
The Species Diversity of Tropical Freshwater Rotifers (Rotifera: Monogononta) in Relation to Environmental Factors


1
DN-EBR, The University of Da Nang—University of Science and Education, Da Nang City 550000, Vietnam
2
Faculty of Biology and Environmental Sciences, The University of Da Nang—University of Science and Education, 459 Ton Duc Thang St., Danang City 550000, Vietnam
*
Authors to whom correspondence should be addressed.
Water 2021, 13(9), 1156; https://doi.org/10.3390/w13091156
Submission received: 15 March 2021
/
Revised: 9 April 2021
/
Accepted: 15 April 2021
/
Published: 22 April 2021
(This article belongs to the Special Issue Zooplankton Diversity in Water Bodies)
Abstract
:This study aims to evaluate the diversity of rotifers in various freshwater habitats in Da Nang City, Vietnam, and to investigate the relationship between community structure and environmental conditions. A total of 75 rotifer species belonging to 25 genera and 17 families were recorded, out of which seven species are new to Vietnam. Species from the families Brachionidae and Lecanidae are abundant (containing about 50% and 69.44%, respectively) in the total rotifer species in both lacustrine and riverine habitats. The number of rotifers recorded in the lakes (64 taxa) was much higher than those in rivers (36 taxa), and this is possibly due to the conditions in lakes being more favorable for the development of small zooplanktons. The significant influences of environmental factors (temperature, pH, turbidity, and trophic state) on the rotifer community were well-reflected in the distribution of commonly found species in lakes (p-value < 0.05). Our study results enrich the data on the diversity and the ecology of rotifers in Vietnam.
1. Introduction
Rotifers represent an important zooplankton group since they can contribute more than 50% of zooplankton productivity [1,2]. The rotifer species can be found in all kinds of water bodies worldwide due to their high adaptability to a wide range of environmental conditions and their ability to occupy open niches quickly. In aquatic systems, rotifers play a crucial role in the energy flows of that system. This group of organisms is responsible for transferring carbon from picoplankton and nanoplankton to macro-zooplankton; they act as links between microbial loops and the larger aquatic food web [3,4,5]. Additionally, rotifers are considered effective indicators for environmental conditions because of their sensitivity to the changes in that particular environment [6,7,8,9]. It was well-documented that the rotifer community structure is influenced by factors such as water temperature, salinity, transparency, trophic status, and predators [10,11]. The rotifer species composition, its total abundance, and its diversity indices have been widely applied in order to assess water quality [12,13,14].
Numerous studies have focused on seasonal or periodic changes in the rotifer community in relation to water characteristics in a single type of water body. However, very few studies compared their structures in different water bodies within the same area. Liang et al. (2019) [15] revealed that the rotifer community structure differs significantly among three types of water bodies: river downstream, reservoir, and tidal creek. These differences could be attributed to the heterogeneity in the trophic state, the salinity, the turbidity, and the hydrological regime of the water body.
Despite several efforts made recently, the data on rotifer species distribution in tropical freshwater, whether in different countries in general or in Vietnam specifically, are still relatively limited. Moreover, less attention has been paid to the relationship between the rotifer community structure and the environmental conditions. The aims of this study were to examine and investigate the following: (1) the diversity of rotifers in freshwater bodies in Da Nang city, Vietnam, and (2) the influences of some environmental factors on the distribution of rotifers in different aquatic systems in this particular area.
2. Materials and Methods
2.1. Study Area
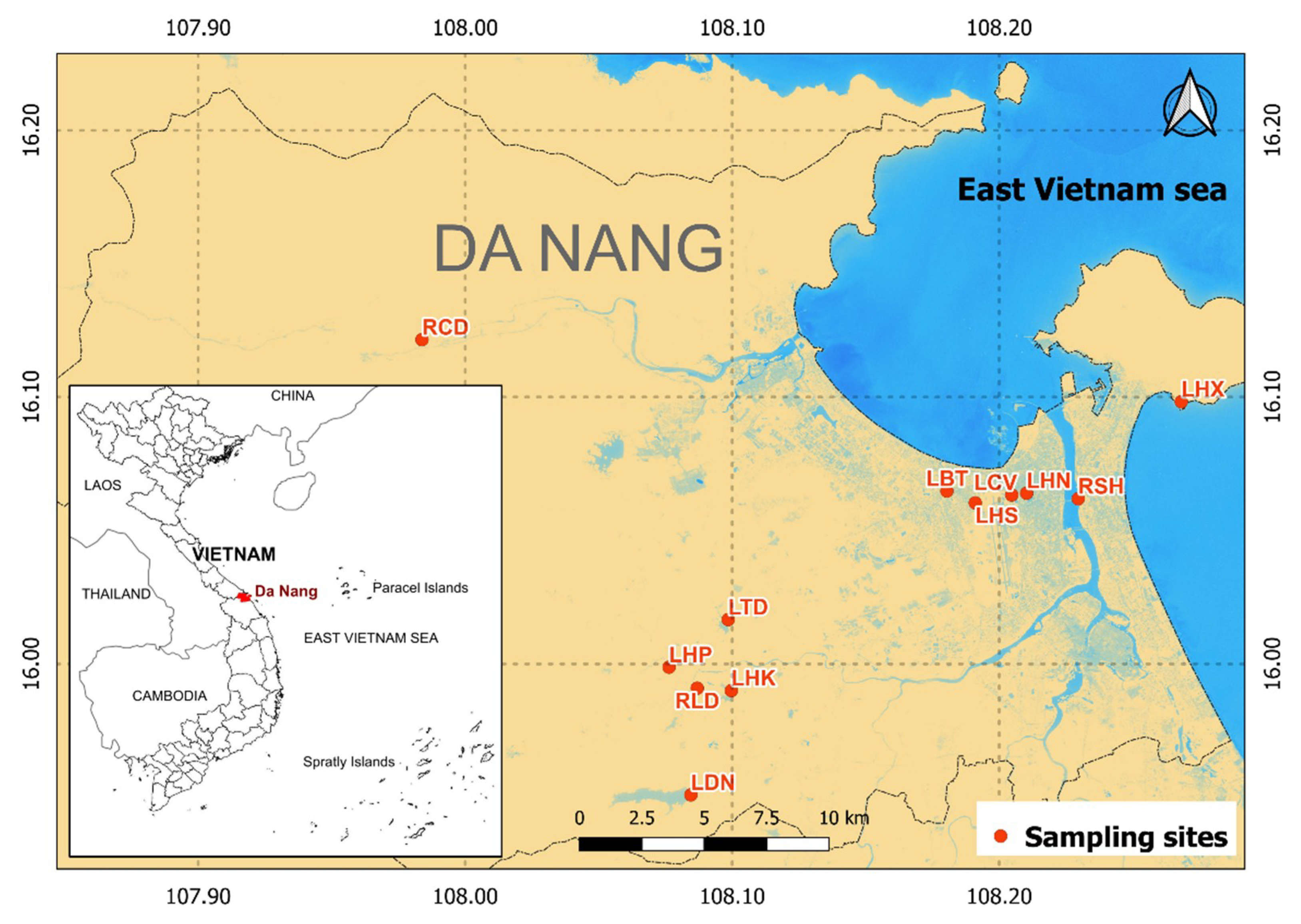
The sampling sites consisted of eight freshwater lakes (8 sites) and two rivers (Han River and Cu De river) (4 sites) in both rural and urban areas of Da Nang city. The sites represent a variety of lotic and lentic freshwater habitats with different natural conditions and anthropogenic impacts (Figure 1). The water quality characteristics of the sampling sites during the sampling campaign were identified by the authors (Table 1). These habitats had a trophic state ranging from oligotrophic to eutrophic, their water temperatures ranged from 24.74 °C to 27.32 °C, and their turbidity ranged from 0.9 to 26.5 NTU. The electrical conductivity values were generally low (≤1), which indicates that all of the sampling sites were freshwater. The pH of water fluctuated from around neutral values to weakly alkaline values, specifically, from 6.61 to 7.78.
2.2. Sampling
The sampling campaign took place during the dry season in August 2018. Qualitative samples of rotifers were collected by dragging a 50 μm mesh size cast-net at 15–20 cm depth and were then preserved with 4% formaldehyde. Rotifer specimens were then sorted and examined under a Hund Wetzlar H600 microscope. The trophi of rotifers were observed by adding a drop of sodium–hypochlorite (NaOCl) to dissolve and isolate the hard trophic parts. Rotifer taxa were identified in the following major work: Koste and Shiel (1987, 1989a, 1989b, 1990a, and 1990b) [16,17,18,19,20], Shiel and Koste (1992) [21], and Segers (1995) [22]. The taxonomic classification and nomenclatures of the rotifers followed the work of Segers (2007) [23].
