
Who’s Next? Non-Indigenous Cnidarian and Ctenophoran Species Approaching to the Italian Waters



1
Department of Biological and Environmental Sciences and Technologies, DiSTeBA, University of Salento, Lecce 73100, Italy
2
CoNISMa, Consorzio Nazionale Interuniversitario per le Scienze del Mare, Rome 00196, Italy
*
Author to whom correspondence should be addressed.
Water 2021, 13(8), 1062; https://doi.org/10.3390/w13081062
Submission received: 26 February 2021
/
Revised: 2 April 2021
/
Accepted: 6 April 2021
/
Published: 12 April 2021
(This article belongs to the Special Issue Implementation of Biodiversity and Ecosystem Services in Marine Ecosystem Management)
Abstract
:The aims of the present paper were to review the knowledge about the Mediterranean non-indigenous species of the taxa Cnidaria and Ctenophora (CC NIS), to screen the risk of 98 species for their potential invasiveness in the Mediterranean Sea and their approach to the Italian waters. Of these, 38% are well established in the basin, 4% are known for their invasiveness, 44% are casual, 11% have a taxonomic status unresolved, and 3% are included in the category ”cryptogenic”. The biodiversity CC NIS of the Mediterranean Sea has changed considerably in the last two decades and 27 out of 98 Mediterranean CC NIS are present in the Italian waters. Fifteen CC NIS, some equipped with high invasive potential, should be regarded as good candidates to become future immigrants of the Italian waters. Anticipatory NIS forecast based on biogeographical and ecological analyses may provide a useful tool for targeted management of the CC NIS issue and for the assessment of the second descriptor of Good Environmental Status. On the other hand, conservation and management of marine ecosystem should be based on the conservation of the essential environmental conditions for the functioning of these ecosystems instead of the contamination or eradication of alien species.
1. Introduction
Marine non-indigenous species (NIS) represent a potential risk to the host environments [1,2] inducing ecosystem alterations often with socioeconomic effects in coastal areas [3,4,5].
Dispersal and invasion of species in new habitats have occurred throughout the history of marine life, but an important point is the fast rate at which invasions are recently taking place facilitated by anthropic activities [6]. The rate of introduction and the spread of invasive alien species increased rapidly in recent decades, so that these species are now considered one of the first anthropogenic threats for the marine communities [7,8,9]. Worldwide revisions and inventories highlight that biological diversity consists of both long-term native species and NIS that in some regions can contribute to local biodiversity with up to more than half of all the species [10]. Today, more than 980 alien species have been counted in the most invaded marine region of the world situated between Spanish and Middle East Mediterranean waters [1]. The best-known Mediterranean invasions are linked to macroalgae with negative impacts on native communities, biodiversity reduction, and ecological relationships alteration [11,12,13,14].
Mediterranean NIS have been examined by the scientific community through spatial and temporal patterns flanked by revision or species inventories [1,5,6,15,16,17,18,19,20,21,22,23,24,25,26,27]. Moreover, breaking of ecological barriers and excavating of canals promoted invasions of marine NIS leading to inclusion of the Mediterranean Sea into the Indo-Pacific region [28]. In the last decades, climate change affects the vulnerability of the region to biological invasions [29,30,31], influencing native species’ geographical and depth ranges [32,33] or the possibility that these species may be replaced by NIS [34,35]. Changing climate is leading indeed the Mediterranean Sea into a new stage characterized by an increase in the number of NIS with warm water affinities [16,36].
Several studies have covered the native distribution range of the Mediterranean marine NIS, but most of them have examined it only at country or regional level [2,37,38,39]. Only a few studies have provided a large-scale estimate of the native distribution of NIS across the Mediterranean and the other European Seas [1,27,40]. Furthermore, the revolution of Good Environmental Status (GES), 11 descriptors of the European Marine Strategy Framework Directive, introduced a new approach to define the status of the environment considering the effects of the putative causes of impact on the living component [33].
In this study, the Mediterranean non-indigenous species of the taxa Cnidaria and Ctenophora (CC NIS) have been considered as a good proxy for changes in biodiversity in the plankton and in the benthos, because of their structuring and functional roles. They inhabit all aquatic ecosystems displaying a wide array of life-cycle strategies with potential negative impacts on biodiversity, ecosystem, tourism, fisheries, fish farms, and power plants. Their native distribution and the main introduction pathways have been examined jointly to time trends of species introductions in relation to the prevailing native distribution areas. Moreover, evaluation of their invasiveness, also through the analysis of some case studies, could allow anticipatory NIS forecast based on biogeographical and ecological analyses of the CC NIS identified as good candidates to become future immigrants of the Italian waters that, to date, host more than 160 marine and brackish NIS along its coastline [39].
2. Materials and Methods
An inclusive bibliographic survey was performed to gather data for this review (indexed and non-indexed journals, checklists, on-line databases, and grey literature). Taxonomically, our survey is based on the extensive revision of the information found in the World Register of Marine Species [41]. Only CC NIS detected in marine and brackish waters were considered.
We identified Mediterranean CC NIS by examining records from the 19th century to 2020 to trace their origin, date, method of introduction, current distribution and establishment status, and global distribution. A database with more than 18,200 records was organized to provide the following information: species, family, collector, life-cycle phase, reproductive state, location, date of collection, publication year of the article, substrate type, water depth, synonymy, and cited references.
The following data (Supplementary Materials) are provided for each CC NIS:
- –
- establishment success in the Mediterranean Sea according to Zenetos et al. [15]: established (widely recorded at some sites), invasive (able to disseminate from their area of initial introduction), casual (few records), cryptogenic (species with no definite evidence of their native or introduced status), questionable (species with taxonomic status unresolved), unknown;
- –
- first record in the Mediterranean Sea (date, locality, reference: the date and location of the first observation) of each species in the Mediterranean Sea were extracted from the literature; if possible, the actual date of first record was reported, along with its publication date;
- –
- native distribution range: following the global marine biogeographic realms proposed by Spalding et al. [42];
- –
- distribution per Mediterranean subregion (following MSFD, Marine Strategy Framework Directive, 2017) [43]: Aegean-Levantine Sea, Ionian Sea and the Central Mediterranean Sea, Adriatic Sea, Western Mediterranean Sea;
- –
- extra-Mediterranean distribution;
- –
- the taxonomic position of each species (Class, Order, Family);
- –
- the primary pathway(s) of introduction in Europe based on CBD [44]: indicating the frequency if low (L), medium (M) or high (H);
- –
- possible notes;
- –
- code of the main pathway(s) of introduction: A (aquaculture), G (range expansion through the Strait of Gibraltar), Lp (Lessepsian), S (shipping), U (unknown).
Temporal trends of CC NIS introductions in the Mediterranean Sea per 5-year intervals have been performed for the time period 1950–2014, whose native distribution corresponds to at least one of the five most important realms of the Mediterranean CC NIS’ native distribution ranges (Temperate Northern Atlantic, Central Indo-Pacific, Western Indo-Pacific, Tropical Atlantic, Temperate Australasia). Due to the time lag between observation date of a new CC NIS and its subsequent reporting, temporal trends only reports up to 2014 were taken into consideration.
Finally, CC NIS have been investigated through the evaluation of their invasiveness (depending on the species ability to rapidly conquer new spaces, spread and generate various impacts) by identifying the possible future immigrants of the Italian waters, and considering also the distribution of CC NIS already established in the Italian seas through the following steps: (1) summarize the data about CC NIS in the Italian waters; (2) determine the origin regions for the CC invaders; (3) analyze CC biodiversity data in each region; (4) determine of new potential Italian invaders according to the data about CC invasive species in neighboring regions (information extracted from Supplementary Materials).
3. Results
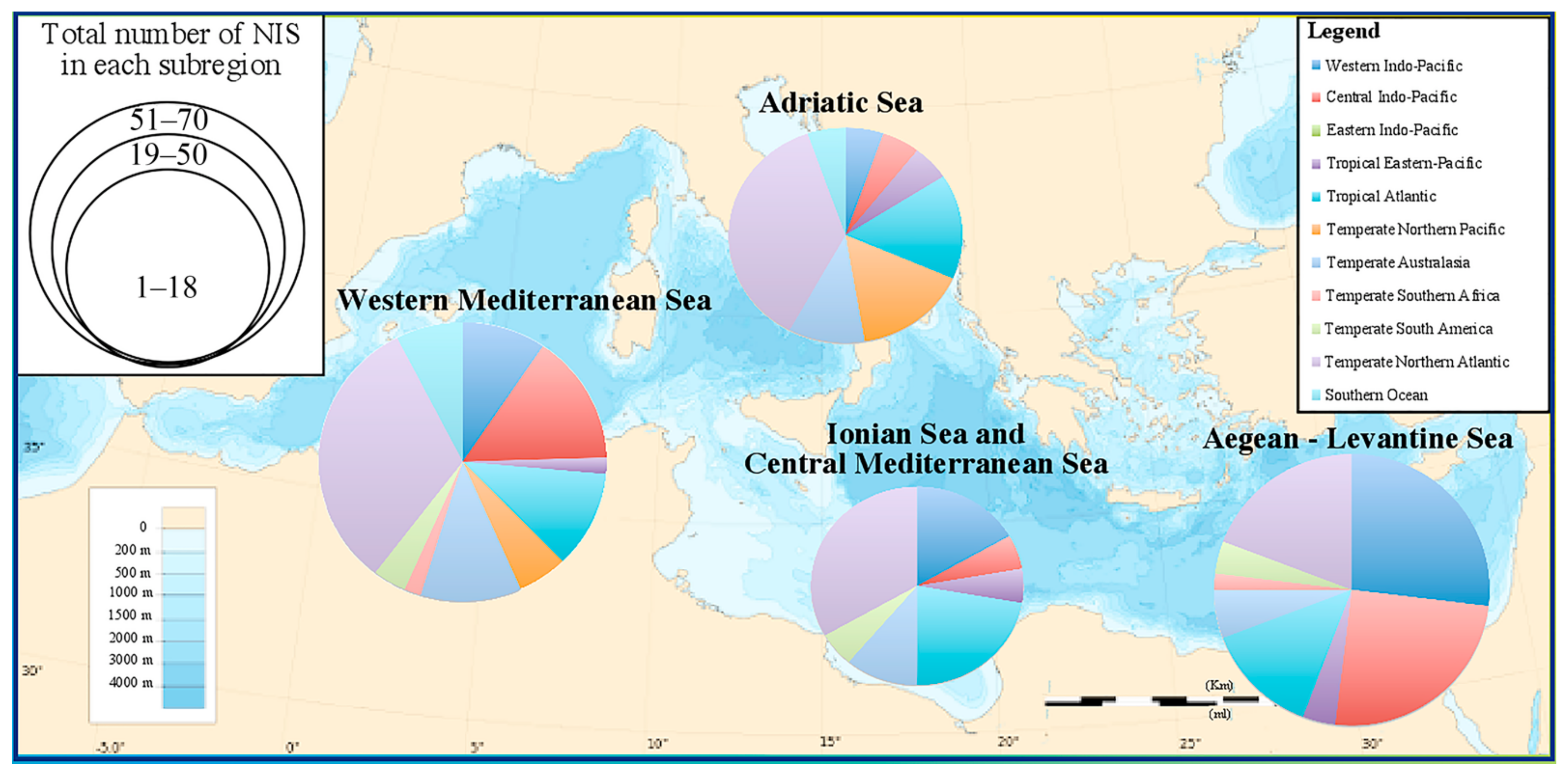
Most CC NIS are concentrated in the Aegean-Levantine region (61 species, 38%) followed by the Western Mediterranean Sea (59 species, 36%), Adriatic Sea (22 species, 14%), and Ionian Sea and Central Mediterranean Sea (20 species, 12%). Most CC NIS, established in one or more Mediterranean subregion [43], have their native distribution range in the following Indo-Pacific realms: Central Indo-Pacific (20 species, 21%), Western Indo-Pacific (19 species, 19%), Temperate Australasia (6 species, 6%), and Tropical Eastern-Pacific (2 species, 2%). On the other hand, the Temperate Northern Atlantic (27 species, 28%) and the Tropical Atlantic (10 species, 10%) constitute also important realms of Mediterranean NIS native distributions (Figure 1, Supplementary Materials). Very few CC NIS have their native distribution in the Temperate Northern Pacific (3 species, 3%), Temperate South Africa (2 species, 2%), Temperate South America (2 species, 2%), and Southern Ocean (4 species, 4%); 3 species have an unknown origin (3%). More biogeographical details of the CC NIS with native distribution in each MSFD Mediterranean subregion are provided in Supplementary Materials.
