
Combined Effects of Summer Water Temperature and Current Velocity on the Distribution of a Cold-Water-Adapted Sculpin (Cottus nozawae)



Abstract
:1. Introduction
2. Materials and Methods
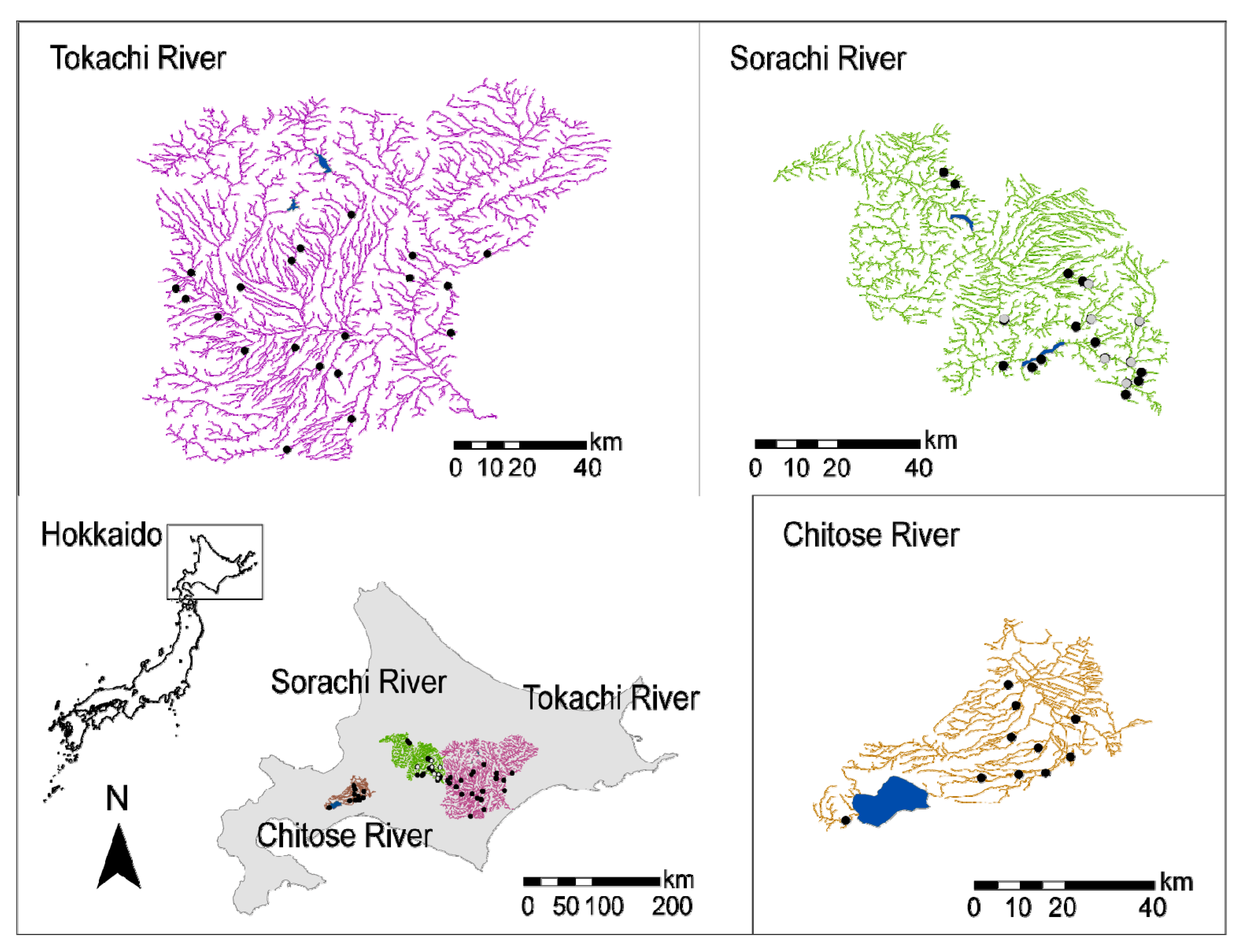
2.1. Abundance Survey
2.2. Occurrence Survey
2.3. Statistical Analyses
3. Results
3.1. Abundance Response to Thermal Gradients
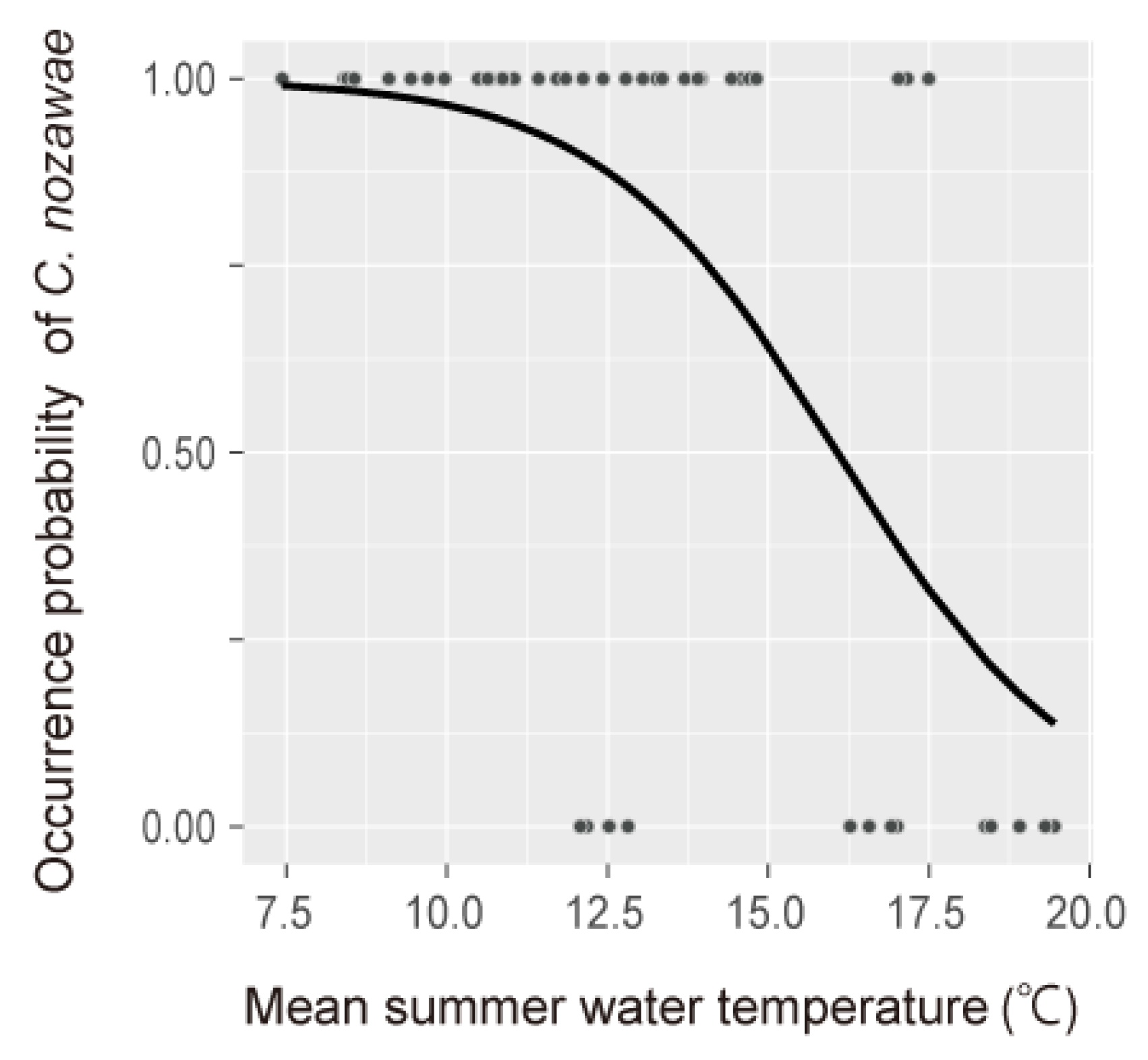
3.2. Occurrence Response to Thermal Gradients
4. Discussion
4.1. Effects of Summer Water Temperature
4.2. Combined Effects of Summer Water Temperature and Current Velocity
4.3. Management Implications
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Buisson, L.; Thuiller, W.; Lek, S.; Lim, P.U.Y.; Grenouillet, G. Climate Change Hastens the Turnover of Stream Fish Assemblages. Glob. Chang. Biol. 2008, 14, 2232–2248. [Google Scholar] [CrossRef]
- Isaak, D.J.; Luce, C.H.; Horan, D.L.; Chandler, G.L.; Wollrab, S.P.; Nagel, D.E. Global Warming of Salmon and Trout Rivers in the Northwestern US: Road to Ruin or Path Through Purgatory? Trans. Am. Fish. Soc. 2018, 147, 566–587. [Google Scholar] [CrossRef]
- Nakano, S.; Kitano, F.; Maekawa, K. Potential Fragmentation and Loss of Thermal Habitats for Charrs in the Japanese Archipelago Due to Climatic Warming. Freshwat. Biol. 1996, 36, 711–722. [Google Scholar] [CrossRef]
- Almodóvar, A.; Nicola, G.G.; Ayllón, D.; Elvira, B. Global Warming Threatens the Persistence of Mediterranean Brown Trout. Glob. Chang. Biol. 2012, 18, 1549–1560. [Google Scholar] [CrossRef] [Green Version]
- Eby, L.A.; Helmy, O.; Holsinger, L.M.; Young, M.K. Evidence of Climate-Induced Range Contractions in Bull Trout Salvelinus Confluentus in a Rocky Mountain Watershed, USA. PLoS ONE 2014, 9, e98812. [Google Scholar] [CrossRef]
