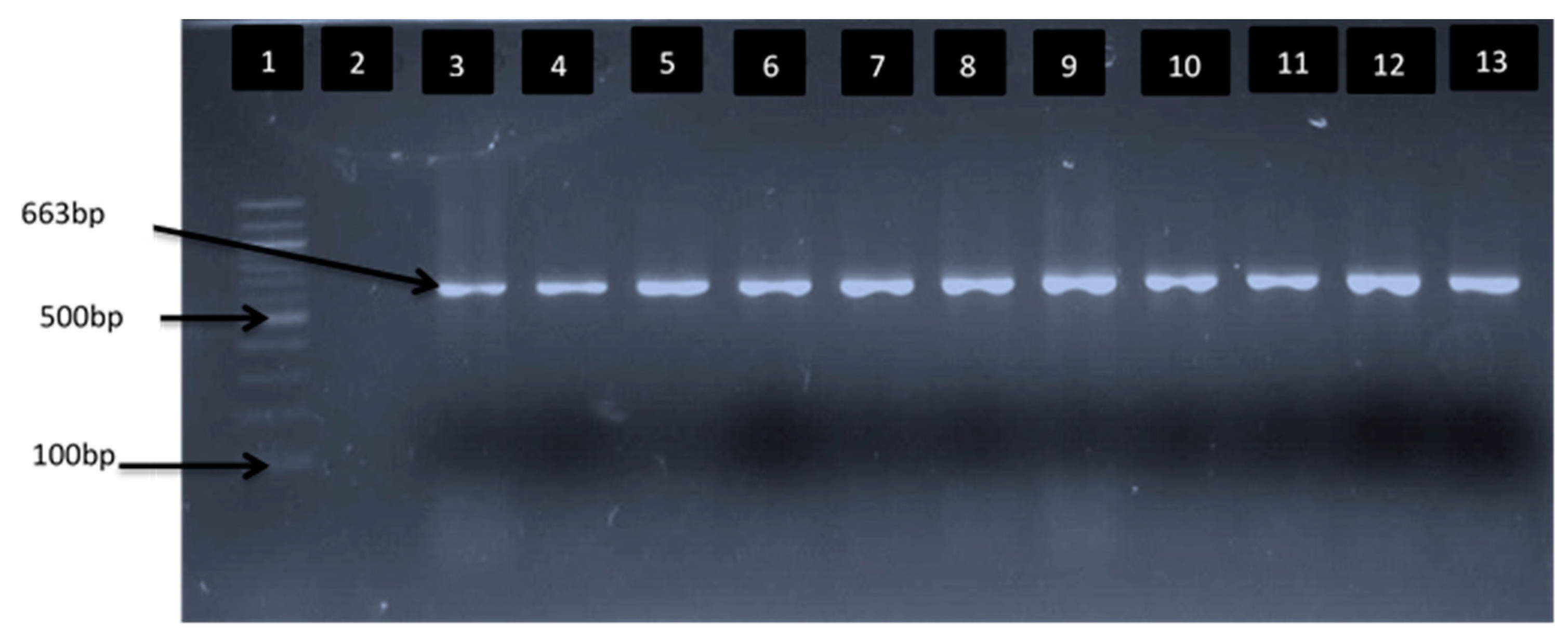
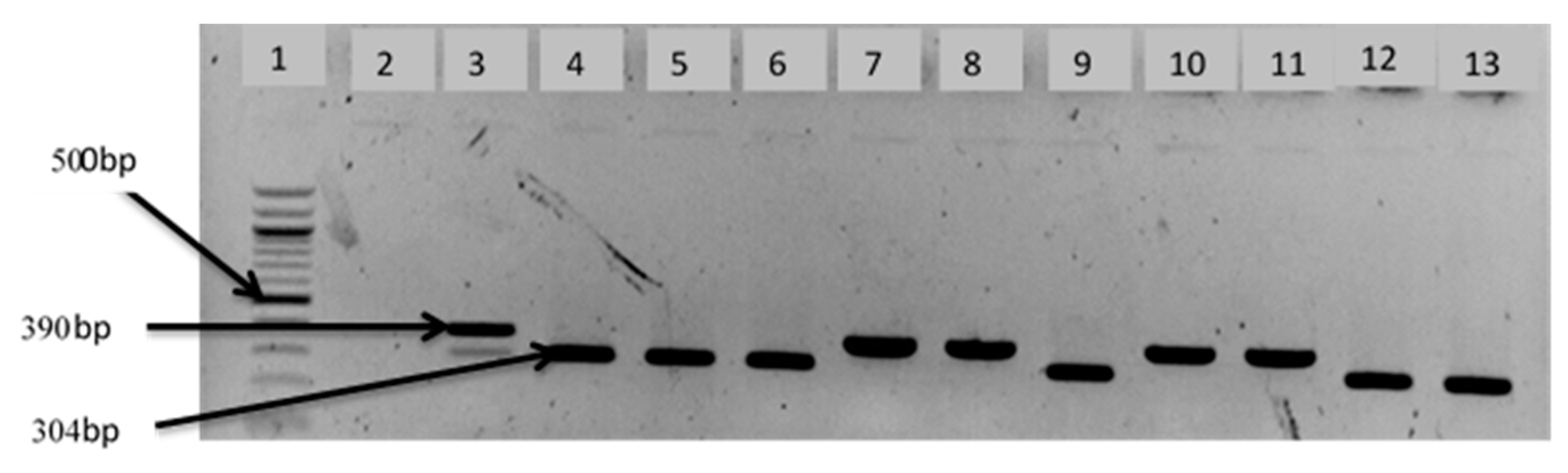
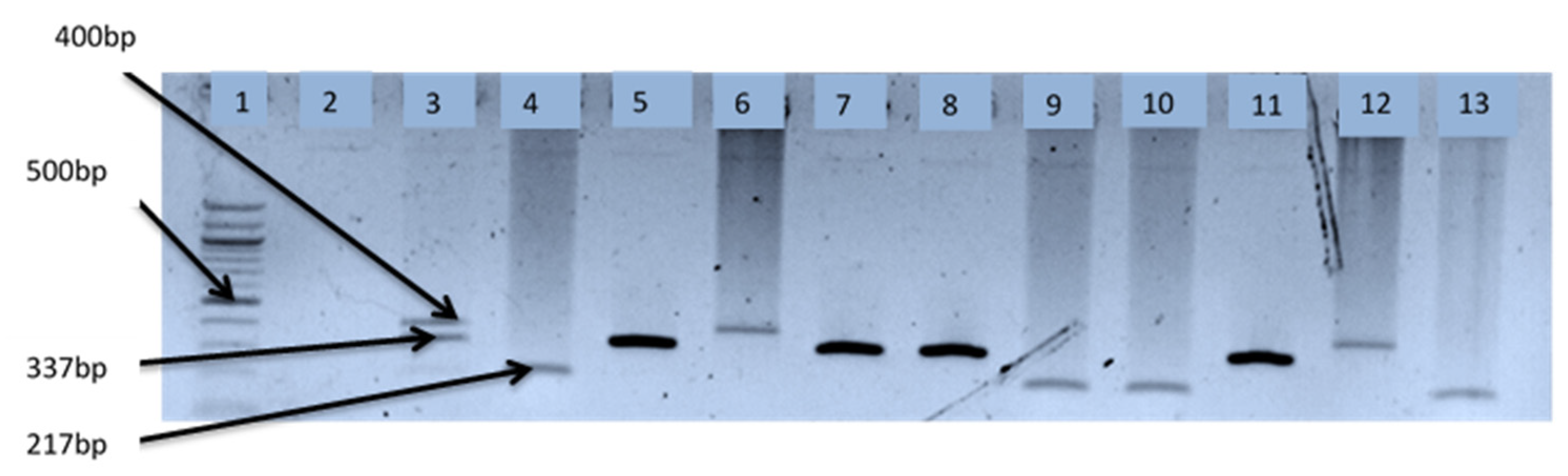
2.6. Delineation of Vibrio Species Isolates
2.7. Statistical Analysis
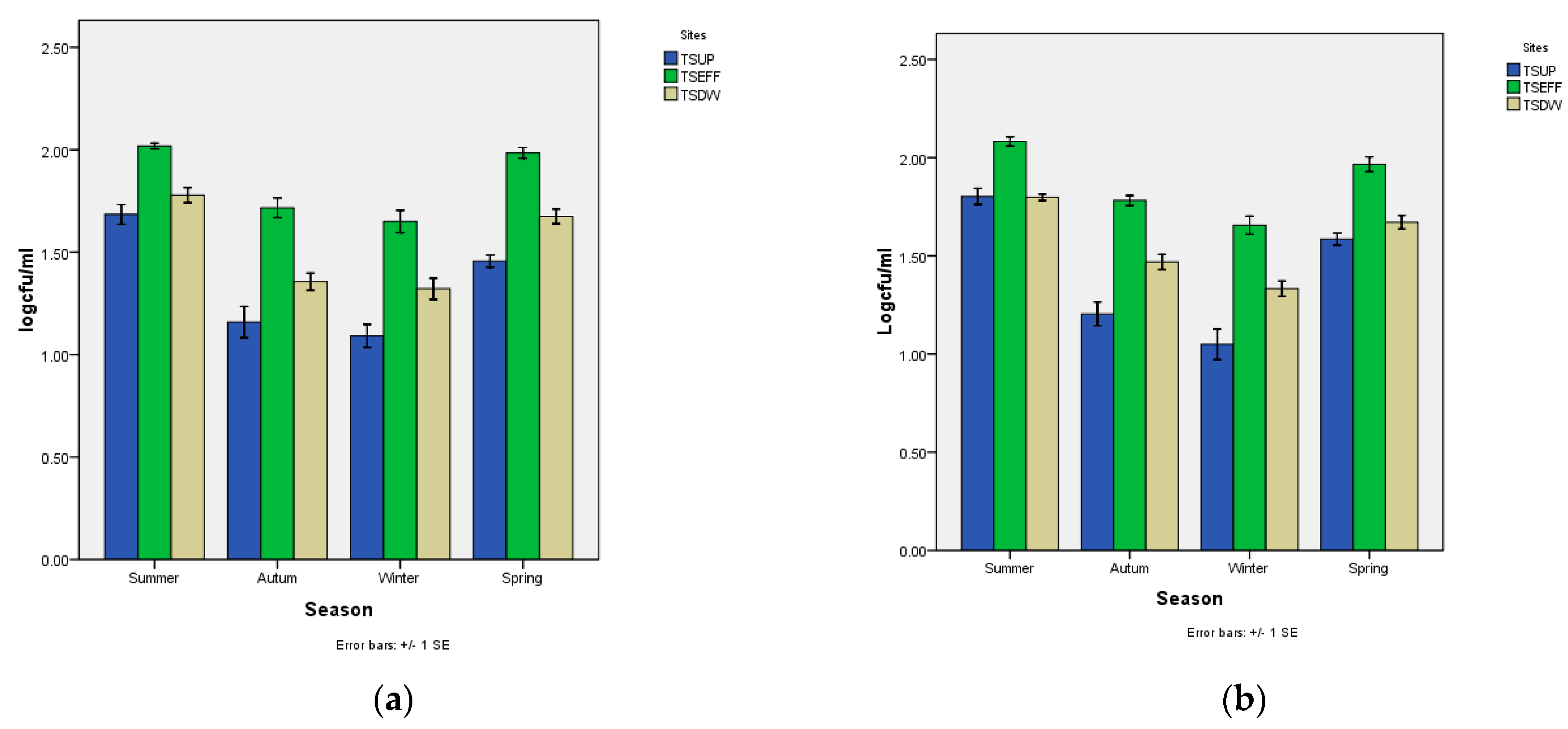
3. Results
4. Discussions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Al-Gheethi, A.A.; Efaq, A.N.; Bala, J.D.; Norli, I.; Abdel-Monem, M.O.; Kadir, M.A. Removal of pathogenic bacteria from sewage-treated effluent and biosolids for agricultural purposes. Appl. Water Sci. 2018, 82, 74. [Google Scholar] [CrossRef] [Green Version]
- Pandey, P.K.; Kass, P.H.; Soupir, M.L.; Biswas, S.; Singh, V.P. Contamination of water resources by pathogenic bacteria. AMB Express 2014, 4, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Cai, L.; Zhang, T. Detecting human bacterial pathogens in wastewater treatment plants by a high-throughput shotgun sequencing technique. Environ. Sci. Technol. 2013, 47, 5433–5441. [Google Scholar] [CrossRef]
- Cantalupo, P.G.; Calgua, B.; Zhao, G.; Hundesa, A.; Wier, A.D.; Katz, J.P.; Grabe, M.; Hendrix, R.W.; Girones, R.; Wang, D.; et al. Raw sewage harbors diverse viral populations. mBio 2011, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chorev, N. The World Health Organization between North and South; Cornell University Press: Ithaca, NY, USA, 2012. [Google Scholar]
