
Flow Intermittency Affects Leaf Decomposition and Benthic Consumer Communities of Alpine Streams: A Case Study along the Po River

 ,
,  , , ,
, , ,  , and
, and
Abstract
:
1. Introduction
2. Materials and Methods
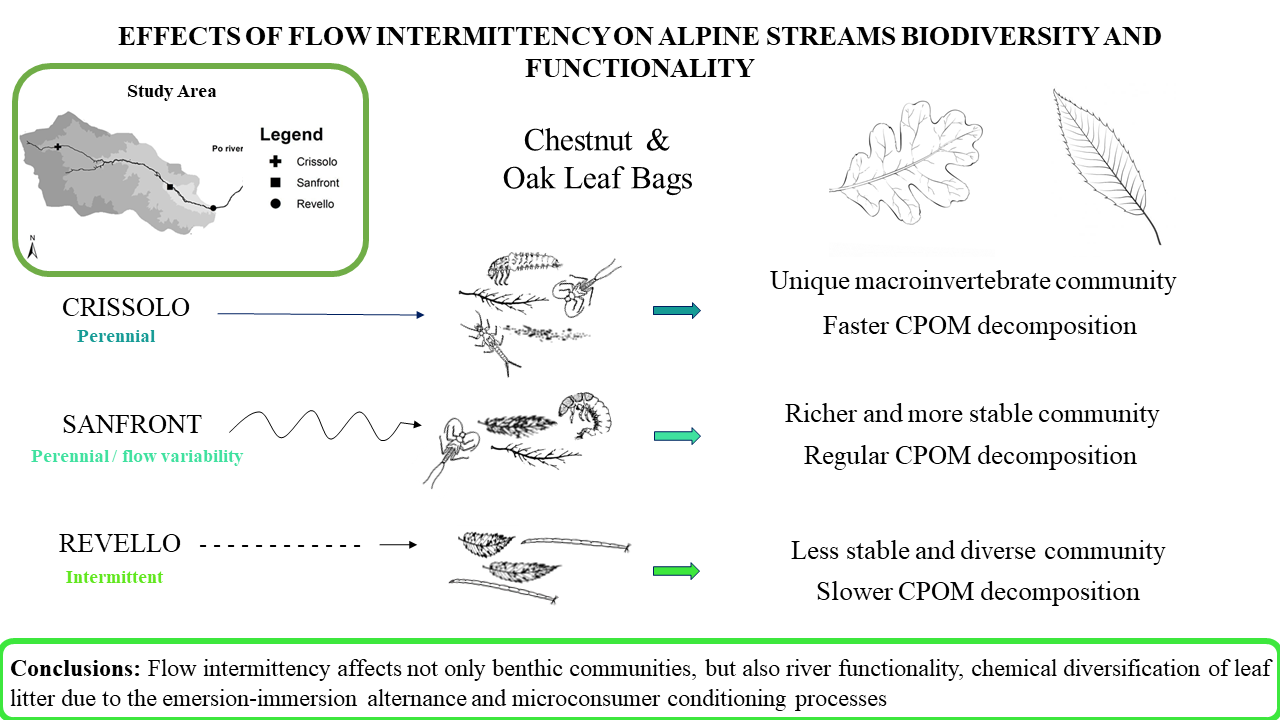
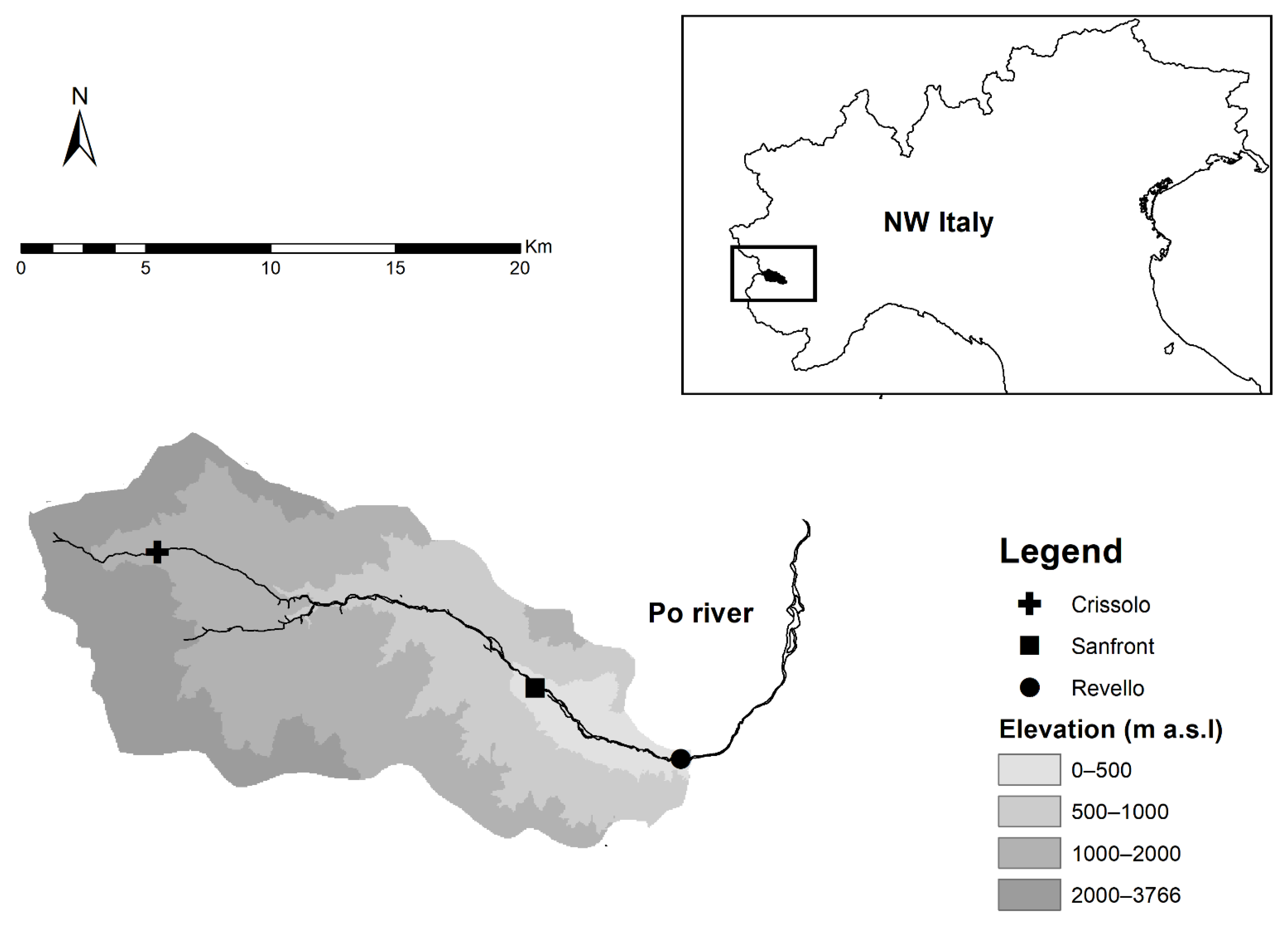
2.1. Study Site
2.2. Experimental Design
2.3. Data Analysis
3. Results
3.1. Environmental Conditions
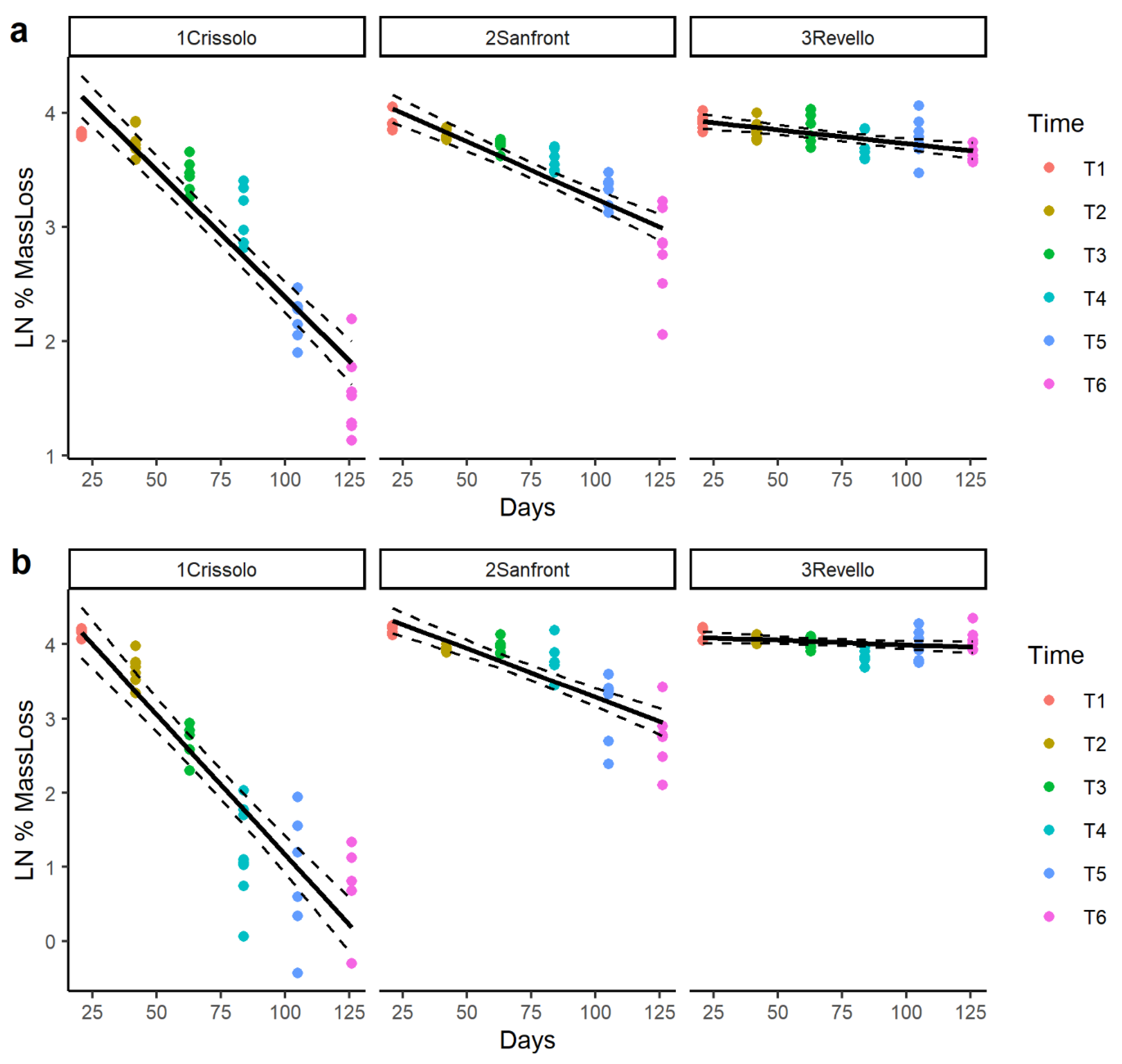
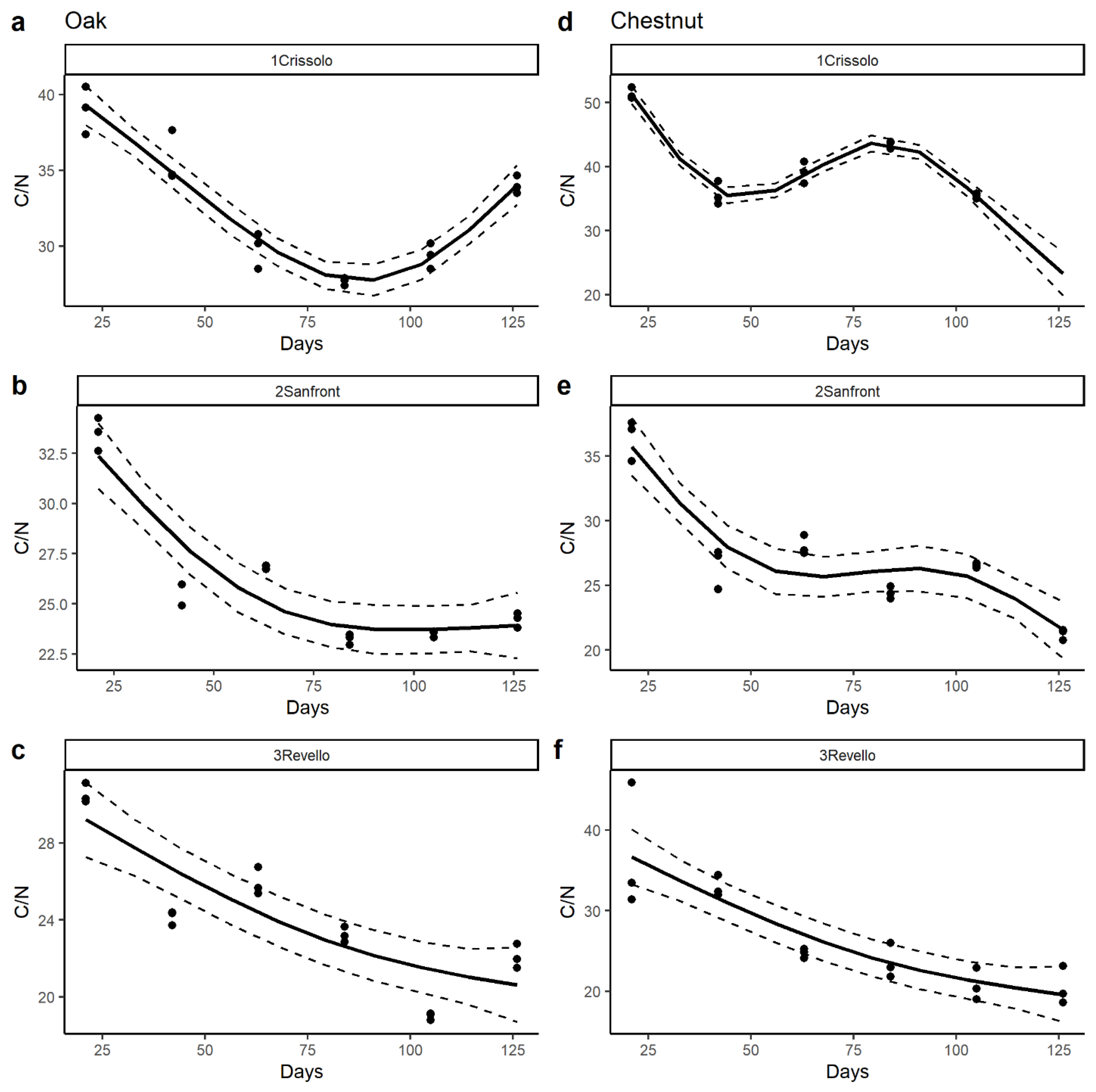
3.2. Leaf Decomposition
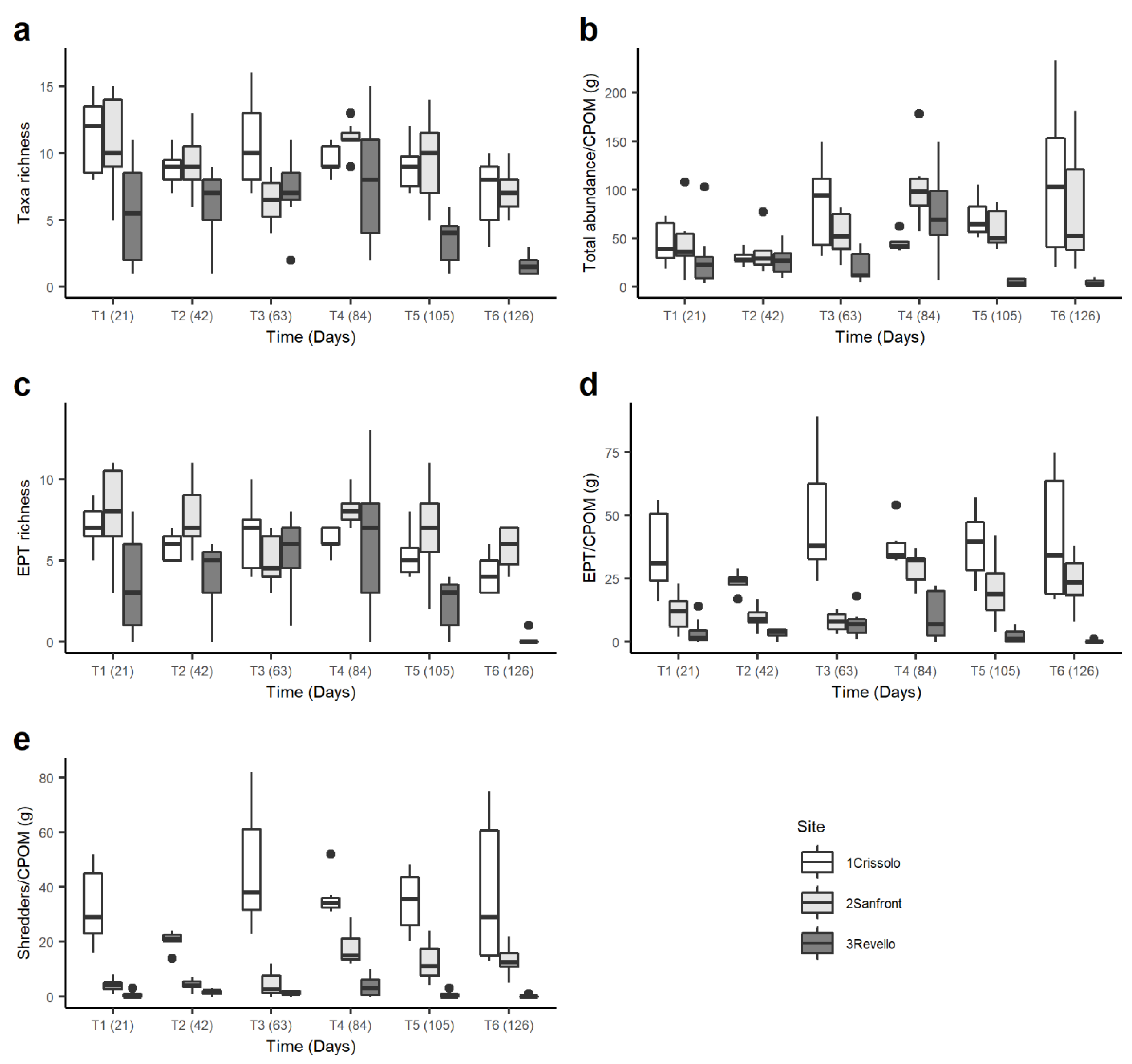
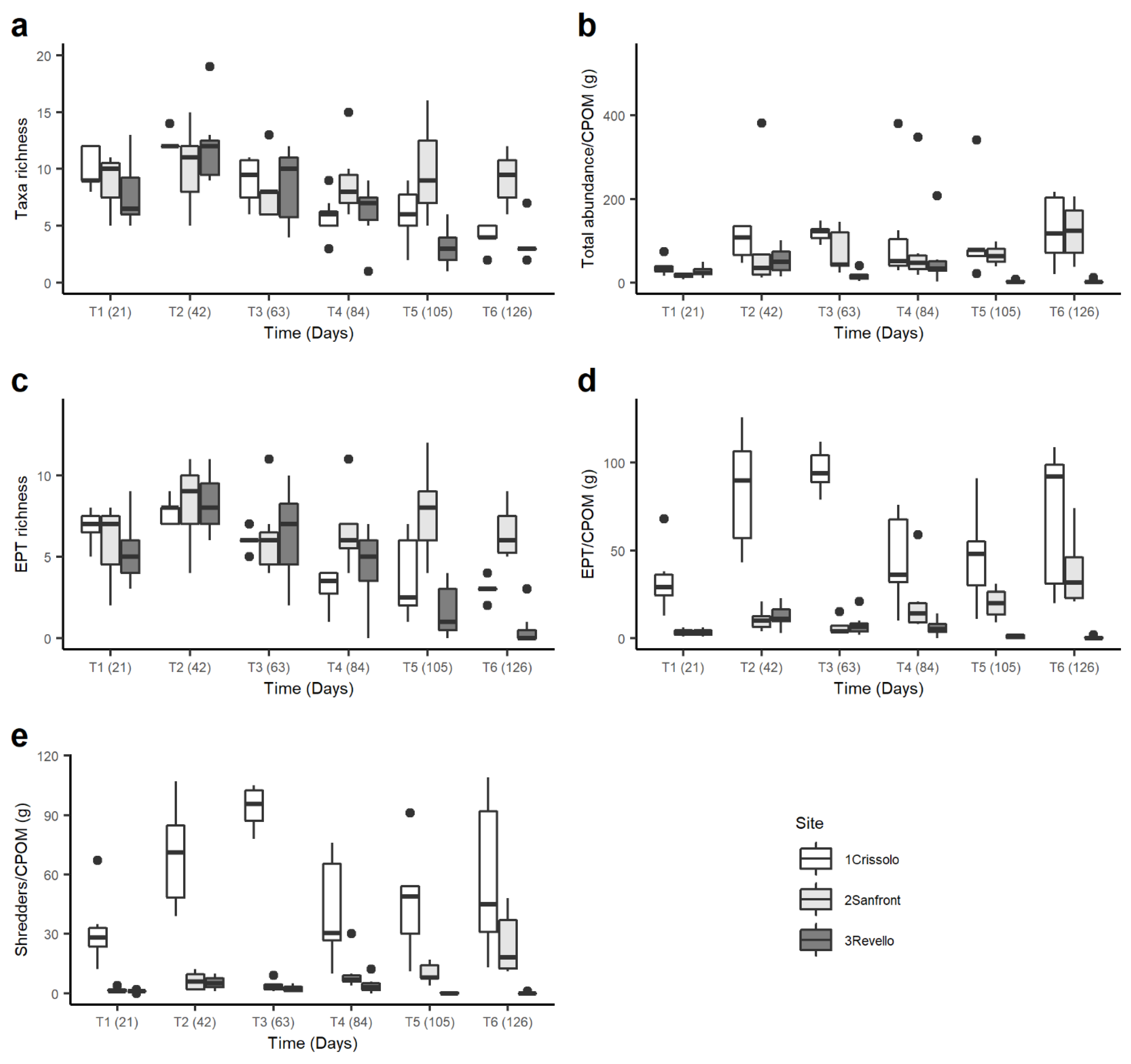
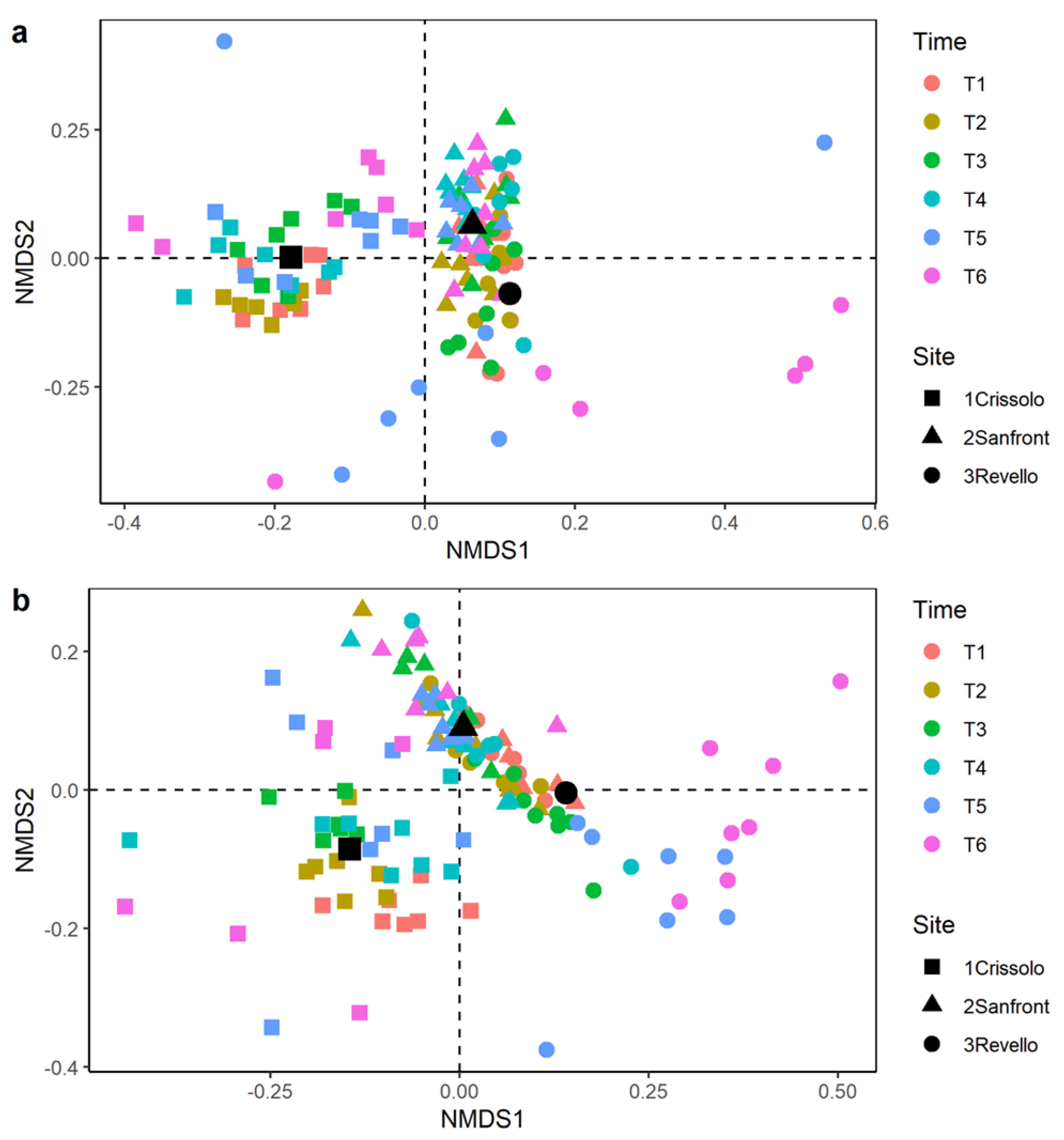
3.3. Macroinvertebrate Community
4. Discussion
4.1. Leaf Litter Decomposition
4.2. Macroinvertebrate Community
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fenoglio, S.; Bo, T.; Cucco, M.; Mercalli, L.; Malacarne, G. Effects of global climate change on freshwater biota: A review with special emphasis on the Italian situation. Ital. J. Zool. 2010, 77, 374–383. [Google Scholar] [CrossRef]
- Lencioni, V. Glacial influence and stream macroinvertebrate biodiversity under climate change: Lessons from the Southern Alps. Sci. Total Environ. 2018, 622, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Rogora, M.; Frate, L.; Carranza, M.L.; Freppaz, M.; Stanisci, A.; Bertani, I.; Bottarin, R.; Brambilla, A.; Canullo, R.; Carbognani, M.; et al. Assessment of climate change effects on mountain ecosystems through a cross-site analysis in the Alps and Apennines. Sci. Total Environ. 2018, 624, 1429–1442. [Google Scholar] [CrossRef] [Green Version]