- Atlas, W.I.; Seitz, K.M.; Jorgenson, J.W.N.; Millard-Martin, B.; Housty, W.G.; Ramos-Espinoza, D.; Burnett, N.J.; Reid, M.; Moore, J.W. Thermal Sensitivity and Flow-Mediated Migratory Delays Drive Climate Risk for Coastal Sockeye Salmon. FACETS 2021, 6, 71–89. [Google Scholar] [CrossRef]
- Rahel, F.J.; Nibbelink, N.P. Spatial Patterns in Relations Among Brown Trout (Salmo trutta) Distribution, Summer Air Temperature, and Stream Size in Rocky Mountain Streams. Can. J. Fish. Aquat. Sci. 1999, 56, 43–51. [Google Scholar] [CrossRef]
- Terui, A.; Miyazaki, Y. Combined Effects of Immigration Potential and Habitat Quality on Diadromous Fishes. Limnology 2017, 18, 121–129. [Google Scholar] [CrossRef]
- Jackson, M.C.; Loewen, C.J.G.; Vinebrooke, R.D.; Chimimba, C.T. Net Effects of Multiple Stressors in Freshwater Ecosystems: A Meta-Analysis. Glob. Chang. Biol. 2016, 22, 180–189. [Google Scholar] [CrossRef]
- Johnson, S.L.; Penaluna, B.E. Climate Change and Interactions with Multiple Stressors in Rivers. In Multiple Stressors in River Ecosystems; Elsevier: Amsterdam, The Netherlands, 2019; pp. 23–44. [Google Scholar]
- Shaffer, M.L. Minimum Population Sizes for Species Conservation. Bioscience 1981, 31, 131–134. [Google Scholar] [CrossRef]
- Ishiyama, N.; Koizumi, I.; Yuta, T.; Nakamura, F. Differential Effects of Spatial Network Structure and Scale on Population Size and Genetic Diversity of the Ninespine stickleback in a Remnant Wetland System. Freshwat. Biol. 2015, 60, 733–744. [Google Scholar] [CrossRef] [Green Version]
- Ishiyama, N.; Nagayama, S.; Akasaka, T.; Nakamura, F. Habitat use by Endangered Japanese Crayfish in Low-Gradient Streams of Southern Hokkaido, Japan: Reach and Microhabitat-Scale Analysis. Hydrobiologia 2012, 686, 257–266. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W. Predicting Species Distribution: Offering More than Simple Habitat Models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef]