- Oluseyi Osunmakinde, C.; Selvarajan, R.; Mamba, B.B.; Msagati, T.A. Profiling bacterial diversity and potential pathogens in wastewater treatment plants using high-throughput sequencing analysis. Microorganisms 2019, 7, 506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aghalari, Z.; Dahms, H.U.; Sillanpää, M.; Sosa-Hernandez, J.E.; Parra-Saldívar, R. Effectiveness of wastewater treatment systems in removing microbial agents: A systematic review. Glob. Health 2020, 16, 13. [Google Scholar] [CrossRef] [PubMed]
- Becerra-Castro, C.; Lopes, A.R.; Vaz-Moreira, I.; Silva, E.F.; Manaia, C.M.; Nunes, O.C. Wastewater reuse in irrigation: A microbiological perspective on implications in soil fertility and human and environmental health. Environ. Int. 2015, 75, 117–135. [Google Scholar] [CrossRef] [Green Version]
- Osunla, C.A.; Okoh, A.I. Vibrio pathogens: A public health concern in rural water resources in Sub-Saharan Africa. Int. J. Environ. Res. Public Health 2017, 14, 1188. [Google Scholar] [CrossRef] [Green Version]
- Miyoshi, S.I. Extracellular proteolytic enzymes produced by human pathogenic Vibrio species. Front. Microbiol. 2013, 4, 339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lutz, C.; Erken, M.; Noorian, P.; Sun, S.; McDougald, D. Environmental reservoirs and mechanisms of persistence of Vibrio cholerae. Front. Microbiol. 2013, 4, 375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eja, M.E.; Abriba, C.; Etok, C.A.; Ikpeme, E.M.; Arikpo, G.E.; Enyi-Idoh, K.H.; Ofor, U.A. Seasonal occurrence of Vibrios in water and shellfish obtained from the Great Kwa River Estuary, Calabar, Nigeria. Bull. Environ. Contam. Toxicol. 2008, 81, 245–248. [Google Scholar] [CrossRef]
- Osuolale, O.; Okoh, A. Isolation and antibiotic profile of Vibrio spp. in final effluents of two wastewater treatment plants in the Eastern Cape of South Africa. BioRxiv 2018, 330456. [Google Scholar] [CrossRef] [Green Version]
- Baron, S.; Larvor, E.; Chevalier, S.; Jouy, E.; Kempf, I.; Granier, S.A.; Lesne, J. Antimicrobial susceptibility among urban wastewater and wild shellfish isolates of non-O1/Non-O139 Vibrio cholerae from La Rance Estuary (Brittany, France). Front. Microbiol. 2017, 8, 1637. [Google Scholar] [CrossRef] [Green Version]
- Baker-Austin, C.; Trinanes, J.; Gonzalez-Escalona, N.; Martinez-Urtaza, J. Non-cholera vibrios: The microbial barometer of climate change. Trends Microbiol. 2017, 25, 76–84. [Google Scholar] [CrossRef] [Green Version]
