
Comparison of Invasive Apple Snail (Pomacea canaliculata) Behaviors in Different Water Temperature Gradients


1
Nakdonggang National Institute of Biological Resources, Sangju 37242, Korea
2
Department of Biology, Kyung Hee University, Dongdaemun, Seoul 02447, Korea
*
Author to whom correspondence should be addressed.
Water 2021, 13(9), 1149; https://doi.org/10.3390/w13091149
Submission received: 31 December 2020
/
Revised: 19 April 2021
/
Accepted: 19 April 2021
/
Published: 22 April 2021
(This article belongs to the Section Biodiversity and Functionality of Aquatic Ecosystems)
Abstract
:Pomacea canaliculata (known as invasive apple snail) is a freshwater snail native to South America that was introduced into many countries (including Asia and North America) as a food source or for organic farming systems. However, it has invaded freshwater ecosystems and become a serious agricultural pest in paddy fields. Water temperature is an important factor determining behavior and successful establishment in new areas. We examined the behavioral responses of P. canaliculata with water temperature changes from 25 °C to 30 °C, 20 °C, and 15 °C by quantifying changes in nine behaviors. At the acclimated temperature (25 °C), the mobility of P. canaliculata was low during the day, but high at night. Clinging behavior increased as the water temperature decreased from 25 °C to 20 °C or 15 °C. Conversely, ventilation and food consumption increased when the water temperature increased from 25 °C to 30 °C. A self-organizing map (an unsupervised artificial neural network) was used to classify the behavioral patterns into seven clusters at different water temperatures. These results suggest that the activity levels or certain behaviors of P. canaliculata vary with the water temperature conditions. Understanding the thermal biology of P. canaliculata may be crucial for managing this invasive snail.
1. Introduction
The apple snail, Poamacea canaliculata (Gastropoda: Ampullariidae), is native to the southern parts of South America [1,2,3]. Although this snail was initially introduced into Asian countries as human food, the venture was unsuccessful. Many snails escaped from snail farms and invaded rice paddy fields and their neighboring freshwater ecosystems. These invasive apple snails, which are especially hazardous for young rice plants, have become serious agricultural pests globally [4,5,6,7], causing major ecological damage, such as the depletion of macrophytes in natural wetlands [8]. Hence, the apple snail is listed on the 100 worst invaders worldwide in the International Union for the Conservation of Nature’s list [9].
The Pomacea species has superior biological and ecological traits which make it a successful invader, such as high growth and reproductive rates, polyphagous feeding habits, mass propagation (e.g., egg spawning), high emergence rate (95.8%) and amphibious respiration and aestivating capacity, which make it highly adaptable and resistant to diverse environmental conditions when compared with native apple snails (e.g., Cipangopaludina chinensis malleata) [10,11,12,13,14] in freshwater ecosystems. However, their traits such as growth rate (e.g., [15]), reproductive maturity (e.g., [16]), spawning (e.g., [17]), and behavior (e.g., [18,19]) are strongly influenced by water temperature [18,19]. Any behavior depends on an organism’s responses to external (environmental) as well as internal (physiological) factors. Therefore, measuring behavioral changes in apple snails helps us understand their ecology in different environmental conditions. For example, apple snails do not produce eggs above 35 °C and do not exhibit reproductive activity below 15 °C [20]. The minimal temperature for activity is 10 °C [18]. Since apple snails are tolerant of water temperatures between 2 °C and 38 °C [21], they may expand their habitat and colonize new freshwater ecosystems with global warming [22,23]. According to Lei et al. [24], their suitable habitat may potentially increase by 3.8% in the 2080s based on the Representative Concentration Pathway (RCP) 2.6 scenario of the Fifth Assessment Report (AR5) of the Intergovernmental Panel on Climate Change (IPCC). Their habitat has the potential to increase by 10.3% based on the RCP 8.5 scenario, indicating that global warming will enable them to enlarge their invasion area, threatening the survival of native apple snails which are weaker competitors. Previous studies [13,18,19,20,21,22,25] have focused on the differences in activity levels (or behaviors) of apple snails across temperatures they are already acclimated to. Therefore, we aimed to determine the influence of changing water temperatures (i.e., increase or decrease from the acclimated temperature) on the behavior of the invasive apple snail.
2. Materials and Methods
2.1. Test Organisms and Behavior Observation
Specimens of apple snail (P. canaliculata) (about 5 kg of adult individuals) were obtained from an apple snail farm (37°48′10″ N, 127°29′40″ E). The stock populations were maintained in an aquarium with dechlorinated tap water at a temperature of 25 ± 1 °C, a light/dark cycle of 16:8 h, and a neutral pH condition ranging from 7.3 to 7.6 for at least 2 months prior to the experiment [26]. Three days before starting the experiment, the specimens were put in the experimental system at the same acclimation temperature to exclude displacement stress from the behavioral analyses. To further reduce irregular behavior (e.g., spawning and mating), only male specimens were used in the experiment [13,25]. Males were identified by the visible testicle (through the translucent shell) [27] or a humped operculum [28]. Sexually undifferentiated snails (smaller than 25 mm) were excluded from the experiment [18,25].
The behavior of the apple snails was observed in a test aquarium (30 × 30 cm with 15 cm water depth) for two days. During the experiment, the water temperature was kept constant at 25 °C (same as the rearing temperature) for the first 2 h and then changed (1 °C/h) to the desired temperature (15, 20, or 30 °C). When the desired temperature was reached, the target temperature was constantly maintained for the remainder of the experimental period. The experimental aquarium was supplied with sediment (3 cm depth of sand), similar to the rearing condition. The rearing experience identified an appropriate diet. We provided a pre-determined amount of lettuce (Lactuca sativa L.) as the food source during the experiment. The lettuce was replaced once a day (i.e., every 24 h) in controlled experimental conditions (25 °C). The same volume was provided in the water-temperature-decreased conditions (from 25 °C to 15 °C and from 25 °C to 20 °C). The food source was provided twice a day (i.e., every 12 h) in the water-temperature-increased conditions (from 25 °C to 30 °C) due to the increase in activity and metabolism (see Result section).
All snails were marked with synthetic polish on the shell to identify the individual specimens in each treatment. Four test specimens were placed in an observation aquarium, and the behaviors of the apple snails were recorded using a web-cam (C905, Logitech, Newark, CA, USA) and checked every ten minutes for two days. New snail specimens were used in each experimental treatment. We identified nine categories of behaviors in the recorded video: floating (FO), clinging to the side of the aquarium (CL), crawling on the bottom of the aquarium (CB), crawling on the side of the aquarium (CS), feeding (FE), folding a funnel with its food (FF), swimming (SW), ventilating the lung (VE), and falling (FA) [13,19]. Based on other research, we categorized the behaviors into inactive (FO and CL) and active (CB, CS, FE, FF, SW, VE and FA) [14,15]. In total, we observed the behaviors of 16 specimens at each temperature. Dead apple snails were removed daily every morning.
