
Aquatic Invertebrate Community Resilience and Recovery in Response to a Supra-Seasonal Drought in an Ecologically Important Naturally Saline Lake

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
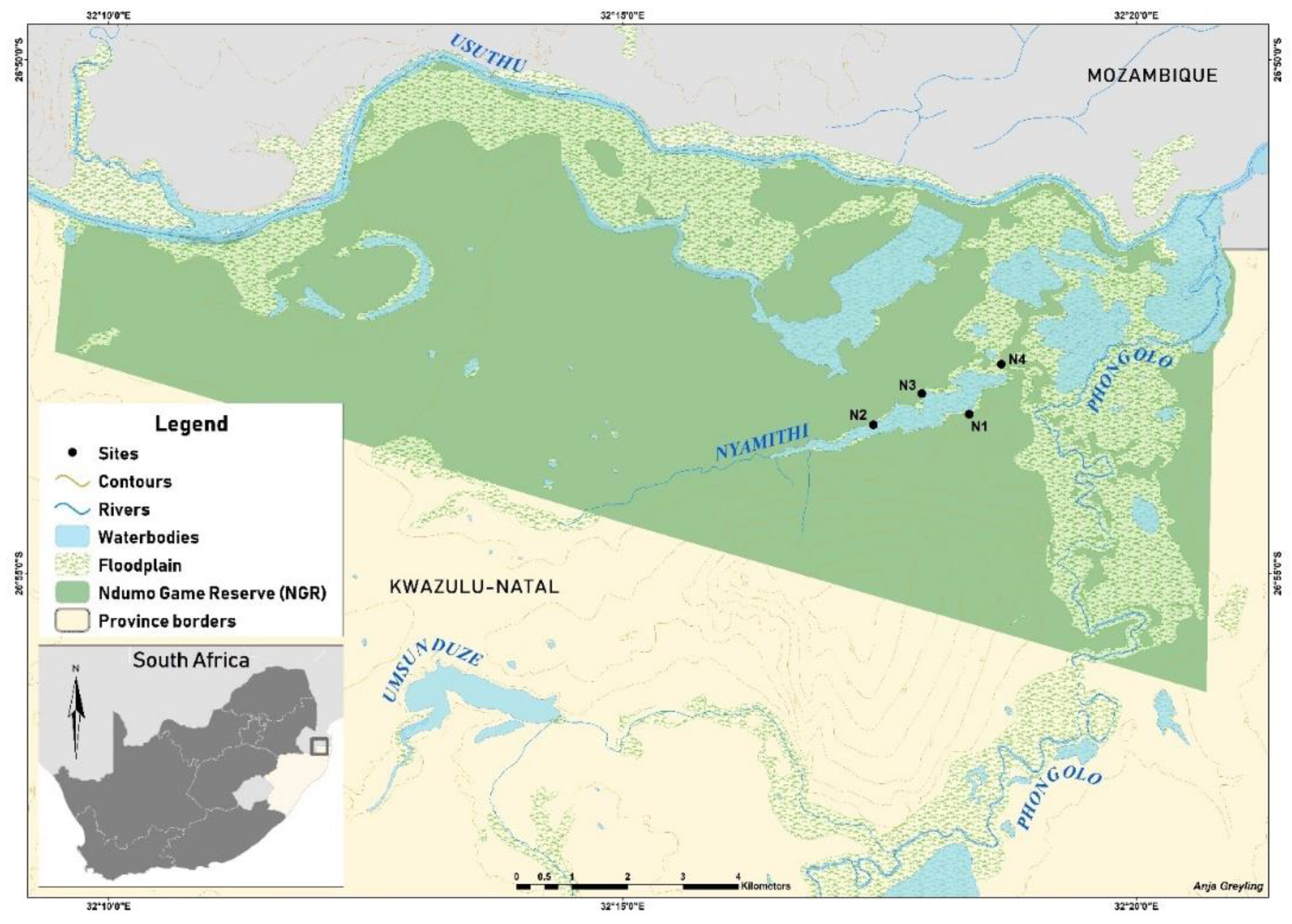
2.1. Study Area
2.2. Field Sampling
2.3. Laboratory Analysis
2.4. Data Analysis
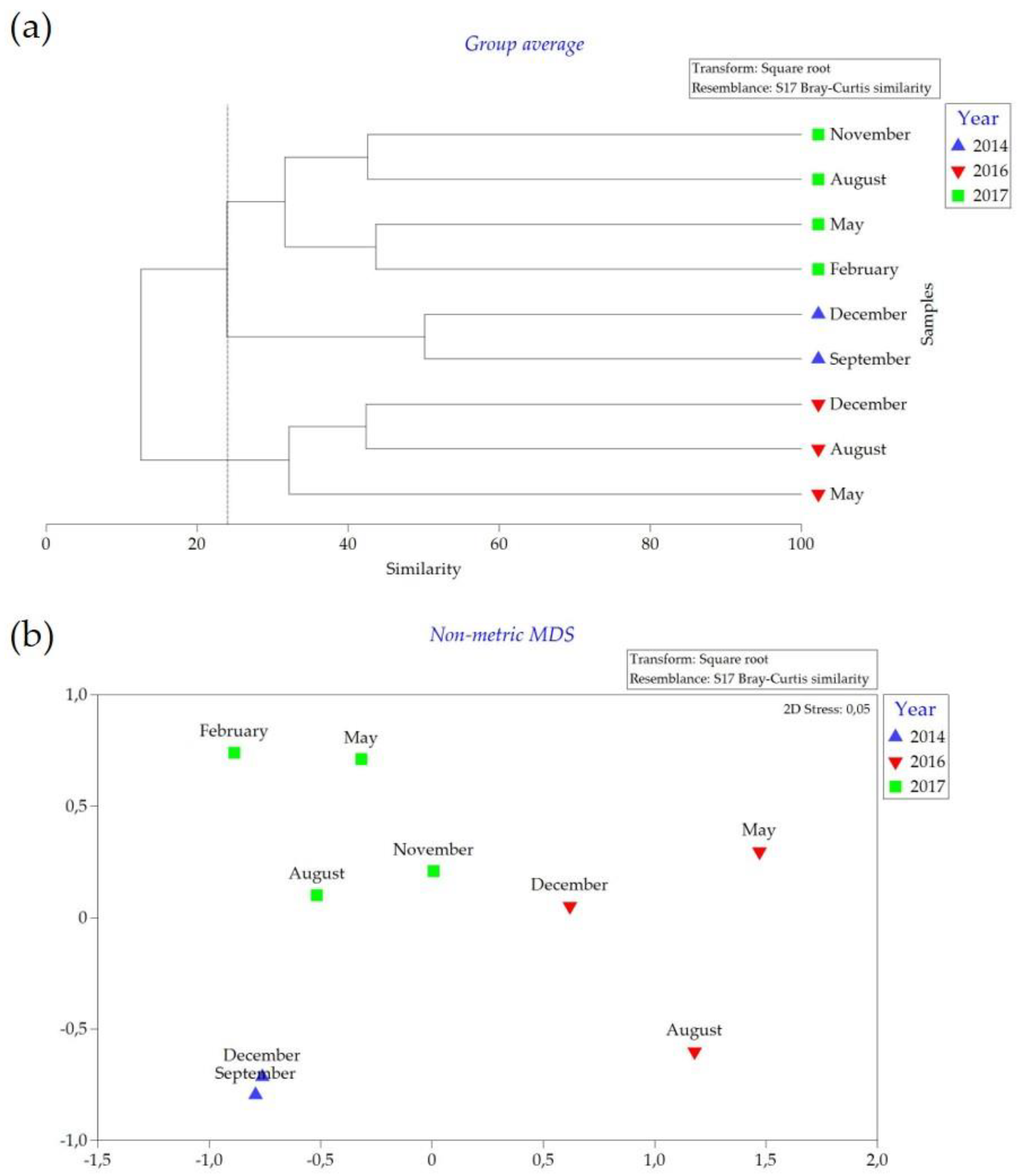
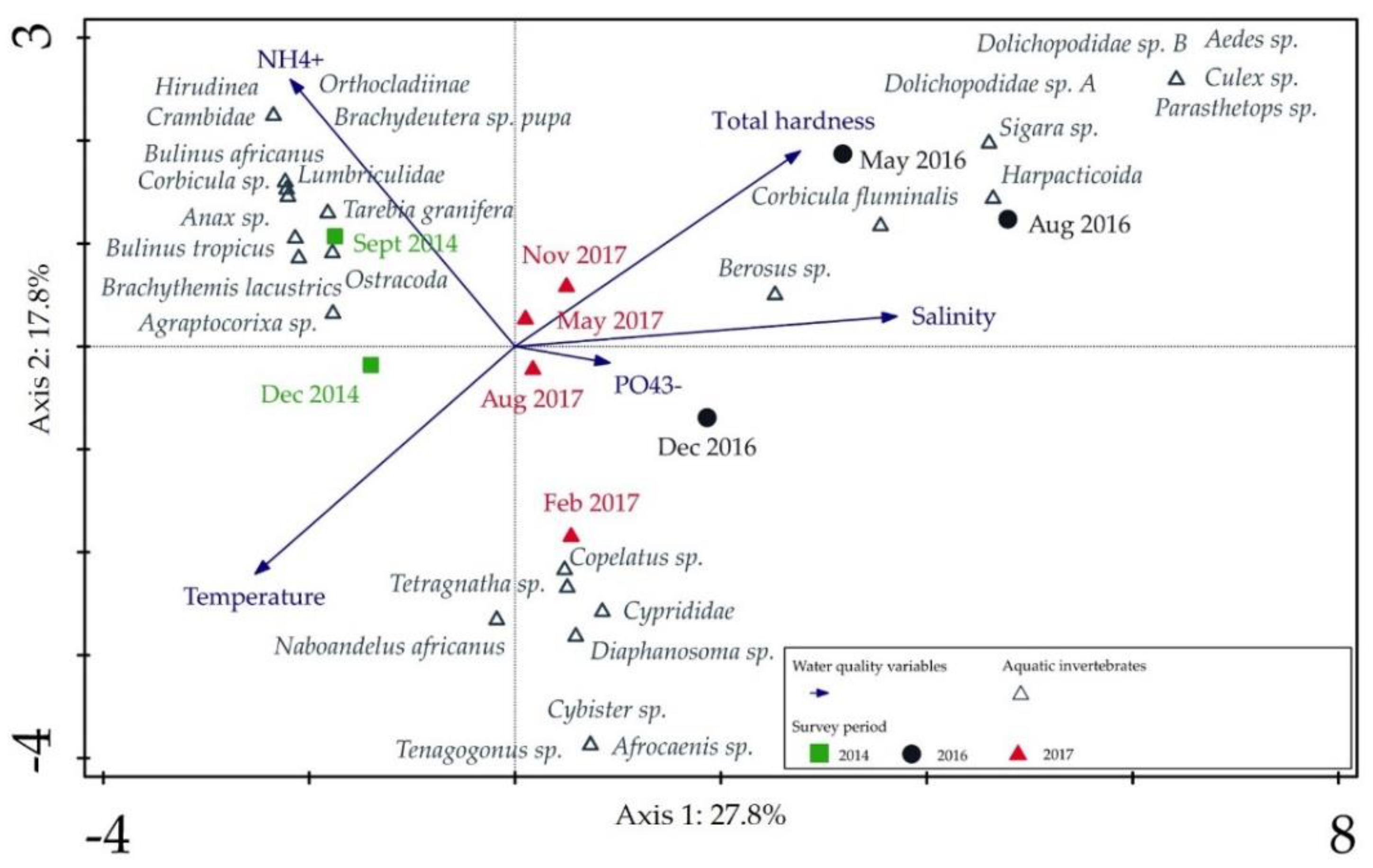
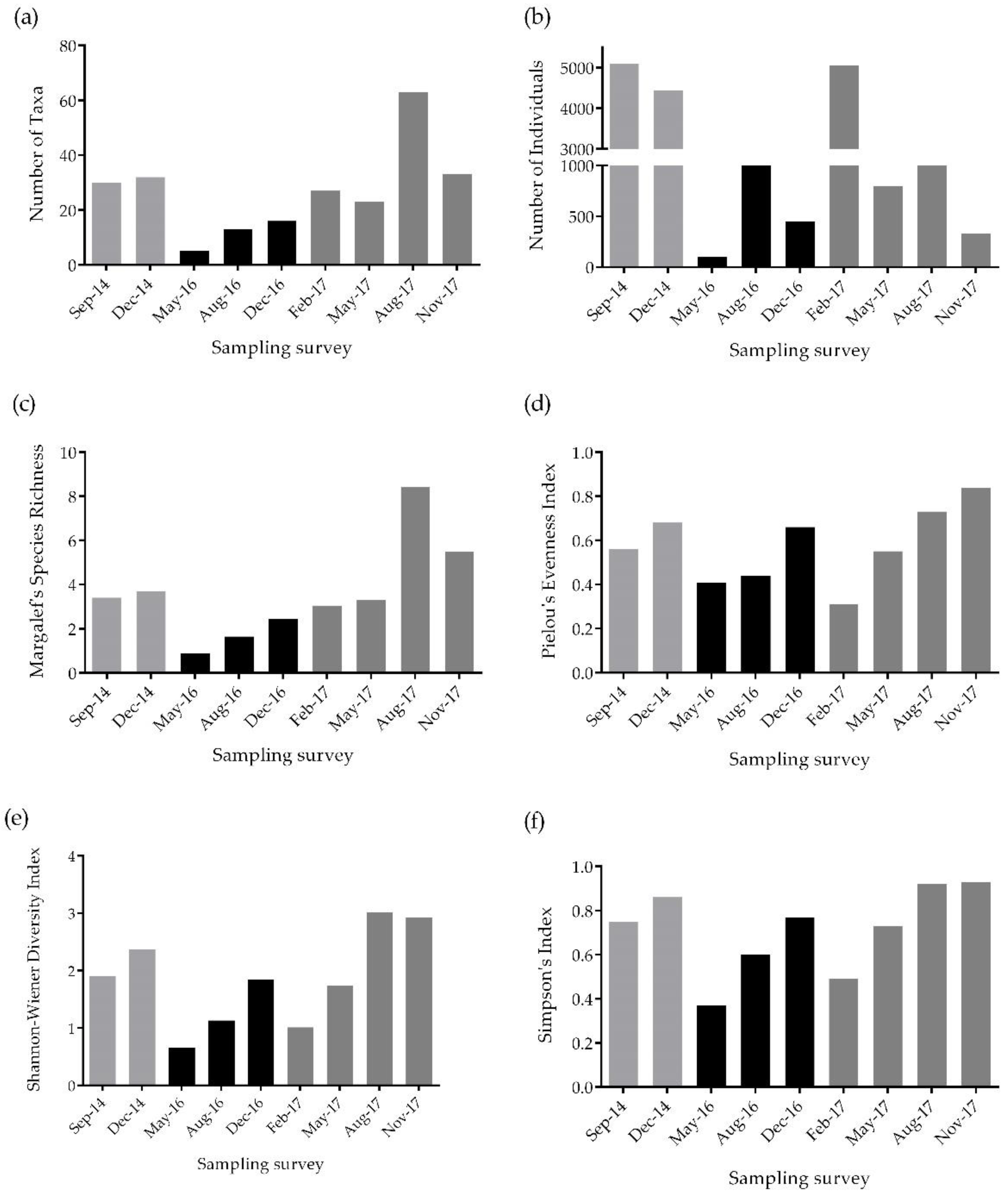
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Williams, W.D. Environmental threats to salt lakes and the likely status of inland saline ecosystems in 2025. Environ. Conserv. 2002, 29, 154–167. [Google Scholar] [CrossRef] [Green Version]
- Messager, M.L.; Lehner, B.; Grill, G.; Nedeva, I.; Schmitt, O. Estimating the volume and age of water stored in global lakes using a geo-statistical approach. Nat. Commun. 2016, 7, 13603. [Google Scholar] [CrossRef]
- Wurtsbaugh, W.A.; Miller, C.; Null, S.E.; DeRose, R.J.; Wilcock, P.; Hahnenberger, M.; Howe, F.; Moore, F. Decline of the world’s saline lakes. Nat. Geosci. 2017, 10, 816–821. [Google Scholar] [CrossRef]
- Zimmerman-Timm, H. Salinisation of inland waters. In Water Uses and Human Impacts on the Water Budget; Lozan, J., Grssl, H., Hupfer, P., Menzel, L., Schönwiese, C., Eds.; Verlag Wissenschaftliche Auswertungen/GEO: Hamburg, Germany, 2007; pp. 133–136. [Google Scholar]
- Williams, W.D.; Kokkinn, M.J. The biogeographical affinities of the fauna in episodically filled salt lakes: A study of Lake Eyre South, Australia. Hydrobiologia 1988, 158, 227–236. [Google Scholar] [CrossRef]
- Karagianni, A.; Stamou, G.; Katsiapi, M.; Polykarpou, P.; Dörflinger, G.; Michaloudi, E. Zooplankton communities in Mediterranean temporary lakes: The case of saline lakes in Cyprus. Ann. Limnol. 2018, 54, 14. [Google Scholar] [CrossRef]
- Williams, W.D. Conservation of salt lakes. Hydrobiologia 1993, 267, 291–306. [Google Scholar] [CrossRef]
- Williams, W.D. Salinity as a determinant of the structure of biological communities in salt lakes. Hydrobiologia 1998, 381, 191–201. [Google Scholar] [CrossRef]
- Moore, J.N. Recent desiccation of western Great Basin saline lakes: Lessons from Lake Abert, Oregon, USA. Sci. Total Environ. 2016, 554, 142–154. [Google Scholar] [CrossRef] [PubMed]
- Jellison, R.; Williams, W.D.; Timms, B.; Alcocer, J.; Aladin, N.V. Salt lakes: Values, threats and future. In Aquatic Ecosyst.; Poluninm, N., Ed.; Cambridge University Press: Cambridge, UK, 2008; pp. 94–110. [Google Scholar]