Of the 98 CC NIS, 38% are well established in the basin, 4% are known for their invasiveness, 44% are casual, 11% have a taxonomic status unresolved, and 3% are included in the category “cryptogenic”.
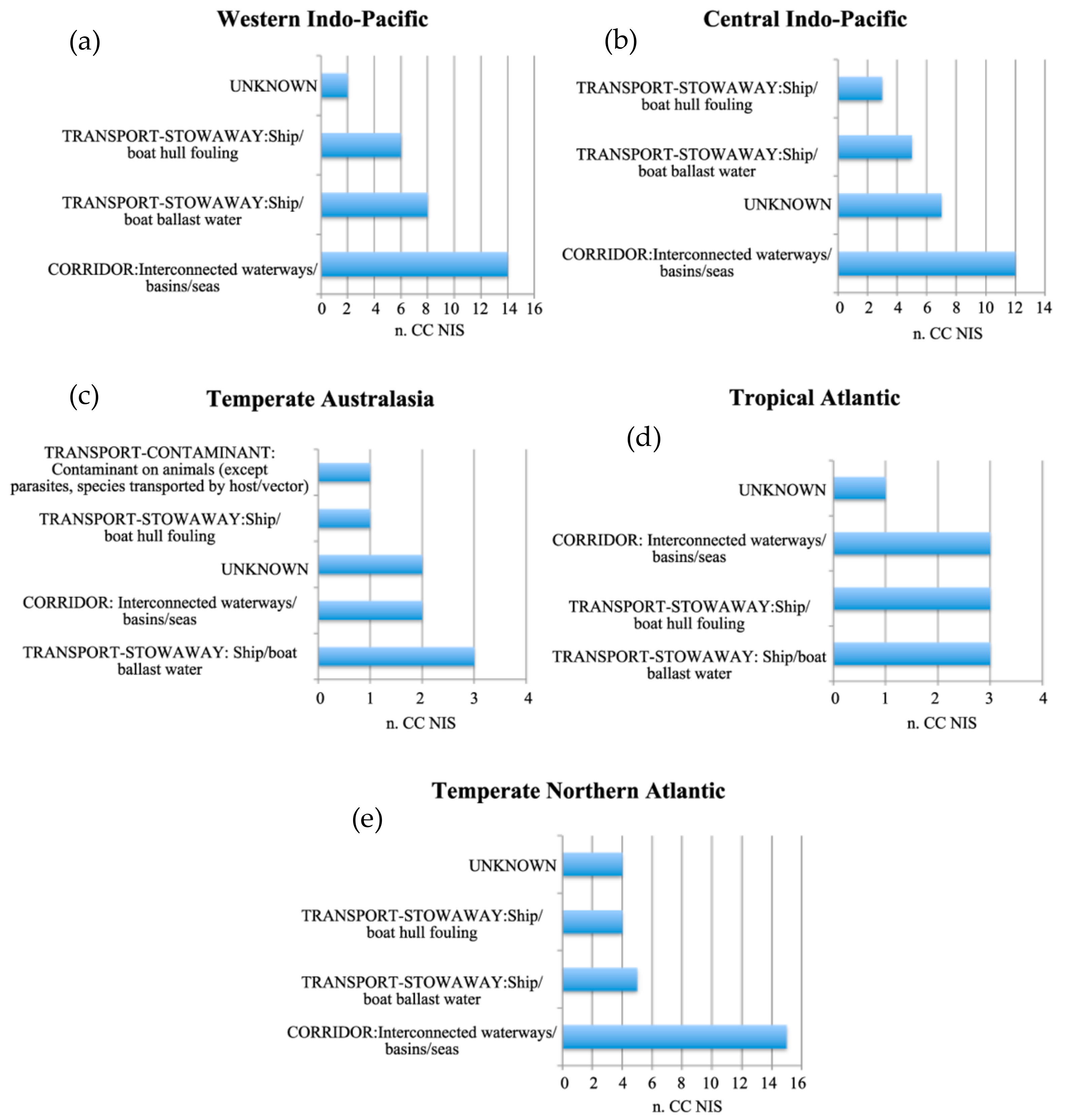
The analysis has shown different patterns of the most important pathways among the native distribution realms (Figure 2).
CC NIS with native distribution in the Western and Central Indo-Pacific have been introduced into the Mediterranean Sea mostly through “Corridor: interconnected waterways/basins/Seas”, corresponding to the Suez Canal. On the other hand, CC NIS with native distribution in the Temperate Australasia have been introduced mainly through shipping, including both “Transport-stowaway: ship/boat ballast water” (hereafter referred to as shipping-ballast) and “Transport-stowaway: ship/boat hull fouling” (hereafter referred to as shipping-fouling). “Transport-contaminant: contaminant on animals (except parasites, species transported by host/vector)” (hereafter referred to as aquaculture-contamination) concerns a few CC NIS with Temperate Australasia native realm. Finally, most CC NIS with native distribution in the Tropical Atlantic are associated with shipping (both ballast and fouling) and with “Corridor: interconnected waterways/basins/Seas”. Similarly, CC NIS related to the Temperate Northern Atlantic realm are mainly linked to interconnection across the Strait of Gibraltar and shipping-ballast/fouling.
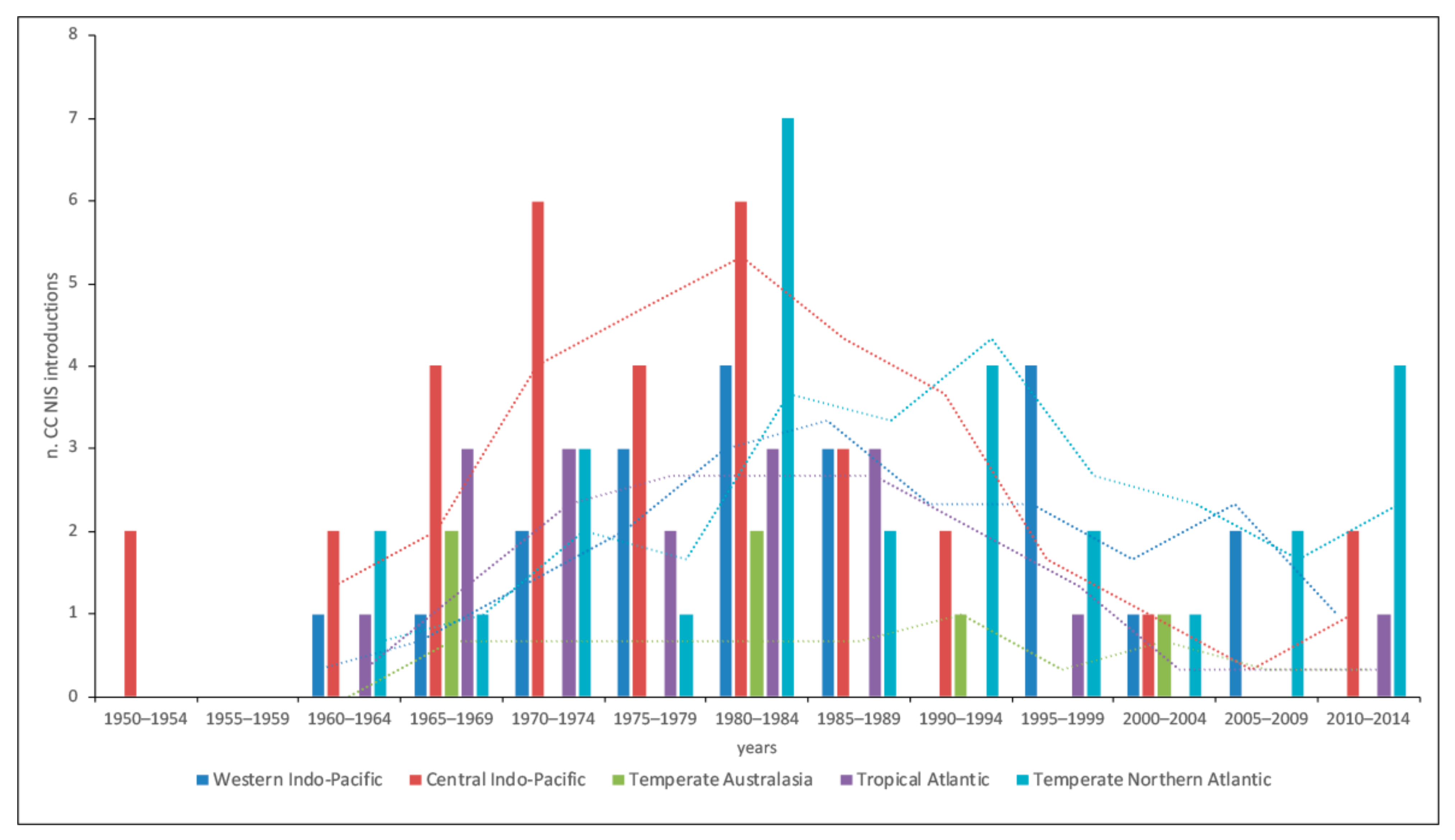
Up to 1980–1984, temporal analysis revealed an overall increasing trend of new CC NIS introductions with native distribution in the realms: Western and Central Indo-Pacific, Temperate Australasia, Tropical Atlantic, and Temperate Northern Atlantic. After this period, most of them present a constant or decreasing trend, less for CC NIS introduced from the Indo-Pacific and Temperate Northern Atlantic (Figure 3).
