
Effect of Water Supply on Physiological Response and Phytonutrient Composition of Chili Peppers

 ,
, 
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Conditions
2.2. Irrigation System and Management
2.3. Measurements and Harvest
2.4. Physiological Responses
- Fo = initial fluorescence
- Fm = maximal fluorescence
- Fv = variable fluorescence (Fm − Fo).
2.5. Chemical Analyses of Phytonutrients
2.6. Statistical Analysis
3. Results
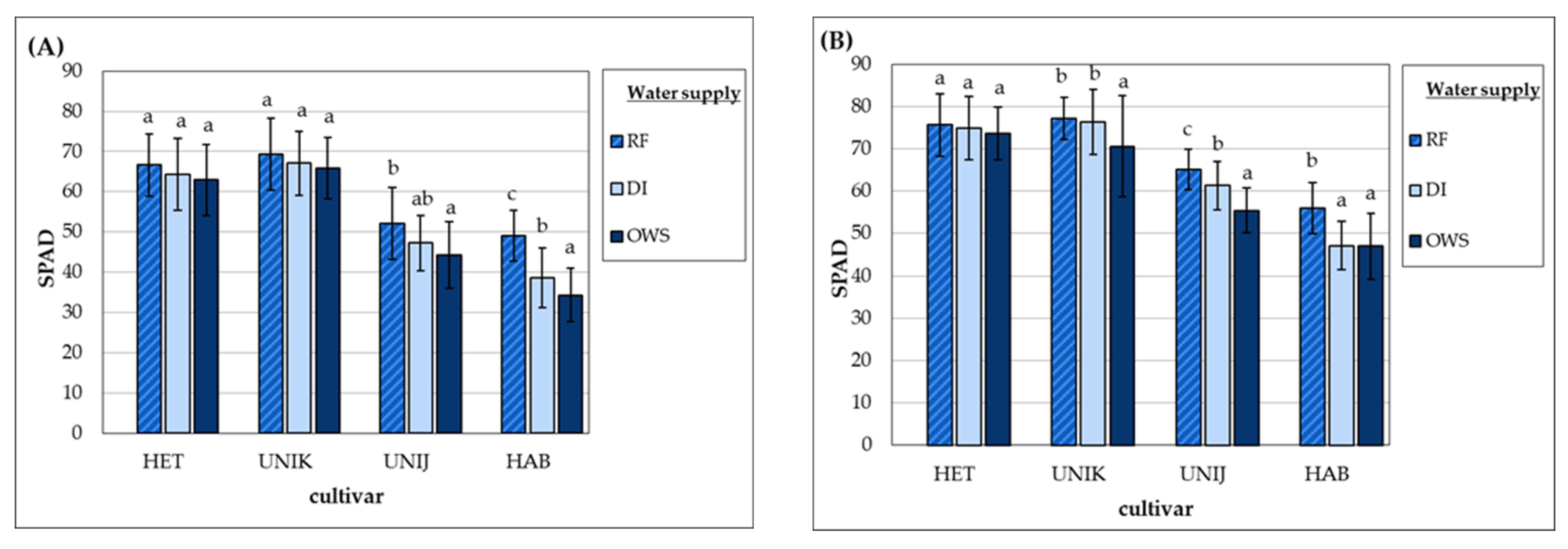
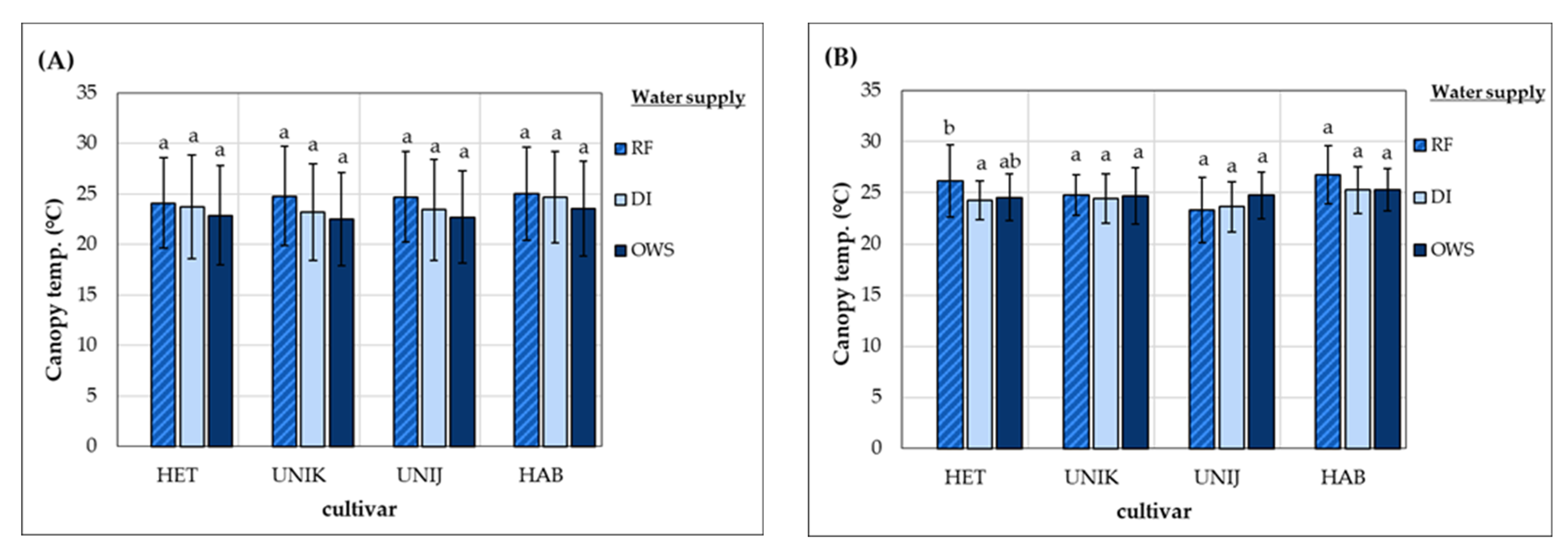
3.1. Effect of Water Supply on Physiological Responses of Cultivars during the Growth Period
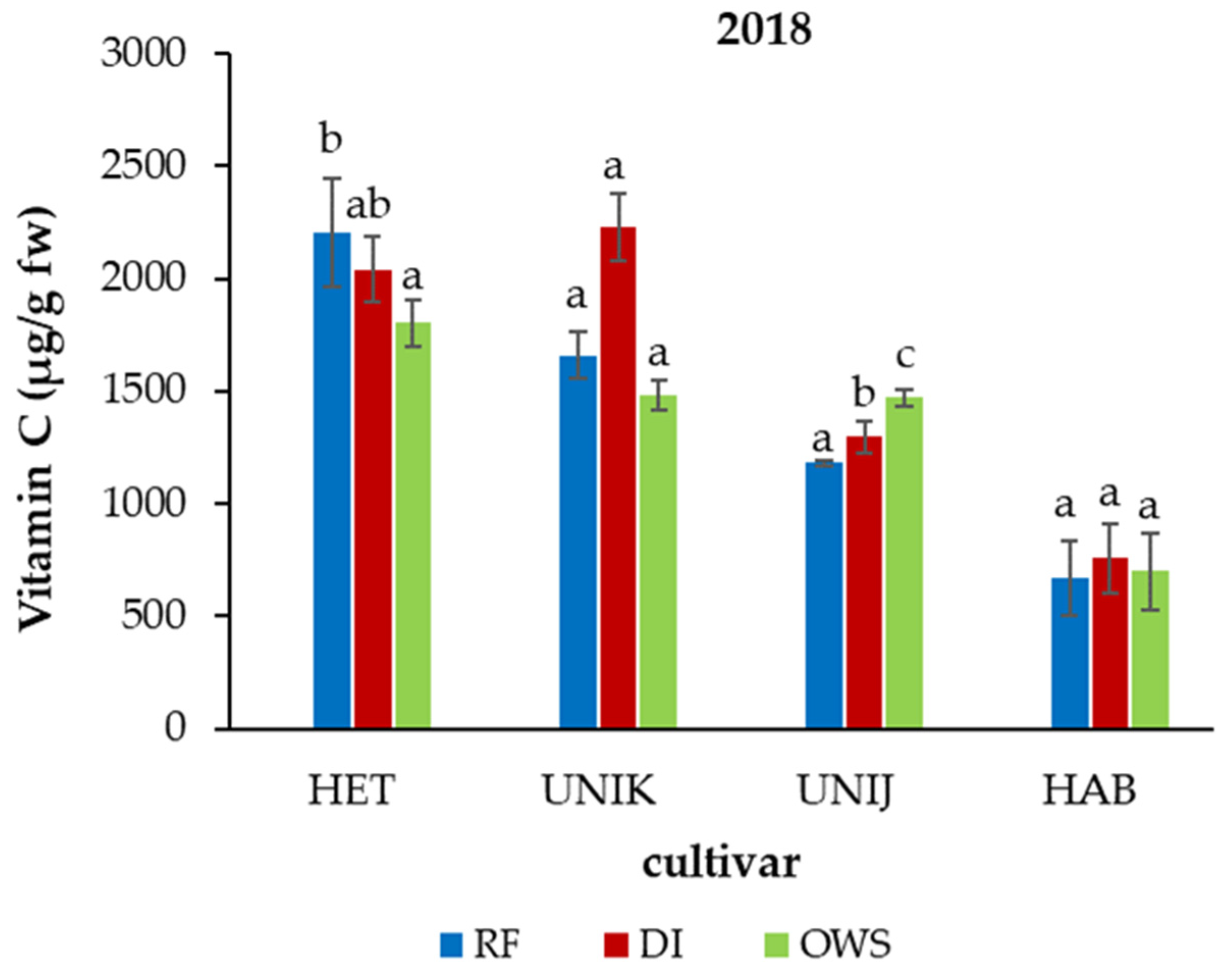
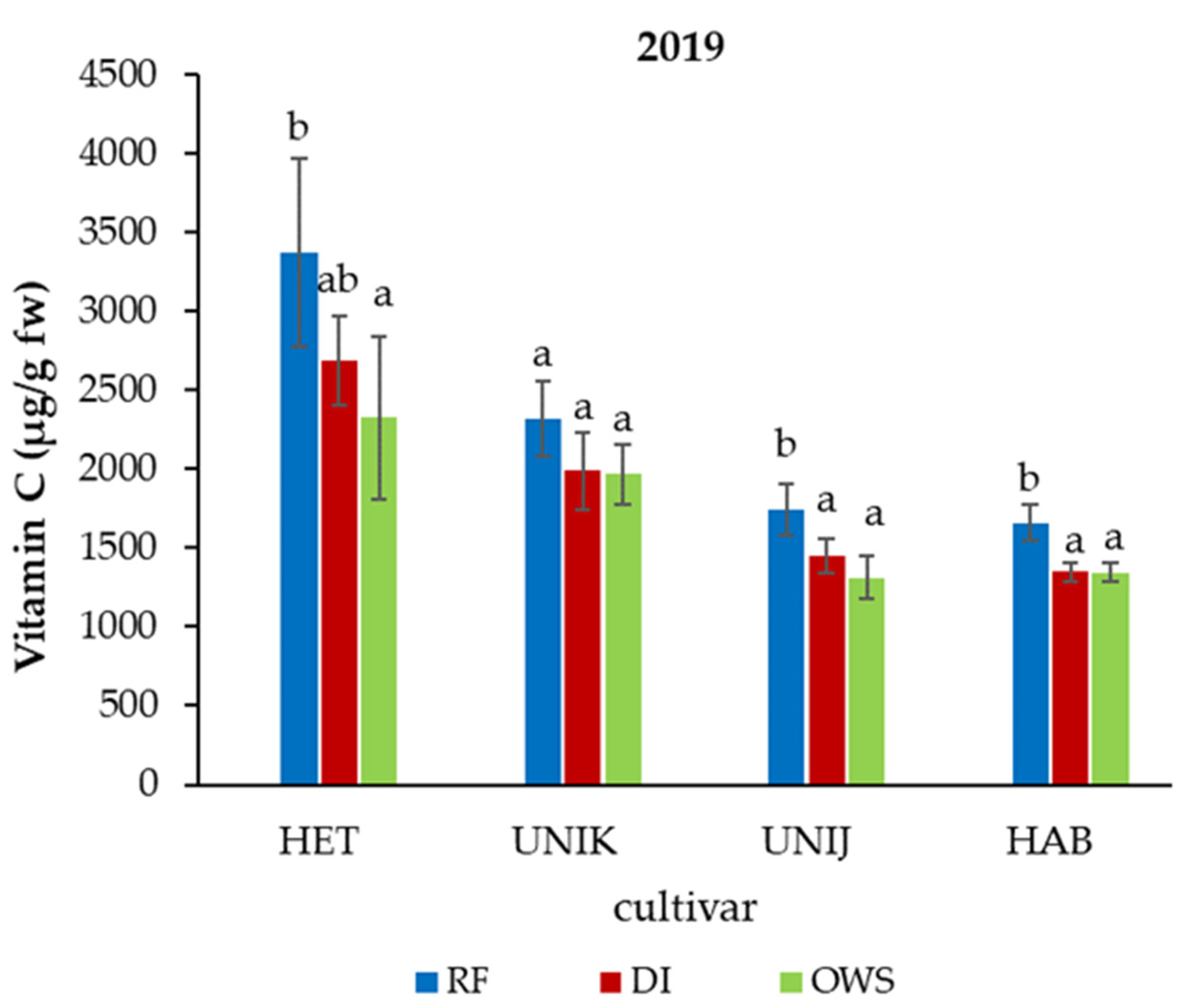
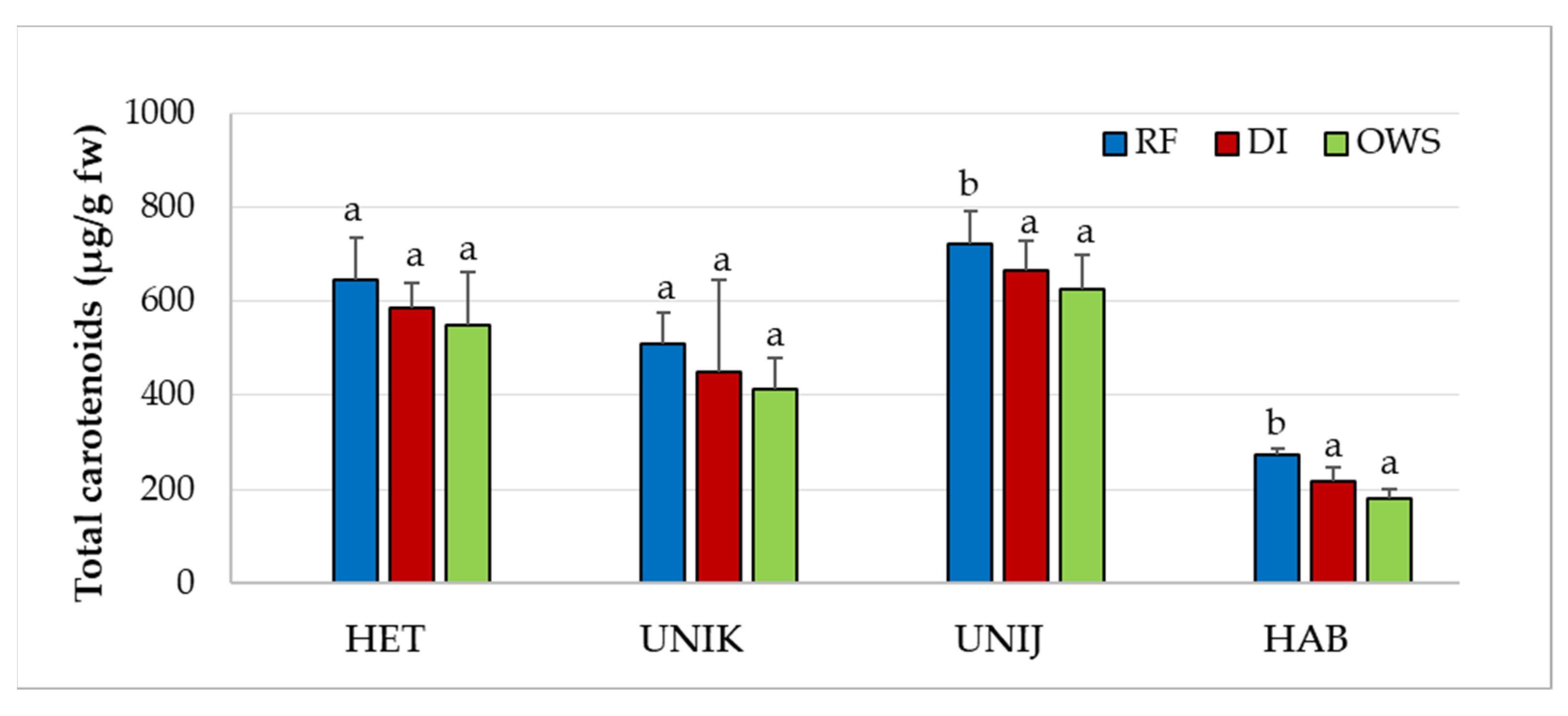
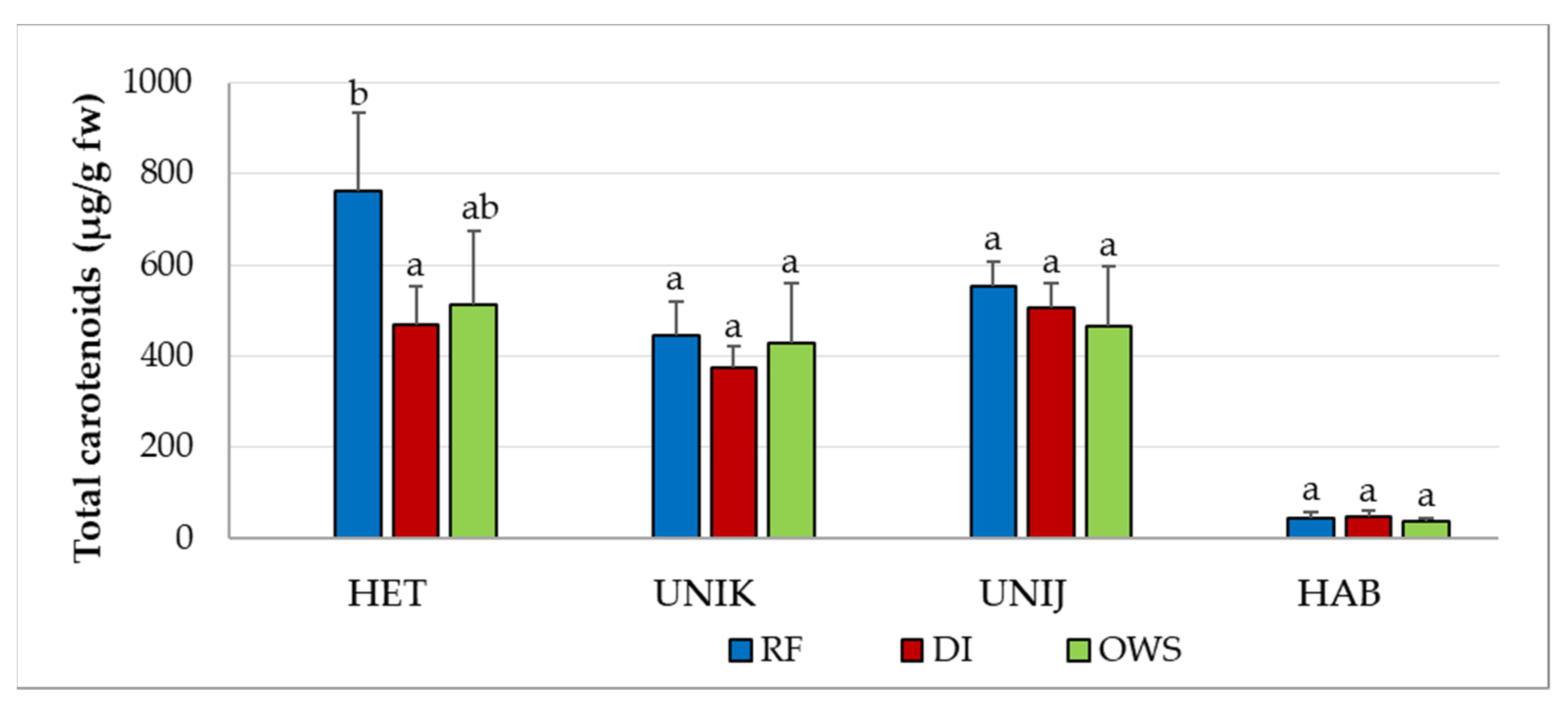
3.2. Effect of Water Supply on Phytonutrients in Chili Cultivars
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smith, M.; Jusztin, M. Paprika: The Spice of Life in Hungary. Spices Tour. Destin. Attract. Cuis. 2014, 38, 53. [Google Scholar]
- Fukuda, N.; Fujita, M.; Ohta, Y. Directional Blue Light Irradiation Triggers Epidermal Cell Elongation of Abaxial Side Resulting in Inhibition of Leaf Epinasty in Geranium under Red Light Condition. Sci. Hortic. 2008, 115, 176–182. [Google Scholar] [CrossRef]
- Dong, C.; Fu, Y.; Liu, G.; Liu, H. ScienceDirect Low Light Intensity Effects on the Growth, Photosynthetic Characteristics, Antioxidant Capacity, Yield and Quality of Wheat (Triticum aestivum L.) at Different Growth Stages in BLSS. Adv. Space Res. 2014, 53, 1557–1566. [Google Scholar] [CrossRef]
- González-Dugo, V.; Orgaz, F.; Fereres, E. Responses of Pepper to Deficit Irrigation for Paprika Production. Sci. Hortic. 2007, 114, 77–82. [Google Scholar] [CrossRef]
- Kang, Y.; Khan, S.; Ma, X. Climate Change Impacts on Crop Yield, Crop Water Productivity and Food Security—A Review. Prog. Nat. Sci. 2009, 19, 1665–1674. [Google Scholar] [CrossRef]
- Sezen, S.M.; Yazar, A.; Eker, S. Effect of Drip Irrigation Regimes on Yield and Quality of Field Grown Bell Pepper. Agric. Water Manag. 2006, 81, 115–131. [Google Scholar] [CrossRef]
- Stanghellini, C.; Kempkes, F.L.K.; Knies, P. Enhancing Environmental Quality in Agricultural Systems. Environ. Sci. 2003, 277–283. [Google Scholar] [CrossRef] [Green Version]
- Kirnak, H.; Naim Demirtas, M. Effects of Different Irrigation Regimes and Mulches on Yield and Macronutrition Levels of Drip-Irrigated Cucumber Under Open Field Conditions. J. Plant. Nutr. 2006, 29, 1675–1690. [Google Scholar] [CrossRef]
- Maisiri, N.; Senzanje, A. On Farm Evaluation of the Effect of Low Cost Drip Irrigation on Water and Crop Productivity Compared to Conventional Surface Irrigation System. Phys. Chem. Earth Parts A/B/C 2005, 30, 783–791. [Google Scholar] [CrossRef]
- Thornton, K.P.; Ericksen, J.P.; Herrero, M.; Challinor, J.A. Climate Variability and Vulnerability to Climate Change: A Review. Glob. Chang. Biol. 2014, 3313–3328. [Google Scholar] [CrossRef]