- Comte, L.; Buisson, L.; Daufresne, M.; Grenouillet, G. Climate-Induced Changes in the Distribution of Freshwater fish: Observed and Predicted Trends. Freshwat. Biol. 2013, 58, 625–639. [Google Scholar] [CrossRef] [Green Version]
- Kawanabe, Y.; Mizuno, N. Freshwater Fishes of Japan; Yama to Keikoku-sha: Tokyo, Japan, 1998. (In Japanese) [Google Scholar]
- Goto, A. Geographic Distribution and Variations of Two Types of Cottus nozawae in Hokkaido, and Morphological Characteristics of C. amblystomopsis from Sakhalin. Jpn. J. Ichthyol. 1980, 27, 97–105. [Google Scholar]
- Goto, A. Ecological and Morphological Divergence of the Freshwater Sculpin, Cottus nozawae Synder-I. Spawning Behavior and Process of the Development in the Post-Hatching Stage. Bull. Fac. Fish. Hokkaido Univ. 1975, 26, 31–37, (In Japanese with English Abstract). [Google Scholar]
- Ishiyama, N.; Watanabe, K.; Nagayama, S.; Nakamura, F.; Kenmotsu, H.; Takahashi, H.; Maruoka, N.; Iwase, H. The Influence of Bedrock Outcrop on Stream Fish Habitat, and the Assessment of Restoration in a Bedrock Streamchannel. Ecol. Civil. Eng. 2009, 12, 57–66, (In Japanese with English Abstract). [Google Scholar] [CrossRef]
- Watanabe, K.; Nakamura, F.; Kamura, K.; Yamada, H.; Watanabe, Y.; Tsuchiya, S. Influence of Stream Alteration on the Abundance and Distribution of Benthic Fish. Ecol. Civ. Eng. 2001, 4, 133–146, (In Japanese with English Abstract). [Google Scholar] [CrossRef]
- JMA. Climate Change Monitoring Report. 2019. Available online: https://www.data.jma.go.jp/cpdinfo/monitor/2019/pdf/ccmr2019_all.pdf (accessed on 15 January 2021). (In Japanese).