- Momba, M.; Azab El-Liethy, M. Vibrio cholerae and Cholera biotypes. Glob. Water Pathog. Proj. Available online: https://www.waterpathogens.org/sites/default/files/Vibrio%20cholerae%20and%20Cholera%20biotypes_4.pdf (accessed on 22 September 2020).
- Olaniran, A.O.; Nzimande, S.B.; Mkize, N.G. Antimicrobial resistance and virulence signatures of Listeria and Aeromonas species recovered from treated wastewater effluent and receiving surface water in Durban, South Africa. BMC Microbiol. 2015, 15, 1–10. [Google Scholar] [CrossRef]
- Verlicchi, P.; Grillini, V. Surface water and groundwater quality in South Africa and mozambique—Analysis of the Most critical pollutants for drinking purposes and challenges in water treatment selection. Water 2020, 12, 305. [Google Scholar] [CrossRef] [Green Version]
- American Public Health Association; American Water Works Association; Water Pollution Control Federation; Water Environment Federation. Standard Methods for the Examination of Water and Wastewater; American Public Health Association: Waldorf, MD, USA, 1915. [Google Scholar]
- SABS. SANS 5221: 2011 South African National Standard Microbiological Analysis of Water General Test Methods; South African Bureau of Standards: Pretoria, South Africa, 2011; pp. 7–11.