- Dorji, K.; Behboudian, M.H.; Zegbe-Domínguez, J.A. Water Relations, Growth, Yield, and Fruit Quality of Hot Pepper under Deficit Irrigation and Partial Rootzone Drying. Sci. Hortic. 2005, 104, 137–149. [Google Scholar] [CrossRef]
- Alvarez-Parrilla, E.; de la Rosa, L.A.; Amarowicz, R.; Shahidi, F. Antioxidant Activity of Fresh and Processed Jalapeno and Serrano Peppers. J. Agric. Food Chem. 2011, 59, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, M.W.; Momin, C.M.; Acharya, P.; Kabir, J.; Debnath, M.K.; Dhua, R.S. Dynamics of Changes in Bioactive Molecules and Antioxidant Potential of Capsicum Chinense Jacq. Cv. Habanero at Nine Maturity Stages. Acta Physiol. Plant 2013, 35, 1141–1148. [Google Scholar] [CrossRef]
- Ochi, T.; Takaishi, Y.; Kogure, K.; Yamauti, I. Antioxidant Activity of a New Capsaicin Derivative from Capsicum Annuum. J. Nat. Prod. 2003, 66, 1094–1096. [Google Scholar] [CrossRef] [PubMed]
- Topuz, A.; Ozdemir, F. Assessment of Carotenoids, Capsaicinoids and Ascorbic Acid Composition of Some Selected Pepper Cultivars (Capsicum Annuum L.) Grown in Turkey. J. Food Compos. Anal. 2007, 20, 596–602. [Google Scholar] [CrossRef]
- Materska, M.; Perucka, I. Antioxidant Activity of the Main Phenolic Compounds Isolated from Hot Pepper Fruit (Capsicum Annuum L.). J. Agric. Food Chem. 2005, 53, 1750–1756. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Peng, J.; Li, Y. Recent Advances in the Study on Capsaicinoids and Capsinoids. Eur. J. Pharmacol. 2011, 650, 1–7. [Google Scholar] [CrossRef]
- Satyanarayana, M.N. Capsaicin and Gastric Ulcers. Crit. Rev. Food Sci. Nutr. 2006, 46, 275–328. [Google Scholar] [CrossRef]
- Prabhat, C.Y.; Han, K.M.K.; Choi, K.; Lee, C.K.C. Capsaicin and Tocopherol in Red Pepper Seed Oil Enhances the Thermal Oxidative Stability during Frying. J. Food Sci. Technol. 2010, 47, 162–165. [Google Scholar] [CrossRef] [Green Version]
- Bae, H.; Jayaprakasha, G.K.; Crosby, K.; Sun, K.; Leskovar, D.I.; Jifon, J.; Patil, B.S. Ascorbic Acid, Capsaicinoid, and Flavonoid Aglycone Concentrations as a Function of Fruit Maturity Stage in Greenhouse-Grown Peppers. J. Food Compos. Anal. 2014, 33, 195–202. [Google Scholar] [CrossRef]
- Deng, L.Z.; Yang, X.H.; Mujumdar, A.S.; Zhao, J.H.; Wang, D.; Zhang, Q.; Wang, J.; Gao, Z.J.; Xiao, H.W. Red Pepper (Capsicum Annuum L.) Drying: Effects of Different Drying Methods on Drying Kinetics, Physicochemical Properties, Antioxidant Capacity, and Microstructure. Dry. Technol. 2018, 36, 893–907. [Google Scholar] [CrossRef]
- Krinsky, N.I.; Johnson, E.J. Carotenoid Actions and Their Relation to Health and Disease. Mol. Asp. Med. 2005, 26, 459–516. [Google Scholar] [CrossRef] [PubMed]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration-Guidelines for Computing Crop Water Requirements-FAO Irrigation and Drainage Paper 56; FAO: Rome, Italy, 1998; Volume 300, p. D05109. [Google Scholar]