- Frank, M.G.; Conover, M.R. Weather and prey availability affect the timing of fall migration of Eared Grebes (Podiceps nigricollis) from Great Salt Lake. Wilson J. Ornithol. 2017, 129, 98–111. [Google Scholar] [CrossRef]
- Acosta, A.A.; Netherlands, E.C.; Retief, F.; de Necker, L.; du Preez, L.; Truter, M.; Alberts, R.; Gerber, R.; Wepener, V.; Malherbe, W.; et al. Conserving freshwater biodiversity in an African subtropical wetland: South Africa’s lower Phongolo river and floodplain. In Managing Wildlife in a Changing World; Kideghesho, J., Ed.; IntechOpen: London, UK, 2020. [Google Scholar]
- Zheng, M. On salinology. Hydrobiologia 2001, 466, 339–347. [Google Scholar]
- Shadrin, N.V. The alternative saline lake ecosystem states and adaptive environmental management. J. Oeceanol. Limnol. 2018, 36, 2010–2017. [Google Scholar] [CrossRef]
- Williams, W.D. What future for salt lakes. Environ. Sci. Policy 1996, 38, 12–39. [Google Scholar]
- Jeppesen, E.; Brucet, S.; Naselli-Flores, L.; Papastergiadou, E.; Stefanidis, K.; Nõges, T.; Nõges, P.; Attayde, J.L.; Zohary, T.; Coppens, J.; et al. Ecological impacts of global warming and water abstraction on lakes and reservoirs due to changes in water level and related changes in salinity. Hydrobiologia 2015, 750, 201–227. [Google Scholar] [CrossRef]
- Waterkeyn, A.; Grillas, P.; Vanschoenwinkel, B.; Brendonck, L. Invertebrate community patterns in Mediterranean temporary wetlands along hydroperiod and salinity gradients. Freshw. Biol. 2008, 53, 1808–1822. [Google Scholar] [CrossRef]
- Waterkeyn, A.; Vanschoenwinkel, B.; Grillas, P.; Brendonck, L. Effect of salinity on seasonal community patterns of Mediterranean temporary wetland crustaceans: A mesocosm study. Limnol. Oceanogr. 2010 55, 1712–1722. [CrossRef]
- Afonina, E.Y.; Tashlykova, N.A. Plankton community and the relationship with the environment in saline lakes of Onon-Torey plain, Northeastern Mongolia. Saudi J. Biol. Sci. 2018, 25, 399–408. [Google Scholar] [CrossRef]
- Brown, P. Salinity tolerance of Artemia and Ephydra: Uncertainty and discrepancies. In Utah Division of Wildlife Resources Great Salt Lake Ecosystem Program; Salt Lake City, UT, USA, Unpublished report; 2010. [Google Scholar]
- Senner, N.R.; Moore, J.N.; Seager, S.T.; Dougill, S.; Kreuz, K.; Senner, S.E. A salt lake under stress: Relationships among birds, water levels, and invertebrates at a Great Basin saline lake. Biol. Conserv. 2018, 220, 320–329. [Google Scholar] [CrossRef]