- Zolezzi, G.; Bellin, A.; Bruno, M.C.; Maiolini, B.; Siviglia, A. Assessing hydrological alterations at multiple temporal scales: Adige River, Italy. Water Resour. Res. 2009, 45, 12. [Google Scholar] [CrossRef]
- Ravazzani, G.; Barbero, S.; Salandin, A.; Senatore, A.; Mancini, M. An integrated hydrological model for assessing climate change impacts on water resources of the upper Po river basin. Water Resour. Manag. 2015, 29, 1193–1215. [Google Scholar] [CrossRef]
- Monroy, S.; Martínez, A.; López-Rojo, N.; Pérez-Calpe, A.V.; Basaguren, A.; Pozo, J. Structural and functional recovery of macroinvertebrate communities and leaf litter decomposition after a marked drought: Does vegetation type matter? Sci. Total Environ. 2017, 599, 1241–1250. [Google Scholar] [CrossRef] [PubMed]
- Gessner, M.O.; Chauvet, E. A case for using litter breakdown to assess functional stream integrity. Ecol. Appl. 2002, 12, 498–510. [Google Scholar] [CrossRef]
- Young, R.G.; Matthaei, C.D.; Townsend, C.R. Organic matter breakdown and ecosystem metabolism: Functional indicators for assessing river ecosystem health. J. N. Am. Benthol. Soc. 2008, 27, 605–625. [Google Scholar] [CrossRef]
- Ledger, M.E.; Brown, L.E.; Edwards, F.K.; Milner, A.M.; Woodward, G. Drought alters the structure and functioning of complex food webs. Nat. Clim. Chang. 2013, 3, 223–227. [Google Scholar] [CrossRef]
- Cummins, K.W. Structure and function of stream ecosystems. BioScience 1974, 24, 631–641. [Google Scholar] [CrossRef]