- Ahmadi, S.H.; Mosallaeepour, E.; Kamgar-Haghighi, A.A.; Sepaskhah, A.R. Modeling Maize Yield and Soil Water Content with AquaCrop under Full and Deficit Irrigation Managements. Water Resour. Manag. 2015, 29, 2837–2853. [Google Scholar] [CrossRef]
- Jifon, J.L.; Syvertsen, J.P.; Whaley, E. Growth Environment and Leaf Anatomy Affect Nondestructive Estimates of Chlorophyll and Nitrogen in Citrus Sp. Leaves. J. Am. Soc. Hortic. Sci. 2005, 130, 152–158. [Google Scholar] [CrossRef] [Green Version]
- Van Goethem, D.; De Smedt, S.; Valcke, R.; Potters, G.; Samson, R. Seasonal, Diurnal and Vertical Variation of Chlorophyll Fluorescence on Phyllostachys Humilis in Ireland. PLoS ONE 2013, 8, e72145. [Google Scholar] [CrossRef]
- Nagy, Z.; Daood, H.; Ambrózy, Z.; Helyes, L. Determination of Polyphenols, Capsaicinoids, and Vitamin C in New Hybrids of Chili Peppers. J. Anal. Methods Chem. 2015, 2015. [Google Scholar] [CrossRef] [Green Version]
- Daood, H.G.; Halasz, G.; Palotás, G.; Palotás, G.; Bodai, Z.; Helyes, L. HPLC Determination of Capsaicinoids with Cross-Linked C18 Column and Buffer-Free Eluent. J. Chromatogr. Sci. 2015, 53, 135–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daood, H.G.; Palotás, G.; Palotás, G.; Somogyi, G.; Pék, Z.; Helyes, L. Carotenoid and Antioxidant Content of Ground Paprika from Indoor-Cultivated Traditional Varieties and New Hybrids of Spice Red Peppers. FRIN 2014, 65, 231–237. [Google Scholar] [CrossRef]
- Daood, H.G.; Vinkler, M.; Markus, F.; Hebshi, E.A.; Biacs, P.A. Antioxidant Vitamin Content of Spice Red Pepper (Paprika) as Affected by Technological and Varietal Factors. Food Chem. 1996, 55, 365–372. [Google Scholar] [CrossRef]
- Minguez-Mosquera, M.I.; Hornero-Mendez, D. Formation and Transformation of Pigments during the Fruit Ripening of Capsicum Annuum Cv. Bola and Agridulce. J. Agric. Food Chem. 1994, 42, 38–44. [Google Scholar] [CrossRef]
- Alonso, M.; Rozados, M.J.; Vega, J.A.; Perez-Gorostiaga, P.; Cuinas, P.; Fonturbel, M.T.; Fernandez, C. Biochemical Responses of Pinus Pinaster Trees to Fire-Induced Trunk Girdling and Crown Scorch: Secondary Metabolites and Pigments as Needle Chemical Indicators. J. Chem. Ecol. 2002, 28, 687–700. [Google Scholar] [CrossRef]
- Rufty, T.W.; Huber, S.C.; Volk, R.J. Alterations in Leaf Carbohydrate Metabolism in Response to Nitrogen Stress. Plant Physiol. 1988, 88, 725–730. [Google Scholar] [CrossRef] [Green Version]
- Radin, J.W.; Parker, L.L. Water Relations of Cotton Plants under Nitrogen Deficiency: I. Dependence upon Leaf Structure. Plant Physiol. 1979, 64, 495–498. [Google Scholar] [CrossRef]
- Bolhar-Nordenkampf, H.R.; Long, S.P.; Baker, N.R.; Oquist, G.; Schreiber, U.; Lechner, E.G. Chlorophyll Fluorescence as a Probe of the Photosynthetic Competence of Leaves in the Field: A Review of Current Instrumentation. Funct. Ecol. 1989, 497–514. [Google Scholar] [CrossRef]
- Agyemang Duah, S.; Nagy, Z.; Helyes, L. The Effect of Different Shading Net on the Quantum Efficiency of PS II in Chilli Pepper Cultivar ‘Star Flame’. Acta Agrar. Debr. 2019, 21–23. [Google Scholar] [CrossRef]