Quantitative samples of the rotifers were collected by filtering 25 L of water through a 50 μm mesh size cast-net and preserved with 4% formaldehyde. The density of rotifers in the samples were determined by using a counting chamber.
The water quality parameters included temperature, pH, total dissolved solids (TDS), electrical conductivity (EC), and turbidity. These parameters were directly measured in situ using a multiparameter water quality sonde YSI 6920. The nutrients (the total nitrogen and total phosphorus) and chlorophyll-a concentrations in water were identified in the laboratory by standard methods. The TRIX trophic index, which is calculated based on the values of nitrogen, phosphorous, and chlorophyll-a, as shown below, was used to evaluate the trophic status of water bodies [24]:
where
TRIX = [log(Chl-a × aD%O × TN × TP) − (−1.5)]/1.2
Chl-a—chlorophyll-a concentration (μg/L);
aD%O—oxygen as absolute% deviation from saturation;
TN—total nitrogen (μg/L);
TP—total phosphorus (μg/L).
2.3. Data Analysis
The species accumulator and species richness estimators were calculated using the vegan package [25] in the R program [26]. Out of these estimators, Jackknife 2 [27], Chao 2 [28], and Bootstrap [29] richness estimators were selected to estimate the expected diversity of planktonic rotifers.
In order to account for the variation in environmental factors among study sites, Principal Component Analysis (PCA) was used. The relation between the distribution of rotifers and environmental factors was investigated by using canonical correspondence analysis (CCA). Explanatory environmental variables were selected by forward selection in the CCA, while selecting only those variables that were significantly related to taxonomic abundances (p-value ≤ 0.05) according to the Monte Carlo permutation tests. The rotifer taxa that were observed in less than 25% of the samples were excluded from CCA to prevent these rare taxa from having an inordinate influence on the statistical results.
3. Results
3.1. Species Diversity of Planktonic Rotifers
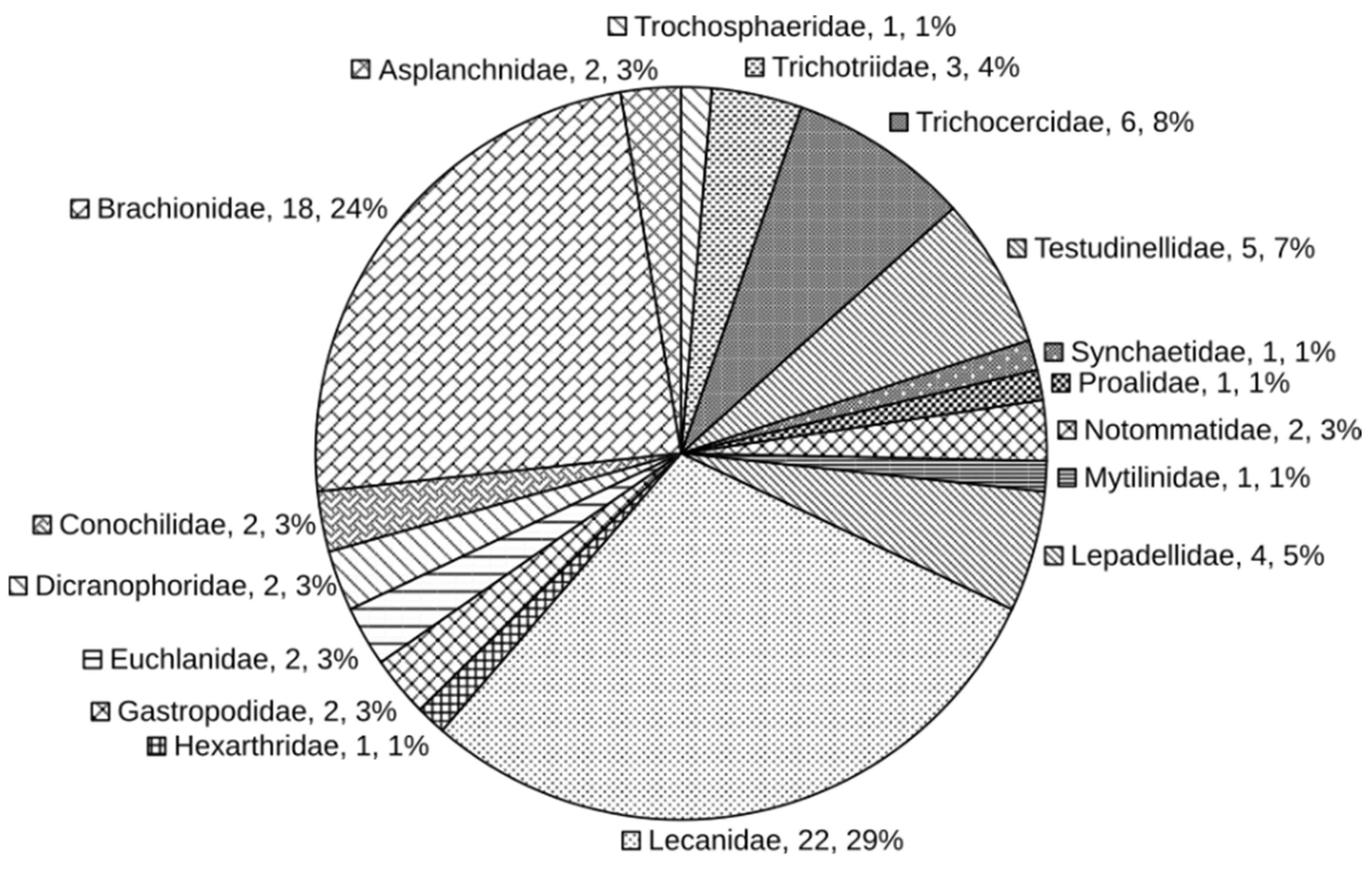
A total of 75 rotifer species, belonging to 25 genera, 17 families, and 2 orders were identified from 12 freshwater bodies in Da Nang (Table 2). Among them, seven species were recognized as new to Vietnam, including Testudinella incisa (Ternetz, 1892), Dicranophorus claviger (Hauer 1965), Keratella americana Carlin 1943, Lecane punctata (Murray, 1913), Lecane yatseni Wei and Xu 2010, Tripleuchlanis plicata (Levander 1894), and Trichocerca stylata (Gosse 1851). The structure of the planktonic rotifer community is shown in Figure 2. Lecanidae (22 taxa, 29%) and Brachionidae (18 taxa, 24%) were the two dominant families in the community. Three families had more than five species in this list: Trichocercidae, Testudinellidae, and Lepadellidae. The rest were composed of 12 families, with only one to three species each.