- Timms, B.V. Salt lakes in Australia: Present problems and prognosis for the future. Hydrobiologia 2005, 552, 1–15. [Google Scholar] [CrossRef]
- Ginatullina, E.; Atwell, L.; Saito, L. Resilience and resistance of zooplankton communities to drought-induced salinity in freshwater and saline lakes of Central Asia. J. Arid Environ. 2017, 144, 1–11. [Google Scholar] [CrossRef]
- Rouault, M.; Richard, Y. Spatial extension and intensity of droughts since 1922 in South Africa. Water SA 2003, 29, 489–500. [Google Scholar]
- Bogan, M.T.; Boersma, K.S.; Lytle, D.A. Resistance and resilience of invertebrate communities to seasonal and supraseasonal drought in arid--land headwater streams. Freshw. Biol. 2015, 60, 2547–2558. [Google Scholar] [CrossRef]
- Lake, P.S. Disturbance, patchiness, and diversity in streams: Journal of the North American. Benthol. Soc. 2000, 19, 573–592. [Google Scholar] [CrossRef] [Green Version]
- Isbell, F.; Craven, D.; Connolly, J.; Loreau, M.; Schmid, B.; Beierkuhnlein, C.; Bezemer, T.M.; Bonin, C.; Bruelheide, H.; De Luca, E.; et al. Biodiversity increases the resistance of ecosystem productivity to climate extremes. Nature 2015, 526, 574–577. [Google Scholar] [CrossRef]
- Bogan, M.T.; Hwan, J.L.; Cervantes-Yoshida, K.; Ponce, J.; Carlson, S.M. Aquatic invertebrate communities exhibit both resistance and resilience to seasonal drying in an intermittent coastal stream. Hydrobiologia 2017, 799, 123–133. [Google Scholar] [CrossRef]
- Jeppesen, E.; Appelt, M.; Hastrup, K.; Grønnow, B.; Mosbech, A.; Smol, J.P.; Davidson, T.A. Living in an oasis: Rapid transformations, resilience, and resistance in the North Water Area societies and ecosystems. Ambio 2018, 47, 296–309. [Google Scholar] [CrossRef] [Green Version]
- Seidl, R.; Rammer, W.; Spies, T.A. Disturbance legacies increase the resilience of forest ecosystem structure, composition, and functioning. Ecol. Appl 2014, 24, 2063–2077. [Google Scholar] [CrossRef] [Green Version]
- James, K.R.; Cant, B.; Ryan, T. Responses of freshwater biota to rising salinity levels and implications for saline water management: A review. Aust. J. Bot. 2003, 51, 703–713. [Google Scholar] [CrossRef]
- Fritz, K.M.; Dodds, W.K. Resistance and resilience of macroinvertebrate assemblages to drying and flood in a tallgrass prairie stream system. Hydrobiologia 2004, 527, 99–112. [Google Scholar] [CrossRef]
- Pimm, S.L. The complexity and stability of ecosystems. Nature 1984, 307, 321–326. [Google Scholar] [CrossRef]