- Demmig-Adams, B.; Adams, W.W.I.I.I.; Logan, B.A.; Verhoeven, A.S. Xanthophyll Cycle-Dependent Energy Dissipation and Flexible Photosystem II Efficiency in Plants Acclimated to Light Stress. Funct. Plant Biol. 1995, 22, 249–260. [Google Scholar] [CrossRef]
- Morison, J.I.L. Sensitivity of Stomata and Water Use Efficiency to High CO2. Plant Cell Environ. 1985, 8, 467–474. [Google Scholar] [CrossRef]
- Loveys, B.R.; Egerton, J.J.G.; Ball, M.C. Higher Daytime Leaf Temperatures Contribute to Lower Freeze Tolerance under Elevated CO2. Plant Cell Environ. 2006, 29, 1077–1086. [Google Scholar] [CrossRef]
- Howard, L.R.; Talcott, S.T.; Brenes, C.H.; Villalon, B. Changes in phytochemical and antioxidant activity of selected pepper cultivars (Capsicum species) as influenced by maturity. J. Agric. Food Chem. 2000, 48, 1713–1720. [Google Scholar] [CrossRef]
- Navarro, J.M.; Flores, P.; Garrido, C.; Martinez, V. Food Chemistry Changes in the Contents of Antioxidant Compounds in Pepper fruits at different ripening stages, as affected by salinity. Food Chem. 2006, 96, 66–73. [Google Scholar] [CrossRef]
- Davies, M.B.; Austin, J.; Partridge, D.A. Vitamin C: Its Chemistry and Biochemistry; Royal Society of Chemistry: Cambridge, UK, 1991. [Google Scholar]
- Lee, S.K.; Kader, A.A. Preharvest and Postharvest Factors Influencing Vitamin C Content of Horticultural Crops. Postharvest Biol. Technol. 2000, 20, 207–220. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, A.F.; Yu, H.; Yang, X.; Jiang, W. Deficit Irrigation Affects Growth, Yield, Vitamin C Content, and Irrigation Water Use Efficiency of Hot Pepper Grown in soilless culture. HortScience 2014, 49, 722–728. [Google Scholar] [CrossRef]
- Gnayfeed, M.H.; Daood, H.G.; Biacs, P.A.; Alcaraz, C.F. Content of Bioactive Compounds in Pungent Spice Red Pepper (Paprika) as Affected by Ripening and Genotype. J. Sci. Food Agric. 2001, 1585, 1580–1585. [Google Scholar] [CrossRef]
- Pérez-López, A.J.; del Amor, F.M.; Serrano-Martínez, A.; Fortea, M.I.; Núñez-Delicado, E. Influence of Agricultural Practices on the Quality of Sweet Pepper Fruits as Affected by the Maturity Stage. J. Sci. Food Agric. 2007, 87, 2075–2080. [Google Scholar] [CrossRef]
- Kurian, A.L.; Starks, A.N. HPLC Analysis of Capsaicinoids Extracted from Whole Orange Habañero Chili Peppers. J. Food Sci. 2002, 67, 956–962. [Google Scholar] [CrossRef]
- Ruiz-Lau, N.; Medina-Lara, F.; Minero-García, Y.; Zamudio-Moreno, E.; Guzmán-Antonio, A.; Echevarría-Machado, I.; Martínez-Estévez, M. Water Deficit Affects the Accumulation of Capsaicinoids in Fruits of Capsicum Chinense Jacq. HortScience 2011, 46, 487–492. [Google Scholar] [CrossRef] [Green Version]
- Phimchan, P.; Techawongstien, S.; Chanthai, S.; Bosland, P.W. Impact of Drought Stress on the Accumulation of Capsaicinoids in Capsicum Cultivars with Different Initial Capsaicinoid Levels. HortScience 2012, 47, 1204–1209. [Google Scholar] [CrossRef] [Green Version]