- JMA. The Climate Records in Japan. Available online: https://www.data.jma.go.jp/obd/stats/etrn/index.php (accessed on 19 March 2021). (In Japanese).
- Ishiyama, N.; Miura, K.; Inoue, T.; Sueyoshi, M.; Nakamura, F. Geology-Dependent Impacts of Forest Conversion on Stream Fish Diversity. Conserv. Biol. 2020. [Google Scholar] [CrossRef]
- Sueyoshi, M.; Ishiyama, N.; Nakamura, F. β-Diversity Decline of Aquatic Insects at the Microhabitat Scale Associated with Agricultural Land Use. Landsc. Ecol. Eng. 2016, 12, 187–196. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model. Selection and Multimodel Inference: A Practical Information-Theoretic Approach; Springer: New York, NY, USA, 2002. [Google Scholar]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S.; Christensen, R.H.B.; Singmann, H. lme4: Linear mixed-effects models using Eigen and S; R Package Version 1.4; 2014; Available online: https://www.researchgate.net/publication/279236477_Package_Lme4_Linear_Mixed-Effects_Models_Using_Eigen_and_S4 (accessed on 1 April 2021).
- Barton, K. MuMIn: Multi-Model Inference; R Package Version 1.43.17; 2020; Available online: https://cran.r-project.org/web/packages/MuMIn/index.html (accessed on 1 April 2021).
- Ishiyama, N.; Ryo, M.; Kataoka, T.; Nagayama, S.; Sueyoshi, M.; Terui, A.; Mori, T.; Akasaka, T.; Nakamura, F. Predicting the Ecological Impacts of Large-dam Removals on a River Network Based on Habitat-Network Structure and Flow Regimes. Conserv. Biol. 2018, 32, 1403–1413. [Google Scholar] [CrossRef]
- Carstensen, B.; Plummer, M.; Laara, E.; Hills, M. Epi: A Package for Statistical Analysis in Epidemiology; R Package Version 2.44; 2021; Available online: https://cran.r-project.org/web/packages/Epi/index.html (accessed on 1 April 2021).