- Beketov, M.A.; Schäfer, R.B.; Marwitz, A.; Paschke, A.; Liess, M. Long-term stream invertebrate community alterations induced by the insecticide thiacloprid: Effect concentrations and recovery dynamics. Sci. Total Environ. 2008, 405, 96–108. [Google Scholar] [CrossRef] [PubMed]
- Brock, M.A.; Nielsen, D.L.; Shiel, R.J.; Green, J.D.; Langley, J.D. Drought and aquatic community resilience: The role of eggs and seeds in sediments of temporary wetlands. Freshw. Biol. 2003, 48, 1207–1218. [Google Scholar] [CrossRef]
- Bogan, M.T.; Lytle, D.A. Severe drought drives novel community trajectories in desert stream pools. Freshw. Biol 2011, 56, 2070–2081. [Google Scholar] [CrossRef]
- Boulton, A.J. Parallels and contrasts in the effects of drought on stream macroinvertebrate assemblages. Freshw. Biol 2003, 48, 1173–1185. [Google Scholar] [CrossRef] [Green Version]
- Boulton, A.J.; Lake, P.S. Effects of drought on stream insects and its ecological consequences. In Aquatic Insects: Challenges to Populations: Proceedings of the Royal Entomological Society’s 24th Symposium; Lancaster, J., Briers, R.A., Eds.; Cromwell Press: Trowbridge, UK, 2008; pp. 81–102. [Google Scholar]
- Chadd, R.P.; England, J.A.; Constable, D.; Dunbar, M.J.; Extence, C.A.; Leeming, D.J.; Murray-Bligh, J.A.; Wood, P.J. An index to track the ecological effects of drought development and recovery on riverine invertebrate communities. Ecol. Indic. 2017, 82, 344–356. [Google Scholar] [CrossRef] [Green Version]
- Lake, P.S. Ecological effects of perturbation by drought in flowing waters. Freshw. Biol. 2003, 48, 1161–1172. [Google Scholar] [CrossRef]
- Brendonck, L.; de Meester, L. Egg banks in freshwater zooplankton: Evolutionary and ecological archives in the sediment. Hydrobiologia 2003, 491, 65–84. [Google Scholar] [CrossRef]
- Nielsen, D.L.; Brock, M.A.; Rees, G.N.; Baldwin, D.S. Effects of increasing salinity on freshwater ecosystems in Australia. Aust. J. Bot. 2003, 51, 655–665. [Google Scholar] [CrossRef]
- Hammer, U.T. Saline Lake Ecosystems of the World; Dr W. Junk Publishers: Dordrecht, The Netherlands, 1986. [Google Scholar]
- Herbst, D.B. Gradients of salinity stress, environmental stability and water chemistry as a templet for defining habitat types and physiological strategies in inland salt waters. In Saline Lakes; Melack, J.M., Jellison, R., Herbst, D.B., Eds.; Springer: Dordrecht, The Netherlands, 2001; pp. 209–219. [Google Scholar]