- Maugeri, T.L.; Carbone, M.; Fera, M.T.; Gugliandolo, C. Detection and differentiation of Vibrio vulnificus in seawater and plankton of a coastal zone of the Mediterranean Sea. Res. Microbiol. 2006, 157, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Kwok, A.Y.; Wilson, J.T.; Coulthart, M.; Ng, L.K.; Mutharia, L.; Chow, A.W. Phylogenetic study and identification of human pathogenic Vibrio species based on partial hsp 60 gene sequences. Can. J. Microbiol. 2002, 48, 903–910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okoh, A.I.; Sibanda, T.; Nongogo, V.; Adefisoye, M.; Olayemi, O.O.; Nontongana, N. Prevalence and characterisation of non-cholerae Vibrio spp. in final effluents of wastewater treatment facilities in two districts of the Eastern Cape Province of South Africa: Implications for public health. Environ. Sci. Pollut. Res. 2015, 22, 2008–2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menezes, F.G.; Neves, S.D.; Sousa, O.V.; Vila-Nova, C.M.; Maggioni, R.; Theophilo, G.N.; Hofer, E.; Vieira, R.H. Detection of virulence genes in environmental strains of Vibrio cholerae from estuaries in northeastern Brazil. Rev. Inst. Med. Trop. São Paulo 2014, 56, 427–432. [Google Scholar] [CrossRef]
- Goel, A.K.; Jain, M.; Kumar, P.; Jiang, S.C. Molecular characterization of Vibrio cholerae outbreak strains with altered El Tor biotype from southern India. World J. Microbiol. Biotechnol. 2010, 26, 281–287. [Google Scholar] [CrossRef] [Green Version]
- Tarr, C.L.; Patel, J.S.; Puhr, N.D.; Sowers, E.G.; Bopp, C.A.; Strockbine, N.A. Identification of Vibrio isolates by a multiplex PCR assay and rpoB sequence determination. J. Clin. Microbiol. 2007, 45, 134–140. [Google Scholar] [CrossRef] [Green Version]
- Osorio, C.R.; Klose, K.E. A region of the transmembrane regulatory protein ToxR that tethers the transcriptional activation domain to the cytoplasmic membrane displays wide divergence among Vibrio species. J. Bacteriol. 2000, 182, 526–528. [Google Scholar] [CrossRef] [Green Version]
- Guardiola-Avila, I.; Acedo-Felix, E.; Sifuentes-Romero, I.; Yepiz-Plascencia, G.; Gomez-Gil, B.; Noriega-Orozco, L. Molecular and genomic characterization of Vibrio mimicus isolated from a frozen shrimp processing facility in Mexico. PLoS ONE 2016, 11, e0144885. [Google Scholar] [CrossRef]
- Wei, S.; Zhao, H.; Xian, Y.; Hussain, M.A.; Wu, X. Multiplex PCR assays for the detection of Vibrio alginolyticus, Vibrio parahaemolyticus, Vibrio vulnificus and Vibrio cholerae with an internal amplification control. Diagn. Microbiol. Infect. Dis. 2014, 79, 115–118. [Google Scholar] [CrossRef]
- Abioye, O.E.; Okoh, A.I. Limpet (Scutellastra cochlear) Recovered From Some Estuaries in the Eastern Cape Province, South Africa Act as Reservoirs of Pathogenic Vibrio Species. Front. Public Health 2018, 6, 237. [Google Scholar] [CrossRef]
- Patel, C.B.; Shanker, R.; Gupta, V.K.; Upadhyay, R.S. Q-PCR based culture-independent enumeration and detection of Enterobacter: An emerging environmental human pathogen in riverine systems and potable water. Front. Microbiol. 2016, 7, 172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pruzzo, C.; Gallo, G.; Canesi, L. Persistence of vibrios in marine bivalves: The role of interactions with haemolymph components. Environ. Microbiol. 2005, 7, 761–772. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Lin, H.; Wang, X.; Austin, B. Significance of Vibrio species in the marine organic carbon cycle—A review. Sci. China Earth Sci. 2018, 61, 1357–1368. [Google Scholar] [CrossRef]
- Rubio-Portillo, E.; Kersting, D.K.; Linares, C.; Ramos-Esplá, A.A.; Antón, J. Biogeographic differences in the microbiome and pathobiome of the coral Cladocora caespitosa in the Western Mediterranean Sea. Front. Microbiol. 2018, 9, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorrasi, S.; Pasqualetti, M.; Franzetti, A.; Pittino, F.; Fenice, M. Vibrio communities along a salinity gradient within a marine saltern hypersaline environment (Saline di Tarquinia, Italy). Environ. Microbiol. 2020, 22, 4356–4366. [Google Scholar] [CrossRef]
- Preisner, M. Surface Water Pollution by Untreated Municipal Wastewater Discharge Due to a Sewer Failure. Environ. Process. 2020, 7, 767–780. [Google Scholar] [CrossRef]
- Ibekwe, A.M.; Ma, J.; Murinda, S.E. Bacterial community composition and structure in an Urban River impacted by different pollutant sources. Sci. Total Environ. 2016, 566, 1176–1185. [Google Scholar] [CrossRef] [Green Version]
- Haque, M.A.; Jewel, M.A.; Sultana, M.P. Assessment of physicochemical and bacteriological parameters in surface water of Padma River, Bangladesh. Appl. Water Sci. 2019, 9, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Chase, E.; Harwood, V.J. Comparison of the effects of environmental parameters on growth rates of Vibrio vulnificus biotypes I, II, and III by culture and quantitative PCR analysis. Appl. Environ. Microbiol. 2011, 77, 4200–4207. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez-Salazar, G.J.; Molina-Garza, Z.J.; Hernández-Acosta, M.; García-Salas, J.A.; Mercado-Hernández, R.; Galaviz-Silva, L. Pathogens in Pacific white shrimp (Litopenaeus vannamei Boone, 1931) and their relationship with physicochemical parameters in three different culture systems in Tamaulipas, Mexico. Aquaculture 2011, 321, 34–40. [Google Scholar] [CrossRef]