- Edwards, P.A.; Cunjak, R.A. Influence of Water Temperature and Streambed Stability on the Abundance and Distribution of Slimy Sculpin (Cottus cognatus). Environ. Biol. Fishes 2007, 80, 9–22. [Google Scholar] [CrossRef]
- Legalle, M.; Santoul, F.; Figuerola, J.; Mastrorillo, S.; Céréghino, R. Factors Influencing the Spatial Distribution Patterns of the Bullhead (Cottus gobio L., Teleostei cottidae): A Multi-Scale Study. Biodivers. Conserv. 2005, 14, 1319–1334. [Google Scholar] [CrossRef]
- Yagami, T.; Goto, A. Patchy Distribution of a Fluvial sculpin, Cottus nozawae, in the Gakko River System at the Southern Margin of its Native Range. Ichthyol. Res. 2000, 47, 27–286. [Google Scholar] [CrossRef]
- Dorts, J.; Bauwin, A.; Kestemont, P.; Jolly, S.; Sanchez, W.; Silvestre, F. Proteasome and Antioxidant Responses in Cottus gobio during a Combined Exposure to Heat Stress and Cadmium. Comp. Biochem. Physiol. Toxicol. Pharmacol. 2012, 155, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Schinegger, R.; Palt, M.; Segurado, P.; Schmutz, S. Untangling the Effects of Multiple Human Stressors and Their Impacts on Fish Assemblages in European Running Waters. Sci. Total Environ. 2016, 573, 1079–1088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Townsend, C.R.; Uhlmann, S.S.; Matthaei, C.D. Individual and Combined Responses of Stream Ecosystems to Multiple Stressors. J. Appl. Ecol. 2008, 45, 1810–1819. [Google Scholar] [CrossRef]
- Poff, N.L.; Allan, J.D.; Bain, M.B.; Karr, J.R.; Prestegaard, K.L.; Richter, B.D.; Sparks, R.E.; Stromberg, J.C. The Natural Flow Regime. Bioscience 1997, 47, 769–784. [Google Scholar] [CrossRef]
- Yamamoto, E.; Goto, A.; Nakaya, K.; Amaoka, K. Aquatic Insect Fauna and Feeding Habits of Two Cottus Species in the Daitobetsu River of Southern Hokkaido. Bull. Fac. Fish. Hokkaido Univ. 1988, 39, 237–256, (In Japanese with English Abstract). [Google Scholar]
- Brittain, J.E.; Eikeland, T.J. Invertebrate Drift—A Review. Hydrobiologia 1988, 166, 77–93. [Google Scholar] [CrossRef]
- Gibbins, C.N.; Vericat, D.; Batalla, R.J. Relations between Invertebrate Drift and Flow Velocity in Sand-Bed and Riffle Habitats and the Limits Imposed by Substrate Stability and Benthic Density. J. N. Am. Benthol. Soc. 2010, 29, 945–958. [Google Scholar] [CrossRef]
- Jiménez-Valverde, A.; Lobo, J.M. Threshold Criteria for Conversion of Probability of Species Presence to Either–or Presence–Absence. Acta Oecol. 2007, 31, 361–369. [Google Scholar] [CrossRef]
- Manel, S.; Williams, H.C.; Ormerod, S.J. Evaluating Presence–Absence Models in Ecology: The Need to Account for Prevalence. J. Appl. Ecol. 2001, 38, 921–931. [Google Scholar] [CrossRef]
- De Chazal, J.; Rounsevell, M.D.A. Land-Use and Climate Change Within Assessments of Biodiversity Change: A Review. Glob. Environ. Chang. 2009, 19, 306–315. [Google Scholar] [CrossRef]