- Jeeatid, N.; Techawongstien, S.; Suriharn, B.; Chanthai, S.; Bosland, P.W.; Techawongstien, S. Influence of Water Stresses on Capsaicinoid Production in Hot Pepper (Capsicum Chinense Jacq.) Cultivars with Different Pungency Levels. Food Chem. 2018, 245, 792–797. [Google Scholar] [CrossRef] [PubMed]
- Harvell, K.P.; Bosland, P.W. The Environment Produces a Significant Effect on Pungency of Chiles. HortScience 1997, 32, 1292. [Google Scholar] [CrossRef] [Green Version]
- González-Zamora, A.; Sierra-Campos, E.; Luna-Ortega, J.G.; Pérez-Morales, R.; Ortiz, J.C.R.; García-Hernández, J.L. Characterization of Different Capsicum Varieties by Evaluation of Their Capsaicinoids Content by High Performance Liquid Chromatography, Determination of Pungency and Effect of High Temperature. Molecules 2013, 18, 13471–13486. [Google Scholar] [CrossRef]
- Gurung, T.; Techawongstien, S.; Suriharn, B.; Techawongstien, S. Impact of Environments on the Accumulation of Capsaicinoids in Capsicum spp. HortScience 2011, 46, 1576–1581. [Google Scholar] [CrossRef]
- Iqbal, Q.; Amjad, M.; Asi, M.R.; Ariño, A. Characterization of Capsaicinoids and Antioxidants in Hot Peppers as Influenced by Hybrid and Harvesting Stage. Plant Foods Hum. Nutr. 2013, 358–363. [Google Scholar] [CrossRef]
- Arimboor, R.; Natarajan, R.B. Red Pepper (Capsicum annuum) Carotenoids as a Source of Natural Food Colors: Analysis and Stability—A Review. J. Food Sci. Technol. 2015, 52, 1258–1271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Azevedo-Meleiro, C.H.; Rodriguez-Amaya, D.B. Qualitative and Quantitative Differences in the Carotenoid Composition of Yellow and Red Peppers Determined by HPLC-DAD-MS. J. Sep. Sci. 2009, 32, 3652–3658. [Google Scholar] [CrossRef] [PubMed]
- Davies, B.H.; Matthews, S.; Kirk, J.T.O. The Nature and Biosynthesis of the Carotenoids of Different Colour Varieties of Capsicum Annuum. Phytochemistry 1970, 9, 797–805. [Google Scholar] [CrossRef]
- Deli, J.; Matus, Z.; Szabolcs, J. Carotenoid Composition in the Fruits of Black Paprika (Capsicum Annuum Variety Longum Nigrum) during Ripening. J. Agric. Food Chem. 1992, 40, 2072–2076. [Google Scholar] [CrossRef]
- Ha, S.-H.; Kim, J.-B.; Park, J.-S.; Lee, S.-W.; Cho, K.-J. A Comparison of the Carotenoid Accumulation in Capsicum Varieties That Show Different Ripening Colours: Deletion of the Capsanthin-Capsorubin Synthase Gene Is Not a Prerequisite for the Formation of a Yellow Pepper. J. Exp. Bot. 2007, 58, 3135–3144. [Google Scholar] [CrossRef] [Green Version]
- Ornelas-Paz, J.D.J.; Cira-chávez, L.A.; Gardea-béjar, A.A.; Guevara-arauza, J.C.; Sepúlveda, D.R.; Reyes-hernández, J.; Ruiz-cruz, S. Effect of Heat Treatment on the Content of Some Bioactive Compounds and Free Radical-Scavenging Activity in Pungent and Non-Pungent Peppers. FRIN 2013, 50, 519–525. [Google Scholar] [CrossRef]
- Carnevale, J.; Cole, E.R.; Crank, G. Photocatalyzed Oxidation of Paprika Pigments. J. Agric. Food Chem. 1980, 28, 953–956. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Mean Temperature (°C) | Mean Relative Humidity (%) | Precipitation (mm) | Irrigation (mm) | Total Water Received by Plant Stands (mm) 1 | |||