- Herbst, D.B. Salinity controls on trophic interactions among invertebrates and algae of solar evaporation ponds in the Mojave Desert and relation to shorebird foraging and selenium risk. Wetlands 2006, 26, 475–485. [Google Scholar] [CrossRef]
- Downing, A.L.; Leibold, M.A. Species richness facilitates ecosystem resilience in aquatic food webs. Freshw. Biol 2010, 55, 2123–2137. [Google Scholar] [CrossRef]
- Khan, F.R.; Keller, W.; Yan, N.D.; Welsh, P.G.; Wood, C.M.; McGeer, J.C. Application of biological ligand and toxic unit modeling approaches to predict improvements in zooplankton species richness in smelter-damaged lakes near Sudbury, Ontario. Environ. Sci. Technol 2012, 46, 1641–1649. [Google Scholar] [CrossRef] [PubMed]
- Seaman, M.T.; Ashton, P.J.; Williams, W.D. Inland salt waters of southern Africa. Hydrobiologia 1991, 210, 75–91. [Google Scholar] [CrossRef]
- Heeg, J.; Breen, C.M. Man and the Pongolo floodplain. In National Scientific Programmes Unit; Council for Scientific and Industrial Research: Pretoria, South Africa, 1982. [Google Scholar]
- South African Wetlands Conservation Programme (SA Wetlands Conservation Programme). Ndumo Game Reserve, South. Africa, Information sheet. South. African Wetlands Conservation Programme; Document No: 24/21/3/3/3/15; Department of Environmental Affairs and Tourism: Pretoria, South Africa, 1996. [Google Scholar]
- De Necker, L.; Neswiswi, T.; Greenfield, R.; van Vuren, J.; Brendonck, L.; Wepener, V.; Smit, N. Long-term water quality patterns of a flow regulated tropical lowland river. Water 2020, 12, 37. [Google Scholar] [CrossRef] [Green Version]
- Baudoin, M.A.; Vogel, C.; Nortje, K.; Naik, M. Living with drought in South Africa: Lessons learnt from the recent El Niño drought period. Int. J. Disaster Risk Reduct. 2017, 23, 128–137. [Google Scholar] [CrossRef]
- Abbas, H.A.; Bond, W.J.; Midgley, J.J. The worst drought in 50 years in a South African savannah: Limited impact on vegetation. Afr. J. Ecol. 2019, 57, 490–499. [Google Scholar] [CrossRef]
- Welicky, R.L.; De Swardt, J.; Gerber, R.; Netherlands, E.C.; Smit, N.J. Drought-associated absence of alien invasive anchorworm, Lernaea cyprinacea (Copepoda: Lernaeidae), is related to changes in fish health. Int. J. Parasitol. Parasites Wildl. 2017, 6, 430–438. [Google Scholar] [CrossRef]
- Ndumo Game Reserve: Integrated Management Plan: 2009–2013 (55); (Version 1.0); Ezemvelo KZN Wildlife: Pietermaritzburg, South Africa, 2009.