- Jin, D.; Kong, X.; Cui, B.; Jin, S.; Xie, Y.; Wang, X.; Deng, Y. Bacterial communities and potential waterborne pathogens within the typical urban surface waters. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef]
- Vezzulli, L.; Colwell, R.R.; Pruzzo, C. Ocean warming and spread of pathogenic vibrios in the aquatic environment. Microb. Ecol. 2013, 65, 817–825. [Google Scholar] [CrossRef]
- Montánchez, I.; Ogayar, E.; Plágaro, A.H.; Esteve-Codina, A.; Gómez-Garrido, J.; Orruño, M.; Arana, I.; Kaberdin, V.R. Analysis of Vibrio harveyi adaptation in sea water microcosms at elevated temperature provides insights into the putative mechanisms of its persistence and spread in the time of global warming. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Vezzulli, L.; Grande, C.; Reid, P.C.; Hélaouët, P.; Edwards, M.; Höfle, M.G.; Brettar, I.; Colwell, R.R.; Pruzzo, C. Climate influence on Vibrio and associated human diseases during the past half-century in the coastal North Atlantic. Proc. Natl. Acad. Sci. USA 2016, 113, E5062–E5071. [Google Scholar] [CrossRef] [Green Version]
- Brenzinger, S.; van der Aart, L.T.; van Wezel, G.P.; Lacroix, J.M.; Glatter, T.; Briegel, A. Structural and proteomic changes in viable but non-culturable Vibrio cholerae. Front. Microbiol. 2019, 10, 793. [Google Scholar] [CrossRef] [Green Version]
- Dechet, A.M.; Yu, P.A.; Koram, N.; Painter, J. Nonfoodborne Vibrio infections: An important cause of morbidity and mortality in the United States, 1997–2006. Clin. Infect. Dis. 2008, 46, 970–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makuwa, S.; Tlou, M.; Fosso-Kankeu, E.; Green, E. Evaluation of fecal coliform prevalence and physicochemical indicators in the effluent from a wastewater treatment plant in the north-west province, South Africa. Int. J. Environ. Res. Public Health 2020, 17, 6381. [Google Scholar] [CrossRef]
- Manning, T.M.; Wilson, S.P.; Chapman, J.C. Toxicity of chlorine and other chlorinated compounds to some Australian aquatic organisms. Bull. Environ. Contam. Toxicol. 1996, 56, 971–976. [Google Scholar] [CrossRef] [PubMed]
- Government Gazette. National Water Act: Revision of General Authorisations, No. 36820; Government Gazette: Pretoria, South Africa, 2013; pp. 3–31.
- Momba, M.N.; Osode, A.N.; Sibewu, M. The impact of inadequate wastewater treatment on the receiving water bodies–Case study: Buffalo City and Nkokonbe Municipalities of the Eastern Cape Province. Water SA 2006, 32. [Google Scholar] [CrossRef] [Green Version]
- Martone-Rocha, S.; Piveli, R.P.; Matté, G.R.; Dória, M.C.; Dropa, M.; Morita, M.; Peternella, F.A.; Matté, M.H. Dynamics of Aeromonas species isolated from wastewater treatment system. J. Water Health 2010, 8, 703–711. [Google Scholar] [CrossRef] [Green Version]
- Lafdal, M.Y.; Malang, S.; Toguebaye, B.S. Antimicrobial susceptibility of aeromonads and coliforms before and after municipal wastewater treatment by activated sludge under arid climate. Int. J. Microbiol. Res. 2012, 3, 174–180. [Google Scholar]
- Dungeni, M.; van Der Merwe, R.R.; Momba, M. Abundance of pathogenic bacteria and viral indicators in chlorinated effluents produced by four wastewater treatment plants in the Gauteng Province, South Africa. Water SA 2010, 36. [Google Scholar] [CrossRef] [Green Version]
- Ye, L.; Zhang, T. Bacterial communities in different sections of a municipal wastewater treatment plant revealed by 16S rDNA 454 pyrosequencing. Appl. Microbiol. Biotechnol. 2013, 97, 2681–2690. [Google Scholar] [CrossRef] [Green Version]
- Ho, H.; Amin, H.; Lee, H.R. Vibrio Infections; Medscape; Texas, Tech University Health Sciences Center: Houston, TX, USA, 2009; 12p. [Google Scholar]
- Igbinosa, E.O.; Obi, L.C.; Tom, M.; Okoh, A.I. Detection of potential risk of wastewater effluents for transmission of antibiotic resistance from Vibrio species as a reservoir in a peri-urban community in South Africa. Int. J. Environ. Health Res. 2011, 21, 402–414. [Google Scholar] [CrossRef]
- Kokashvili, T.; Whitehouse, C.A.; Tskhvediani, A.; Grim, C.J.; Elbakidze, T.; Mitaishvili, N.; Janelidze, N.; Jaiani, E.; Haley, B.J.; Lashkhi, N.; et al. Occurrence and diversity of clinically important Vibrio species in the aquatic environment of Georgia. Front. Public Health 2015, 3, 232. [Google Scholar] [CrossRef]
- DWAF: Department of Water Affairs. 2013 Green Drop Report; 2016. Available online: https://www.dws.gov.za/Documents/GreenDropReport2009_ver1_web.pdf (accessed on 15 September 2020).