3.2. Comparison of Rotifer Community Structures among Habitat Types
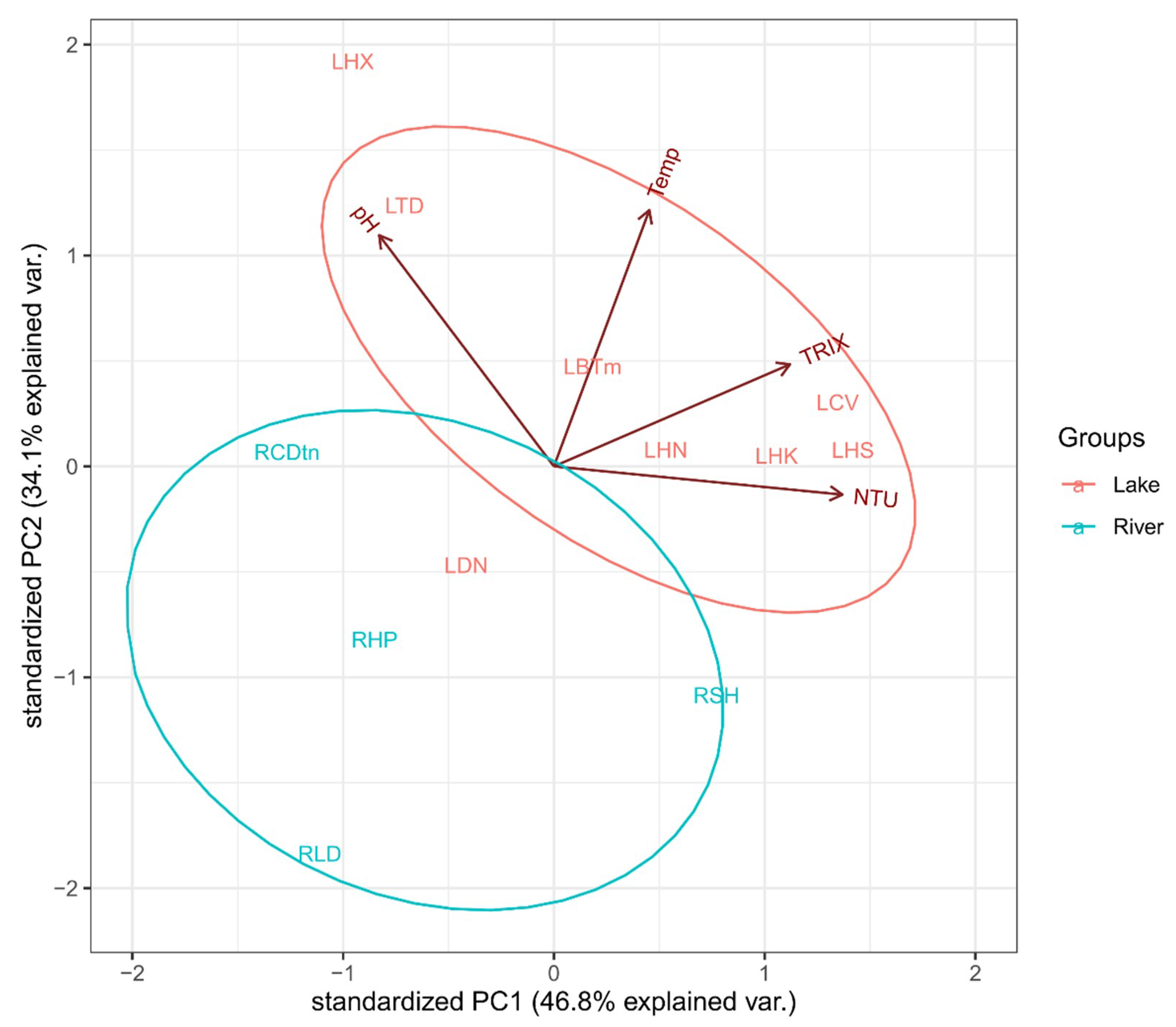
The first and second axes combinedly explained 80.9% of the differences in the physical and chemical characteristics of the water from the investigated systems (Figure 3). TRIX and turbidity (expressed by Nephelometric Turbidity Unit—NTU) were found to be associated with the first principal component (the scores were 0.56 and 0.68, respectively), while the second principal component was determined by water temperature and pH (the scores were 0.71 and 0.64, respectively). The results from the principal component analysis revealed a discrepancy between two types of aquatic ecosystems—the lake ecosystem and the river ecosystem. More specifically, the lakes’ water was typically characterized by higher TRIX, higher temperature, and higher turbidity values compared to those of rivers (Table 1). Only the pH value remained quite similar between the two types of aquatic ecosystems. In this analysis, electrical conductivity was eliminated due to high collinearity with turbidity. Consequently, the rotifer assemblage inhabiting these two types of aquatic habitats were expected to be different in terms of structure and in terms of the level of diversity. Thus, we divided our data into two categories for further investigation: the lake category and the river category. More noticeably, it is also possible to categorize 12 sampling sites into urban and suburban areas based on strong discrimination solely by assessing the first environmental gradient (PC1 = 46.8%), which is closely related to turbidity and TRIX gradients (Figure 3 and Table 1).
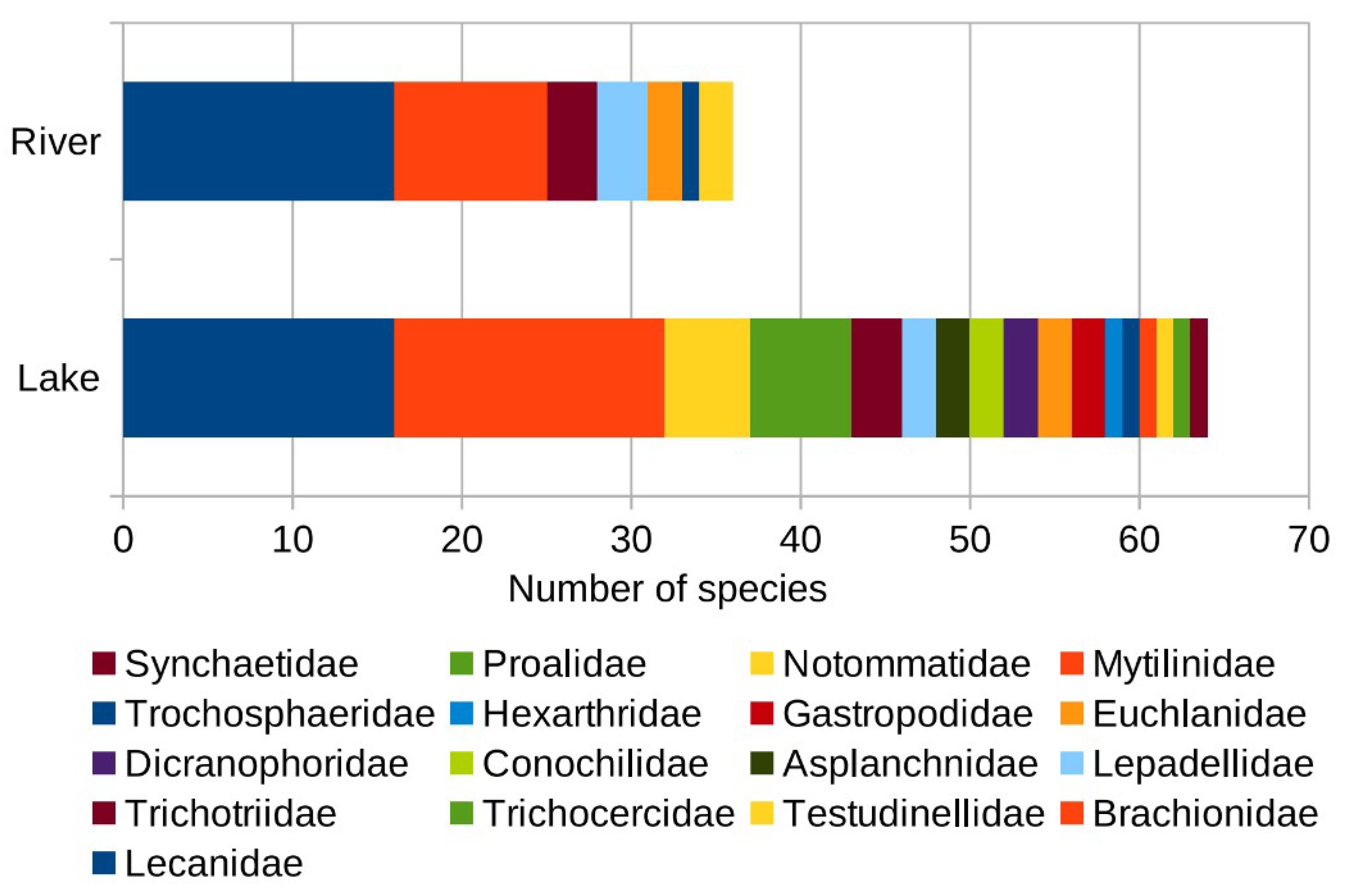
A comparison between the planktonic rotifer communities in lakes and in rivers showed that the lake system was much more diverse in terms of the rotifer community than that of the river system (Figure 4). Seventeen recorded families were found in lakes, while only seven recorded families appeared in rivers. In addition to this, the number of rotifer taxa recorded in lakes was much higher than in rivers (64 taxa vs. 36 taxa). However, these two aquatic ecosystems were similar in terms of the common dominant families, which were Lecanidae and Brachionidae. These two families accounted for 69.44% and 50% of the total number of species in rivers and lakes, respectively.