- Leff, L.G. Microbial Ecology of Streams. Encycl. Water Sci. Technol. Soc. 2019, 1–13. [Google Scholar] [CrossRef]
- Allan, J.D.; Castillo, M.M.; Capps, K.A. Stream Ecology: Structure and Function of Running Waters; Springer Nature: Cham, Switzerland, 2020. [Google Scholar]
- Zhang, M.; Cheng, X.; Geng, Q.; Shi, Z.; Luo, Y.; Xu, X. Leaf litter traits predominantly control litter decomposition in streams worldwide. Glob. Ecol. Biogeogr. 2019, 28, 1469–1486. [Google Scholar] [CrossRef]
- Bastias, E.; Ribot, M.; Romaní, A.M.; Mora-Gómez, J.; Sabater, F.; López, P.; Martí, E. Responses of microbially driven leaf litter decomposition to stream nutrients depend on litter quality. Hydrobiologia 2018, 806, 333–346. [Google Scholar] [CrossRef]
- Santschi, F.; Gounand, I.; Harvey, E.; Altermatt, F. Leaf litter diversity and structure of microbial decomposer communities modulate litter decomposition in aquatic systems. Funct. Ecol. 2018, 32, 522–532. [Google Scholar] [CrossRef] [Green Version]
- Tank, J.L.; Rosi-Marshall, E.J.; Griffiths, N.A.; Entrekin, S.A.; Stephen, M.L. A review of allochthonous organic matter dynamics and metabolism in streams. J. N. Amer. Benthol. Soc. 2010, 29, 118–146. [Google Scholar] [CrossRef] [Green Version]
- Follstad Shah, J.J.; Kominoski, J.S.; Ardón, M.; Dodds, W.K.; Gessner, M.O.; Griffiths, N.A.; Hawkins, C.P.; Johnson, S.L.; Lecerf, A.; LeRoy, C.J.; et al. Global synthesis of the temperature sensitivity of leaf litter breakdown in streams and rivers. Glob. Chang. Biol. 2017, 23, 3064–3075. [Google Scholar] [CrossRef] [Green Version]