3.1. Cnidaria and Ctenophora Potential Invaders of the Italian Waters
Among the most invasive species, Mnemiopsis leidyi A. Agassiz, 1865 and Rhopilema nomadica Galil, Spanier & Ferguson, 1990 have been recently reported in the Italian waters [45,46,47,48,49]. Other CC NIS (15 species), well established in some areas of the Mediterranean basin (some of these equipped with invasive potential), could soon reach the Italian waters (Table 1).
Figure 4 shows the distribution of CC NIS in the Italian seas. The majority of CC NIS are concentrated in the South (16 species) and Central Tyrrhenian Sea (9 species), in the Ligurian Sea (14 species) and in the North Adriatic Sea (9 species).
3.2. Oculina patagonica, a Positive Alien Species?
In 1966, Oculina patagonica de Angelis, 1908 has been registered for the first time in the Mediterranean waters (Savona, Italy) [94,95]. This species expanded its presence, being found in Alicante (Spain, 1973), Egypt (1981) and in the Eastern Mediterranean (1992), as well as in the Croatian waters in 2011 [95]. Recently, it has been detected also in the Sicilian coasts (Giovanni Giallongo personal observation), suggesting an invasive trend in the Italian waters. Its widespread presence is due to its capacity to adapt to harsh conditions and almost to every kind of substrate [96], in vertical or horizontal surfaces and in a wide depth range [97]. Artificial substrata are especially suitable for this invasive species [95,98], which seems to have little problems even in polluted conditions [99,100].
Probably one of the keys to understand the successful invasion of this anthozoan is related to its early sexual reproduction (1–2 years, [95]) and the bailout propagation of the species [99]. Gametogenesis is fast, taking five months in the case of female colonies and three months in the case of male colonies [101], similar to the parasitic alcyonarian Alcyonium coralloides [102]. This kind of reproduction may be an advantage in a fast-changing sea in which cnidarians may have problems in the stabilization of their populations due to climate change [103]. However, another potential advantage in the species expansion is the relationship with sea urchins and the erosion of the hard bottom substrate [100]. It has been shown that, in seven years (2003–2010), the population density increased between 172–276% [100], concurrently with an increase in abundance of the Arbaxia lixula and Paracentrotus lividus sea urchins. The surface eroded by the sea urchins is not suitable for most of the sessile species, but it seems to be an ideal substrate for Oculina patagonica expansion.
The presence of the sea urchins seems to be an essential point for the invasive species, which takes advantage of the absence of the macroalgae in shallow waters. However, one interesting point is that, once settled, O. patagonica may share the space with new incoming algae that are not grazed by the sea urchins [100]. So, in a certain sense, we are in front of a facilitator species, which in some way partially “protects” some of the autochthonous benthic algae from the excessive sea urchin grazing. The system recovers and may also enhance biodiversity on some occasions due to the presence of this eco-engineering species [104]. O. patagonica is thus a good monitored example of how a NIS may partially transform the habitat.
4. Discussion and Conclusions
The Mediterranean Sea is widely colonized by marine NIS hosting the highest number of them in the world [5]. Mediterranean cnidarian biodiversity is changing, and seawater warming is impacting marine ecosystems, but as highlighted by Bianchi et al. [105], an accurate evaluation of changes requires the availability of long-term biological data series.
The Mediterranean high number of CC NIS is mainly linked to the presence of the Lessepsian immigrants, heavy shipping traffic (fouling and ballast waters), but also to the long history of marine monitoring [1,2,15,16,17,18,40,106].
According to Galil et al. [28] and Tsiamis et al. [1], the patterns concerning the Mediterranean CC NIS native distribution differ among the Mediterranean marine subregions, following the history and traits of the dominating primary pathways of introduction in each subregion. In fact, the vast majority of the marine Mediterranean CC NIS have their native distribution in the Central and Western Indo-Pacific mostly associated with their introductions into the Mediterranean Sea through the Suez Canal [16,25,40]. As a result, 38% of all the Mediterranean CC NIS primary introductions were reported first in the Aegean-Levantine Sea. On the other hand, CC NIS reported first from the Western Mediterranean Sea have their native distribution mainly in the Temperate Northern Atlantic (28%) as pathway of introduction the entrance through the Strait of Gibraltar and shipping (ballast/fouling) traffic [86,107,108]. Other species of Mediterranean CC NIS have their native distribution in the Tropical Atlantic (10%) and Temperate Australasia (6%) with shipping (both ballast and fouling) as most responsible pathway for their introductions into the Mediterranean basin. On the other hand, very few Mediterranean CC NIS are associated to the Southern Ocean (4%), possibly due to climate differences and the limited pathways presence in this marine realm.
A surge in the records in the 1980s reflects probably the publication of the results of specific programs in the Levant Sea (see Galil [109] for more details). Moreover, the increase in shipping transported CC NIS may be attributed to the increase in shipping volume throughout the Mediterranean basin resulting in new shipping routes, a result of a significant shift in global economy trends. In the same way, the increase in commercial introductions follows the increase of shellfish production in aquaculture facilities [110].
According to several researchers [1,23,24,25], during the last years an overall negative trend in species introductions is noted in the Mediterranean Sea. However, the observed general decrease in NIS introductions should be considered with caution because could be attributed to the time lag between the observation date of a NIS and its subsequent reporting [1,111,112]. Moreover, this decrease should be attributed to the fewer introductions of CC NIS associated with aquaculture-contamination [17] and shipping, presumably due to compulsory measures implemented at a national or European level [113,114], and the recent adoption by the International Maritime Organization (IMO) of the “International Convention for the Control and Management of Ships’ Ballast Water and Sediments” (BWM Convention). The approval of the guidance of the Marine Environment Protection Committee [115] should be encouraged since it concerns the shipping that has an important role in the CC NIS introductions [1,116,117,118,119]. We must consider also the permanence and spread of existing CC NIS, and their role in the ecosystem functioning. Is not only a matter of ‘new introduced species’, but also which role they may have in the habitat composition, energy fluxes and biogeochemical cycles in the near future [103].
According to the CC NIS invasiveness, and the distance from a region where their introductions were registered, three of the fifteen CC NIS potential invaders of the Italian waters (Table 1) deserve special attention.
The first species, Macrorhynchia philippina Kirchenpauer, 1872, native of the southern Pacific Ocean region, is one of the alien hydroid species most widely spread and well established in the Levant Sea [70,120]. This Lessepsian invader, found in shallow waters, can create dense populations and has been recorded also in the Aegean Sea introduced by shipping [70,121] being characterized by a high invasive potential. It is now a worldwide species in all tropical and sub-tropical oceans [108] and there are also records from temperate regions [122]. M. phylippina is a stinging species and the increase of its density and abundance in the Mediterranean basin could have a negative impact on local economies [70,121].
The second species, Oulastrea crispata (Lamarck, 1816), a non-indigenous zooxanthellate scleractinian coral, has been found in shallow water on the west coast of Corsica [92]. It is a species native on near-shore coral reefs in the central Indo-Pacific and a successful colonizer being able to settle on a wide variety of substrata and utilizes various reproductive strategies. Being widespread in temperate and subtropical waters, it is likely that it will be able to find a suitable temperature regime in the Mediterranean basin for further range expansion due to the ”tropicalization” of this area [105].
Finally, the third species, Eucheilota paradoxica Mayer, 1900 is native of the tropical Atlantic region and its medusa stage has been recorded near the Italian coasts (along the Croatian coast [51] and the French coast [56,57] in the ’70s and ’90s, respectively), while the most recent records occurred along the African coasts [60].
Another aspect to consider is the spatial scale. According to several authors [123,124], the biodiversity of native and exotic species is often negatively correlated at small scale, but positively related at large scale. At broader spatial scale the complexity of natural community environments reduces the resistance to the spread of NIS provided by high species richness [125,126]. Recently, research on positive or neutral effects of NIS has received particular attention even if, generally, their positive impacts may be underestimated [5,127,128,129,130]. In fact, there are many examples in the literature regarding the NIS introduction accounting its effects on diversity, structure and functioning of marine ecosystems [124,131,132,133,134] where the problem is magnified because complex life cycles facilitate connectivity among distant environments. An assessment of ecological and economic impacts is still lacking in the marine environment [135,136,137,138]. In European seas an attempt to treat the impact of NIS on biodiversity and ecosystem functioning was addressed by Katsanevakis et al. [5]. The authors found most of the ecological services (food provision, water purification, recreation and tourism) were negatively impacted by the presence of NIS. The results of a few macro-ecological studies [124,139] have revealed the presence of positive relationships between alien and native species richness in marine environments focusing attention on some large-scale features. These factors, not detectable by studies carried out on a single species or habitat, can contribute toward the development to a more complete understanding of the impacts of alien species from an ecosystem perspective [124,140,141]. Moreover, the general perception that NIS are a threat to biodiversity is true [142,143] but it should be considered that in most cases the time of investigation has not been long enough to clarify their real effects on the habitat composition and functioning [139,144]. On the one hand, the impact of NIS on biodiversity cannot be generalized because they cause different effects at several spatial and temporal scales [13,124,145,146,147,148,149,150,151] being difficult to predict the fate of NIS in new habitat [152]. On the other hand, there is the possibility that some endangered species could also benefit from their transport towards different environments. In fact, considering that some invaders could survive better than endemic species in changing habitats that may compensate the decrease in the number of species caused by climate change and may act as ‘reservoirs of diversity’ [139,153]. According to Davis et al. [154], the ‘practical value of the native-versus-alien species dichotomy’ in conservation is setting. As suggested by Giangrande et al. [139], therefore, it is essential to distinguish early NIS from naturalized species defining a temporal baseline that can be used for this purpose.
The monitoring bias is another issue that should not be neglected: inevitably more NIS primary introductions in the Mediterranean Sea have been reported for well-known taxonomic groups (e.g., mollusks, fish, macroalgae) [133,155,156,157]. In addition, more focused studies are needed to examine pathway-introduction of most Mediterranean CC NIS because for the majority of introductions the pathway certainty is not sufficient, with the exception of the Lessepsian immigrants.
The information concerning the pathways could indeed be useful for CC NIS management per marine subregion according to the MSFD, directing where priority should be given to avoid new introductions. Furthermore, risk screening of the potential invasiveness of non-native species in the Mediterranean Sea can aid managers in making informed decisions on targeting species for management [158].
The study of new arrivals at an ecosystem level should involve interdisciplinary synergism and accurate analysis of species distribution. This requires international cooperation to define issues of provenance and to assess possible risks related to commercial exchanges [139].