2.2. Feeding
Digital image processing was used evaluate the food consumption by measuring the difference in lettuce leaf area before and after the experiment (Figure 1). We placed lettuce leaves on four acrylic plates (7 × 15 cm) and placed them on the bottom of the observation aquarium. The acrylic plates were fixed in the aquarium during the experiment. After the lettuce was exchanged, digital images of the remaining lettuce were taken, and processed by changing the images to white (fed area) and black (unfed area) using a threshold method in MATLAB ver. 7.2 [29]. The numbers of pixels in the black and white areas were measured to calculate the amount of food consumed during the experiment.
2.3. Data Analysis
We analyzed the behavioral data in four steps. First, to interpret the overall behavioral differences among different temperature conditions, we used the Kruskal–Wallis (K-W) test [30], followed by a Dunn’s multiple comparison test to identify significant differences among the temperature conditions (at the level of p < 0.05). Second, a Mann–Whitney (M–W) U test was applied to compare the behavioral differences between day and night in each temperature condition as these snails are nocturnal. Third, a M–W U test was used to compare the food consumption between the first and second experimental days in each of the water temperature treatments. We expected the food consumption between the first and second days would vary because the first day included temperature changes (ramping up or down to treatment conditions) whereas the second day had a constant temperature. Fourth, we used a self-organizing map (SOM) [31,32] to identify differences in the patterns of the nine behavioral types (i.e., CL, CB, CS, FE, FF, SW, FO, VE, and FA) exhibited by apple snails when exposed to changes in water temperature.
SOM is an unsupervised learning algorithm of artificial neural networks consisting of input and output layers connected with computational weights [31,32,33,34]. The SOM trains the input data by updating the weights based on the connection intensity between the input and output layers. When the input data (nine categories of behaviors in this study) were initially assigned to the network, each output neuron calculated the distance between the weight vector and the input vector. The number of output neurons was set to 150 (15 × 10) in a two-dimensional hexagonal grid based on our preliminary studies. Among the 150 output neurons, the best matching neuron (BMU) that had the minimum distance between the weight and the input neuron was considered the winner. The weight vectors were updated for the BMU and its neighborhood units according to the SOM learning rule. The input vectors were classified according to the closest weight vectors, using the SOM learning rule. We applied these functions in the SOM toolbox (http://www.cis.hut.fi/projects/somtoolbox/ (accessed on 05 October 2020)) in MATLAB ver. 7.2 [29]. After the SOM learning process, the SOM units were classified using the U-matrix and the Ward linkage method based on the Euclidean distance [35]. The multi-response permutation procedure (MRPP) was applied to test whether there were significant differences among clusters in the SOM using PC-ORD for Windows ver. 5.0 [36]. Then, behavioral differences among clusters in the SOM results were compared using K-W and then Dunn’s multiple comparison tests.
All statistical analyses (i.e., K-W, Dunn’s multiple comparison test, and M–W U test) were performed using STATISTICA for Windows ver. 7 [37].
3. Results
3.1. Comparion of Behavioral Differences in Different Temperature Conditions
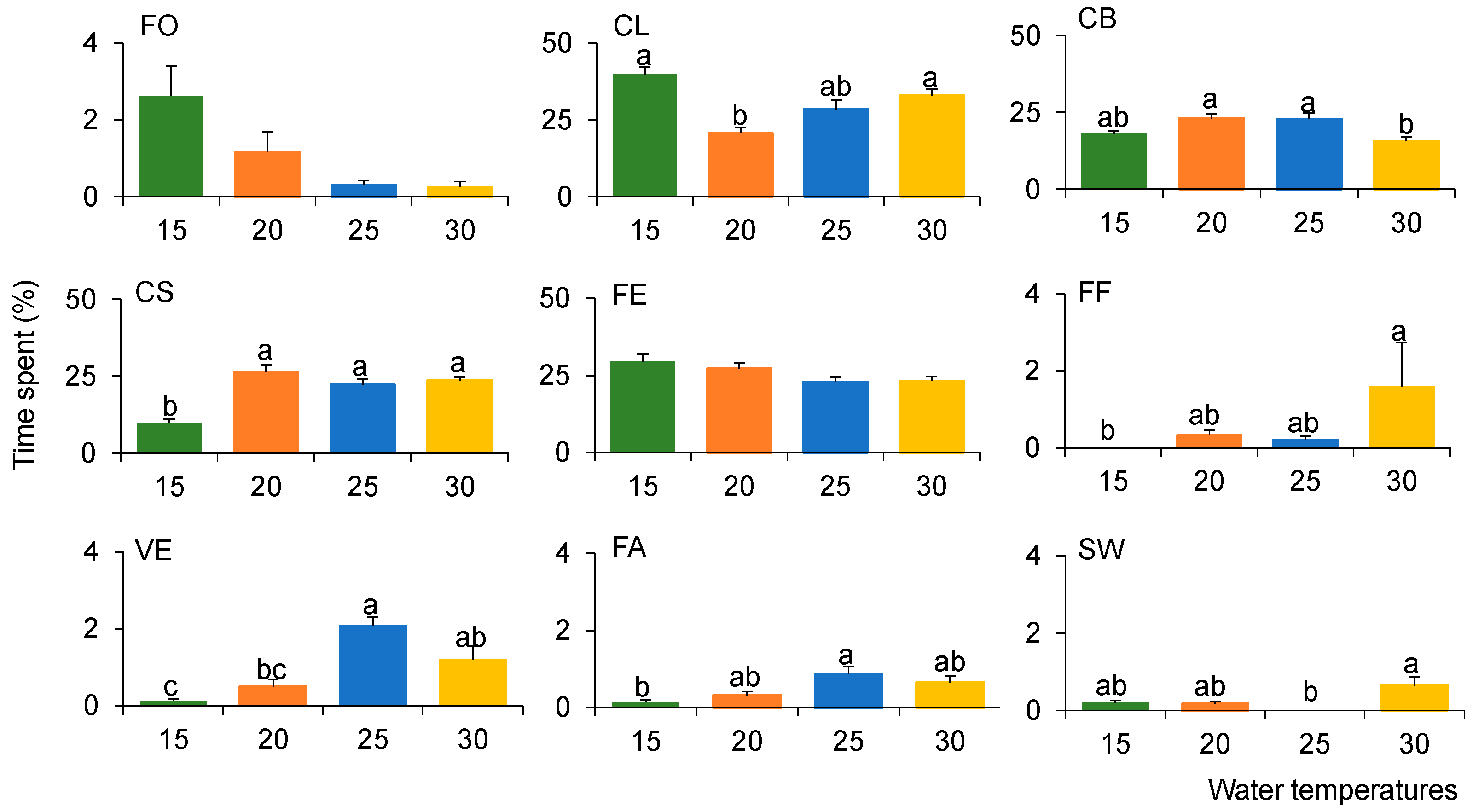
Among the nine behaviors, CL, CB, CS and FE were the dominant behaviors observed in all temperature treatments (Figure S1). When combined, they represented 95.7% (25 °C to 30 °C) to 97.5% (25 °C to 20 °C) of the observed behaviors (Figure 2).