- Leigh, C.; Bonada, N.; Boulton, A.J.; Hugueny, B.; Larned, S.T.; Vander Vorste, R.; Datry, T. Invertebrate assemblage responses and the dual roles of resistance and resilience to drying in intermittent rivers. Aquat. Sci. 2016, 78, 291–301. [Google Scholar] [CrossRef] [Green Version]
- Chessman, B.C. Dissolved-oxygen, current and temperature preferences of stream invertebrates estimated from field distributions: Application to assemblage responses to drought. Hydrobiologia 2018, 809, 141–153. [Google Scholar] [CrossRef]
- Doretto, A.; Piano, E.; Falasco, E.; Fenoglio, S.; Bruno, M.C.; Bona, F. Investigating the role of refuges and drift on the resilience of macroinvertebrate communities to drying conditions: An experiment in artificial streams. River Res. Appl. 2018, 34, 777–785. [Google Scholar] [CrossRef]
- Piano, E.; Doretto, A.; Falasco, E.; Fenoglio, S.; Gruppuso, L.; Nizzoli, D.; Viaroli, P.; Bona, F. If Alpine streams run dry: The drought memory of benthic communities. Aquat. Sci. 2019, 81, 32. [Google Scholar] [CrossRef]
- Tonin, A.M.; Hepp, L.U.; Restello, R.M.; Gonçalves, J.F. Understanding of colonization and breakdown of leaves by invertebrates in a tropical stream is enhanced by using biomass as well as count data. Hydrobiologia 2014, 740, 79–88. [Google Scholar] [CrossRef]
- Datry, T.; Corti, R.; Claret, C.; Philippe, M. Flow intermittence controls leaf litter breakdown in a French temporary alluvial river: The “drying memory”. Aquat. Sci. 2011, 73, 471–483. [Google Scholar] [CrossRef]