In conclusion, as suggested by Corriero et al. [124] and Buonocore et al. [120], from a conservation point of view, diverse marine communities have unquestioned conservation and ecological value, and provide economic benefits such as diving experiences, nursering effect, shelter of fisheries biomass, carbon sequesters. Therefore, two different models should be distinguished during marine monitoring, the first aimed at promptly reporting the arrival of NIS in hot spots of introduction and the second at evaluating the success of these species in marine communities. Finally, according to Ekebom [159] and considering the unpredictability of the invasions processes, methods to detect impacts should be improved by implementing mensurative and experimental studies at different spatial and temporal scales.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/w13081062/s1.
Author Contributions
Conceptualization, C.G. and S.R.; methodology, C.G.; validation, C.G. and S.R.; formal analysis, C.G.; investigation, C.G. and S.R.; resources, C.G. and S.R.; data curation, C.G. and S.R.; writing—original draft preparation, C.G. and S.R.; writing—review and editing, C.G. and S.R.; visualization, C.G. and S.R.; supervision, C.G. and S.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
We thank Stefano Piraino (University of Salento) for his contribution (advice and discussions) to the paper. Thanks to Francesco Cozzoli for consultancy on the processing of data relating to temporal trends.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tsiamis, K.; Zenetos, A.; Deriu, I.; Gervasini, E.; Cardoso, A.C. The native distribution of the European marine non-indigenous species. Aquat. Invasions 2018, 13, 187–198. [Google Scholar] [CrossRef]
- Servello, G.; Andaloro, F.; Azzurro, E.; Castriota, L.; Catra, M.; Chiarore, A.; Crocetta, F.; D’alessandro, M.; Denitto, F.; Froglia, C.; et al. Marine Alien Species in Italy: A Contribution to the Implementation of Descriptor D2 of The Marine Strategy Framework Directive. Mediterr. Mar. Sci. 2019, 20, 1–48. [Google Scholar] [CrossRef] [Green Version]
- Wallentinus, I.; Nyberg, C.D. Introduced marine organisms as habitat modifiers. Mar. Pollut. Bull. 2007, 55, 323–332. [Google Scholar] [CrossRef]
- Molnar, J.L.; Gamboa, R.L.; Revenga, C.; Spalding, M.D. Assessing the global threat of invasive species to marine biodiversity. Front. Ecol. Environ. 2008, 6, 485–492. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Wallentinus, I.; Zenetos, A.; Leppäkoski, E.; Çinar, M.E.; Oztürk, B.; Cardoso, A.C. Impacts of marine invasive alien species on ecosystem services and biodiversity: A pan-European review. Aquat. Invasions 2014, 9, 391–423. [Google Scholar] [CrossRef]
- Ojaveer, H.; Galil, B.G.; Carlton, J.T.; Alleway, H.; Goulletquer, P.; Lehtiniemi, M.; Marchini, A.; Miller, W.; Occhipinti-Ambrogi, A.; Peharda, M.; et al. Historical baselines in marine bioinvasions: Implications for policy and management. PLoS ONE 2018, 13, e0202383. [Google Scholar] [CrossRef] [Green Version]
- Nellemann, C.; Hain, S.; Alder, J. Rapid Response Assessment in Dead Water—Merging of Climate Change with Pollution, Over-Harvest, and Infestations in the World’s Fishing Grounds; United Nations Environment Programme, GRID-Arendal: Arendal, Norway, 2008. [Google Scholar]
- Costello, M.J.; Coll, M.; Danovaro, R.; Halpin, P.; Ojaveer, H.; Miloslavich, P. A census of marine biodiversity knowledge, resources, and future challenges. PLoS ONE 2010, 5, e12110. [Google Scholar] [CrossRef] [Green Version]
- Coll, M.; Piroddi, M.; Albouy, C.; Ben Rais Lasram, F.; Cheung, W.W.L.; Christensen, V.; Karpouzi, V.S.; Guilhaumon, F.; Mouillot, D.; Paleczny, M.; et al. The Mediterranean Sea under siege: Spatial overlap between marine biodiversity, cumulative threats and marine reserves. Glob. Ecol. Biogeogr. 2012, 21, 465–480. [Google Scholar] [CrossRef]
- Schlaepfer, M.A. Do non-native species contribute to biodiversity? PLoS Biol. 2018, 16, e2005568. [Google Scholar] [CrossRef] [Green Version]
- Boudouresque, C.F.; Verlaque, M. Biological pollution in the Mediterranean Sea: Invasive versus introduced macrophytes. Mar. Pollut. Bull. 2002, 44, 32–38. [Google Scholar] [CrossRef]
- Piazzi, L.; Meinesz, A.; Verlaque, M.; Akçali, B.; Antolić, B.; Argyrou, M.; Balata, D.; Ballesteros, E.; Calvo, S.; Cinelli, F.; et al. Invasion of Caulerpa racemosa var. cylindracea (Caulerpales, Chlorophyta) in the Mediterranean Sea: An assessment of the spread. Cryptogam. Algol. 2005, 26, 189–202. [Google Scholar]
- Piazzi, L.; Balata, D. Invasion of alien macroalgae in different Mediterranean habitats. Biol. Invasions 2009, 11, 193–204. [Google Scholar] [CrossRef]
- Thomsen, M.S.; Wernberg, T.; Tuya, F.; Silliman, B.R. Evidence for impact of nonindigenous macroalgae: A meta-analysis of experimental and field studies. J. Phycol. 2009, 45, 812–819. [Google Scholar] [CrossRef] [PubMed]
- Zenetos, A.; Çinar, M.E.; Pancucci-Papadopoulou, M.A.; Harmelin, J.G.; Furnari, G.; Andaloro, F.; Bellou, N.; Streftaris, N.; Zibrowius, H. Annotated list of marine alien species in the Mediterranean with records of the worst invasive species. Mediterr. Mar. Sci. 2005, 6, 63–118. [Google Scholar] [CrossRef]
- Zenetos, A.; Gofas, S.; Morri, C.; Rosso, A.; Violanti, D.; García Raso, J.E.; Çinar, M.E.; Almogi-Labin, A.; Ates, A.S.; Azzurro, E.; et al. Alien species in the Mediterranean Sea by 2012. A contribution to the application of European Union’s Marine Strategy Framework Directive (MSFD). Part 2. Introduction trends and pathways. Mediterr. Mar. Sci. 2012, 13, 328–352. [Google Scholar] [CrossRef] [Green Version]
- Katsanevakis, S.; Zenetos, A.; Belchior, C.; Cardoso, A.C. Invading European seas: Assessing pathways of introduction of marine aliens. Ocean Coast. Manag. 2013, 76, 64–74. [Google Scholar] [CrossRef]
- Galil, B.S.; Marchini, A.; Occhipinti-Ambrogi, A.; Minchin, D.; Narščius, A.; Ojaveer, H.; Olenin, S. International arrivals: Widespread bioinvasions in European Seas. Ethol. Ecol. Evol. 2014, 26, 152–171. [Google Scholar] [CrossRef] [Green Version]
- Nunes, A.L.; Katsanevakis, S.; Zenetos, A.; Cardoso, A.C. Gateways to alien invasions in the European Seas. Aquat. Invasions 2014, 9, 133–144. [Google Scholar] [CrossRef]
- Cardeccia, A.; Marchini, A.; Occhipinti-Ambrogi, A.; Galil, B.; Gollasch, S.; Minchin, D.; Narščius, A.; Olenin, S.; Ojaveer, H. Assessing biological invasions in European Seas: Biological traits of the most widespread non-indigenous species. Estuar. Coast. Shelf Sci. 2016, 201, 17–28. [Google Scholar] [CrossRef]
- Ojaveer, H.; Olenin, S.; Narščius, A.; Florin, A.B.; Ezhova, E.; Gollasch, S.; Jensen, K.R.; Lehtiniemi, M.; Minchin, D.; Normant-Saremba, M.; et al. Dynamics of biological invasions and pathways over time: A case study of a temperate coastal sea. Biol. Invasions 2016, 19, 799–813. [Google Scholar] [CrossRef]
- Galil, B.S.; Marchini, A.; Occhipinti-Ambrogi, A.; Ojaveer, H. The enlargement of the Suez Canal—Erythraean introductions and management challenges. Manag. Biol. Invasions 2017, 8, 141–152. [Google Scholar] [CrossRef] [Green Version]
- Zenetos, A. Progress in Mediterranean bioinvasions two years after the Suez Canal enlargement. Acta Adriat. 2017, 58, 347–358. [Google Scholar] [CrossRef]
- Zenetos, A.; Çinar, M.E.; Crocetta, F.; Golani, D.; Rosso, A.; Servello, G.; Shenkar, N.; Turon, X.; Verlaque, M. Uncertainties and validation of alien species catalogues: The Mediterranean as an example. Estuar. Coast. Shelf Sci. 2017, 191, 171–187. [Google Scholar] [CrossRef]
- Galil, B.S.; Marchini, A.; Occhipinti-Ambrogi, A. East is east and West is west? Management of marine bioinvasions in the Mediterranean Sea. Estuar. Coast. Shelf Sci. 2018, 201, 7–16. [Google Scholar] [CrossRef]
- Zenetos, A.; Corsini-Foka, M.; Crocetta, F.; Gerovasileiou, V.; Karachle, P.K.; Simboura, N.; Tsiamis, K.; Pancucci, A. Deep cleaning of alien and cryptogenic species records in the Greek Seas (2018 update). Manag. Biol. Invasions 2018, 9, 209–226. [Google Scholar] [CrossRef] [Green Version]
- Katsanevakis, S.; Poursanidis, D.; Hoffman, R.; Rizgalla, J.; Rothman, S.B.-S.; Levitt-Barmats, Y.; Hadjioannou, L.; Trkov, D.; Garmendia, M.K.; Rizzo, M.; et al. Unpublished Mediterranean records of marine alien and cryptogenic species. BioInvasions Rec. 2020, 9, 165–182. [Google Scholar] [CrossRef]
- Galil, B.S.; Boero, F.; Campbell, M.L.; Carlton, J.T.; Cook, E.; Fraschetti, S.; Gollasch, S.; Hewitt, C.L.; Jelmert, A.; Macpherson, E.; et al. ‘Double trouble’: The expansion of the Suez Canal and marine bioinvasions in the Mediterranean Sea. Biol. Invasions 2015, 17, 973–976. [Google Scholar] [CrossRef] [Green Version]
- Masters, G.; Norgrove, L. Climate change and invasive alien species. CABI Work. Pap. 2010, 1, 30. [Google Scholar]
- Occhipinti-Ambrogi, A.; Galil, B.S. Marine alien species as an aspect of global change. Adv. Oceanogr. Limnol. Adv. Oceanogr. Limnol. 2010, 1, 199–218. [Google Scholar] [CrossRef]
- Gravili, C. Alien jellyfish in expansion: The contribution of taxonomy to ecology. In Jellyfish: Ecology, Distribution Patterns and Human Interactions; Mariottini, G.L., Ed.; Nova Publishers: New York, NY, USA, 2017; pp. 27–49. [Google Scholar]
- Ponti, M.; Perlini, R.A.; Ventra, V.; Grech, D.; Abbiati, M.; Cerrano, C. Ecological shifts in Mediterranean coralligenous assemblages related to gorgonian forest loss. PLoS ONE 2014, 9, e102782. [Google Scholar] [CrossRef]
- Boero, F. The future of the Mediterranean Sea ecosystem: Towards a different tomorrow. Rend. Fis. Acc. Lincei 2015, 26, 3–12. [Google Scholar] [CrossRef]
- Dukes, J.S.; Mooney, H.A. Does global change increase the success of biological invaders? Trends Ecol. Evol. 1999, 14, 135–139. [Google Scholar] [CrossRef]
- Cox, G.W. Alien Species and Evolution: The Evolutionary Ecology of Exotic Plants, Animals, Microbes, and Interacting Native Species; Island Press: Washington, DC, USA, 2004. [Google Scholar]
- Mannino, A.; Balistreri, P.; Deidun, A. The marine biodiversity of the Mediterranean Sea in a changing climate: The impact of biological invasions. In Mediterranean Identities—Environment, Society, Culture; Fuerst-Bjeliš, B., Ed.; IntechOpen: Zagabria, Croatia, 2017; pp. 101–127. [Google Scholar]
- Galil, B.S. Seeing Red: Alien species along the Mediterranean coast of Israel. Aquat. Invasions 2007, 2, 281–312. [Google Scholar] [CrossRef]
- Zenetos, A.; Pancucci-Papadopoulou, M.A.; Zogaris, S.; Papastergiadou, E.; Vardakas, L.; Aligizaki, K.; Economou, A. Aquatic alien species in Greece: Tracking sources, patterns and effects on the ecosystem. J. Biol. Res. Thessalon. 2009, 12, 135–172. [Google Scholar]
- Occhipinti-Ambrogi, A.; Marchini, A.; Cantone, G.; Castelli, A.; Chimenz, C.; Cormaci, M.; Froglia, C.; Furnari, G.; Gambi, M.C.; Giaccone, G.; et al. Alien species along the Italian coasts: An overview. Biol. Invasions 2011, 13, 215–237. [Google Scholar] [CrossRef]
- Zenetos, A.; Gofas, S.; Verlaque, M.; Çinar, M.E.; García Raso, J.E.; Bianchi, C.N.; Morri, C.; Azzurro, E.; Bilecenoglu, M.; Froglia, C.; et al. Alien species in the Mediterranean Sea by 2010. A contribution to the application of European Union’s Marine Strategy Framework Directive (MSFD). Part I. Spatial distribution. Mediterr. Mar. Sci. 2010, 112, 381–493. [Google Scholar] [CrossRef] [Green Version]
- Appeltans, W.; Bouchet, P.; Boxshall, G.A.; De Broyer, C.; de Voogd, N.J.; Gordon, D.P.; Hoeksema, B.W.; Horton, T.; Kennedy, M.; Mees, J.; et al. (Eds.) World Register of Marine Species. 2012. Available online: http://www.marinespecies.org (accessed on 17 October 2020).