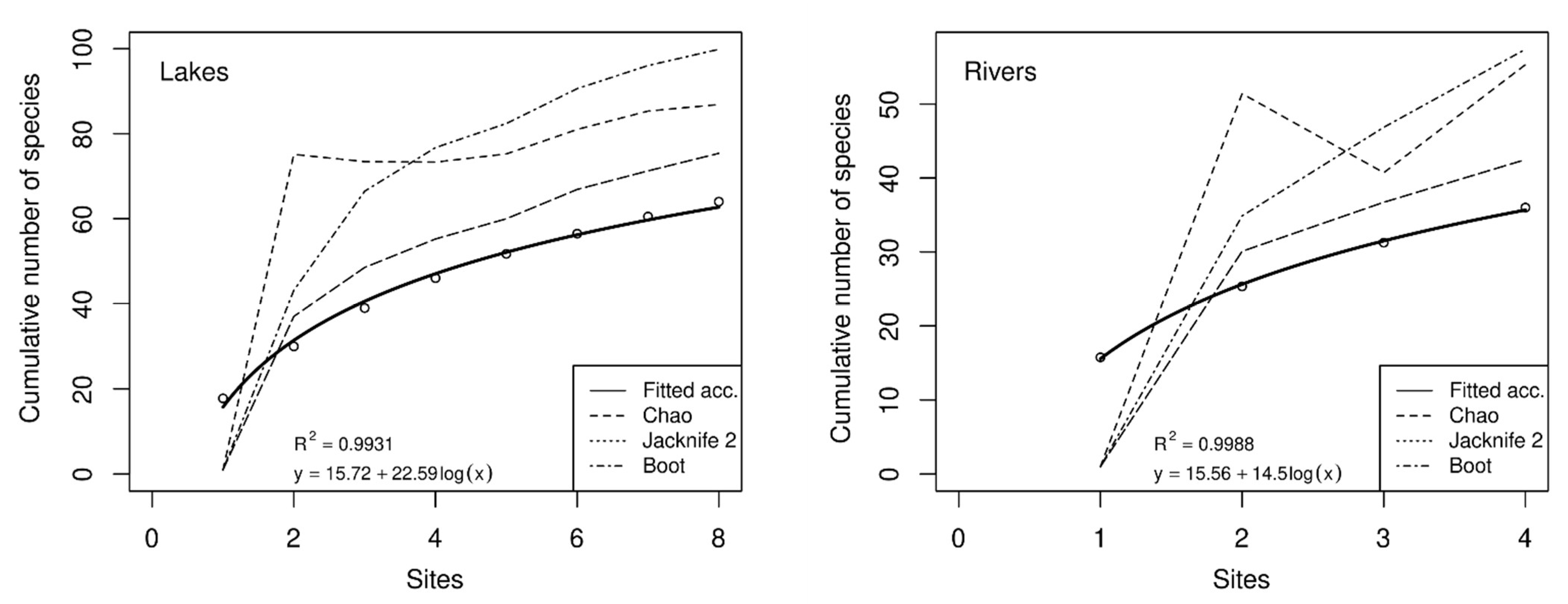
The differences in diversity and species richness between lacustrine and riverine rotifer assemblages were further reflected by the species accumulation curves and species richness estimators, as shown in Figure 5. The species accumulation curves of both groups shared a similar pattern, which increased rapidly with a slope of 22.59 for the lake group and of 14.5 for the river group. This demonstrated that lentic habitats should host a more diverse community of planktonic rotifers. Nonetheless, the average number of taxa found per sample was comparable (15.72 and 15.56). The estimation of rotifer species richness using three different statistical techniques (Jackknife 2, Chao 2, and Bootstrap) in lakes ranged from 75 to 100 taxa and from 42 to 57 taxa in rivers. Notwithstanding, all other richness estimators indicated that the numbers of species could be higher than those currently recorded in both types of aquatic systems.
We further noticed a distinction in the physical and chemical characteristics of water samples collected from urban and rural freshwater bodies, which is possibly attributed to their basin (surrounding areas). The lakes and rivers in rural areas are surrounded by layers of vegetation, while in urban areas, they are normally isolated in residential areas. The lack of natural water filtration and heat insulation systems culminated with a high rate of nutrient-rich discharge from its surroundings makes urban surface water more eutrophic, more turbid, and higher in temperature. Canonical correspondence analysis was implemented to investigate the relationships between environmental variations and the distribution of rotifer taxa in lakes and in rivers from urban and rural areas. There were no significant correlations between the environmental condition and the biological data found for river samples, although the model of planktonic rotifers and environmental factors in lakes was statistically significant at a confidence level of 5% (p-value = 0.024) (Figure 6).
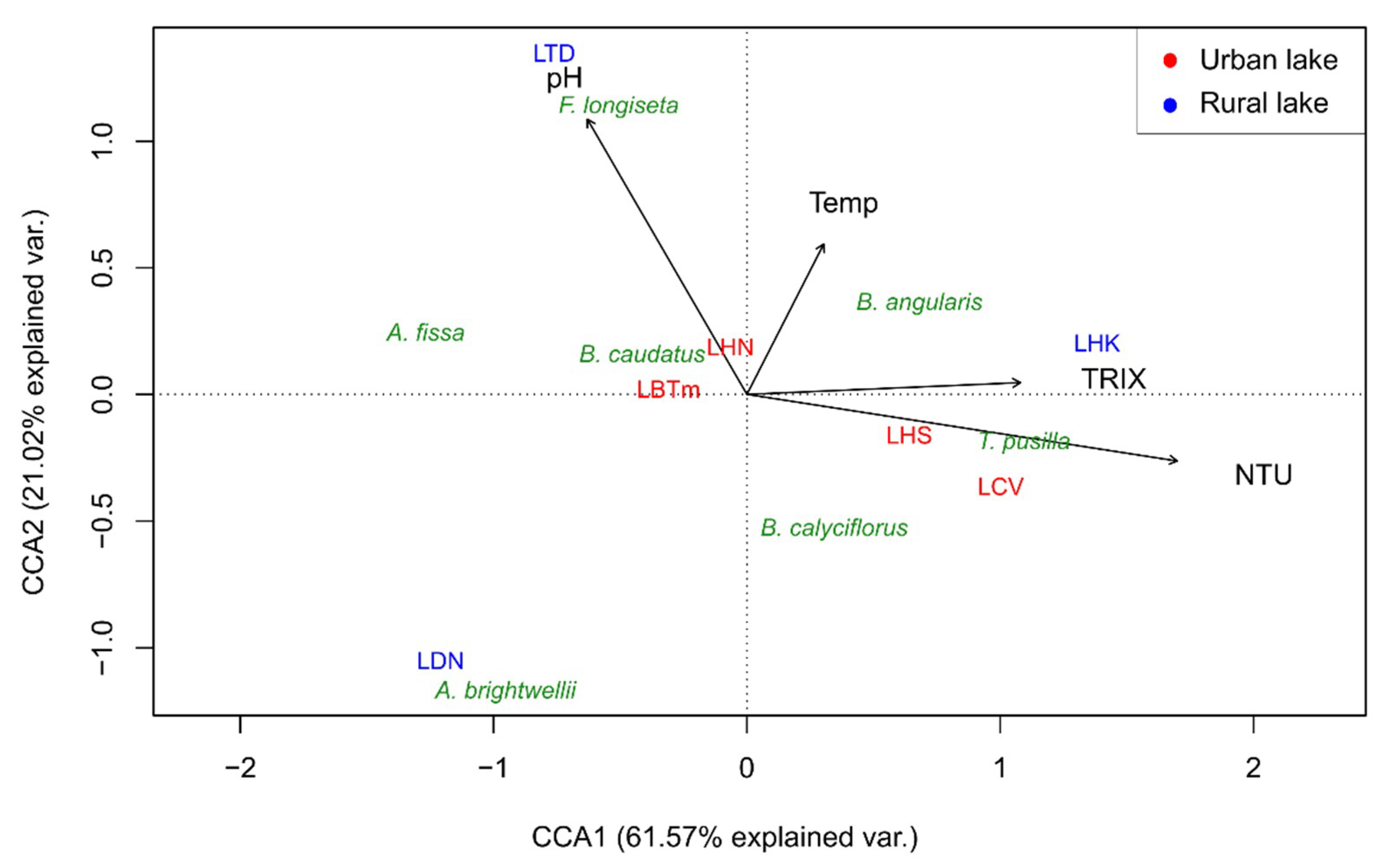
In general, 84.50% of the variation in the distribution of common rotifer species in Da Nang lakes was attributed to the combined influences of four environmental factors, including pH levels, temperature, turbidity, and TRIX. Among these variables, TRIX and temperature were statistically significant at a confidence level of 95%, with p-values of 0.042 and 0.022, respectively, while pH was only significant at a confidence level of 90% (p-value = 0.069). The first two axes in total were responsible for 82.53% of the variation in the biological data, yet only CCA1 was considered significant (p-value = 0.02). This gradient was mainly defined by TRIX, with a score of 0.6. The turbidity also established strong correlations with this axis, with a score of 0.95; however, this was not statistically significant. Meanwhile, CCA2 was considerably correlated with both temperature (score = 0.44) and pH (score = 0.80).