- Ledger, M.E.; Edwards, F.K.; Brown, L.E.; Milner, A.M.; Woodward, G.U.Y. Impact of simulated drought on ecosystem biomass production: An experimental test in stream mesocosms. Glob. Chang. Biol. 2011, 17, 2288–2297. [Google Scholar] [CrossRef]
- Gonçalves, A.L.; Lírio, A.V.; Graça, M.A.; Canhoto, C. Fungal species diversity affects leaf decomposition after drought. Int. Rev. Hydrobiol. 2016, 101, 78–86. [Google Scholar] [CrossRef]
- Gionchetta, G.; Oliva, F.; Menéndez, M.; Lopez Laseras, P.; Romaní, A.M. Key role of streambed moisture and flash storms for microbial resistance and resilience to long-term drought. Freshw. Biol. 2019, 64, 306–322. [Google Scholar] [CrossRef]
- Gionchetta, G.; Romaní, A.M.; Oliva, F.; Artigas, J. Distinct responses from bacterial, archaeal and fungal streambed communities to severe hydrological disturbances. Sci. Rep. 2019, 9, 13506. [Google Scholar] [CrossRef]
- Arias-Real, R.; Gutiérrez-Cánovas, C.; Munoz, I.; Pascoal, C.; Menéndez, M. Fungal biodiversity mediates the effects of drying on freshwater ecosystem functioning. Ecosystems 2021, 24, 1–15. [Google Scholar] [CrossRef]
- Fenoglio, S.; Bo, T.; Cammarata, M.; López-Rodríguez, M.J.; Tierno de Figueroa, J.M. Seasonal variation of allochthonous and autochthonous energy inputs in an alpine stream. J. Limnol. 2015, 74, 272–277. [Google Scholar] [CrossRef] [Green Version]