- Spalding, M.D.; Fox, H.E.; Allen, G.R.; Davidson, N.; Ferdaña, Z.A.; Finlayson, C.; Halpern, B.S.; Jorge, M.A.; Lombana, A.; Lourie, S.A.; et al. Marine ecoregions of the world: A bioregionalization of coastal and shelf areas. BioScience 2007, 57, 573–583. [Google Scholar] [CrossRef] [Green Version]
- MSFD. Technical Document on the Delineation of MSFD Article 4 Marine Regions and Subregions; Jensen, H.M., Panagiotidis, P., Eds.; 16/05/2017. Version 1.0; European Environment Agency: København, Denmark, 2017; 21p. [Google Scholar]
- CBD. Pathways of Introduction of Invasive Species, Their Prioritization and Management; UNEP/CBD/SBSTTA/18/9/Add.1; CBD: Montreal, QC, Canada, June 2014; 18p. [Google Scholar]
- Boero, F.; Putti, M.; Trainito, E.; Prontera, E.; Piraino, S.; Shiganova, T.A. First records of Mnemiopsis leidyi (Ctenophora) from the Ligurian, Thyrrhenian and Ionian Seas (Western Mediterranean) and first record of Phyllorhiza punctata (Cnidaria) from the Western Mediterranean. Aquat. Invasions 2009, 4, 675–680. [Google Scholar] [CrossRef]
- Crocetta, F.; Agius, D.; Balistreri, P.; Bariche, M.; Bayhan, Y.K.; Çakir, M.; Ciriaco, S.; Corsini-Foka, M.; Deidun, A.; El Zrelli, R.; et al. New Mediterranean Biodiversity Records (October 2015). Mediterr. Mar. Sci. 2015, 16, 682–702. [Google Scholar] [CrossRef] [Green Version]
- Boero, F.; Brotz, L.; Gibbons, M.J.; Piraino, S.; Zampardi, S. Impacts and effects of ocean warming on jellyfish. In Explaining Ocean Warming: Causes, Scale, Effects and Consequences; Lagìffoley, D., Baxter, J.M., Eds.; IUCN: Gland, Switzerland, 2016; pp. 213–237. [Google Scholar]
- Balistreri, P.; Spiga, A.; Deidun, A.; Gueroun, S.K.; Yahia, M.N.D. Further spread of the venomous jellyfish Rhopilema nomadica Galil, Spannier & Ferguson, 1990 (Rhizostomeae, Rhizostomatidae) in the western Mediterranean. BioInvasions Rec. 2017, 6, 19–24. [Google Scholar] [CrossRef]
- Malej, A.; Tirelli, V.; Lučić, D.; Paliaga, P.; Vodopivec, M.; Goruppi, A.; Sara, A.; Benzi, M.; Bettoso, N.; Camatti, E.; et al. Mnemiopsis leidyi in the northern Adriatic: Here to stay? J. Sea Res. 2017, 124, 10–16. [Google Scholar] [CrossRef]
- Morri, C.; Puce, S.; Bianchi, C.N.; Bitar, G.; Zibrowius, H.; Bavestrello, G. Hydroids (Cnidaria: Hydrozoa) from the Levant Sea (mainly Lebanon), with emphasis on alien species. J. Mar. Biol. Assoc. U.K. 2009, 89, 49–62. [Google Scholar] [CrossRef]
- Schmidt, H.E.; Benović, A. Notes on the Hydromedusae (Cnidaria) from the Adriatic Sea. J. Mar. Biol. Assoc. U.K. 1977, 57, 635–640. [Google Scholar] [CrossRef]
- Dallot, S.; Goy, J.; Carré, C. Peuplements de carnivores planctoniques gélatineux et structures productives en Méditerranée occidentale. Oceanol. Acta 1988, SP, 193–209. [Google Scholar]
- Goy, J.; Lakkis, S.; Zeidane, R. Les Méduses de la Méditerranée orientale. Rapp. Comm. int. Mer Méditerr. 1988, 31, 299. [Google Scholar]
- Goy, J.; Lakkis, S.; Zeidane, R. Les Méduses de la Méditerranée orientale. In A Propos des Migrations Lessepsiennes; Godeaux, J., Ed.; Bulletin de l’Institut Océanographique, No. Spéc.; Musée Océanographique: Monaco-Ville, Monaco, 1990; Volume 7, pp. 79–88. [Google Scholar]
- Goy, J.; Lakkis, S.; Zeidane, R. Les méduses (Cnidaria) des eaux Libanaises. The medusae Cnidaria of Lebanese waters. Ann. Inst. Océanogr. Monaco 1991, 67, 99–128. [Google Scholar]
- Carré, D.; Carré, C. Complex reproductive cycle in Eucheilota paradoxica (Hydrozoa: Leptomedusae): Medusae, polyps and frustules produced from medusa stage. Mar. Biol. Berl. 1990, 104, 303–310. [Google Scholar] [CrossRef]
- Buecher, E.; Gibbons, M.J. Temporal persistence in the vertical structure of the assemblage of planktonic medusae in the NW Mediterranean Sea. Mar. Ecol. Prog. Ser. 1999, 189, 105–115. [Google Scholar] [CrossRef]
- Touzri, C.; Yahia, O.K.D.; Hamdi, H.; Goy, J.; Yahia, M.N.D. Spatio-temporal distribution of Medusae (Cnidaria) in the Bay of Bizerte (South Western Mediterranean Sea). Cah. Biol. Mar. 2010, 51, 167–176. [Google Scholar]
- Ounifi-Ben Amor, K.; Rifi, M.; Ghanem, R.; Draief, I.; Zaouali, J.; Ben Soussi, J. Update of alien fauna and new records from Tunisian marine waters. Mediterr. Mar. Sci. 2016, 17, 124–143. [Google Scholar] [CrossRef]
- Kherchouche, A.; Hafferssas, A. Species composition and distribution of Medusae (Cnidaria: Medusozoa) in the Algerian coast between 2°e and 7°e (SW Mediterranean Sea). Mediterr. Mar. Sci. 2020, 21, 52–61. [Google Scholar] [CrossRef]
- Lakkis, S.; Zeidane, R. Les hydroméduses des eaux néritiques libanaises: Composition et distribution. Rapp. Procès-Verbaux Réun. Comm. Int. Explor. Sci. Mer Méditerr. 1985, 29, 179–180. [Google Scholar]
- Benović, A.; Lucić, D.; Onofri, V.; Batistić, M.; Njire, J. Bathymetric distribution of medusae in the open waters of the middle and south Adriatic Sea during spring 2002. J. Plankton Res. 2005, 27, 79–89. [Google Scholar] [CrossRef]
- Casanova, J.-P. Campagnes du “Meteor” dans l’Atlantique, N.-E. Siphonophores, Méduses et Thécosomes. Distribution verticale et comparaisons faunistiques avec la Méditerranée. Meteor Forsch.-Erbgeb. 1980, 32, 15–32. [Google Scholar]
- Gili, J.M. Estudio bionómico y ecológico de los cnidarios bentónicos de las islas Medes (Girona). Oecol. Aquat. 1981, 5, 105–123. [Google Scholar]
- Gili, J.M. Estudio Sistematico y Faunistico de los Cnidarios de la Costa Catalana. Ph.D. Thesis, University of Barcelona, Barcelona, Spain, 1986; pp. 1–565. [Google Scholar]
- Gili, J.M.; Pagès, F.; Vives, F. Distribution and ecology of a population of planktonic cnidarians in the western Mediterranean. In Modern Trends in the Systematics, Ecology and Evolution of Hydroids and Hydromedusae; Bouillon, J., Boero, F., Cicogna, F., Cornelius, P.F.S., Eds.; Clarendon Press: Oxford, UK, 1987; pp. 157–170. [Google Scholar]
- Gili, J.M.; Bouillon, J.; Pagès, F.; Palanques, A.; Puig, P.; Heussner, S. Origin and biogeography of the deep-water Mediterranean Hydromedusae including the description of two new species collected in submarine canyons of Northwestern Mediterranean. Sci. Mar. 1998, 62, 113–134. [Google Scholar] [CrossRef] [Green Version]
- Gili, J.M.; Pagès, F.; Bouillon, J.; Palanques, A.; Puig, P.; Heussner, S.; Calafat, A.; Canals, M.; Monaco, A. A multidisciplinary approach to the understanding of hydromedusan populations inhabiting Mediterranean submaryne canyons. Deep Sea Res. Part I 2000, 47, 1513–1533. [Google Scholar] [CrossRef]
- Touzri, C.; Hamdi, H.; Goy, J.; Yahia, M.N.D. Diversity and distribution of gelatinous zooplankton in the Southwestern Mediterranean Sea. Mar. Ecol. 2012, 33, 393–406. [Google Scholar] [CrossRef]
- Bitar, G.; Bitar-Kouli, S. Impact de la pollution sur la répartition des peuplements de substrat dur à Beyrouth (Liban—Mediterranée Orientale). Rapp. Comm. Int. Mer Méditerr. 1995, 34, 19. [Google Scholar]
- Zibrowius, H.; Bitar, G. Invertebres marins exotiques sur la côte du Liban. Leban. Sci. J. 2003, 4, 67–74. [Google Scholar]
- Çinar, M.E.; Bilecenoglu, M.; Öztürk, B.; Can, A. New records of alien species on the Levantine coast of Turkey. Aquat. Invasions 2006, 1, 84–90. [Google Scholar] [CrossRef]
- Uysal, I.; Turan, C. Impacts and risk of venomous and sting marine alien species in Turkish marine waters. Biharean Biol. 2019, 14, 41–48. [Google Scholar]
- Zakaria, H.Y. Pelagic coelenterates in the waters of the western part of the Egyptian Mediterranean coast during summer and winter. Oceanologia 2004, 46, 253–268. [Google Scholar]
- García Corrales, P.; Aguirre Inchaurbe, A.; González Mora, D. Contribución al conocimiento de los hidrozoos de las costas españolas. Parte I: Halecidos, Campanularidos y Plumularidos. Boln. Inst. Esp. Oceanogr. 1978, 4, 5–73. [Google Scholar]
- Roca, I.; Moreno, I. Distribución de los cnidarios bentónicos litorales en tres localidades de la margen W de la Bahía de Palma de Mallorca. Boll. Soc. Hist. Nat. Balear. 1985, 29, 19–30. [Google Scholar]
- Roca Martinez, I. Estudio de los Cnidarios Bentónicos de las Aguas Costeras de Mallorca. Bachelor’s Thesis, Universitat de les Illes Balears, Facultat de Ciències, Palma de Mallorca, Spain, 1986; pp. 1–32. [Google Scholar]