- DWAF: Department of Water Affairs. 2014 Green Drop Report; 2016. Available online: https://www.dws.gov.za/Documents/GreenDropReport2009_ver1_web.pdf (accessed on 15 September 2020).
- Mara, D.; Horan, N.J. (Eds.) Handbook of Water and Wastewater Microbiology; Elsevier: Amsterdam, The Netherlands, 2003. [Google Scholar]
- Nongogo, V.; Okoh, A.I. Occurrence of Vibrio pathotypes in the final effluents of five wastewater treatment plants in Amathole and Chris Hani District Municipalities in South Africa. Int. J. Environ. Res. Public Health 2014, 11, 7755–7766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janelidze, N.; Jaiani, E.; Lashkhi, N.; Tskhvediani, A.; Kokashvili, T.; Gvarishvili, T.; Jgenti, D.; Mikashavidze, E.; Diasamidze, R.; Narodny, S.; et al. Microbial water quality of the Georgian coastal zone of the Black Sea. Mar. Pollut. Bull. 2011, 62, 573–580. [Google Scholar] [CrossRef]
- Morris, J.G., Jr. Non-O group 1 Vibrio cholerae: A look at the epidemiology of an occasional pathogen. Epidemiol. Rev. 1990, 12, 179–191. [Google Scholar] [CrossRef]
- Redondo-Salvo, S.; Fernández-López, R.; Ruiz, R.; Vielva, L.; de Toro, M.; Rocha, E.P.; Garcillán-Barcia, M.P.; de la Cruz, F. Pathways for horizontal gene transfer in bacteria revealed by a global map of their plasmids. Nat. Commun. 2020, 11, 1–13. [Google Scholar] [CrossRef]
- Beshiru, A.; Okareh, O.T.; Okoh, A.I.; Igbinosa, E.O. Detection of antibiotic resistance and virulence genes of Vibrio strains isolated from ready-to-eat shrimps in Delta and Edo States, Nigeria. J. Appl. Microbiol. 2020, 129, 17–36. [Google Scholar] [CrossRef] [PubMed]
- Newton, A.; Kendall, M.; Vugia, D.J.; Henao, O.L.; Mahon, B.E. Increasing rates of vibriosis in the United States, 1996–2010: Review of surveillance data from 2 systems. Clin. Infect. Dis. 2012, 54, S391–S395. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WWTPs | AL | TS |
|---|---|---|
| Municipality | Sarah Baartman District | Chris Hani District |
| Technology | Activated Sludge | Stabilization pond |
| Geographic location | S33.31612626°, E26.107717° | S32.045122, E27.810904 |
| Design capacity (ML/d) | 1.1 | NI |
| Receiving river | Boesman | Tsomo |
| Population statistics (2016) | 138, 182 households | 194, 291 households |
| Specie | Sequence | Size bp | References |
|---|---|---|---|
| Vibrio genus | F: CGG TGA AAT GCG TAG AGA T R: TTA CTA GCG ATT CCG AGT TC | 663 | [22,23] |
| V. cholerae | F: CAC CAA GAA GGT GAC TTT ATT GTG R: GGT TTG TCG AAT TAG CTT CAC C | 304 | [24,25] |
| V. parahaemolyticus | F: GCA GCT GAT CAA AAC GTT GAG T R: ATT ATC GAT CGT GCC ACT CAC | 897 | [23,26] |
| V. vulnificus | F: GTC TTA AAG CGG TTG CTG C R: CGC TTC AAG TGC TGG TAG AAG | 410 | [23] |
| V. Fluvialis | F: GAC CAG GGC TTT GAG GTG GAC GAC R: AGG ATA CGG CAC TTG AGT AAG ACT C | 217 | [23,27] |
| V. Mimicus | F: GGT AGC CAT CAG TCT TAT CAC G R: ATC GTG TCC CAA TAC TTC ACC G | 390 | [28] |
| V. alginolyticus | F: GAG AAC CCG ACA GAA GCG AAG R: CCT AGT GCG GTG ATC AGT GTT G | 337 | [29] |
| TS WWTPs | ||||||
|---|---|---|---|---|---|---|