With regard to the distribution of some common rotifer taxa, it seems that Trichocerca pusilla, Brachionus angularis, and Brachionus calyciflorus were favored by high trophic states and high levels of turbidity. On the other hand, Anuraeopsis fissa and Asplanchna brightwellii tended to prefer oligotrophic and clear water conditions. Concurrently, Filinia longiseta was found to be abundant when the second environmental gradient increased, and this correlated well with higher pH and temperature levels.
More interestingly, urban lakes were observed to share comparable environmental conditions: a high trophic state, high turbidity, and a neutral pH. Urban lakes thus, have a similar community structure to the dominant Brachionus genus. In contrast, there were no general patterns in the rural lakes’ condition; therefore, the compositions of rotifer communities were highly variable among rural lakes. For example, Trichocerca pusilla was abundant in Hoi Khe Lake, while Anuraeopsis fissa and Asplanchna brightwellii dominated the rotifer assemblage of Dong Nghe Lake.
4. Discussion
4.1. Community Structure
Recently, more attention has been paid to studies on rotifer community composition and distribution in Vietnamese aquatic and psammon habitats [30,31,32,33,34,35], leading to a recognition of the high diversity of rotifers in Vietnam. This study continues these efforts by providing a taxonomic list of rotifers from a variety of lotic and lentic freshwater habitats in Da Nang and by discovering seven new species in Vietnam. Our records of the community structure for freshwater bodies in Da Nang were quite similar to that of other areas in Vietnam [31,32,33,34,35], in Thailand [36], and in Cambodia [37]: with Brachionidae, Lecanidae, Trichocercidae, and Lepadellidae being the dominant families. Lecanidae and Brachionidae were represented by a large number of tropic-centered genera, specifically Lecane and Brachionus, in both lakes and rivers in Da Nang. The dominance of these two families is commonly observed in freshwater bodies in Southeast Asia [31,32,35,38,39,40]. This similarity is possibly due to the homogeneity of the climate regime within the region.
The number of rotifer species recorded in Da Nang lakes (63 taxa) was lower in comparison with the other lakes in Central Vietnam, such as lake Bau Thiem (89 taxa) [32] and lake Thuy Tien (82 taxa) [31,32]. The number of rotifer species recorded in Da Nang lakes (64 taxa) was higher than that in Southern Vietnamese lakes and reservoirs, such as Tra Su (9 taxa), Bung Binh Thien (24 taxa), Tri An (33 taxa), Suoi Trau (45 taxa), and Da Ban (16 taxa) [34,35]. With respect to riverine habitats, there were 36 rotifer taxa identified from two rivers in Da Nang that were lower than that in Nhu Y river in Hue (49 taxa) [31] but greater than that in some rivers in Southern Vietnam (14–29 taxa) [34]. However, these comparisons were mainly for reference since the sampling intensity and strategy applied in each study were not the same. Moreover, the actual diversity of rotifers in Da Nang freshwater bodies might be higher than our observation since some small species (body size < 50 µm) might be lost due to the choice of sampling mesh size. Nevertheless, it should be noted that several small species, e.g., Anuraeopsis fissa, were still found in this study. Taking environmental conditions into our considerations, our study demonstrates a clear discrepancy between lakes and rivers in Figure 3. Lentic and lotic environments are proven to be different from one another in terms of hydrology and chemistry, resulting in differences in ecology, specifically trophic level [15,41,42]. It is expected that most lentic habitats possess higher diversity in terms of plants, algae, and fish species given the wide range of specific niches. Our results agree with this expectation, as both the abundance and diversity levels of Rotifera communities of lacustrine systems were much greater than that of the riverine systems.
4.2. Relation to Environmental Factors
From the principal component analysis, it is possible to relate this finding to higher values of TRIX and turbidity. The trophic state is considered one of the most important characteristics of an aquatic ecosystem [43] and commonly plays a key role in determining the composition of rotifer communities [13]. Rotifers are normally found to be increasingly dominant in more productive systems [44,45]. Nonetheless, it is noteworthy to point out that the diversity index for the whole zooplankton assemblage was evidenced to decrease with an increasing degree of eutrophication [45,46,47]. In our study, however, the highest diversity of rotifers (23 taxa) was observed in oligotrophic water bodies and then followed by mesotrophic water bodies (14 taxa). Eutrophic water bodies had the lowest diversity (12 taxa).
Aside from the trophic condition, turbidity has been shown to have various influences on aquatic biota such as the ecological condition, resource availability, and species interactions [48]. For example, suspended solids may provide refuge for zooplankton from visual predators, yet high mineral turbidity may reduce phytoplankton production through light limitation, which in turn leads to a reduction in food availability for zooplankton. Some investigated lakes in our study showed a higher turbidity level compared to rivers, which diverged from the common pattern. This may be due to the fact that our sampling campaign was taken during the low water period (August 2018—the dry season in Da Nang). Thus, the influences of turbulent inflows into the rivers were very minor; on the other hand climate conditions were favorable for the development of phytoplankton and led to an elevation in turbidity in lentic systems. Many studies indicated a deleterious effect of the increased concentration of suspended solids on Cladocera populations while rotifer populations were unaffected [49,50,51], resulting in an expansion of rotifer assemblage in turbid water. Our results were in line with these findings: lake habitats with higher turbidity (11.3 NTU, 63 taxa) might create the ideal conditions for rotifers to develop than in riverine systems with lower turbidity (6.6 NTU, 36 taxa).
In summary, we suggest that the condition of lacustrine systems in comparison with riverine systems in Da Nang during the studied period was more favorable for the development of Rotifera. The higher species richness and the abundance of rotifers in lakes might be the result of increased food availability and reduced competition for living resources with other planktonic animals.
The environmental data retrieved from the lake samples were abundant and covered a wide range of conditions. Thus, a canonical correspondence analysis was applied to identify the influences of environmental variables on patterns of variation in the rotifer assemblage, which is represented by some commonly found species. A statistically significant model was obtained, which indicated that the densities of the concerned species were indeed regulated by TRIX and pH.
It is well-known that the community structure and the abundance of rotifers are considerably affected by total abundance (hence rotifer species composition) and that diversity indices have been widely used as bioindicators for assessing water quality [7,12,13,14,52]. May and O’Hare (2005) argued that rotifer abundance reflects lake trophicity better than species composition because it is more sensitive to changes in the trophic state. Good indicators of eutrophic condition have been listed in several studies, and are Brachionus sp., Trichocerca pusilla, and Filinia longiseta [7,20,52,53], as they are normally found with high densities in nutrient-rich water. Consistent with these findings, a high number of T. pusilla, B. angularis, and B. calyciflorus individuals were found in eutrophic urban lakes in our study. Grazing strategy, including food selection and collection, should be taken into account when explaining the variations in rotifer dominance [15]. For example, B. angularis, which is classified as fine particle sedimentations based on their malleate trophi and preference for food items of less than 5 μm in size [54,55], is favored by environments with a high concentration of dissolved organic matter—which is enhanced by eutrophication. Anuraeopsis fissa is considered a microfilter-feeder, mostly feeding on bacteria-laden detritus particles and nanophytoplankton that are in suspension [56]; thus, they tend to be numerous in oligotrophic water. This is confirmed by our results as we recorded a relatively high number of A. fissa individuals in two oligotrophic lakes—Dong Nghe (7040 individuals·m−3) and Truoc Dong (1920 individuals·m−3).