|---|---|---|---|---|---|---|---|---|
| DI | OWS | RF | DI | OWS | ||||
| 2018 | 23.8 | 71.0 | 347.8 | 132.6 | 272.2 | 347.8 | 480.4 | 620.0 |
| 2019 | 25.8 | 72.3 | 132.6 | 152.2 | 289.0 | 132.6 | 284.4 | 421.6 |
| Cultivar | Water Supply | NDC (µg/g) | CAP (µg/g) | DC (µg/g) | HCAP (µg/g) | iDC (µg/g) | HDCs (µg/g) |
|---|---|---|---|---|---|---|---|
| HET | RF | 9.6 ± 0.2 a | 58.2 ± 14.4 a | 33.6 ± 3.5 a | 1.17 ± 0.1 a | 4.1 ± 0.5 a | 5.32 ± 0.7 a |
| DI | 9.3 ± 1.8 a | 69.7 ± 11.6 a | 43.7 ± 2.3 a | 1.24 ± 0.1 a | 3.5 ± 0.6 a | 4.17 ± 0.7 a | |
| OWS | 8.9 ± 1.9 a | 62.1 ± 12.2 a | 41.1 ± 4.4 a | 1.2 ± 0.2 a | 3.7 ± 0.7 a | 4.11 ± 0.4 a | |
| F-value | 0.119 | 0.749 | 0.286 | 0.061 | 0.516 | 0.720 | |
| p value | 0.889 | 0.500 | 0.758 | 0.941 | 0.614 | 0.513 | |
| UNIK | RF | 2.8 ± 0.1 a | 17.3 ± 1.8 a | 13.5 ± 1.8 a | ND | 1.05 ± 0.40 a | 1.0 ± 0.1 a |
| DI | 4.0 ± 0.3 a | 29.2 ± 2.9 b | 19.8 ± 3.2 a | ND | 1.6 ± 0.35 a | 1.6 ± 0.2 a | |
| OWS | 3.7 ± 0.4 a | 30.3 ± 4.9 b | 18.5 ± 1.2 a | ND | 0.9 ± 0.35 a | 1.7 ± 0.2 a | |
| F-value | 1.360 | 6.908 | 2.640 | 3.900 | 2.053 | ||
| p value | 0.305 | 0.015 | 0.125 | 0.060 | 0.184 | ||
| UNIJ | RF | 116.4 ± 11.6 a | 1282.7 ± 137.3 a | 796.2 ± 36.9 a | ND | 41.1 ± 3.4 a | 35.1 ± 2.2 a |
| DI | 117.2 ± 9.3 a | 1239.9 ± 142.3 a | 766.5 ± 61.2 a | ND | 44.6 ± 4.1 a | 36.2 ± 4.8 a | |
| OWS | 112.4 ± 14.5 a | 1213.6 ± 153.4 a | 744.6 ± 71.4 a | ND | 31.0 ± 2.61 a | 33.2 ± 4.2 a | |
| F-value | 0.341 | 0.157 | 0.790 | 1.033 | 0.103 | ||
| p value | 0.720 | 0.857 | 0.483 | 0.394 | 0.903 | ||
| HAB | RF | 60.3 ± 7.5 a | 1822.1 ± 121.3 a | 996.4 ± 108.6 a | ND | 45.0 ± 5.4 a | 22.7 ± 2.2 a |
| DI | 79.8 ± 10.3 a | 2191.8 ± 247 a | 1080.6 ± 119.2 a | ND | 43.8 ± 4.40 a | 24.4 ± 5.3 a | |
| OWS | 75.6 ± 13.2 a | 2130.6 ± 216.3 a | 1029.8 ± 189.1 a | ND | 64.8 ± 7.8 a | 25.2 ± 7.5 a | |
| F-value | 0.407 | 0.696 | 0.301 | 0.694 | 0.221 | ||
| p value | 0.677 | 0.524 | 0.747 | 0.524 | 0.806 |
| Cultivar | Water Supply | NDC (µg/g) | CAP (µg/g) | DC (µg/g) | HCAP (µg/g) | iDC (µg/g) | HDCs (µg/g) |
|---|---|---|---|---|---|---|---|
| HET | RF | 42.9 ± 10.5 b | 405.1 ± 67.0 a | 250.5 ± 46.6 b | 2.1 ± 0.3 a | 7.2 ± 1.0 b | 24.4 ± 4.1 b |
| DI | 27.1 ± 6.4 a | 255.6 ± 29.4 a | 151.0 ± 25.8 a | 1.5 ± 0.1 a | 4.0 ± 1.0 a | 14.1 ± 2.9 a | |
| OWS | 20.3 ± 4.2 a | 238.7 ± 27.7 a | 124.3 ± 20.1 a | 1.5 ± 0.1 a | 2.4 ± 0.40 a | 11.1 ± 1.8 a | |
| F-value | 9.456 | 3.595 | 6.298 | 0.600 | 29.328 | 9.318 | |
| p value | 0.006 | 0.071 | 0.019 | 0.569 | ≤0.001 | 0.006 | |
| UNIK | RF | 18.2 ± 2.1 a | 232.4 ± 31.1 a | 99.4 ± 18.3 a | 1.8 ± 0.1 a | 2.8 ± 0.3 b | 13.3 ± 1.3 a |
| DI | 17.8 ± 2.9 a | 131.6 ± 17.3 a | 80.1 ± 9.3 a | 1.5 ± 0.1 a | 2.2 ± 0.3 b | 12.7 ± 2.7 a | |
| OWS | 13.8 ± 2.3 a | 98.2 ± 16.5 a | 58.6 ± 6.8 a | 1.2 ± 0.1 a | 1.7 ± 0.1 a | 7.3 ± 1.1 a | |
| F-value | 1.564 | 3.709 | 1.075 | 3.268 | 8.587 | 2.060 | |