Apple snails spent the highest proportion of time in CL in the 25 °C to 15 °C treatment (38.4%, K-W test, p < 0.05), while they exhibited the lowest amount of time in CS (14.7%, K-W test, p < 0.05). On the other hand, apple snails spent the highest amount of time in FF under the temperature condition from 25 °C to 30 °C treatment (K-W test, p < 0.05). Even though the time spent in FE and FO was not significantly different across the different temperature conditions, FO (2.6%) was highest in the temperature condition of the 25 °C to 15 °C treatment. The snails also spent more time in VE (2.1%) and FA (0.9%) in the control treatment (K-W test, p < 0.05).
3.2. Temporal Patterns of Behaviors in Different Temperature Regimes
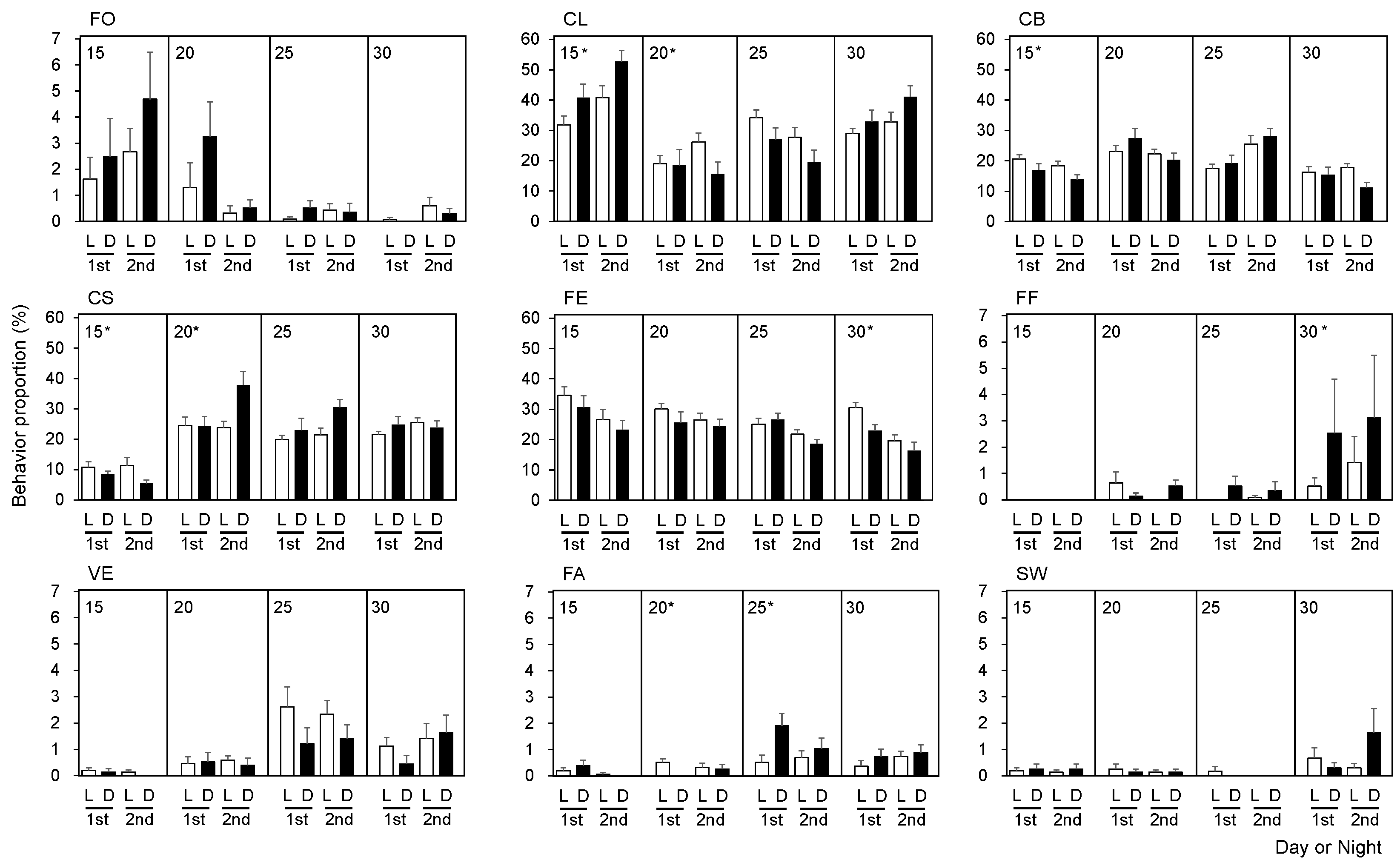
Overall, the proportion of inactive behaviors such as FO and CL increased in the 25 °C to 15 °C treatment, although there was no significant difference in FO (Figure 3). CL in the control treatment (25 °C) was relatively higher during the light periods compared with the dark periods. Conversely, CL was greater in dark periods in the 25 °C to 15 °C treatment (M–W U test, p < 0.05). The proportion of CB and CS was higher during the light periods in the 25 °C to 15 °C treatment (M–W U test, p < 0.05). The proportion of CS was higher during the dark periods in the 25 °C to 20 °C treatment and the control experiment (M–W U test, p < 0.05). The proportion of FE decreased over time in the 25 °C to 30 °C treatment (M–W U test, p < 0.05). On the other hand, the proportion of another feeding behavior (i.e., FF) was higher (especially at night) in the 25 °C to 30 °C treatment than in all other treatments.
3.3. Food Consumption
The apple snails consumed 76.8% of the food in the total experimental period in the control treatment (Table 1). Food consumption was significantly lower in decreasing temperature conditions (i.e., 25 °C to 20 °C and 25 °C to 15 °C), contrary to the time spent feeding (K-W, p < 0.05). While the amount of food consumed was not significantly different between the experimental days 1 and 2 in the control treatment and the 25 °C to 30 °C (M–W U test, p < 0.05). The food consumption was lower on the second day in the 25 °C to 15 °C treatment (M–W U test, p < 0.05).