- Mantyka-Pringle, C.S.; Martin, T.G.; Moffatt, D.B.; Linke, S.; Rhodes, J.R. Understanding and Predicting the Combined Effects of Climate Change and Land-use Change on Freshwater Macroinvertebrates and Fish. J. Appl. Ecol. 2014, 51, 572–581. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Variable | AIC |
|---|---|
| (Abundance) | |
| -Temperature- | |
| Mean summer water temperature | 241.47 |
| Maximum summer water temperature | 245.34 |
| -Substrate- | |
| Proportion of large substrate | 247.97 |
| Proportion of small substrate | 248.01 |
| (Occurrence) | |
| -Temperature- | |
| Mean summer water temperature | 43.01 |
| Maximum summer water temperature | 49.41 |
| Variables | df | AIC | ∆AIC | Weight |
|---|---|---|---|---|
| −Temp + Velocity + Temp × Velocity | 5 | 232.6 | 0.0 | 0.3 |
| −Temp + Velocity | 4 | 234.9 | 2.3 | 0.1 |
| −Temp + Velocity + Temp × Velocity + Substrate | 6 | 235.5 | 2.9 | 0.1 |
| Variables | df | AIC | ∆AIC | Weight |
|---|---|---|---|---|
| −Temp + Area − Catchment −Temp × Catchment | 5 | 42.6 | 0.0 | 0.06 |
| −Temp + Area − Catchment | 4 | 42.6 | 0.0 | 0.06 |
| −Temp + Area − Slope − Catchment | 5 | 42.8 | 0.3 | 0.05 |
| −Temp | 2 | 43.0 | 0.5 | 0.05 |
| −Temp − Slope + Temp * Slope | 4 | 43.5 | 0.9 | 0.04 |
| −Temp + Area − Slope − Catchment − Temp * Catchment | 6 | 43.5 | 0.9 | 0.04 |
| −Temp −Slope | 3 | 43.6 | 1.1 | 0.03 |
| −Temp + Area − Catchment + Farmland − Temp * Farmland | 6 | 43.8 | 1.3 | 0.03 |
| −Temp + Area | 3 | 43.9 | 1.3 | 0.03 |
| −Temp − Slope − Catchment | 4 | 44.0 | 1.4 | 0.03 |
| −Temp + Area − Catchment − Farmland − Temp * Catchment | 6 | 44.0 | 1.5 | 0.03 |
| −Temp + Area − Slope − Catchment + Temp*Slope | 6 | 44.1 | 1.5 | 0.03 |
| −Temp + Area − Slope − Catchment + Farmland − Temp * Farmland | 7 | 44.2 | 1.6 | 0.03 |
| −Temp − Catchment | 3 | 44.3 | 1.7 | 0.02 |
| −Temp + Area − Slope − Catchment − Farmland | 6 | 44.4 | 1.8 | 0.02 |
| −Temp + Area − Catchment − Farmland | 5 | 44.5 | 1.9 | 0.02 |
| Variable | 95% CIs | ||
|---|---|---|---|
| Temp | −3.7 | - | −0.5 |
| Area | −0.6 | - | 3.4 |
| Slope | −1.7 | - | 0.5 |
| Temp * Slope | −0.6 | - | 2.1 |
| Catchment | −2.9 | - | 0.6 |
| Temp * Catchment | −5.5 | - | 1.9 |
| Farmland | −1.4 | - | 1.5 |
| Temp * Farmland | −3.6 | - | 0.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suzuki, K.; Ishiyama, N.; Koizumi, I.; Nakamura, F. Combined Effects of Summer Water Temperature and Current Velocity on the Distribution of a Cold-Water-Adapted Sculpin (Cottus nozawae). Water 2021, 13, 975. https://doi.org/10.3390/w13070975
Suzuki K, Ishiyama N, Koizumi I, Nakamura F. Combined Effects of Summer Water Temperature and Current Velocity on the Distribution of a Cold-Water-Adapted Sculpin (Cottus nozawae). Water. 2021; 13(7):975. https://doi.org/10.3390/w13070975
Chicago/Turabian StyleSuzuki, Kaiji, Nobuo Ishiyama, Itsuro Koizumi, and Futoshi Nakamura. 2021. "Combined Effects of Summer Water Temperature and Current Velocity on the Distribution of a Cold-Water-Adapted Sculpin (Cottus nozawae)" Water 13, no. 7: 975. https://doi.org/10.3390/w13070975