- Calapez, A.R.; Elias, C.L.; Almeida, S.F.; Feio, M.J. Extreme drought effects and recovery patterns in the benthic communities of temperate streams. Limnetica 2014, 33, 281–296. [Google Scholar] [CrossRef]
- Elias, C.L.; Calapez, A.R.; Almeida, S.F.; Feio, M.J. From perennial to temporary streams: An extreme drought as a driving force of freshwater communities’ traits. Mar. Freshw. Res. 2015, 66, 469–480. [Google Scholar] [CrossRef]
- Falasco, E.; Piano, E.; Doretto, A.; Fenoglio, S.; Bona, F. Lentification in Alpine rivers: Patterns of diatom assemblages and functional traits. Aquat. Sci. 2018, 80, 36. [Google Scholar] [CrossRef]
- Piano, E.; Doretto, A.; Falasco, E.; Gruppuso, L.; Fenoglio, S.; Bona, F. The role of recurrent dewatering events in shaping ecological niches of scrapers in intermittent Alpine streams. Hydrobiologia 2019, 841, 177–189. [Google Scholar] [CrossRef]
- ARPA Agenzia Regionale per la Protezione dell’Ambiente. Idrologia in Piemonte nel 2012; Regione: Piemonte, Italy, 2013; p. 23. Available online: http://www.arpa.piemonte.it (accessed on 3 November 2021).
- Campaioli, S.; Ghetti, P.F.; Minelli, A.; Ruffo, S. Manuale per il Riconoscimento dei Macroinvertebrati Delle Acque Dolci Italiane; Provincia Autonoma di Trento: Trento, Italy, 1994; Volume I. [Google Scholar]
- Campaioli, S.; Ghetti, P.F.; Minelli, A.; Ruffo, S. Manuale per il Riconoscimento dei Macroinvertebrati Delle Acque Dolci Italiane; Provincia Autonoma di Trento: Trento, Italy, 1999; Volume II. [Google Scholar]