3.4. Characterization of Behavioral Patterns
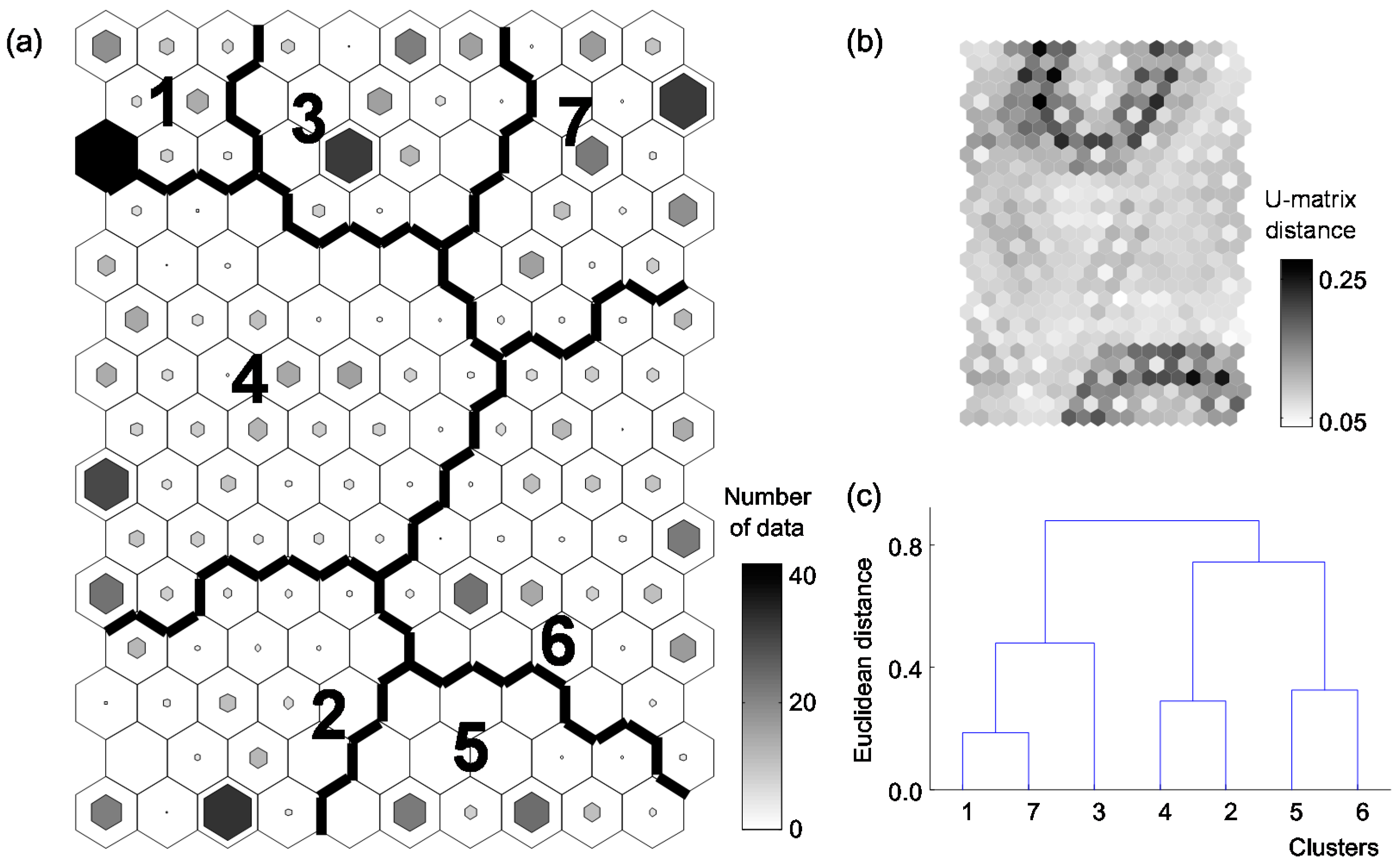
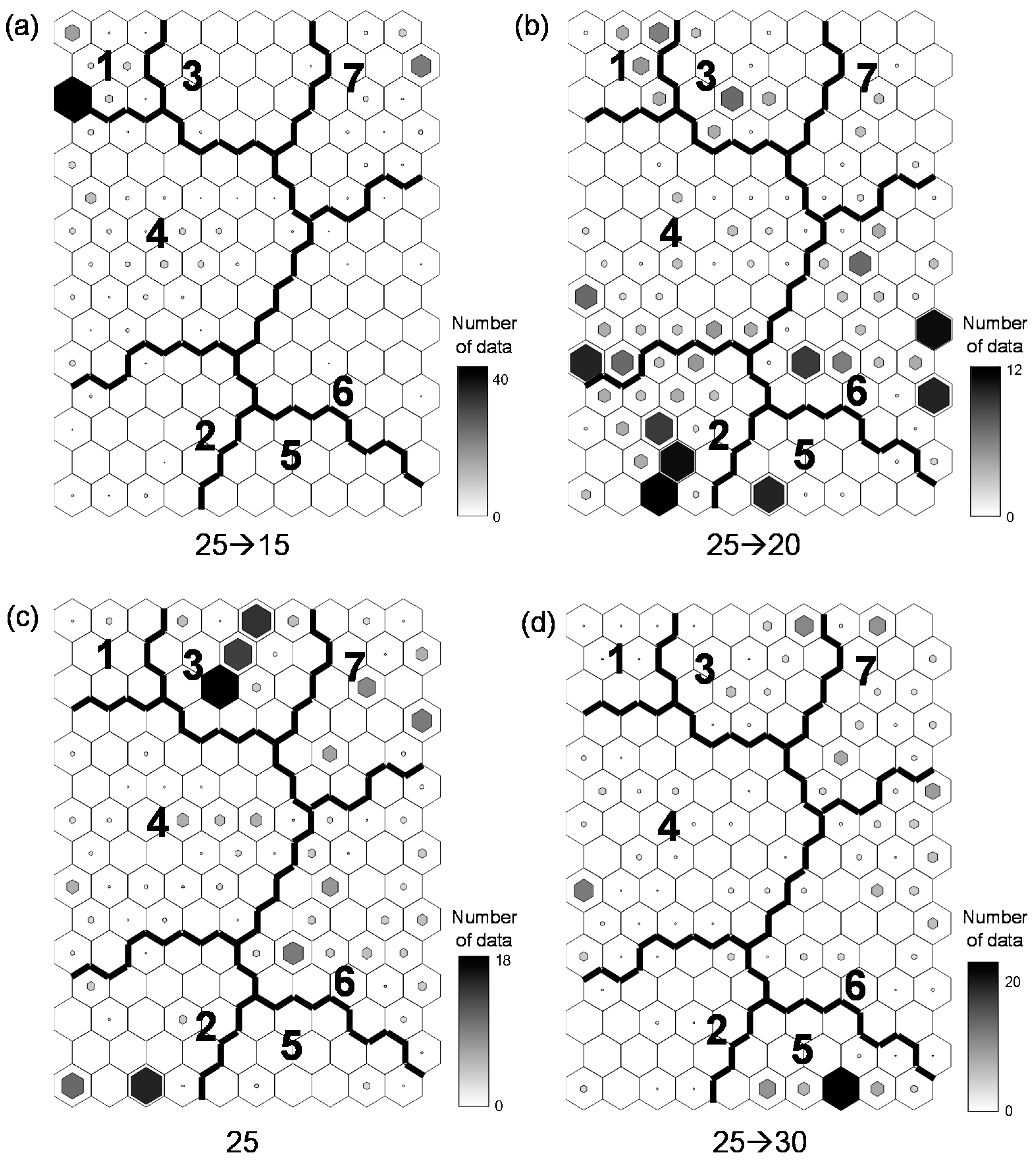
Behavioral responses to water temperature differences were characterized using the similarities in behavioral categories with the SOM results (Figure 4). This analysis revealed differences in behavioral responses to different changes in temperature. The SOM output units were classified into seven clusters based on a U-matrix and hierarchical cluster analysis using the Ward linkage method with Euclidean distance measures. The MRPP showed significant differences among the clusters (A = 0.26, p < 0.001). The behavioral data in the upper left area (clusters 1 and 4) responded to decreasing temperatures ranging from 25 °C to 15 °C (Figure 5a). The behavioral data of temperature changes from 25 °C to 20 °C were located in the lower area (clusters 2 and 6, especially) (Figure 5b). Even though the control data (25 °C) tended to be scattered in the SOM map (except cluster 1), most of the data were located in cluster 3 (Figure 5c). The behavioral data from increasing temperature (25 °C to 30 °C) were located mainly in the right-side area of the SOM map (clusters 5, 6, and 7) (Figure 5d).