The density of Filinia longiseta seemed to be boosted by the alkaline conditions in Lake Truoc Dong. Berziņš and Pejler (1989) [57] reported a bimodal pH tolerance in this species, with its peak abundance occurring at pH levels from 7–9. Most noticeably, the abundance of Asplanchna brightwellii seemed to be negatively correlated with those of the Brachionus species. Specifically, in Lake Dong Nghe, the density of Asplanchna brightwellii was very high, with 9240 individuals/m3, while the number of total Brachionus individuals was much fewer, with only 2640 individuals/m3, and half of them were B. calyciflorus. In fact, Asplanchna brightwellii is a predator feeding on small herbivorous rotifers (e.g., Brachionus) [58,59], yet it tends to avoid prey that are able to develop defenses such as spines (Brachionus calyciflorus) and setae (Filinia longiseta) [58]. This relationship can evidently be seen in lake Dong Nghe since this habitat provides a zooplanktonic community with a low level of food (Chl-a = 3.55 mg·m−3), which allows for the dominance of small rotifers.
5. Conclusions
A total of 75 species of rotifers, belonging to 25 genera and 17 families, were recorded in eight lakes and two rivers in Da Nang, and seven of these species were new to Vietnam. The community structures in both rivers and lakes were mainly contributed by Lecanidae and Brachionidae, with 69% of the total species in rivers and 50% in lakes belonging to these two families. The differences in the natural conditions of the lacustrine and riverine systems might be drivers of dissimilarity between the rotifer communities in lakes and rivers. The higher species richness and diversity in lakes in comparison to those in rivers could be attributed to the higher values of trophic level, temperature, and the turbidity of lake water. Furthermore, the distribution of rotifers in lakes was found to be significantly related to abiotic factors (temperature, TRIX, pH, and turbidity) at a confidence level of 95%. These findings have contributed important information to the body of knowledge on rotifers in Vietnam as well as in tropical regions.
Author Contributions
Conceptualization, M.T.-D., Q.A.T.-N., N.-T.P., and Q.H.D.; methodology, M.T.-D., Q.H.D., and Q.A.T.-N.; data curation, M.T.-D., N.-T.P., and Q.H.D.; software: M.T.-D. and N.-T.P.; supervision: M.T.-D.; validation: M.T.-D.; writing—original draft: Q.A.T.-N. and N.-T.P.; writing—review and editing: M.T.-D. and Q.A.T.-N. All authors have read and agreed to the published version of the manuscript.
Funding
This research is funded by Funds for Science and Technology Development of the Ministry of Education and Training, Vietnam, grant number B2020-DNA-08.
Acknowledgments
We would like to thank the Faculty of Biology and Environmental Science at the University of Science and Education (UDN) for providing us with the research facilities.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Herzig, A. The analysis of planktonic rotifer populations: A plea for long-term investigations. In Rotifer Symposium IV; Springer: Dordrecht, The Netherlands, 1987; pp. 163–180. [Google Scholar] [CrossRef]
- Walz, N. Rotifer populations in plankton communities: Energetics and life history strategies. Experientia 1995, 51, 437–453. [Google Scholar] [CrossRef]
- Wallace, R.L.; Snell, T.W.; Ricci, C.N.; Nogrady, T. Rotifera Biology, ecology and systematics. In Guides to the Identification of the Microinvertebrates of the Continental Waters of the World 23; Segers, H., Dumont, H.J., Eds.; Kenobi Productions, Ghent & Backhuys Publishers: Leiden, The Netherlands, 2006; p. 165. ISBN 9789057821783. [Google Scholar]
- Arndt, H. Rotifers as predators on components of the microbial web (bacteria, heterotrophic flagellates, ciliates)—A review. In Rotifer Symposium VI; Springer: Dordrecht, The Netherlands, 1993; pp. 231–246. [Google Scholar] [CrossRef]
- Devetter, M.; Seda, J. Rotifer fecundity in relation to components of microbial food web in a eutrophic reservoir. Hydrobiologia 2003, 504, 167–175. [Google Scholar] [CrossRef]
- Duggan, I.C.; Green, J.D.; Shiel, R.J. Distribution of rotifer assemblages in North Island, New Zealand, lakes: Relationships to environmental and historical factors. Freshw. Biol. 2002, 47, 195–206. [Google Scholar] [CrossRef]
- Sládeček, V. Rotifers as indicators of water quality. Hydrobiologia 1983, 100, 169–201. [Google Scholar] [CrossRef]
- Vilaclara, G.; Sládeček, V. Mexican rotifers as indicators of water quality with description of Collotheca riverai, n. sp. Arch. Hydrobiol. 1989, 115, 257–263. [Google Scholar]
- May, L.; Spears, B.M.; Dudley, B.J.; Gunn, I.D.M. The response of the rotifer community in Loch Leven, UK, to changes associated with a 60% reduction in phosphorus inputs from the catchment. Int. Rev. Hydrobiol. 2014, 99, 65–71. [Google Scholar] [CrossRef] [Green Version]
- Devetter, M. Influence of environmental factors on the rotifer assemblage in an artificial lake. Hydrobiologia 1998, 387, 171–178. [Google Scholar] [CrossRef]
- Khaleqsefat, E.; Malekzadeh-Viayeh, R. Effect of season and environmental variables on rotifer community structure: Evidence from two selected freshwaters in Northwest Iran. Xenotransplantation 2013, 17, 61–70. [Google Scholar]
- May, L.; O’Hare, M. Changes in rotifer species composition and abundance along a trophic gradient in Loch Lomond, Scotland, UK. Hydrobiologia 2005, 546, 397–404. [Google Scholar] [CrossRef]
- Wen, X.-L.; Xi, Y.-L.; Qian, F.-P.; Zhang, G.; Xiang, X.-L. Comparative analysis of rotifer community structure in five subtropical shallow lakes in East China: Role of physical and chemical conditions. Hydrobiologia 2010, 661, 303–316. [Google Scholar] [CrossRef]
- Gutkowska, A.; Paturej, E.; Kowalska, E. Rotifer trophic state indices as ecosystem indicators in brackish coastal waters. Oceanologia 2013, 55, 887–899. [Google Scholar] [CrossRef]
- Liang, D.; Wei, N.; Wang, Q.; Jersabek, C.D.; He, X.; Yang, Y. Influence of Hydrological Heterogeneity on Rotifer Community Structure in Three Different Water Bodies in Shantou Area, Guangdong (China). Zool. Stud. 2019, 58, e23. [Google Scholar] [CrossRef]
- Koste, W.; Shiel, R.J. Rotifera from Australian inland waters. III. Euchlanidae, Mytilinidae and Trichotriidae (Rotifera: Monogononta). Trans. R. Soc. Aust. 1989, 113, 85–114. [Google Scholar]
- Koste, W.; Shiel, R.J. Rotifera from Australian inland waters. IV. Colurellidae (Rotifera: Monogononta). Trans. R. Soc. South. Aust. 1989, 113, 119–143. [Google Scholar]
- Koste, W.; Shiel, R.J. Rotifera from Australian inland waters. II. Epiphanidae and Brachionidae (Rotifera: Monogononta). Invertebr. Syst. 1987, 1, 949–1021. [Google Scholar] [CrossRef]
- Koste, W.; Shiel, R. Rotifera from Australian inland waters V. Lecanidae (Rotifera: Monogononta). Trans. R. Soc. South. Aust. 1990, 114, 1–36. [Google Scholar]
- Koste, W.; Shiel, R.J. Rotifera from Australian inland waters. VI. Proalidae, Lindiidae (Rotifera: Monogononta). Trans. R. Soc. South. Aust. 1990, 114, 129–143. [Google Scholar]
- Shiel, R.J.; Koste, W. Rotifera from Australian inland waters. VIII. Trichocercidae (Monogononta). Trans. R. Soc. South. Aust. 1992, 116, 1–27. [Google Scholar]
- Segers, H. Rotifera Vol. 2: The Lecanidae (Monogononta). In Guides to the Identification of the Microinvertebrates of the Continental Waters of the World 18; Nogrady, T., Dumont, H.J., Eds.; SPB Academic Publishing: The Hague, The Netherlands, 1995; pp. 1–225. ISBN 9051030916. [Google Scholar]
- Segers, H. Annotated checklist of the rotifers (Phylum Rotifera), with notes on nomenclature, taxonomy and distribution. Zootaxa 2007, 1564, 1–104. [Google Scholar] [CrossRef]
- Vollenweider, R.A.; Giovanardi, F.; Montanari, G.; Rinaldi, A. Characterization of the trophic conditions of marine coastal waters with special reference to the NW Adriatic Sea: Proposal for a trophic scale, turbidity and generalized water quality index. Environmetrics 1998, 9, 329–357. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: An Introduction to Ordination. Vegan Community Ecol. Packag. R Packag. version 2.4-1. 2016. Available online: https://CRAN.R-project.org/package=vegan (accessed on 10 January 2011).