- Smit, N.J.; Vlok, W.; van Vuren, J.H.J.; du Preez, L.; van Eeden, E.; O’Brien, G.C.; Wepener, V. Socio-Ecological System Management of the Lower Phongolo River and Floodplain Using Relative Risk Methodology; WRC Report No 2185/1/16; Water Research Commission: Pretoria, South Africa, 2016. [Google Scholar]
- Whittington, M.; Malan, G.; Panagos, M.D. Trends in waterbird diversity at Banzi, Shokwe and Nyamithi pans, Ndumo game reserve, South Africa. Ostrich 2013, 84, 47–61. [Google Scholar] [CrossRef]
- Calverley, P.M.; Downs, C.T. Population status of Nile crocodiles in Ndumo Game Reserve, KwaZulu-Natal, South Africa (1971–2012). Herpetologica 2014, 70, 417–425. [Google Scholar] [CrossRef]
- Batzer, D.P.; Wissinger, S.A. Ecology of insect communities in nontidal wetlands. Annu. Rev. Entomol 1996, 41, 75–100. [Google Scholar] [CrossRef] [PubMed]
- Tokeshi, M.; Arakaki, S. Habitat complexity in aquatic systems: Fractals and beyond. Hydrobiologia 2012, 685, 27–47. [Google Scholar] [CrossRef]
- Harabiš, F.; Tichanek, F.; Tropek, R. Dragonflies of freshwater pools in lignite spoil heaps: Restoration management, habitat structure and conservation value. Ecol. Eng. 2013, 55, 51–61. [Google Scholar] [CrossRef]
- Collier, K.J.; Probert, P.K.; Jeffries, M. Conservation of aquatic invertebrates: Concerns, challenges and conundrums. Aquat. Coserv. Mar. Freshw. Ecosyst. 2016, 26, 817–837. [Google Scholar] [CrossRef]
- Merck KGaA (Merck Kommanditgesellschaft auf Aktien). Spectroquant Pharo 300 Operation Manual; Merck KGaA: Darmstadt, Germany, 2014. [Google Scholar]
- Day, J.A.; Stewart, B.A.; de Moor, I.J.; Louw, A.E. Guides to the freshwater invertebrates of Southern Africa. Volume 2: Crustacea I (Notostraca, Anostraca, Conchostraca and Cladocera); WRC Report TT 121/00; Water Research Commission: Pretoria, South Africa, 1999. [Google Scholar]
- Day, J.A.; de Moor, I.J.; Stewart, B.A.; Louw, A.E. Guides to the Freshwater Invertebrates of Southern Africa. Volume 3: Crustacea II (Ostracoda, Copepoda and Branchiura); WRC Report TT 148/01; Water Research Commission: Pretoria, South Africa, 2001. [Google Scholar]
- Day, J.A.; Stewart, B.A.; de Moor, I.J.; Louw, A.E. Guides to the Freshwater invertebrates of Southern Africa. Volume 4: Crustacea III (Bathynellacea, Amphipoda, Isopoda, Spelaeogriphacea, Tanaidacea, Decapoda); WRC Report TT 141/01; Water Research Commission: Pretoria, South Africa, 2001. [Google Scholar]
- Day, J.A.; de Moor, I.J. Guides to Freshwater Invertebrates of Southern Africa. Volume 5: Non–Arthropods (The Protozoans, Porifera, Cnidaria, Platyhelminthes, Nemertea, Rotifera, Nematoda, Nematomorpha, Gastrotrichia, Bryozoa, Tardigrada, Polychaeta, Oligochaeta and Hirudinae); WRC Report TT 167/02; Water Research Commission: Pretoria, South Africa, 2002. [Google Scholar]
- Day, J.A.; de Moor, I.J. Guides to the Freshwater Invertebrates of Southern Africa. Volume 6: Arachnida and Mollusca (Araneae, Water Mites and Mollusca); WRC Report TT 182/02; Water Research Commission: Pretoria, South Africa, 2002. [Google Scholar]
- Day, J.A.; Harrison, A.D.; de Moor, I.J. Guides to the Freshwater Invertebrates of Southern Africa. Volume 9: Diptera; WRC Report TT 201/02; Water Research Commission: Pretoria, South Africa, 2003. [Google Scholar]
- De Moor, I.J.; Day, J.A.; de Moor, F.C. Guides to the Freshwater Invertebrates of Southern Africa. Volume 7: Insecta I (Ephemeroptera, Odonata and Plecoptera); WRC Report TT 207/03; Water Research Commission: Pretoria, South Africa, 2003. [Google Scholar]
- De Moor, I.J.; Day, J.A.; de Moor, F.C. Guides to the Freshwater Invertebrates of Southern Africa. Volume 8: Insecta II (Hemiptera, Megaloptera, Neuroptera, Trichoptera and Lepidoptera); WRC Report TT 214/03; Water Research Commission: Pretoria, South Africa, 2003. [Google Scholar]
- Stals, R.; de Moor, I.J. Guides to the Freshwater Invertebrates of Southern Africa Volume 10: Coleoptera; WRC Report TT 320/07; Water Research Commission: Pretoria, South Africa, 2008. [Google Scholar]
- Clarke, K.R.; Warwick, R.M. Change in marine communities: An approach to statistical analysis and interpretation. In Manual for the PRIMER Statistical Programme; Natural Environment Research Council: Plymouth, UK, 2001. [Google Scholar]
- Quinn, G.P.; Keough, M.J. Experimental Design and Data Analysis for Biologists; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Margalef, R. Perspectives in Ecological Theory; University of Chicago Press: Chicago, IL, USA, 1968. [Google Scholar]
- Pielou, E.E. Mathematical Ecology; Wiley Publishing: New York, NY, USA, 1977. [Google Scholar]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1963. [Google Scholar]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. Getting Started with PRIMER v7; PRIMER-E: Plymouth, UK, 2015. [Google Scholar]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- RStudio Team. RStudio: Integrated Development for R; RStudio, Inc.: Boston, MA, USA, 2019. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer: New York, NY, USA, 2009. [Google Scholar]
- Šmilauer, P.; Lepš, J. Multivariate Analysis of Ecological Data using Canoco 5; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- De Cáceras, M.; Jansen, F.; de Cáceras, M.M. Package Indicspecies: Relationship Between Species and Groups of Sites; R Package Version 1.7.6; Boston, MA, USA, 2016; Available online: https://mran.microsoft.com/snapshot/2018-04-23/web/packages/indicspecies/index.html (accessed on 15 March 2021).