All behavioral categories (except SW and FA) were significantly different among the clusters (Table 2). FO (7.8%) was the highest in cluster 1 whereas CS were the lowest in cluster 1 (12.0%) and cluster 4 (13.3%). CB and CS were highest in cluster 2 (35.6%) and cluster 6 (37.7%), respectively. FF was highest in cluster 5 (7.6%) and VE (8.3%) was high in cluster 3. CL was the highest in cluster 7 (52.4%) whereas crawling on the bottom of the aquarium (14.1%) and feeding behavior (18.3%) were the lowest.
4. Discussion
We examined the behavioral responses of apple snails to changes in water temperature. Distinct behaviors were observed during the transitions in water temperature and these behaviors were clearly classified according to the temperature change treatment based on the SOM analysis.
The proportion of CL during the light periods was higher than during the dark periods under constant temperature conditions at 25 °C. Prey commonly uses this behavioral strategy to avoid predators. Lee and Oh [38] showed that active behavior such as crawling was restricted during the day in ampullariids (e.g., Lanistes solidus and Lamiste nyassanus). In addition, Wada and Yoshida [39] showed that P. canaliculata exhibited greater burrowing behavior during the daytime and emerged from the soil only at night, to avoid diurnal predators. Similarly, Heiler et al. [40] observed a decrease in the mobility of P. canaliculata in the daytime in rearing conditions. We observed changes in the frequency of clinging behavior across all the temperature regimes. While we observed a continuous increase in CL under both 25 °C to 15 °C and 25 °C to 20 °C, the CL also unexpectedly increased on the second day in the 25 °C to 30 °C treatment. This result may be explained by our experimental design wherein we temporarily disconnected the aeration pump to clearly observe their behavior. This disconnection may have resulted in water fouling, which could interfere with the normal perfusion in gill activation [41]. On the second day, apple snails clung to the side of the aquarium at the water surface with their siphon outside of the water, even though they did not ventilate their lungs as temperature rose from 25 °C to 30 °C.
Feeding behavior also gradually decreased as the temperature increased from 25 °C to 30 °C. However, we observed a higher use of an alternative feeding method (where apple snails fold a funnel to gather food) in this temperature treatment compared to the 25 °C to 20 °C and 25 °C to 15 °C treatments. Moreover, the decrease in feeding behavior did not result in a decrease in the amount of lettuce consumed. The feeding behavior frequency decreased over time with both increasing and decreasing temperatures. However, the amount of lettuce eaten decreased sharply under decreasing temperature but increased in the increasing temperature treatment. Our results indicate a higher lettuce consumption with increasing temperatures. Feeding is closely related to movement, movement velocity, and metabolic rates as temperature increases. For example, according to Freiburg and Hazelwood [42], Santos et al. [43], and Ramakrishnan [44], the oxygen consumption of Pomacea sp. increases with temperature (15 °C to 25 °C). In addition, Costil and Bailey [19] reported that feeding behavior in P. carneus increased with increasing temperature, indicating a compensatory increase in the feeding rate in stressful conditions [18].
Seuffert et al. [18] reported that the time P. canaliculta spent crawling was not significantly related to temperature differences. However, our study focused on the behavior of P. canaliculta under changing water temperature conditions. This is also contrary to Heiler et al. [40] who focused on their behavior in different acclimation temperatures. In addition, we subdivided the crawling behavior into two categories (crawling on the bottom or side of the aquarium), which is contrary to Seuffert et al. [18]. We found that the frequency as well as the time spent crawling on the side of the aquarium under decreasing temperature conditions (25 °C to 15 °C) were significantly lower than that under other temperature conditions. This could be associated with the reduced activity levels of the apple snail with decreasing temperature. The activity of apple snails sharply decreased in temperatures ranging from 15 °C to 10 °C (in the Curanmala Grande stream) and from 18 °C to 12 °C (in the Cochico Chico stream) under field conditions [19]. Similarly, in a previous study by Stevens et al. [37], no activity was observed in P. paludos in water temperatures below 13 °C. In our study, behaviors such as CL and CS were strongly affected by the temperature in the lowest decreasing temperature treatment (25 °C to 15 °C). In addition, the frequency of aerial respiration related to the increase in CS. Apple snails typically crawl on the side of the aquarium and periodically respire in the air using their siphon in normal conditions [34]. However, the decrease in temperature reduces the aerial respiration as well as CS.
The relative proportion of time spent inactive was extremely low in our study compared to previous studies (e.g., [19,40]). We did not observe a lack of movement on the aquarium bottom with closed operculum or in a heat-coma state during the experiments. This phenomenon may be explained by the different experimental designs between the studies. While previous studies examined behaviors at temperatures to which the snails were already acclimated [13,18,25,40], we observed the behavior of the snails at gradually changing water temperatures. We found differences in the frequency of inactive behavior in our study compared to previous studies [18,40,45] in spite of a similar experimental design. This could be explained by the differences in the definition of inactive behavior. Alternatively, these differences may be due to differences in P. canaliculata strains used by different research groups [18]. Moreover, the duration of acclimation also differed among the studies, ranging from three days [19] to several weeks [13,25,40,45].
In any behavioral study, the edge effect is an issue to consider [46] because the movement behavior is influenced by the shape of the observation cage. To avoid this issue, we used a round aquarium as the observation cage. However, it was not easy to observe the behaviors of specimens in the round cage, particularly with video observations. In addition, behaviors were not changed significantly by the edge effect in a preliminary study. For instance, when apple snails crawled on the side of the aquarium and reached the edge, they continued the crawling behavior. The majority of the papers in the literature on apple snail behavior involved rectangular aquariums [47,48,49]. However, further research is required to justify the edge effects on response behaviors in apple snails to temperature changes.
As an invasive species, apple snails become active, feed more, and disperse more widely with global warming [20,24]. Therefore, we must understand the ecology of apple snails to establish an efficient and effective management policy for these invasive snails. Our research observed the behavioral differences with temperature conditions for two days. Our results provide basic information for us to understand their activity as well as their behavioral transition with temperature changes. However, our research was limited to only observing their behavior for a short duration (two days), even though the observation interval was more frequent (10 min intervals) than in previous research (e.g., every hour). Further research, including long-term monitoring studies, examining different behaviors periodicity, and daily rhythms among different temperature conditions are necessary to shed greater insight into the behavioral patterns of apple snails.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/w13091149/s1, Figure S1: Behavior patterns of Pomacea canaliculata under different temperature-changing conditions (15 °C, 20 °C, 25 °C (control) and 30 °C).