- R Development Core Team. R Development Core Team, R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; ISBN 3900051070. [Google Scholar]
- Palmer, M.W. Estimating species richness: The second-order jackknife reconsidered. Ecology 1991, 72, 1512–1513. [Google Scholar] [CrossRef]
- Chao, A. Estimating the population size for Capture-Recapture data with unequal catchability. Biometrics 1987, 43, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.P.; van Belle, G. Nonparametric estimation of species richness. Biometrics 1984, 40, 119–129. [Google Scholar] [CrossRef] [Green Version]
- Trinh-Dang, M.; Van Vo, M.; Nguyen, A.; Tran, Q.; Thi, H.; Le, N.; Tran, S.N. Species diversity of rotifers (Rotifera: Eurotatoria) of Phu Ninh Lake with five new records from Vietnam. Int. J. Aquat. Biol. 2019, 7, 38–44. [Google Scholar]
- Trinh-Dang, M.; Segers, H.; Sanoamuang, L. Rotifers from Thuy Tien lake and Nhu Y river in central Vietnam, with a description of Ploesoma asiaticum new species (Rotifera: Monogononta). J. Limnol. 2013, 72, 376–386. [Google Scholar] [CrossRef] [Green Version]
- Trinh-Dang, M.; Segers, H.; Sanoamuang, L.O. Psammon rotifers in central Vietnam, with the descriptions of three new species (Rotifera: Monogononta). Zootaxa 2015, 4018, 249–265. [Google Scholar] [CrossRef]
- Trinh-Dang, M.; Phan, D.D.; Vo, V.M. A new rotifer species: Lecane langsenensis n. sp. (Rotifera: Monogononta) from Vietnam. Zootaxa 2019, 4615, 155–163. [Google Scholar] [CrossRef]
- Phan, D.D.; Le, T.N.N. Diversity on Rotifera species compatitions in fresh inland waters of Southern Vietnam and some new records for zooplankton fauna of Vietnam. J. Biol. 2012, 34, 13–20. [Google Scholar]
- Zhdanova, S.M. The species composition of rotifers in the water reservoirs of Central Vietnam. Inl. Water Biol. 2011, 4, 425–434. [Google Scholar] [CrossRef]
- Sa-Ardrit, P.; Pholpunthin, P.; Segers, H. A checklist of the freshwater rotifer fauna of Thailand (Rotifera, Monogononta, Bdelloidea). J. Limnol. 2013, 72, 361–375. [Google Scholar] [CrossRef] [Green Version]
- Meas, S.; Sanoamuang, L. Rotifer Communities in the Cambodian Mekong River Basins. Khon Kaen Univ. J. 2007, 08, 18–30. [Google Scholar] [CrossRef]
- Segers, H. Zoogeography of the Southeast Asian Rotifera. Hydrobiologia 2001, 446/447, 233–246. [Google Scholar] [CrossRef]
- Fernando, C.H.; Nora, P. The Rotifera of Malaysia and Singapore with remarks on some species. Hydrobiologia 1981, 78, 205–219. [Google Scholar] [CrossRef]
- Sanoamuang, L.; Segers, H.; Dumont, H.J. Additions to the rotifer fauna of south-east Asia: New and rare species from north-eas. Hydrobiologia 1995, 313/314, 35–45. [Google Scholar] [CrossRef]
- Dias, J.D.; Simões, N.R.; Meerhoff, M.; Lansac-Tôha, F.A.; Velho, L.F.M.; Bonecker, C.C. Hydrological dynamics drives zooplankton metacommunity structure in a Neotropical floodplain. Hydrobiologia 2016, 781, 109–125. [Google Scholar] [CrossRef]
- Wan Maznah, W.O.; Intan, S.; Sharifah, R.; Lim, C.C. Lentic and lotic assemblages of zooplankton in a tropical reservoir, and their association with water quality conditions. Int. J. Environ. Sci. Technol. 2018, 15, 533–542. [Google Scholar] [CrossRef]
- Andronikova, I.N. Zooplankton characteristics in monitoring of Lake Ladoga. In The First International Lake Ladoga Symposium; Springer: Dordrecht, The Netherlands, 1996; pp. 173–179. [Google Scholar]
- Park, G.S.; Marshall, H.G. Estuarine relationships between zooplankton community structure and trophic gradients. J. Plankton Res. 2000, 22, 121–135. [Google Scholar] [CrossRef]
- Paturej, E. Assessment of the Trophic State of A Restored Urban Lake Based on Zooplankton Community Structure and Zooplankton-Related Indices. Pol. J. Nat. Sci. 2008, 23, 440–449. [Google Scholar] [CrossRef]
- Starling, F.L. Comparative study of the zooplankton composition of six lacustrine ecosystems in central Brazil during the dry season. Rev. Bras. Biol. 2000, 60, 101–111. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Geng, H. Effect of Microcystis aeruginosa on the rotifer Brachionus calyciflorus at different temperatures. Bull. Environ. Contam. Toxicol. 2012, 88, 20–24. [Google Scholar] [CrossRef]
- Hart, R.C. Zooplankton distribution in relation to turbidity and related environmental gradients in a large subtropical reservoir: Patterns and implications. Freshw. Biol. 1990, 24, 241–263. [Google Scholar] [CrossRef]
- Kirk, K.L.; Gilbert, J.J. Suspended clay and the population dynamics of planktonic rotifers and cladocerans. Ecology 1990, 71, 1741–1755. [Google Scholar] [CrossRef]
- Pace, M.L.; Findlay, S.E.G.; Lints, D. Zooplankton in Advective Environments: The Hudson River Community and a Comparative Analysis. Can. J. Fish. Aquat. Sci. 1992, 49, 1060–1069. [Google Scholar] [CrossRef]
- Thorp, J.H.; Black, A.R.; Haag, K.H.; Wehr, J.D. Zooplankton assemblages in the Ohio River: Seasonal, tributary, and navigation dam effects. Can. J. Fish. Aquat. Sci. 1994, 51, 1634–1643. [Google Scholar] [CrossRef]
- Radwan, S. Planktonic rotifers as indicators of lake trophy. Ann. UMCS Sect. C 1976, 31, 227–235. [Google Scholar]
- Duggan, I.C. The ecology of periphytic rotifers. Hydrobiologia 2001, 446/447, 139–148. [Google Scholar] [CrossRef]
- Fontaneto, D.; Melone, G. Postembryonic Development of Hard Jaws (Trophi) in a Species Belonging to the Brachionus plicatilis Complex (Rotifera, Monogononta): A Morphometric Analysis. Microsc. Res. Tech. 2008, 71, 146–157. [Google Scholar] [CrossRef]
- Virro, T.; Haberman, J.; Haldna, M.; Blank, K. Diversity and structure of the winter rotifer assemblage in a shallow eutrophic northern temperate Lake Võrtsjärv. Aquat. Ecol. 2009, 43, 755–764. [Google Scholar] [CrossRef]
- Špoljar, M.; Habdija, I.; Primc-Habdija, B.; Sipos, L. Impact of environmental variables and food availability on rotifer assemblage in the karstic barrage Lake Visovac (Krka River, Croatia). Int. Rev. Hydrobiol. 2005, 90, 555–579. [Google Scholar] [CrossRef]
- Berziņš, B.; Pejler, B. Rotifer occurrence in relation to pH. Hydrobiologia 1989, 116, 223–231. [Google Scholar] [CrossRef]
- Iyer, N.; Rao, T.R. Responses of the predatory rotifer Asplanchna intermedia to prey species differing in vulnerability: Laboratory and field studies. Freshw. Biol. 1996, 36, 521–533. [Google Scholar] [CrossRef]
- Sarma, S.S.S.; Nandini, S.; Dumont, H.J. Feeding preference and population growth of Asplanchna brightwelli (Rotifera) offered two non-evasive prey rotifers. Hydrobiologia 1997, 361, 77–88. [Google Scholar] [CrossRef]
Figure 1.
The study area.

Figure 2.
Community structure of planktonic rotifers in Da Nang (the values on graph indicate the family name, number of taxa, and percentage).
Figure 2.
Community structure of planktonic rotifers in Da Nang (the values on graph indicate the family name, number of taxa, and percentage).