- Boersma, K.; Bogan, M.T.; Henrichs, B.A.; Lytle, D.A. Invertebrate assemblages of pools in arid-land streams have high functional redundancy and are resistant to severe drying. Freshw. Biol. 2014, 59, 491–501. [Google Scholar] [CrossRef]
- Hart, B.T.; Bailey, P.; Edwards, R.; Hortle, K.; James, K.; McMahon, A.; Meredith, C.; Swadling, K. A review of the salt sensitivity of the Australian freshwater biota. Hydrobiologia 1991, 21, 105–144. [Google Scholar] [CrossRef]
- Aspin, T.W.; Khamis, K.; Matthews, T.J.; Milner, A.M.; O’Callaghan, M.J.; Trimmer, M.; Woodward, G.; Ledger, M.E. Extreme drought pushes stream invertebrate communities over functional thresholds. Glob. Chang. Biol. 2019, 25, 230–244. [Google Scholar] [CrossRef] [PubMed]
- Weyl, O.L.F.; Ellender, B.R.; Wasserman, R.J.; Truter, M.; Dalu, T.; Zengeya, T.A.; Smit, N.J. Alien freshwater fauna in South Africa. In Biological Invasions in South Africa, Invading Nature—Springer Series in Invasion Ecology; van Wilgen, B.W., Measey, J., Richardson, D.M., Wilson, J.R., Zengeya, T.A., Eds.; Springer: Cham, Switzerland, 2020; Volume 14, pp. 153–183. [Google Scholar]
- Miranda, N.A.F.; Perissinotto, R.; Appleton, C.C. Salinity and temperature tolerance of the invasive freshwater gastropod Tarebia granifera. S. Afr. J. Sci. 2010, 106, 1–7. [Google Scholar] [CrossRef]
- Appleton, C.C.; Forbes, A.T.; Demetriades, N.T. The occurrence, bionomics and potential impacts of the invasive freshwater snail Tarebia granifera (Lamarck, 1822) (Gastropoda: Thiaridae) in South Africa. Zool. Meded. 2009, 83, 525–536. [Google Scholar]
- Darwall, W.; Smith, K.; Allen, D.; Holland, R.; Harrison, I.; Brooks, E. The diversity of life in African freshwaters: Underwater, under threat. In An Analysis of the Status and Distribution of Freshwater Species Throughout Mainland Africa; The World Conservation Union: Gland, Switzerland, 2011. [Google Scholar]
- Castillo, A.M.; Sharpe, D.M.; Ghalambor, C.K.; De León, L.F. Exploring the effects of salinization on trophic diversity in freshwater ecosystems: A quantitative review. Hydrobiologia 2018, 807, 1–17. [Google Scholar] [CrossRef]
- Williams, W.D. Inland salt lakes: An introduction. Hydrobiologia 1981, 81, 1–14. [Google Scholar] [CrossRef]
- Velasco, J.; Millán, A.; Hernández, J.; Gutiérrez, C.; Abellán, P.; Sánchez, D.; Ruiz, M. Response of biotic communities to salinity changes in a Mediterranean hypersaline stream. Saline Syst. 2006, 2, 1–15. [Google Scholar] [CrossRef] [Green Version]
- McLachlan, A.J.; Cantrell, M.A. Survival strategies in tropical rain pools. Oecologia 1980, 47, 344–351. [Google Scholar] [CrossRef] [PubMed]
- Samways, M.J. Dragonflies and Damselflies of South Africa; Pensoft Publishers: Sofia, Bulgaria, 2008. [Google Scholar]
- Bird, M.S.; Mlambo, M.C.; Wasserman, R.J.; Dalu, T.; Holland, A.J.; Day, J.A.; Villet, M.H.; Bilton, D.T.; Barber-James, H.M.; Brendonck, L. Deeper knowledge of shallow waters: Reviewing the invertebrate fauna of southern African temporary wetlands. Hydrobiologia 2019, 827, 89–121. [Google Scholar] [CrossRef]
- Martens, K.; Schön, I.; Meisch, C.; Horne, D.J. Global diversity of ostracods (Ostracoda, Crustacea) in freshwater. In Freshwater Animal Diversity Assessment; Balian, E.V., Lévêque, C., Segers, H., Martens, K., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 185–193. [Google Scholar]
- Smith, A.J.; Delorme, L.D. Chapter 19: Ostracoda. In Ecology and Classification of North American Freshwater Invertebrates; Thorp, J.H., Covich, A.P., Eds.; Academic Press: London, UK, 2010; pp. 725–771. [Google Scholar]
- Coull, B.C. Marine Flora and Fauna of the Northeastern United States: Copepoda, Harpacticoida; US Department of Commerce, National Oceanic and Atmospheric Administration, National Marine Fisheries Service: Silver Spring, MD, USA, 1977; Volume 399. [Google Scholar]
- Dimante-Deimantovica, I.; Jensen, T.C.; Walseng, B. Freshwater harpacticoids (Crustacea: Copepoda: Harpacticoida) in Norway–a comprehensive contribution from GO Sars, and a provisional checklist. J. Nat. Hist. 2016, 50, 1773–1795. [Google Scholar] [CrossRef]
- Ruhí, A.; Boix, D.; Sala, J.; Gascón, S.; Quintana, X.D. Spatial and temporal patterns of pioneer macrofauna in recently created ponds: Taxonomic functional approaches. Hydrobiologia 2009, 634, 137–151. [Google Scholar] [CrossRef]
- Brendonck, L.; Pinceel, T.; Ortells, R. Dormancy and dispersal as mediators of zooplankton population and community dynamics along a hydrological disturbance gradient in inland temporary pools. Hydrobiologia 2017, 796, 201–222. [Google Scholar] [CrossRef]
- Wiggins, G.B.; Mackay, R.J.; Smith, I.M. Evolutionary and ecological strategies of animals in annual temporary pools. Arch. Hydrobiol. 1980, 58, 97–206. [Google Scholar]
- Warnock, N.; Page, G.W.; Ruhlen, T.D.; Nur, N.; Takekawa, J.Y.; Hanson, J.T. Management and conservation of San Francisco Bay salt ponds: Effects of pond salinity, area, tide, and season on Pacific Flyway waterbirds. Waterbirds 2002, 25, 79–92. [Google Scholar]
- Anufriieva, E.V.; Shadrin, N.V. Extreme hydrological events destabilize aquatic ecosystems and open doors for alien species. Quat. Int. 2018, 475, 11–15. [Google Scholar] [CrossRef]