- Sansoni, G. Atlante per il Riconoscimento dei Macroinvertebrati dei Corsi D’acqua Italiani; Provincia Autonoma di Trento: Trento, Italy, 1988. [Google Scholar]
- Tachet, H.; Bournaud, M.; Richoux, P. Introduction à L’etude des Macroinvertebres des Eaux Douces, 2nd ed.; Villeurbanne, Universiteé de Lyon I: Lyon, France, 1984. [Google Scholar]
- Usseglio-Polatera, P.; Bournaud, M.; Richoux, P.; Tachet, H. Biological and ecological traits of benthic freshwater macroinvertebrates: Relationships and definition of groups with similar traits. Freshw. Biol. 2000, 43, 175–205. [Google Scholar] [CrossRef]
- Zuur, A.F.; Ieno, E.N.; Elphick, C.S. A protocol for data exploration to avoid common statistical problems. Methods Ecol. Evol. 2010, 1, 3–14. [Google Scholar] [CrossRef]
- Benfield, E.F.; Fritz, K.M.; Tiegs, S.D. Leaf-litter breakdown. In Methods in Stream Ecology; Lamberti, G.A., Hauer, F.R., Eds.; Elsevier Academic Press: Cambridge, MA, USA, 2017. [Google Scholar] [CrossRef]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Anderson, M.J. Distance-based tests for homogeneity of multivariate dispersions. Biometrics 2006, 62, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Piano, E.; Giuliano, D.; Isaia, M. Islands in cities: Urbanization and fragmentation drive taxonomic and functional variation in ground arthropods. Basic Appl. Ecol. 2020, 43, 86–98. [Google Scholar] [CrossRef] [Green Version]
- Dufrêne, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- R Core Development Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 4 October 2021).