| Site | Physicochemical Parameters | Cfu/ mLCorel Coeff | Site | Cfu/ mLCorel Coeff | Site | Cfu/ mLCorel Coeff |
| TSUP | pH | −0.327 | TSEFF | 0.218 | TSDW | 0.27 |
| Cond | −0.076 | −0.232 | 0.026 | |||
| TDS | −0.076 | −0.182 | 0.044 | |||
| Sal | −0.064 | −0.175 | 0.014 | |||
| Temp | 0.545 ** | 0.526 ** | 0.517 ** | |||
| DO | −0.393 * | 0.421 * | −0.012 | |||
| BOD | 0.304 | 0.332 | −0.029 | |||
| TSS | 0.387 * | 0.298 | 0.13 | |||
| Turb | 0.386 * | 0.275 | 0.134 | |||
| FreeCl2 | 0.238 | 0.463 ** | ||||
| AL WWTPs | ||||||
| Sites | Physicochemical parameters | Cfu/ mLCorel coeff | Site | Cfu/ mLCorel coeff | Site | Cfu/ mLCorel coeff |
| ALUP | pH | 0.377 * | ALEFF | 0.497 ** | ALDW | 0.607 ** |
| Cond | 0.449 ** | 0.488 ** | 0.331 * | |||
| TDS | 0.543 ** | 0.452 ** | 0.350 * | |||
| Sal | 0.390 * | 0.459 ** | 0.246 | |||
| Temp | 0.509 ** | 0.615 ** | 0.559 ** | |||
| DO | −0.04 | 0.615 ** | 0.07 | |||
| BOD | 0.212 | 0.575 ** | 0.281 | |||
| TSS | −0.185 | 0.511 ** | 0.103 | |||
| Turb | −0.087 | 0.630 ** | 0.197 | |||
| FreeCl2 | 0.137 | 0.107 | ||||
| TS Wastewater Treatment Plant | AL Wastewater Treatment Plant | ||
|---|---|---|---|
| Site Types | p values | Site Types | p values |
| TSUP vs. TSEFF | <0.0001 * | ALUP vs. ALEFF | <0.0001 * |
| TSUP vs. TSDW | 0.007 * | ALUP vs. ALDW | <0.0001 * |
| TSEFF vs. TSDW | <0.0001 * | ALEFF vs. ALDW | <0.0001 * |
| TS Wastewater Treatment Plant | AL Wastewater Treatment Plant | ||
|---|---|---|---|
| Seasons | p values | Seasons | p values |
| Summ vs. Autu | 0.524 | Summ vs. Autu | 0.394 |
| Summ vs. Wint | <0.0001 * | Summ vs. Wint | <0.0001 * |
| Summ vs. Spri | <0.0001 * | Summ vs. Spri | 0.001 * |
| Autu vs. Wint | <0.0001 * | Autu vs. Wint | <0.0001 * |
| Autu vs. Spri | <0.0001 * | Autu vs. Spri | 0.009 * |
| Wint vs. Spri | 0.011 * | Wint vs. Spri | 0.014 * |
| TS Wastewater Treatment Plant | AL Wastewater Treatment Plant | ||
|---|---|---|---|
| Seasons | p values | Seasons | p values |
| Summ vs. Autu | 0.108 * | Summ vs. Autu | 0.055 * |
| Summ vs. Wint | 0.001 * | Summ vs. Wint | <0.0001 * |
| Summ vs. Spri | 0.003 * | Summ vs. Spri | <0.0001 * |
| Autu vs. Wint | 0.013 * | Autu vs. Wint | 0.002 * |
| Autu vs. Spri | 0.37 | Autu vs. Spri | 0.006 * |
| Wint vs. Spri | 0.516 | Wint vs. Spri | 0.376 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Osunla, A.C.; Abioye, O.E.; Okoh, A.I. Distribution and Public Health Significance of Vibrio Pathogens Recovered from Selected Treated Effluents in the Eastern Cape Province, South Africa. Water 2021, 13, 932. https://doi.org/10.3390/w13070932
Osunla AC, Abioye OE, Okoh AI. Distribution and Public Health Significance of Vibrio Pathogens Recovered from Selected Treated Effluents in the Eastern Cape Province, South Africa. Water. 2021; 13(7):932. https://doi.org/10.3390/w13070932
Chicago/Turabian StyleOsunla, Ayodeji C., Oluwatayo E. Abioye, and Anthony I. Okoh. 2021. "Distribution and Public Health Significance of Vibrio Pathogens Recovered from Selected Treated Effluents in the Eastern Cape Province, South Africa" Water 13, no. 7: 932. https://doi.org/10.3390/w13070932