Author Contributions
Conceptualization, M.-J.B. and Y.-S.P.; methodology, M.-J.B.; software, M.-J.B.; validation, M.-J.B. and Y.-S.P.; formal analysis, M.-J.B.; investigation, M.-J.B.; resources, M.-J.B. and Y.-S.P.; data curation, M.-J.B.; writing—original draft preparation, M.-J.B., E.-J.K. and Y.-S.P.; writing—review and editing, M.-J.B., E.-J.K. and Y.-S.P.; visualization, M.-J.B.; supervision, Y.-S.P.; project administration, Y.-S.P.; funding acquisition, Y.-S.P. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by a grant (NNIBR202002102) from the Nakdonggang National Institute of Biological Resources (NNIBR) funded by the Ministry of Environment (MOE), Republic of Korea and by the National Research Foundation of Korea (NRF) funded by the Korean government (NRF-2019R1A2C1087099).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in this article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Martín, P.R.; Estebenet, A.L.; Cazzaniga, N.J. Factors affecting the distribution of Pomacea canaliculatac (Gastropoda: Ampullariidae) along its southernmost limit. Malacologia 2001, 43, 13–23. [Google Scholar]
- Cowie, R.H. Apple snails (Ampullariidae) as agricultural pests: Their biology, impacts and management. In Molluscs as Crop Pests; Barker, G.M., Ed.; CAB International: Wallingford, UK, 2002; pp. 145–192. [Google Scholar]
- Rawlings, T.A.; Hayes, K.A.; Cowie, R.H.; Collins, T.M. The identity, distribution, and impacts of non-native apple snails in the continental United States. BMC Evol. Biol. 2007, 7, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halwart, M. Fish as Biocontrol Agents in Rice: The Potential of Common Carp Cyprinus carpio (L.) and Nile Tilapia Oreochromis niloticus (L.); Margraf Verlag: Weikersheim, Germany, 1995; p. 169. [Google Scholar]
- Wada, T. Strategies for controlling the apple snails Pomacea canaliculata (Lamarck) (Gastropoda: Ampullariidae) in Japanese direct-sown paddy fields. JARQ 2004, 38, 75–80. [Google Scholar] [CrossRef] [Green Version]
- Cowie, R.H.; Hayes, K.A.; Thiengo, S.C. What are apple snails? Confused taxonomy and some preliminary resolution. In Global Advances in Ecology and Management of Golden Apple Snails; Joshi, R.C., Sebastian, L.S., Eds.; Philippine Rice Research Institute: Nueva Ecija, Philippines, 2006; pp. 3–24. [Google Scholar]
- Hayes, K.A.; Joshi, R.C.; Thiengo, S.C.; Cowie, R.H. Out of South America: Multiple origins of non-native apple snails in Asia. Divers. Distrib. 2008, 14, 701–712. [Google Scholar] [CrossRef]
- Carlsson, N.O.L.; Brönmark, C.; Hansson, L.A. Invading herbivory: The golden apple snail alters ecosystem functioning in Asian wetlands. Ecology 2004, 85, 1575–1580. [Google Scholar] [CrossRef] [Green Version]
- Lowe, S.; Browne, M.; Boudjelas, S.; De Poorter, M. 100 of the World’s Worst Invasive Alien Species—A selection from the Global Invasive Species Database; The Invasive Species Specialist Group (ISSG) a Specialist Group of the Species Survival Commission (SSC) of the World Conservation Union (IUCN): Auckland, New Zealand, 2000; 12p. [Google Scholar]
- Mendoza, R.; Aguilera, C.; Montemayor, J.; Rodriguez, G. Utilization of artificial diets and effect of protein/energy relationship on growth performance of the apple snails Pomacea bridgesi (Prosobranchia: Ampullariidae). Veliger 1999, 42, 101–111. [Google Scholar]
- Estebenet, A.L.; Martín, P.R. Pomacea canaliculata (Gastropoda: Ampullariidae): Life-history traits and their plasticity. Biocell 2002, 26, 83–89. [Google Scholar]
- Ramnarine, I.W. Quantitative protein requirements of the edible snail Pomacea urceus (Müller). J. World Aquacult. Soc. 2004, 35, 253–256. [Google Scholar] [CrossRef]
- Bae, M.J.; Chon, T.S.; Park, Y.-S. Modeling behavior control of golden apple snails at different temperatures. Ecol. Model. 2015, 306, 86–94. [Google Scholar] [CrossRef]
- Teo, S.S. Biology of the golden apple snail, Pomacea canaliculata (Lamarck, 1822), with emphasis on responses to certain environmental conditions in Sabah, Malaysia. Molluscan Res. 2004, 24, 139–148. [Google Scholar] [CrossRef]
- Estebenet, A.L.; Cazzaniga, N.J. Growth and demography of Pomacea canaliculata (Gastropoda: Ampullariidae) under laboratory conditions. Malacol. Rev. 1992, 25, 1–12. [Google Scholar]
- Lach, L.; Britton, D.K.; Rundell, R.J.; Cowie, R.H. Food preference and reproductive plasticity in an invasive freshwater snail. Biol. Invasion 2000, 2, 279–288. [Google Scholar] [CrossRef]
- Albrecht, E.A.; Carreño, N.B.; Castro-Vazquez, A. A study of environmental factors influencing the seasonal onset of reproductive behaviour in the South American apple-snail Pomacea canaliculata (Gastropoda: Ampullariidae). J. Molluscan Stud. 1999, 65, 241–250. [Google Scholar] [CrossRef]
- Seuffert, M.E.; Burela, S.; Martín, P.R. Influence of water temperature on the activity of the freshwater snail Pomacea canaliculata (Caenogastropoda: Ampullariidae) at its southernmost limit (Southern Pampas, Argentina). J. Therm. Biol. 2010, 35, 77–84. [Google Scholar] [CrossRef]
- Costil, K.; Bailey, S.E.R. Influence of water temperature on the activity of Planorbarius corneus (Pulmonata, Planorbidae). Malacologia 1998, 39, 141–150. [Google Scholar]
- Seuffert, M.E.; Martín, P.R. Thermal limits for the establishment and growth of populations of the invasive apple snail Pomacea canaliculata. Biol. Invasions 2017, 19, 1169–1180. [Google Scholar] [CrossRef]
- Oya, S.; Hirai, Y.; Miyahara, Y. Injuring habits of the apple snail, Ampullarius insularus D’Orbigny, to the young rice seedlings. Proc. Assoc. Kyushu Pl. Prot. Res. 1986, 32, 92–95. [Google Scholar] [CrossRef]
- Lv, S.; Zhou, X.N.; Zhang, Y.; Liu, H.X.; Zhu, D.; Yin, W.G.; Steinmann, P.; Wang, X.H.; Jia, T.W. The effect of temperature on the development of Angiostrongylus cantonensis (Chen 1935) in Pomacea canaliculata (Lamarck 1822). Parasitol. Res. 2006, 99, 583–587. [Google Scholar] [CrossRef]
- Bae, M.J.; Kwon, Y.S.; Park, Y.S. Effects of global warming on the distribution of overwintering Pomacea canaliculata (Gastropoda: Ampullariidae) in Korea. Korean J. Ecol. Environ. 2002, 45, 453–458. [Google Scholar]
- Lei, J.; Chen, L.; Li, H. Using ensemble forecasting to examine how climate change promotes worldwide invasion of the golden apple snail (Pomacea canaliculata). Environ. Monit. Assess. 2017, 189, 1–11. [Google Scholar] [CrossRef]
- Bae, M.J.; Park, Y.S. Characterizing the effects of temperature on behavioral periodicity in golden apple snails (Pomacea canaliculata). Ecol. Inform. 2015, 29, 130–138. [Google Scholar] [CrossRef]
- Seuffert, M.E.; Martín, P.R. Juvenile growth and survival of the apple snail Pomacea canaliculata (Caenogastropoda: Ampullariidae) reared at different constant temperatures. Springer Plus 2013, 2, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Takeda, N. Histological studies on the maturation of the reproductive system in the apple snail, Pomacea canaliculata. J. Anal. Biosci. 1999, 22, 425–432. [Google Scholar]
- Estebenet, A.L.; Martín, P.R.; Burela, S. Conchological variation in Pomacea canaliculata and other South American Ampullariidae (Caenogastropoda, Architaenioglossa). Biocell 2006, 30, 329–335. [Google Scholar] [PubMed]
- Mathworks. Matlab. Available online: www.mathworks.com (accessed on 20 January 2014).