| p value | 0.261 | 0.067 | 0.381 | 0.086 | 0.008 | 0.183 | |
| UNIJ | RF | 151.4 ± 21.7 b | 1662.5 ± 235.4 b | 1108.6 ± 156.8 b | ND | 39.4 ± 11.2 b | 53.4 ± 5.9 b |
| DI | 117.8 ± 7.7 a | 1796.7 ± 242.7 b | 1050.5 ± 83.1 a | ND | 23.6 ± 7.8 a | 38.5 ± 1.6 b | |
| OWS | 87.0 ± 18.9 a | 1513.5 ± 139.4 a | 743.7 ± 47.6 a | ND | 16.6 ± 5.2 a | 27.8 ± 1.0 a | |
| F-value | 22.795 | 65.553 | 76.909 | 7.636 | 9.032 | ||
| p value | ≤0.001 | ≤0.001 | ≤0.001 | 0.012 | 0.007 | ||
| HAB | RF | 108.5 ± 19.6 b | 2744.3 ± 316.9 b | 1323.8 ± 155.8 c | 21.7 ± 1.0 a | 56.0 ± 11.3 b | 51 ± 4.5 b |
| DI | 63.9 ± 8.3 a | 2969.7 ± 162.0 b | 1034.2 ± 99.4 b | 14.0 ± 1.6 a | 33.5 ± 8.2 a | 28.8 ± 6.6 a | |
| OWS | 42.9 ± 3.4 a | 2392.2 ± 262.6 a | 742.8 ± 75.9 a | 9.8 ± 0.1 a | 22.3 ± 1.3 a | 22.3 ± 1.0 a | |
| F-value | 29.081 | 5.188 | 25.357 | 0.500 | 9.831 | 27.071 | |
| p value | ≤0.001 | 0.032 | ≤0.001 | 0.622 | 0.005 | ≤0.001 |
| Cultivar | Water Supply | Free Caps. (µg/g) | Free Zeax. (µg/g) | Caps ME (µg/g) | Zeax ME (µg/g) | β-Carotene (µg/g) | Caps DE (µg/g) | Zeax DE (µg/g) |
|---|---|---|---|---|---|---|---|---|
| HET | RF | 36.9 ± 5.4 b | 4.1 ± 1.5 a | 54.5 ± 6.5 b | 7.6 ± 0.7 a | 40.8 ± 5.0 a | 215.7 ± 3.3 a | 59.6 ± 1.7 b |
| DI | 35.4 ± 2.9 b | 2.3 ± 0.3 a | 53.7 ± 5.5 b | 8.0 ± 0.7 a | 36.3 ± 0.6 a | 225.3 ± 22.5 a | 45.2 ± 0.9 b | |
| OWS | 26.5 ± 2.7 a | 2.5 ± 0.9 a | 37.6 ± 5.2 a | 8.6 ± 2.7 a | 28.7 ± 3.2 a | 148.1 ± 16.4 a | 34.7 ± 1.2 a | |
| F value | 8.279 | 3.680 | 11.058 | 0.312 | 0.958 | 2.654 | 84.118 | |
| p value | 0.009 | 0.068 | 0.004 | 0.740 | 0.419 | 0.124 | ≤0.001 | |
| UNIK | RF | 42.8 ± 7.6 a | 6.4 ± 0.8 a | 45.2 ± 6.8 a | 11.3 ± 2.1 a | 38.4 ± 2.7 a | 128.3 ± 12.4 a | 33.6 ± 2.0 a |
| DI | 56.6 ± 8.5 a | 5.9 ± 0.7 a | 49.4 ± 5.2 a | 15.0 ± 1.3 a | 28.9 ± 4.7 a | 126.0 ± 16.8 a | 19.7 ± 3.3 a | |
| OWS | 32.3 ± 3.1 a | 3.8 ± 0.4 a | 38.7 ± 3.5 a | 8.2 ± 1.2 a | 34.7 ± 4.5 a | 120.4 ± 23.2 a | 15.9 ± 0.6 a | |
| F value | 1.359 | 1.503 | 0.699 | 0.770 | 0.152 | 0.176 | 0.739 | |
| p value | 0.305 | 0.273 | 0.522 | 0.491 | 0.861 | 0.842 | 0.504 | |
| UNIJ | RF | 51.2 ± 5.6 b | 8.8 ± 1.8 a | 54.2 ± 7.1 b | 23.0 ± 3.5 b | 64.1 ± 2.7 b | 177.8 ± 16.9 b | 58.5 ± 9.4 a |
| DI | 24.7 ± 3.3 a | 5.9 ± 1.7 a | 31.9 ± 3.7 a | 15.7 ± 2.5 a | 40.1 ± 1.6 a | 114.3 ± 12.2 a | 42.8 ± 4.6 a | |
| OWS | 23.0 ± 3.5 a | 6.7 ± 0.8 a | 29.4 ± 4.3 a | 17.1 ± 2.2 a | 37.1 ± 1.9 a | 114.9 ± 19.9 a | 39.9 ± 5.6 a | |
| F value | 9.079 | 0.836 | 22.890 | 7.809 | 18.957 | 13.558 | 2.063 | |
| p value | 0.007 | 0.464 | ≤0.001 | 0.011 | 0.001 | 0.002 | 0.183 | |
| HAB | RF | 1.9 ± 0.2 b | 0.4 ± 0.0 b | 1.2 ± 0.1 a | ND | 5.3 ± 2.4 a | 3.6 ± 2.1 a | 2.2 ± 0.3 a |
| DI | 0.6 ± 0.1 a | 0.1 ± 0.0 a | 0.7 ± 0.2 a | ND | 6.5 ± 2.9 a | 4.9 ± 2.3 a | 1.1 ± 0.1 a | |
| OWS | 0.6 ± 0.1 a | ND | 0.5 ± 0.1 a | ND | 6.6 ± 1.2 a | 5.7 ± 1.6 a | 1.3 ± 0.1 a | |
| F value | 77.448 | 8.203 | 2.433 | 2.625 | 1.066 | 1.177 | ||
| p value | ≤0.001 | 0.009 | 0.143 | 0.126 | 0.384 | 0.351 |