Figure 3.
Principal component analysis of water characteristics at 12 sampling sites (see Table 1 for the short codes of the sampling sites).
Figure 3.
Principal component analysis of water characteristics at 12 sampling sites (see Table 1 for the short codes of the sampling sites).

Figure 4.
Rotifer community structure at the family level in lakes and rivers in Da Nang.

Figure 5.
Species accumulation curves and species richness estimators of rotifers in lakes (left) and rivers (right) in Da Nang.
Figure 5.
Species accumulation curves and species richness estimators of rotifers in lakes (left) and rivers (right) in Da Nang.

Figure 6.
Canonical correspondence analysis of commonly found rotifer taxa and environmental variables in freshwater lakes in Da Nang (see Table 1 for the short codes of the sampling sites).
Figure 6.
Canonical correspondence analysis of commonly found rotifer taxa and environmental variables in freshwater lakes in Da Nang (see Table 1 for the short codes of the sampling sites).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Water quality characteristics at 12 sampling sites.
| Site | Code | Type | Region | Trophic State | TRIX | Temp (°C) | EC (mS/cm) | pH | Tur. (NTU) |
|---|---|---|---|---|---|---|---|---|---|
| Xanh Lake | LHX | Lake | Rural | Mesotrophic | 5.04 | 27.17 | 0.166 | 7.78 | 0.9 |
| Han River | RSH | River | Urban | Mesotrophic | 5.02 | 25.73 | 1.001 | 6.69 | 26.5 |
| Ham Nghi Lake | LHN | Lake | Urban | Eutrophic | 6.32 | 26.14 | 0.815 | 6.92 | 9.3 |
| Cong Vien Lake | LCV | Lake | Urban | Eutrophic | 6.47 | 26.47 | 0.728 | 6.92 | 24.3 |
| Sen Lake | LHS | Lake | Urban | Eutrophic | 6.58 | 26.27 | 0.469 | 6.86 | 24.6 |
| Bau Trang Lake | LBT | Lake | Urban | Mesotrophic | 5.63 | 26.37 | 0.556 | 7.20 | 13.1 |
| Truoc Dong Lake | LTD | Lake | Rural | Mesotrophic | 5.09 | 26.9 | 0.090 | 7.47 | 1.4 |
| Hoi Khe Lake | LHK | Lake | Rural | Oligotrophic | 4.82 | 27.32 | 0.056 | 6.61 | 25.6 |
| Hoi Phuoc River | RHP | River | Rural | Oligotrophic | 4.75 | 25.35 | 0.067 | 7.02 | 1.2 |
| Luong Dong River | RLD | River | Rural | Oligotrophic | 4.13 | 24.74 | 0.069 | 6.80 | 1.0 |
| Dong Xanh—Dong Nghe Lake | LDN | Lake | Rural | Oligotrophic | 4.6 | 26.39 | 0.050 | 6.72 | 1.7 |
| Cu De River | RCD | River | Rural | Oligotrophic | 4.22 | 25.9 | 0.079 | 7.41 | 2.2 |
Table 2.
Systematic list of the Rotifera species in freshwater habitats in Da Nang City, central Vietnam.
Table 2.
Systematic list of the Rotifera species in freshwater habitats in Da Nang City, central Vietnam.
| Conochilidae | Lecane closterocerca (Schmarda 1859) |
| Conochilus coenobasis (Skorikov 1914) | Lecane crepida (Harring 1914) |
| Conochilus dossuarius (Hudson 1885) | Lecane curvicornis (Murray 1913) |
| Hexarthridae | Lecane elsa Hauer 1931 |
| Hexarthra intermedia (Wiszniewski 1929) | Lecane grandis (Murray 1913) |
| Testudinellidae | Lecane haliclysta (Harring and Myers 1926) |
| Testudinella brevicaudata (Yamamoto 1951) | Lecane hamata (Stokes 1896) |
| Testudinella amphora (Hauer 1938) | Lecane hastata (Murray 1913) |
| Testudinella patina (Hermann 1783) | Lecane hornemanni (Ehrenberg 1834) |
| Testudinella tridentata (Smirnov 1931) | Lecane leontina (Turner 1892) |
| Testudinella incisa (Ternetz 1892) | Lecane ludwigii (Eckstein 1883) |
| Trochosphaeridae | Lecane luna (Müller 1776) |
| Filinia longiseta (Ehrenberg 1834) | Lecane lunaris (Ehrenberg 1832) |
| Asplanchnidae | Lecane obtusa (Murray 1913) |
| Asplanchna priodonta (Gosse 1850) | Lecane papuana (Murray 1913) |
| Asplanchna brightwellii (Gosse 1850) | Lecane punctata (Murray 1913) |
| Brachionidae | Lecane quadridentata (Ehrenberg 1830) |
| Anuraeopsis fissa (Gosse 1851) | Lecane ruttneri (Hauer 1938) |
| Anuraeopsis navicula (Rousselet 1911) | Lecane scutata (Harring and Myers 1926) |
| Keratella americana (Carlin 1943) | Lecane signifera (Jennings 1896) |
| Brachionus angularis (Gosse 1851) | Lecane yatseni (Wei and Xu 2010) |
| Brachionus budapestinensis (Daday 1885) | Lepadellidae |
| Brachionus calyciflorus (Pallas 1766) | Colurella obtusa (Gosse 1886) |
| Brachionus caudatus (Barrois and Daday 1894) | Lepadella ehrenbergii (Perty 1850) |
| Brachionus diversicornis (Daday 1883) | Lepadella ovalis (Müller 1786) |
| Brachionus durgae (Dhanapathi 1974) | Lepadella rhomboides (Gosse 1886) |
| Brachionus falcatus (Zacharias 1898) | Mytilinidae |
| Brachionus forficula (Wierzejski 1891) | Mytilina ventralis (Ehrenberg 1830) |
| Brachionus murphyi (Sudzuki 1989) | Notommatidae |
| Brachionus plicatilis (Müller 1786) | Cephalodella gibba (Ehrenberg 1830) |
| Brachionus quadridentatus (Hermann 1783) | Cephalodella gigantea (Remane 1933) |
| Keratella cochlearis (Gosse 1851) | Proalidae |
| Keratella quadrata (Müller 1786) | Proales sp. |
| Keratella tropica (Apstein 1907) | Synchaetidae |
| Platyias quadricornis (Ehrenberg 1832) | Polyarthra dolichoptera (Idelson 1925) |
| Dicranophoridae | Trichocercidae |
| Dicranophoroides claviger (Hauer 1965) | Trichocerca chattoni (Beauchamp 1907) |
| Dicranophorus prionacis (Harring and Myers 1928) | Trichocerca cylindrica (Imhof 1891) |
| Euchlanidae | Trichocerca dixonnuttalli (Jennings 1903) |
| Euchlanis dilatata (Ehrenberg 1832) | Trichocerca pusilla (Jennings 1903) |
| Tripleuchlanis plicata (Levander 1894) | Trichocerca similis (Wierzejski 1893) |
| Gastropodidae | Trichocerca stylata (Gosse 1851) |
| Ascomorpha ovalis (Bergendal 1892) | Trichotriidae |
| Ascomorpha saltans (Bartsch 1870) | Macrochaetus collinsii (Gosse 1867) |
| Lecanidae | Macrochaetus longipes (Myers 1934) |
| Lecane bulla (Gosse 1851) | Trichotria tetractis (Ehrenberg 1830) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Phan, N.-T.; Duong, Q.H.; Tran-Nguyen, Q.A.; Trinh-Dang, M. The Species Diversity of Tropical Freshwater Rotifers (Rotifera: Monogononta) in Relation to Environmental Factors. Water 2021, 13, 1156. https://doi.org/10.3390/w13091156
AMA Style
Phan N-T, Duong QH, Tran-Nguyen QA, Trinh-Dang M. The Species Diversity of Tropical Freshwater Rotifers (Rotifera: Monogononta) in Relation to Environmental Factors. Water. 2021; 13(9):1156. https://doi.org/10.3390/w13091156
Chicago/Turabian StylePhan, Nhat-Truong, Quang Hung Duong, Quynh Anh Tran-Nguyen, and Mau Trinh-Dang. 2021. "The Species Diversity of Tropical Freshwater Rotifers (Rotifera: Monogononta) in Relation to Environmental Factors" Water 13, no. 9: 1156. https://doi.org/10.3390/w13091156
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.