- Zar, J.H. Biostatistical Analysis; Prentice Hall: Hoboken, NJ, USA, 1999. [Google Scholar]
- Kohonen, T. Self-organized formation of topologically correct feature maps. Biol. Cybern. 1982, 43, 59–69. [Google Scholar] [CrossRef]
- Kohonen, T. Self-Organizing Maps, 3rd ed.; Springer: Berlin, Germany, 2001; 501p. [Google Scholar]
- Bae, M.J.; Kim, J.S.; Park, Y.S. Evaluation of changes in effluent quality from industrial complexes on the Korean nationwide scale using a self-organizing map. Int. J. Environ. Res. Public Health 2012, 9, 1182–1200. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.S.; Chung, Y.J.; Moon, Y.S. Hazard ratings of pine forests to a pine wilt disease at two spatial scales (individual trees and stands) using self-organizing map and random forest. Ecol. Inform. 2013, 13, 40–46. [Google Scholar] [CrossRef]
- Ward, J.H. Hierarchical grouping to optimize an objective function. J. Am. Stat. Assoc. 1963, 58, 236–244. [Google Scholar] [CrossRef]
- McCune, B.; Mefford, M.J. PC-ORD: Multivariate Analysis of Ecological Data; Version 4 for Windows; [User’s Guide]. MjM Software Design; Gleneden Beach: Lincoln, OR, USA, 1999. [Google Scholar]
- StatSoft, M. Statistica for Windows; StatSoft: Tulsa, OK, USA, 2002. [Google Scholar]
- Lee, T.G.; Oh, K.C. Golden apple snails in Korea. In Global Advances in Ecology and Management of Golden Apple Snails; Joshi, R.C., Sebastian, L.S., Eds.; Philippine Rice Research Institute: Science City of Muñoz, Philippines, 2006; p. 588. [Google Scholar]
- Wada, T.; Yoshida, K. Burrowing by the apple snail, Pomacea canaliculata (Lamarck); daily periodicity and factors affecting burrowing. Kyushu Plant Prot. Res. 2000, 46, 88–93. [Google Scholar] [CrossRef]
- Heiler, K.C.M.; von Oheimb, P.V.; Ekschmitt, K.; Albrecht, C. Studies on the temperature dependence of activity and on the diurnal activity rhythm of the invasive Pomacea canaliculata (Gastropoda: Ampullariidae). Mollusca 2008, 26, 73–81. [Google Scholar]
- Seuffert, M.E.; Martín, P.R. Dependence on aerial respiration and its influence on microdistribution in the invasive freshwater snail Pomacea canaliculata (Caenogastropoda, Ampullariidae). Biol. Invasion 2010, 12, 1695–1708. [Google Scholar] [CrossRef]
- Freiburg, M.W.; Hazelwood, D.H. Oxygen consumption of two amphibious snails: Pomacea paludosa and Marisa cornuarietis (Prosobranchia: Ampullariidae). Malacologia 1977, 16, 541–548. [Google Scholar]
- Santos, C.A.Z.; Penteado, C.H.S.; Mendes, E.G. The respiratory responses of an amphibious snail Pomacea lineata (Spix, 1827), to temperature and oxygen tension variations. Comp. Biochem. Physiol. 1987, 86, 409–415. [Google Scholar] [CrossRef]
- Ramakrishnan, V. Salinity, pH, Temperature, Desiccation and Hypoxia Tolerance in the Invasive Freshwater Apple Snail Pomacea insularum. Ph.D. Thesis, University of Texas, Arlington, TX, USA, 2007. [Google Scholar]
- Stevens, A.J.; Welch, Z.C.; Darby, P.C.; Percival, H.F. Temperature effects on Florida applesnail activity: Implications for snail kite foraging success and distribution. Wildl. Soc. 2002, B35, 75–81. [Google Scholar]
- Potts, J.R.; Hillen, T.; Lewis, M.A. The “edge effect” phenomenon: Deriving population abundance patterns from individual animal movement decisions. Theor. Ecol. 2016, 9, 233–247. [Google Scholar] [CrossRef] [Green Version]
- Prabhakaran, G.; Bhore, S.J.; Ravichandran, M. Development of a bait carrier material for apple snail (Pomacea maculata) based on its feed preferences using snail attractant tracking device. J. Nat. Sci. Biol. Med. 2019, 10, 8–15. [Google Scholar] [CrossRef]
- Maldonado, M.A.; Martín, P.R. Dealing with a hyper-successful neighbor: Effects of the invasive apple snail Pomacea canaliculata on exotic and native snails in South America. Curr. Zool. 2018, 65, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Joo, S.; Jung, S.; Lee, S.; Cowie, R.H.; Takagi, D. Freshwater snail feeding: Lubrication-based particle collection on the water surface. J. R. Soc. Interface 2020, 17, 20200139. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Examples representing the feeding area extracted using digital image processing in Matlab. White color: fed area, and black color: unfed area.
Figure 1.
Examples representing the feeding area extracted using digital image processing in Matlab. White color: fed area, and black color: unfed area.

Figure 2.
Average proportion of time spent by apple snails on each of the nine types of behaviors under different water temperature change regimes for two days. Different letters indicate significant differences among temperature conditions based on Dunn’s multiple comparison test (p < 0.05). Error bars indicate standard error of each behavior. We used 432 numbers using 10 min internal data for each individual. Error bars present the standard errors. FO: floating, CL: clinging to the side of the aquarium, CS: crawling on the side of the aquarium, CB: crawling on the bottom of the aquarium, FE: feeding, FF: folding a funnel with its food, VE: ventilating the lung, FA: falling, and SW: swimming.
Figure 2.