- El Beshbeeshy, M. Descriptions of some thecate hydroids (Cnidaria-Hydrozoa) from the Egyptian Mediterranean waters. Part 4. Families: Plumulariidae, Halopteridae & Kirchenpaueriidae. Bull. Nat. Inst. Oceanogr. Fish. 1995, 21, 397–412. [Google Scholar]
- Billard, A. Hydroïdes des côtes de Syrie (Mission Gruvel). In Les États de Syrie. Richesse Marines et Fluviales. Exploitation Actuelle—Avenir; Gruvel, A., Ed.; Bibltque Faune Colon Françaises; 1931; Volume 3, pp. 389–395. [Google Scholar]
- Picard, J. Origines et affinités de la faune d’hydropolypes (Gymnoblastes et Calyptoblastes) et d’hydroméduses (Anthoméduses et Leptoméduses) de la Méditerranée. Rapp. Procès-Verbaux Réun. Comm. Int. Explor. Sci. Mer Méditerr. 1958, 14, 187–199. [Google Scholar]
- Marinopoulos, J. Biological survey of the eastern Mediterranean Sea: Hydroids (preliminary study). Rapp. Procès-Verbaux Réun. Comm. Int. Explor. Sci. Mer Méditerr. 1979, 2526, 119–120. [Google Scholar]
- El Beshbeeshy, M.E. Descriptions of some thecate hydroids (Cnidaria-Hydrozoa) from the Egyptian Mediterranean waters. Part 3 family: Sertulariidae. Bull. Natl. Inst. Oceanogr. Fish. 1994, 20, 145–159. [Google Scholar]
- González-Duarte, M.M.; Megina, C.; Bethencourt, M. Sertularia marginata (Cnidaria: Hydrozoa) in the Mediterranea: An alien species in expansion? Mediterr. Mar. Sci. 2013, 14, 384–389. [Google Scholar] [CrossRef] [Green Version]
- Lakkis, S. Contribution à l’étude du zooplancton des eaux libanaises. Mar. Biol. Berl. 1971, 11, 138–148. [Google Scholar]
- Redier, L. Hydraires et bryozoaires de Méditerranée. II—Banyuls-sur-Mer. Cah. Nat. 1962, 18, 33–38. [Google Scholar]
- Medel, M.D.; Vervoort, W. Plumularian hydroids (Cnidaria: Hydrozoa) from the Strait of Gibraltar and nearly areas. Zool. Verh. Leiden 1995, 300, 1–72. [Google Scholar]
- Galil, B.S.; Gershwin, L.-A.; Zorea, M.; Rahav, A.; Rothman, S.B.-S.; Fine, M.; Lubinevsky, H.; Douek, J.; Paz, G.; Rinkevich, B. Cotylorhiza erythraea Stiasny, 1920 (Scyphozoa: Rhizostomeae: Cepheidae), yet another erythraean jellyfish from the Mediterranean coast of Israel. Mar. Biodiv. 2017, 47, 229–235. [Google Scholar] [CrossRef]
- Galil, B.S.; Gershwin, L.-A.; Douek, J.; Rinkevich, B. Marivagia stellata gen. et sp. nov. (Scyphozoa: Rhizostomeae: Cepheidae), another alien jellyfish from the Mediterranean coast of Israel. Aquat. Invasions 2010, 5, 331–340. [Google Scholar] [CrossRef]
- Zaatari, M. Odd Marine Changes Strike Coast of Sidon. The Daily Star Lebanon. 17 October 2010. Available online: https://www.dailystar.com.lb/News/Lebanon-News/2010/Oct-07/59848-odd-marine-changes-strike-coast-of-sidon.ashx (accessed on 17 October 2020).
- Bitar, G.; Badreddine, A. First record of Marivagia stellata Galil and Gershwin, 2010 (Scyphozoa: Rhizostomeae: Cepheidae) from the Lebanese waters in the eastern Mediterranean Sea. J. Black Sea/Mediterr. Environ. 2019, 25, 172–177. [Google Scholar]
- Mamish, S.; Durgham, H.; Al-Masri, M.S. First record of the new alien sea jelly species Marivagia stellata Galil and Gershwin, 2010 off the Syrian coast. Mar. Biodivers. Rec. 2016, 9, 23. [Google Scholar] [CrossRef] [Green Version]
- Hoeksema, B.W.; Ocaña Vicente, O. First record of the Central Indo-Pacific reef coral Oulastrea crispata in the Mediterranean Sea. Mediterr. Mar. Sci. 2014, 15, 429–436. [Google Scholar] [CrossRef] [Green Version]
- Bianchi, C.N. Proposta di suddivisione dei mari italiani in settori biogeografici. Notiziario SIBM 2004, 46, 57–59. [Google Scholar]
- Zibrowius, H. Oculina patagonica, scléractiniaire hermatypique introduit en Méditerranée. Helgol. Wiss. Meeresun. 1974, 26, 153–173. [Google Scholar] [CrossRef] [Green Version]
- Fine, M.; Zibrowius, H.; Loya, Y. Oculina patagonica: A non-lessepsian scleractinian coral invading the Mediterranean Sea. Mar. Biol. 2001, 138, 1195–1203. [Google Scholar] [CrossRef]
- Salomidi, M.; Katsanevakis, S.; Issaris, Y.; Tsiamis, K. Anthropogenic disturbance of coastal habitats promotes the spread of the introduced scleractinian coral Oculina patagonica in the Mediterranean Sea. Biol. Invasions 2013, 15, 1961–1971. [Google Scholar] [CrossRef]
- Serrano, E.; Coma, R.; Ribes, M. Pattern of Oculina patagonica occurrence along the Iberian peninsula coastline: A first step to understand the factors affecting its invasion dynamics. Rapp. Comm. Int. Mer Méditerr. 2013, 40, 603. [Google Scholar]
- Rubio-Portillo, E.; Vazquez, M.; Valle, C. Growth and bleaching of the coral Oculina patagonica under different environmental conditions in the western Mediterranean Sea. Mar. Biol. 2014, 161, 1333–2343. [Google Scholar] [CrossRef]
- Armoza-Zvuloni, R.; Segal, R.; Kramarsky-Winter, E.; Loya, Y. Repeated bleaching events may result in high tolerance and notable gametogenesis in stony corals: Oculina patagonica as a model. Mar. Ecol. Prog. Ser. 2011, 426, 149–159. [Google Scholar] [CrossRef]
- Coma, R.; Serrano, E.; Linares, C.; Ribes, M.; Díaz, D.; Ballesteros, E. Sea urchins predation facilitates coral invasion in a marine reserve. PLoS ONE 2011, 6, e22017. [Google Scholar] [CrossRef] [Green Version]
- Airi, V.; Gizzi, F.; Marchini, C.; Goffredo, S. Sexual reproduction of Mediterranean scleractinian corals. In The Cnidaria, Past, Present and Future; Goffredo, S., Dubinsky, Z., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 269–278. [Google Scholar] [CrossRef]
- Quintanilla, E.; Gili, J.M.; López-González, P.J.; Tsounis, G.; Madurell, T.; Fiorillo, I.; Rossi, S. Sexual reproductive cycle of the epibiotic soft coral, Alcyonium coralloides (Octocorallia, Alcyonacea), growing on the Mediterranean gorgonian Paramuricea clavata. Aquat. Biol. 2013, 18, 113–124. [Google Scholar] [CrossRef] [Green Version]
- Rossi, S.; Isla, E.; Bosch-Belmar, M.; Galli, G.; Gori, A.; Gristina, M.; Ingrosso, G.; Milisenda, G.; Piraino, S.; Rizzo, L.; et al. Changes of energy fluxes in the marine animal forest of the Anthropocene: Factors shaping the future seascape. ICES J. Mar. Sci. 2019, 76, 2008–2019. [Google Scholar] [CrossRef]
- Serrano, E.; Ribes, M.; Coma, R. Demographics of the zooxanthellate coral Oculina patagonica along the Mediterranean Iberian coast in relation to environmental parameters. Sci. Total Environ. 2018, 634, 1580–1592. [Google Scholar] [CrossRef]
- Bianchi, C.N.; Azzola, A.; Parravicini, V.; Peirano, A.; Morri, C.; Montefalcone, M. Abrupt change in a subtidal rocky reef community coincided with a rapid acceleration of sea water warming. Diversity 2019, 11, 215. [Google Scholar] [CrossRef] [Green Version]
- Dragičević, B.; Anadoli, O.; Angel, D.; Benabdi, M.; Bitar, G.; Castriota, L.; Crocetta, F.; Deidun, A.; Jakov, D.; Edelist, D.; et al. New Mediterranean Biodiversity Records (December 2019). Mediterr. Mar. Sci. 2019, 20, 645–656. [Google Scholar] [CrossRef] [Green Version]
- Ramil, F.; Vervoort, W. Report on the Hydroida collected by the “BALGIM” expedition in and around the Strait of Gibraltar. Zool. Verh. Leiden 1992, 277, 1–262. [Google Scholar]
- Bouillon, J.; Medel, M.D.; Pagès, F.; Gili, J.-M.; Boero, F.; Gravili, C. Fauna of the Mediterranean Hydrozoa. Sci. Mar. 2004, 68 (Suppl. 2), 5–438. [Google Scholar]
- Galil, B.S. Alien species in the Mediterranean Sea—Which, when, where, why? Hydrobiologia 2008, 606, 105–116. [Google Scholar] [CrossRef]
- UNEP/MAP/MED POL. Mariculture in the Mediterranean. MAP Tech. Rep. Ser. 2004, 140, 80. [Google Scholar]
- Azzurro, E.; Maynou, F.; Belmaker, J.; Golani, D.; Crooks, J.A. Lag times in Lessepsian fish invasion. Biol. Invasions 2016, 18, 2761–2772. [Google Scholar] [CrossRef]
- Zenetos, A.; Gratsia, E.; Cardoso, A.; Tsiamis, K. Time lags in reporting of biological invasions: The case of Mediterranean Sea. Mediterr. Mar. Sci. 2019, 20, 469–475. [Google Scholar] [CrossRef]
- EU. Council Regulation (EC) No 708/2007 of 11 June 2007 concerning use of alien and locally absent species in aquaculture. OJEU Off. J. Eur. Union 2007, 168, 1–17. [Google Scholar]
- Savini, D.; Occhipinti-Ambrogi, A.; Marchini, A.; Tricarico, E.; Gherardi, F.; Olenin, S.; Gollasch, S. The top 27 animal alien species introduced into Europe for aquaculture and related activities. J. Appl. Ichthyol. 2010, 26, 1–7. [Google Scholar] [CrossRef]