| Cultivar | Water Supply | Free Caps. (µg/g) | Free Zeax. (µg/g) | Caps ME (µg/g) | Zeax ME (µg/g) | β-Carotene (µg/g) | Caps DE (µg/g) | Zeax DE (µg/g) |
|---|---|---|---|---|---|---|---|---|
| HET | RF | 1.9 ± 0.4 b | 15.1 ± 3.5 b | 57.5 ± 6.1 b | 59.6 ± 1.7 a | 41.9 ± 2.8 a | 256.3 ± 42.0 b | 51.2 ± 5.2 a |
| DI | 1.0 ± 0.3 a | 8.8 ± 2.0 a | 33.9 ± 3.0 a | 45.2 ± 0.9 a | 31.7 ± 0.8 a | 152.1 ± 31.6 a | 34.2 ± 2.8 a | |
| OWS | 1.2 ± 0.4 a | 9.4 ± 3.9 a | 37.7 ± 5.8 ab | 34.7 ± 1.2 a | 31.7 ± 4.0 a | 171.7 ± 31.6 ab | 38.1 ± 8.0 a | |
| F value | 9.196 | 4.559 | 6.091 | 3.731 | 2.926 | 5.416 | 2.314 | |
| p value | 0.007 | 0.043 | 0.021 | 0.066 | 0.105 | 0.029 | 0.155 | |
| UNIK | RF | 1.1 ± 0.5 a | 10.6 ± 2.4 a | 27.0 ± 3.7 a | 33.6 ± 2.0 a | 40.2 ± 4.5 a | 132.8 ± 21.7 a | 13.3 ± 2.1 a |
| DI | 1.0 ± 0.2 a | 8.7 ± 0.9 a | 52.0 ± 7.6 b | 19.7 ± 3.3 a | 29.8 ± 2.6 a | 136.4 ± 12.9 a | 11.8 ± 1.8 a | |
| OWS | 1.7 ± 0.8 a | 11.6 ± 4.7 a | 56.7 ± 6.1 b | 15.9 ± 0.6 a | 31.4 ± 3.2 a | 142.9 ± 18.1 a | 8.9 ± 1.2 a | |
| F value | 1.859 | 0.897 | 9.279 | 0.533 | 0.317 | 0.095 | 1.108 | |
| p value | 0.211 | 0.441 | 0.007 | 0.604 | 0.736 | 0.910 | 0.371 | |
| UNIJ | RF | 1.3 ± 0.5 a | 10.2 ± 1.3 a | 46.9 ± 5.5 a | 58.5 ± 9.4 b | 58.6 ± 4.4 a | 185.7 ± 19.6 b | 34.6 ± 6.9 b |
| DI | 1.2 ± 0.6 a | 9.8 ± 1.0 a | 43.5 ± 4.6 a | 42.8 ± 4.6 a | 43.9 ± 3.4 a | 144.1 ± 9.8 a | 34.8 ± 4.5 b | |
| OWS | 0.8 ± 0.6 a | 8.1 ± 1.3 a | 39.0 ± 4.3 a | 39.9 ± 5.6 a | 40.4 ± 10.0 a | 157.4 ± 17.1 b | 29.4 ± 0.6 a | |
| F value | 0.936 | 0.465 | 1.586 | 17.168 | 0.124 | 29.924 | 31.036 | |
| p value | 0.427 | 0.642 | 0.257 | 0.001 | 0.885 | ≤0.001 | ≤0.001 | |
| HAB | RF | 0.3 ± 0.0 b | tr | 3.9 ± 0.8 ab | 0.7 ± 0.1 a | 5.3 ± 2.4 a | 5.9 ± 0.9 a | 4.6 ± 1.1 b |
| DI | 0.1 ± 0.0 a | tr | 2.2 ± 0.6 a | 0.8 ± 0.1 a | 6.5 ± 2.9 a | 17.2 ± 3.2 b | 3.2 ± 0.7 ab | |
| OWS | ND | tr | 5.4 ± 0.8 b | 0.6 ± 0.1 a | 6.6 ± 1.2 a | 14.9 ± 1.4 b | 2.5 ± 0.3 a | |
| F value | 30.160 | 4.658 | 0.549 | 1.494 | 9.548 | 4.377 | ||
| p value | ≤0.001 | 0.041 | 0.596 | 0.275 | 0.006 | 0.047 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agyemang Duah, S.; Silva e Souza, C.; Nagy, Z.; Pék, Z.; Neményi, A.; Daood, H.G.; Vinogradov, S.; Helyes, L. Effect of Water Supply on Physiological Response and Phytonutrient Composition of Chili Peppers. Water 2021, 13, 1284. https://doi.org/10.3390/w13091284
Agyemang Duah S, Silva e Souza C, Nagy Z, Pék Z, Neményi A, Daood HG, Vinogradov S, Helyes L. Effect of Water Supply on Physiological Response and Phytonutrient Composition of Chili Peppers. Water. 2021; 13(9):1284. https://doi.org/10.3390/w13091284
Chicago/Turabian StyleAgyemang Duah, Stella, Clarice Silva e Souza, Zsuzsa Nagy, Zoltán Pék, András Neményi, Hussein G. Daood, Szergej Vinogradov, and Lajos Helyes. 2021. "Effect of Water Supply on Physiological Response and Phytonutrient Composition of Chili Peppers" Water 13, no. 9: 1284. https://doi.org/10.3390/w13091284