Average proportion of time spent by apple snails on each of the nine types of behaviors under different water temperature change regimes for two days. Different letters indicate significant differences among temperature conditions based on Dunn’s multiple comparison test (p < 0.05). Error bars indicate standard error of each behavior. We used 432 numbers using 10 min internal data for each individual. Error bars present the standard errors. FO: floating, CL: clinging to the side of the aquarium, CS: crawling on the side of the aquarium, CB: crawling on the bottom of the aquarium, FE: feeding, FF: folding a funnel with its food, VE: ventilating the lung, FA: falling, and SW: swimming.

Figure 3.
Comparison of the proportions of nine behavioral categories between day and night conditions in different water temperatures for two days. Error bars indicate standard error of each behavior. (L: Light, D: Dark, 1st: First day, 2nd: Second day). * indicates statistical difference between day and night based on Mann–Whitney U test (p < 0.05).
Figure 3.
Comparison of the proportions of nine behavioral categories between day and night conditions in different water temperatures for two days. Error bars indicate standard error of each behavior. (L: Light, D: Dark, 1st: First day, 2nd: Second day). * indicates statistical difference between day and night based on Mann–Whitney U test (p < 0.05).

Figure 4.
Patterning the behavioral differences in Pomacea canaliculata according to changes in water temperatures (from 25 °C to 15 °C, 20 °C, 25 °C, or 30 °C). (a) Classification of apple snails in the self-organizing map (SOM) trained with nine behavioral categories; (b) U-matrix; and (c) dendrogram of a hierarchical cluster analysis of the SOM units using the Ward linkage method based on the Euclidean distance.
Figure 4.
Patterning the behavioral differences in Pomacea canaliculata according to changes in water temperatures (from 25 °C to 15 °C, 20 °C, 25 °C, or 30 °C). (a) Classification of apple snails in the self-organizing map (SOM) trained with nine behavioral categories; (b) U-matrix; and (c) dendrogram of a hierarchical cluster analysis of the SOM units using the Ward linkage method based on the Euclidean distance.

Figure 5.
Differences in behavioral patterns under different experimental conditions on a self-organizing map. Apple snails were evaluated using four different conditions of changing water temperatures from 25 °C to (a) 15 °C; (b) 20 °C; (c) 25 °C (control); and (d) 30 °C.
Figure 5.
Differences in behavioral patterns under different experimental conditions on a self-organizing map. Apple snails were evaluated using four different conditions of changing water temperatures from 25 °C to (a) 15 °C; (b) 20 °C; (c) 25 °C (control); and (d) 30 °C.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Differences in food consumption at different temperature conditions. Different letters (a, b, c) indicate significant differences between treatments based on Dunn’s multiple comparison test (p < 0.05).
Table 1.
Differences in food consumption at different temperature conditions. Different letters (a, b, c) indicate significant differences between treatments based on Dunn’s multiple comparison test (p < 0.05).
| Experimental Temperature (°C) | Consumed Area (cm2) | ||
|---|---|---|---|
| Total | Day 1 | Day 2 | |
| 15 | 18.2 c | 26.1 * | 10.4 * |
| 20 | 22.5 bc | 26.1 * | 18.9 * |
| 25 | 39.9 ab | 42.7 | 37.2 |
| 30 | 60.9 a | 61.7 | 60.0 |
* indicates a significant difference between first- and second-day food consumption based on a Mann–Whitney U test (p < 0.05).
Table 2.
Differences among nine behavioral categories in different clusters trained in the self-organizing map. FO: floating; CL: clinging to the side of the aquarium; CB: crawling on the bottom of the aquarium; CS: crawling on the side of the aquarium; FE: feeding on lettuce; FF: folding a funnel with its food; SW: swimming; VE: ventilating the lung; and FA: falling.
Table 2.
Differences among nine behavioral categories in different clusters trained in the self-organizing map. FO: floating; CL: clinging to the side of the aquarium; CB: crawling on the bottom of the aquarium; CS: crawling on the side of the aquarium; FE: feeding on lettuce; FF: folding a funnel with its food; SW: swimming; VE: ventilating the lung; and FA: falling.
| Behavior Category | Cluster | ||||||
|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | |
| FO | 7.8 a** | 0.9 b | 0.5 b | 0.3 b | 0.4 b | 0.2 b | 0.1 b |
| (0.3) | (0.2) | (0.2) | (0.1) | (0.2) | (0.1) | (0.1) | |
| CL | 41.0 b | 14.7 d | 27.9 c | 28 c | 27.6 c | 24.8 c | 52.4 a |
| (1.1) | (0.7) | (1.2) | (0.7) | (1.7) | (0.7) | (0.8) | |
| CB | 16.0 cd | 35.6 a | 19.0 b | 20.6 b | 18.6 bc | 15.9 c | 14.1 d |
| (0.8) | (0.8) | (1.0) | (0.6) | (1.2) | (0.6) | (0.6) | |
| CS | 12.0 c | 22.1 b | 20.6 b | 13.3 c | 22.5 b | 37.7 a | 14.1 c |
| (0.9) | (0.7) | (1.1) | (0.5) | (1.3) | (0.6) | (0.6) | |
| FE | 22.7 b | 25.4 b | 22.9 b | 37.2 a | 21.0 bc | 20.5 c | 18.3 c |
| (0.8) | (0.9) | (1.0) | (0.7) | (1.3) | (0.6) | (0.7) | |
| FF | 0.1 b | 0.0 b | 0.1 b | 0.0 b | 7.6 a | 0.0 b | 0.1 b |
| (0.1) | (0.0) | (0.1) | (0.0) | (0.2) | (0.0) | (0.1) | |
| SW | 0.1 | 0.0 | 0.2 | 0.3 | 0.6 | 0.2 | 0.6 |
| (0.1) | (0.0) | (0.1) | (0.1) | (0.2) | (0.1) | (0.2) | |
| VE | 0.1 b | 0.0 b | 8.3 a | 0.0 b | 0.8 b | 0.0 b | 0.1 b |
| (0.1) | (0.0) | (0.2) | (0.0) | (0.3) | (0.0) | (0.1) | |
| FA | 0.2 | 1.3 | 0.5 | 0.3 | 0.9 | 0.6 | 0.2 |
| (0.1) | (0.3) | (0.2) | (0.1) | (0.3) | (0.1) | (0.1) | |
* The numbers in parenthesis represent the number of sampling sizes. ** Values are means (standard error). Different letters (a, b, c, etc.) indicate significant differences among clusters based on Dunn’s multiple comparison tests (p < 0.01).
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Bae, M.-J.; Kim, E.-J.; Park, Y.-S. Comparison of Invasive Apple Snail (Pomacea canaliculata) Behaviors in Different Water Temperature Gradients. Water 2021, 13, 1149. https://doi.org/10.3390/w13091149
AMA Style
Bae M-J, Kim E-J, Park Y-S. Comparison of Invasive Apple Snail (Pomacea canaliculata) Behaviors in Different Water Temperature Gradients. Water. 2021; 13(9):1149. https://doi.org/10.3390/w13091149
Chicago/Turabian StyleBae, Mi-Jung, Eui-Jin Kim, and Young-Seuk Park. 2021. "Comparison of Invasive Apple Snail (Pomacea canaliculata) Behaviors in Different Water Temperature Gradients" Water 13, no. 9: 1149. https://doi.org/10.3390/w13091149
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.