- MEPC. Guidelines for the Control and Management of Ships’ Biofouling to Minimize the Transfer of Invasive Aquatic Species; Annex 26. Resolution MEPC.207(62); Marine Environment Protection Committee: London, UK, 15 July 2011; 25p. [Google Scholar]
- Mineur, F.; Johnson, M.P.; Maggs, C.A. Macroalgal introductions by hull fouling on recreational vessels: Seaweeds and sailors. Environ. Manag. 2008, 42, 667–676. [Google Scholar] [CrossRef]
- Murray, C.C.; Pakhomov, E.A.; Therriault, T.W. Recreational boating: A large unregulated vector transporting marine invasive species. Divers. Distrib. 2011, 17, 1161–1172. [Google Scholar] [CrossRef]
- Gravili, C.; Di Camillo, C.G.; Piraino, S.; Boero, F. Hydrozoan species richness in the Mediterranean Sea: Past and present. Mar. Ecol. 2013, 34 (Suppl. 1), 41–62. [Google Scholar] [CrossRef]
- Foster, V.; Giesler, R.J.; Wilson, A.M.W.; Nall, C.R.; Cook, E.J. Identifying the physical features of marina infrastructure associated with the presence of non-native species in the UK. Mar. Biol. 2016, 163, 173. [Google Scholar] [CrossRef] [Green Version]
- Buonocore, E.; Franzese, P.P.; Russo, G.F. Marine ecosystem services: Understanding the value of nature. Water 2021. In press. [Google Scholar]
- González-Duarte, M.M.; Megina, C.; López-González, P.J.; Galil, B. Cnidarian alien species in expansion. In The Cnidaria, Past, Present and Future. The world of Medusa and her sisters; Goffredo, S., Dubinsky, Z., Eds.; Springer: Cham, Switzerland, 2016; pp. 139–160. [Google Scholar]
- Moura, C.J.; Cuhna, M.R.; Porteiro, F.M.; Rogers, A.D. A molecular phylogenetic appraisal of the systematics of the Aglaopheniidae (Cnidaria: Hydrozoa, Leptothecata) from the north-east Atlantic and west Mediterranean. Zool. J. Linnean Soc. 2012, 164, 717–727. [Google Scholar] [CrossRef] [Green Version]
- Byers, J.E.; Noonburg, E.G. Scale dependent effects of biotic resistance to biological invasion. Ecology 2003, 84, 1428–1433. [Google Scholar] [CrossRef]
- Corriero, G.; Pierri, C.; Accoroni, S.; Alabiso, G.; Bavestrello, G.; Barbone, E.; Bastianini, M.; Bazzoni, A.M.; Bernardi Aubry, F.; Boero, F.; et al. Ecosystem vulnerability to alien and invasive species: A case study on marine habitats along the Italian coast. Aquat. Conserv. Mar. Freshw. Ecosyst. 2015, 26, 392–409. [Google Scholar] [CrossRef]
- Levine, J.M. Species diversity and biological invasions: Relating local process to community pattern. Science 2000, 288, 852–854. [Google Scholar] [CrossRef] [Green Version]
- Shea, K.; Chesson, P. Community ecology theory as a framework for biological invasions. Trends Ecol. Evol. 2002, 17, 170–176. [Google Scholar] [CrossRef]
- Thieltges, D.W.; Strasser, M.; Reise, K. How bad are invaders in coastal waters? The case of the American slipper limpet Crepidula fornicata in western Europe. Biol. Invasions 2006, 8, 1673–1680. [Google Scholar] [CrossRef]
- Schlaepfer, M.A.; Sax, D.F.; Olden, J.D. The potential conservation value of non-native species. Conserv. Biol. 2011, 25, 428–437. [Google Scholar] [CrossRef] [PubMed]
- McLaughlan, C.; Gallardo, B.; Aldridge, D.C. How complete is our knowledge of the ecosystem services impacts of Europe’s top 10 invasive species? Acta Oecol. 2013, 54, 119–130. [Google Scholar] [CrossRef]
- Thomsen, M.S.; Byers, J.E.; Schiel, D.R.; Bruno, J.F.; Olden, J.D.; Wernberg, T.; Silliman, B.R. Impacts of marine invaders on biodiversity depend on trophic position and functional similarity. Mar. Ecol. Prog. Ser. 2014, 495, 39–47. [Google Scholar] [CrossRef] [Green Version]
- Pimentel, D.; Lach, L.; Zuniga, R.; Morrison, D. Environmental and economic costs of nonindigenous species in the United States. BioScience 2000, 50, 53–65. [Google Scholar] [CrossRef]
- Stachowicz, J.J.; Byrnes, J.E. Species diversity, invasion success, and ecosystem functioning: Disentangling the influence of resource competition, facilitation, and extrinsic factors. Mar. Ecol. Prog. Ser. 2006, 311, 251–262. [Google Scholar] [CrossRef]
- Galil, B.S. Loss or gain? Invasive aliens and biodiversity in the Mediterranean Sea. Mar. Pollut. Bull. 2007, 55, 314–322. [Google Scholar] [CrossRef]
- Thomsen, M.S.; Olden, J.D.; Wernberg, T.; Griffin, J.N.; Silliman, B.R. A broad framework to organize and compare ecological invasion impacts. Environ. Res. 2011, 111, 899–908. [Google Scholar] [CrossRef]
- Perrings, C.; Williamson, M.; Barbier, E.B.; Delfino, D.; Dalmazzone, S.; Shogren, J.; Simmons, P.; Watkinson, A. Biological invasion risks and the public good: An economic perspective. Conserv. Ecol. 2002, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- Dick, J.T.A.; Alexander, M.E.; Jeschke, J.M.; Ricciardi, A.; MacIsaac, H.J.; Robinson, T.B.; Kumschick, S.; Weyl, O.L.F.; Dunn, A.M.; Hatcher, M.J.; et al. Advancing impact prediction and hypothesis testing in invasion ecology using a comparative functional response approach. Biol. Invasions 2014, 16, 735–753. [Google Scholar] [CrossRef] [Green Version]
- Ojaveer, H.; Galil, B.S.; Campbell, M.L.; Carlton, J.T.; Canning-Clode, J.; Cook, E.J.; Davidson, A.D.; Hewitt, C.L.; Jelmert, A.; Marchini, A.; et al. Classification of non-indigenous species based on their impacts: Considerations for application in marine management. PLoS Biol. 2015, 13, e1002130. [Google Scholar] [CrossRef] [Green Version]
- Giakoumi, S.; Katsanevakis, S.; Albano, P.G.; Azzurro, E.; Cardoso, A.C.; Cebrian, E.; Deidun, A.; Edelist, D.; Francour, P.; Jimenez, C.; et al. Management priorities for marine invasive species. Sci. Total Environ. 2019, 688, 976–982. [Google Scholar] [CrossRef]
- Giangrande, A.; Pierri, C.; Del Pasqua, M.; Gravili, C.; Gambi, M.C.; Gravina, M.F. The Mediterranean in check: Biological invasions in a changing sea. Mar. Ecol. 2020, 41, e12583. [Google Scholar] [CrossRef]
- Davis, M.A.; Grime, J.P.; Thompson, K. Fluctuating resources in plant communities: A general theory of invisibility. J. Ecol. 2000, 88, 528–534. [Google Scholar] [CrossRef] [Green Version]
- Zaiko, A.; Olenin, S.; Daunys, D.; Nalepa, T. Vulnerability of benthic habitats to the aquatic invasive species. Biol. Invasions 2007, 9, 703–714. [Google Scholar] [CrossRef]
- Paolucci, E.M.; MacIsaac, K.J.; Ricciardi, A. Origin matters: Alien consumers inflict greater damage on prey populations than do native consumers. Divers. Distrib. 2013, 19, 988–995. [Google Scholar] [CrossRef]
- Simberloff, D. Biological invasions: What’s worth fighting and what can be won? Ecol. Eng. 2014, 65, 112–121. [Google Scholar] [CrossRef]
- Carosi, A.; Ghetti, L.; Lorenzoni, M. The role of climate changes in the spread of freshwater fishes: Implications for alien cool and warm-water species in a Mediterranean basin. Water 2021, 13, 347. [Google Scholar] [CrossRef]
- Ceccherelli, G.; Campo, D. Different effects of Caulerpa racemosa on two co-occurring seagrasses in the Mediterranean. Bot. Mar. 2002, 45, 71–76. [Google Scholar] [CrossRef]
- Grosholz, E. Ecological and evolutionary consequences of coastal invasions. Trends Ecol. Evol. 2002, 17, 22–27. [Google Scholar] [CrossRef]
- Arenas, F.; Sanchez, I.; Hawkins, S.; Jenkins, S.R. The invasibility of marine algal assemblages: Role of functional diversity and identity. Ecology 2006, 87, 2851–2861. [Google Scholar] [CrossRef]
- Ricciardi, A.; Cohen, J. The invasiveness of an introduced species does not predict its impact. Biol. Invasions 2007, 9, 309–315. [Google Scholar] [CrossRef]
- Kumschick, S.; Gaertner, M.; Vilà, M.; Essl, F.; Jeschke, J.M.; Pyšek, P.; Ricciardi, A.; Bacher, S.; Blackburn, T.M.; Dick, J.T.A.; et al. Ecological impacts of alien species: Quantification, scope, caveats, and recommendations. Bioscience 2015, 65, 55–63. [Google Scholar] [CrossRef]
- Zwerschke, N.; Hollyman, P.R.; Wild, R.; Strigner, R.; Turner, J.R.; King, J.W. Limited impact of an invasive oyster on intertidal assemblage structure and biodiversity: The importance of environmental context and functional equivalency with native species. Mar. Biol. 2018, 165, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonanno, G.; Orlando-Bonaca, M. Non-indigenous marine species in the Mediterranean Sea—Myth and reality. Environ. Sci. Policy 2019, 96, 123–131. [Google Scholar] [CrossRef]