- Timpane-Padgham, B.L.; Beechie, T.; Klinger, T. A systematic review of ecological attributes that confer resilience to climate change in environmental restoration. PLoS ONE 2017, 12, e0173812. [Google Scholar] [CrossRef] [Green Version]
- Sjöstedt, J.; Langenheder, S.; Kritzberg, E.; Karlsson, C.M.; Lindström, E.S. Repeated disturbances affect functional but not compositional resistance and resilience in an aquatic bacterioplankton community. Environ. Microbiol. Rep. 2018, 10, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Brendonck, L.; Rogers, D.C.; Olesen, J.; Weeks, S.; Hoeh, W.R. Global diversity of large branchiopods (Crustacea: Branchiopoda) in freshwater. In Freshwater Animal Diversity Assessment; Balian, E.V., Lévêque, C., Segers, H., Martens, K., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 167–176. [Google Scholar]
- Zimmer, K.D.; Hanson, M.A.; Wrubleski, D.A. Invertebrates in permanent wetlands (long-hydroperiod marshes and shallow lakes). In Invertebrates in Freshwater Wetlands; Batzer, D., Boix, D., Eds.; Springer: Cham, Switzerland, 2016; pp. 251–286. [Google Scholar]
- Jeffries, M.J.; Epele, L.B.; Studinski, J.M.; Vad, C.F. Invertebrates in temporary wetland ponds of the temperate biomes. In Invertebrates in Freshwater Wetlands; Batzer, D., Boix, D., Eds.; Springer: Cham, Switzerland, 2016; pp. 105–139. [Google Scholar]
- Miranda, N.A.; Perissinotto, R. Benthic assemblages of wetlands invaded by Tarebia granifera (Lamarck, 1822) (Caenogastropoda: Thiaridae) in the iSimangaliso Wetland Park, South Africa. Molluscan Res. 2014, 34, 40–48. [Google Scholar] [CrossRef]
- Jones, R.W.; Hill, J.M.; Coetzee, J.A.; Avery, T.S.; Weyl, O.L.F.; Hill, M.P. The abundance of an invasive freshwater snail Tarebia granifera (Lamarck, 1822) in the Nseleni River, South Africa. Afr. J. Aquat. Sci. 2017, 42, 75–81. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Overall Similarity | Species | Average Abundance | Average Similarity | Percentage Contribution (%) | Cumulative Contribution (%) |
|---|---|---|---|---|---|---|
| 2014 | 50.12% | Ostracoda | 22.2 | 7.7 | 15.3 | 15.3 |
| Tarebia granifera | 32.8 | 7.6 | 15.1 | 30.4 | ||
| Bulinus tropicus | 17.9 | 6.9 | 13.7 | 44.1 | ||
| Bulinus natalensis | 22.9 | 5.1 | 10.3 | 54.4 | ||
| Agraptocorixa sp. A | 14.8 | 3.8 | 7.6 | 61.9 | ||
| Baetidae | 7.7 | 3 | 6.1 | 68 | ||
| Enallagma sp. | 7.4 | 2.3 | 4.5 | 72.6 | ||
| 2016 | 35.55% | Sigara sp. | 12.5 | 15.4 | 43.4 | 43.4 |
| Harpacticoida | 15.5 | 13.3 | 37.3 | 80.8 | ||
| 2017 | 35.43% | Ostracoda | 26.1 | 9.4 | 26.4 | 26.4 |
| Chironominae | 5.5 | 3.5 | 10 | 36.4 | ||
| Hyphydrus sp. A | 7.3 | 3 | 8.4 | 44.8 | ||
| Appasus sp. | 6.6 | 2.9 | 8.2 | 53 | ||
| Thermocyclops sp. | 12.4 | 2.7 | 7.7 | 60.7 | ||
| Pseudagrion sp. | 4 | 2.1 | 5.8 | 66.5 | ||
| Micronecta sp. | 3 | 1.4 | 4 | 70.5 |
| Water Quality Variable | September 2014 | December 2014 | May 2016 | August 2016 | December 2016 | February 2017 | May 2017 | August 2017 | November 2017 |
|---|---|---|---|---|---|---|---|---|---|
| Temperature (˚C) | 26.1 | 29.6 | 23.4 | 17.6 | 28.5 | 29.7 | 23.8 | 22.9 | 33.5 |
| pH | 8.8 | 7.9 | 8.5 | 9.3 | 8.2 | 8.4 | 7.1 | 8.3 | 8.8 |
| Dissolved Oxygen (mg L−1) | 5.8 | 8.3 | 6.0 | 10.3 | 5.5 | 4.5 | 1.9 | 5.3 | * |
| Oxygen Saturation (%) | 72.4 | 105.6 | 72.4 | 105.8 | 70.3 | 56 | 19.5 | 62.5 | * |
| Salinity (g L−1) | 3.9 | 2.3 | 9.8 | 11.5 | 10 | 5.4 | 2.3 | 4.9 | 7.1 |
| PO43− (mg L−1) | 0.1 | 0.2 | 0.5 | 0.7 | 0.3 | 0.1 | 0.03 | 4.0 | 0.1 |
| NO3− (mg L−1) | 2.2 | 0.4 | 0.9 | 1.5 | 1.0 | 1.1 | 1.1 | 8.2 | 0.8 |
| NO2− (mg L−1) | 0.04 | 0.04 | 0.02 | 0.01 | 0.04 | <0.02 | 0.02 | 0.3 | * |
| NH4+(mg L−1) | 2.2 | 0.1 | <0.05 | 0.1 | 0.2 | 0.2 | 0.1 | 0.5 | 0.1 |
| NH3 (mg L−1) | <0.01 | <0.01 | 0.02 | <0.01 | 0.04 | 0.01 | 0.01 | 0.4 | * |
| SO42−(mg L−1) | 313 | 243 | 975 | * | 536 | 611 | 45 | 2283 | 1320 |
| Cl−(mg L−1) | 731 | 567 | 300 | 3280 | 1068 | 193 | 356 | 1348 | 1720 |
| Total Hardness (mg L−1) | 284 | 234 | 1910 | 1130 | 623 | 195 | 188 | 915 | 977 |
| Mg2+ (mg L−1) | 79 | 51 | 1220 | 660 | 355 | 123 | 124 | 333 | 388 |
| Ca2+ (mg L−1) | 206 | 183 | 690 | 470 | 268 | 71 | 64 | 581 | 645 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Necker, L.; Brendonck, L.; van Vuren, J.; Wepener, V.; Smit, N.J. Aquatic Invertebrate Community Resilience and Recovery in Response to a Supra-Seasonal Drought in an Ecologically Important Naturally Saline Lake. Water 2021, 13, 948. https://doi.org/10.3390/w13070948
de Necker L, Brendonck L, van Vuren J, Wepener V, Smit NJ. Aquatic Invertebrate Community Resilience and Recovery in Response to a Supra-Seasonal Drought in an Ecologically Important Naturally Saline Lake. Water. 2021; 13(7):948. https://doi.org/10.3390/w13070948
Chicago/Turabian Stylede Necker, Lizaan, Luc Brendonck, Johan van Vuren, Victor Wepener, and Nico J. Smit. 2021. "Aquatic Invertebrate Community Resilience and Recovery in Response to a Supra-Seasonal Drought in an Ecologically Important Naturally Saline Lake" Water 13, no. 7: 948. https://doi.org/10.3390/w13070948