- Sagoff, M. Do non-native species threaten the natural environment? J. Agric. Environ. Ethics 2005, 18, 215–236. [Google Scholar] [CrossRef]
- Walther, G.R.; Roques, A.; Hulme, P.E.; Sykes, M.T.; Pyšek, P.; Kühn, I.; Zobel, M.; Bacher, S.; Botta-Dukát, Z.; Bugmann, H.; et al. Alien species in a warmer world: Risks and opportunities. Trends Ecol. Evol. 2009, 24, 686–693. [Google Scholar] [CrossRef] [Green Version]
- Davis, M.A.; Chew, M.K.; Hobbs, R.J.; Lugo, A.E.; Ewel, J.J.; Vermeij, G.J.; Brown, J.H.; Rosenzweig, M.L.; Gardener, M.R.; Carroll, S.P.; et al. Don’t judge species on their origins. Nature 2011, 474, 153–154. [Google Scholar] [CrossRef]
- Galil, B.S. Taking stock: Inventory of alien species in the Mediterranean Sea. Biol. Invasions 2009, 11, 359–372. [Google Scholar] [CrossRef]
- Galil, B.S. The alien crustaceans in the Mediterranean Sea: An historical review. In In the Wrong Place—Alien Marine Crustaceans: Distribution, Biology and Impacts; Galil, B.S., Clark, P.F., Carlton, J.T., Eds.; Springer: Cham, Netherlands, 2011; pp. 377–401. [Google Scholar]
- Sala, E.; Kizilkaya, Z.; Yildirim, D.; Ballesteros, E. Alien marine fishes deplete algal biomass in the eastern Mediterranean. PLoS ONE 2011, 6, e17356. [Google Scholar] [CrossRef] [Green Version]
- Killi, N.; Tarkan, A.S.; Kozic, S.; Copp, G.H.; Davison, P.I.; Vilizzi, L. Risk screening of the potential invasiveness of non-native jellyfishes in the Mediterranean Sea. Mar. Pollut. Bull. 2020, 150, 110728. [Google Scholar] [CrossRef]
- Ekebom, J. The long and winding road of the ecosystem approach into marine environmental policies. Aquat. Conserv. Mar. Freshw. Ecosyst 2013, 23, 1–6. [Google Scholar] [CrossRef]
Figure 1.
Proportion of the major native distribution ranges of CC NIS (following Spalding et al. [42] for biogeographic realms classifications) in the Mediterranean subregions. The size of each pie chart represents the total number of NIS primarily introduced in a subregion.
Figure 1.
Proportion of the major native distribution ranges of CC NIS (following Spalding et al. [42] for biogeographic realms classifications) in the Mediterranean subregions. The size of each pie chart represents the total number of NIS primarily introduced in a subregion.

Figure 2.
Number of CC NIS introduced in the Mediterranean Sea through primary introduction pathways, based on CBD [44] scheme associated with each of five major realms of Mediterranean CC NIS’s native distribution: (a) Western Indo-Pacific, (b) Central Indo-Pacific, (c) Temperate Australasia, (d) Tropical Atlantic, (e) Temperate Northern Atlantic. Several species are linked to more than one pathway.
Figure 2.
Number of CC NIS introduced in the Mediterranean Sea through primary introduction pathways, based on CBD [44] scheme associated with each of five major realms of Mediterranean CC NIS’s native distribution: (a) Western Indo-Pacific, (b) Central Indo-Pacific, (c) Temperate Australasia, (d) Tropical Atlantic, (e) Temperate Northern Atlantic. Several species are linked to more than one pathway.

Figure 3.
Temporal trends of new marine CC NIS introductions in Europe per 5-year intervals, whose native distribution corresponds to at least one of the five most important realms of Mediterranean CC NIS’ native distribution ranges for the time period 1950–2014 (dashed lines: mobile averages over 3-year period).
Figure 3.
Temporal trends of new marine CC NIS introductions in Europe per 5-year intervals, whose native distribution corresponds to at least one of the five most important realms of Mediterranean CC NIS’ native distribution ranges for the time period 1950–2014 (dashed lines: mobile averages over 3-year period).

Figure 4.
Distribution of CC NIS in each biogeographic area within the Italian Seas following Bianchi [93]. (A) Ligurian Sea; (B) central Tyrrhenian Sea; (C) south Tyrrhenian Sea; (D) Strait of Messina; (E) south-eastern tip of Sicily, Pelagie Islands; (F) Ionian Sea; (G) south Adriatic Sea; (H) central Adriatic Sea; (I) north Adriatic Sea. For each sector, CC NIS percentage and CC NIS number/Cnidaria and Ctenophora total number are shown.
Figure 4.
Distribution of CC NIS in each biogeographic area within the Italian Seas following Bianchi [93]. (A) Ligurian Sea; (B) central Tyrrhenian Sea; (C) south Tyrrhenian Sea; (D) Strait of Messina; (E) south-eastern tip of Sicily, Pelagie Islands; (F) Ionian Sea; (G) south Adriatic Sea; (H) central Adriatic Sea; (I) north Adriatic Sea. For each sector, CC NIS percentage and CC NIS number/Cnidaria and Ctenophora total number are shown.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Cnidaria potential invaders of the Italian waters.
| Taxa | Origin | Main Way of Introduction | Distribution in Mediterranean Sea and Records | Last Record
(Locality and Year) | Probable Invasiveness |
|---|---|---|---|---|---|
| Hydrozoa | |||||
| Dynamena quadridentata (Ellis & Solander, 1786) | Tropical Atlantic | Suez Canal (Lessepsian species) | Levant Sea [50] | Lebanon coast (2004) | ++ |
| Eucheilota paradoxica Mayer, 1900 | Tropical Atlantic | Unknown | Croatian coast [51], Alborán Sea [52], Levant Sea [53,54,55], French coast [56,57], Tunisian coast [58,59], Algerian coast [60] | Tunisian coast (2015) | ++ |
| Eucheilota ventricularis McCrady, 1859 | North temperate Atlantic | Suez Canal (Lessepsian species) | Levant Sea [53,54,55,61] | Lebanon coast (1989) | + |
| Haliscera bigelowi Kramp, 1947 | Tropical eastern Pacific | Unknown | Croatian coast [51,62], Alborán Sea [63], Spanish coast [64,65,66,67,68], Tunisian coast [59,69] | Tunisian coast (2015) | ++ |
| Macrorhynchia philippina Kirchenpauer, 1872 | Central Indo- Pacific | Suez Canal (Lessepsian species) | Lebanon coast [50,70,71], Levantine coast of Turkey [72], Suez Canal [22], Aegean Sea [73] | Suez Canal (2014) | +++ |
| Moerisia carine Bouillon, 1978 | Central Indo- Pacific | Suez Canal (Lessepsian species) | Levant Sea [53,54,55] | Lebanon coast (1989) | + |
| Olindias singularis Browne, 1905 | Central Indo- Pacific | Suez Canal (Lessepsian species) | Egyptian Mediterranean coast [74] | Egyptian Mediterranean coast (extending from Alexandria in the east to Sidi Barani in the west) (2001) | ++ |
| Plumularia pulchella Bale, 1882 | Temperate Australasia | Unknown | Alborán Sea [65], Spanish coast [75,76,77], Egyptian Mediterranean waters [78] | Egyptian Mediterranean waters (1977) | + |
| Sertularia marginata (Kirchenpauer, 1864) | Western Indo- Pacific | Suez Canal (Lessepsian species) | Levant Sea [50,79,80,81,82], Alborán Sea [83] | Alborán Sea (2012) | +++ |
| Sertularia tongensis (Stechow, 1919) as S. thecocarpa | Western Indo- Pacific | Suez Canal (Lessepsian species) | Levant Sea [50] | Lebanon coast (2004) | ++ |
| Tetrorchis erythrogaster Bigelow, 1909 | Tropical eastern Pacific | Unknown | Beyrouth [53,54,55,61,84] | Lebanon coast (1982) | + |
| Gymnangium montagui (Billard, 1912) | Temperate North Atlantic | Espansion natural range | French coast [85], Alborán Sea [86] | Strait of Gibraltar and nearly areas (1993) | ++ |
| Scyphozoa | |||||
| Cotylorhiza erythraea Stiasny, 1920 | Western Indo- Pacific | Suez Canal (Lessepsian species) | Levant Sea [87] | Israel coast (2015) | +++ |
| Marivagia stellata Galil & Gershwin, 2010 | Western Indo- Pacific | Shipping (ballast waters) (Lessepsian species) | Levant Sea [88,89,90,91] | Syrian coast (2015), Lebanese waters (2015) | +++ |
| Anthozoa | |||||
| Oulastrea crispata (Lamarck, 1816) | Central Indo- Pacific | Unknown | Corsica [92] | Corsica (2014) | ++ |
+: low; ++: medium; +++: high.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gravili, C.; Rossi, S. Who’s Next? Non-Indigenous Cnidarian and Ctenophoran Species Approaching to the Italian Waters. Water 2021, 13, 1062. https://doi.org/10.3390/w13081062
AMA Style
Gravili C, Rossi S. Who’s Next? Non-Indigenous Cnidarian and Ctenophoran Species Approaching to the Italian Waters. Water. 2021; 13(8):1062. https://doi.org/10.3390/w13081062
Chicago/Turabian StyleGravili, Cinzia, and Sergio Rossi. 2021. "Who’s Next? Non-Indigenous Cnidarian and Ctenophoran Species Approaching to the Italian Waters" Water 13, no. 8: 1062. https://doi.org/10.3390/w13081062
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.