- Bates, D.; Machler, M.; Bolker, B. Lme4: Linear Mixed-Effects Models Using S4 Classes, R package version 0.999375–42; 2011. Available online: http://CRAN.R-project.org/package=lme4 (accessed on 4 October 2021).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Wagner, H. Vegan: Community Ecology Package, R Package Version 2.2-1; 2015. Available online: https://cran.rproject.org/package=vegan (accessed on 4 October 2021).
- Cáceres, M.D.; Legendre, P. Associations between species and groups of sites: Indices and statistical inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer Nature: New York, NY, USA, 2016. [Google Scholar] [CrossRef]
- Kassambara, A. ggpubr: “ggplot2” Based Publication Ready Plots. R Package Version 0.1, 6. 2017. Available online: https://cran.r-project.org/package=ggpubr (accessed on 4 October 2021).
- Northington, R.M.; Webster, J.R. Experimental reductions in stream flow alter litter processing and consumer subsidies in headwater streams. Freshw. Biol. 2017, 62, 737–750. [Google Scholar] [CrossRef]
- Duarte, S.; Mora-Gómez, J.; Romaní, A.M.; Cássio, F.; Pascoal, C. Responses of microbial decomposers to drought in streams may depend on the environmental context. Environ. Microbiol. Rep. 2017, 9, 756–765. [Google Scholar] [CrossRef] [PubMed]
- Mas-Martí, E.; Sanpera-Calbet, I.; Muñoz, I. Bottom-up effects of streambed drying on consumer performance through changes in resource quality. Aquat. Sci. 2017, 79, 719–731. [Google Scholar] [CrossRef] [Green Version]
- Mora-Gómez, J.; Duarte, S.; Cássio, F.; Pascoal, C.; Romaní, A.M. Microbial decomposition is highly sensitive to leaf litter emersion in a permanent temperate stream. Sci. Total Environ. 2018, 621, 486–496. [Google Scholar] [CrossRef] [PubMed]
- Simões, S.; Canhoto, C.; Bärlocher, F.; Gonçalves, A.L. Hydrological contraction patterns and duration of drying period shape microbial-mediated litter decomposition. Sci. Total Environ. 2021, 785, 147312. [Google Scholar] [CrossRef] [PubMed]
- Santonja, M.; Pellan, L.; Piscart, C. Macroinvertebrate identity mediates the effects of litter quality and microbial conditioning on leaf litter recycling in temperate streams. Ecol. Evol. 2018, 8, 2542–2553. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, V.; Chauvet, E.; Canhoto, C. Effects of experimental warming, litter species, and presence of macroinvertebrates on litter decomposition and associated decomposers in a temperate mountain stream. Can. J. Fish. Aquat. Sci. 2015, 72, 206–216. [Google Scholar] [CrossRef] [Green Version]
- Júnior, E.S.A.; Martinez, A.; Gonçalves, A.L.; Canhoto, C. Combined effects of freshwater salinization and leaf traits on litter decomposition. Hydrobiologia 2020, 847, 3427–3435. [Google Scholar] [CrossRef]
- López, E.S.; Pardo, I.; Felpeto, N. Seasonal differences in green leaf breakdown and nutrient content of deciduous and evergreen tree species and grass in a granitic headwater stream. Hydrobiologia 2001, 464, 51–61. [Google Scholar] [CrossRef]
- Villanueva, V.D.; Albariño, R.; Canhoto, C. Detritivores feeding on poor quality food are more sensitive to increased temperatures. Hydrobiologia 2011, 678, 155–165. [Google Scholar] [CrossRef]
- Del Campo, R.; Corti, R.; Singer, G. Flow intermittence alters carbon processing in rivers through chemical diversification of leaf litter. Limnol. Oceanogr. Lett. 2020, 6, 232–242. [Google Scholar] [CrossRef]
- Di Sabatino, A.; Coscieme, L.; Cristiano, G. Effects of antecedent drying events on structure, composition and functional traits of invertebrate assemblages and leaf-litter breakdown in a former perennial river of Central Apennines (Aterno River, Abruzzo, Central Italy). Ecohydrology 2021, 14, e2358. [Google Scholar] [CrossRef]
- Piano, E.; Doretto, A.; Mammola, S.; Falasco, E.; Fenoglio, S.; Bona, F. Taxonomic and functional homogenisation of macroinvertebrate communities in recently intermittent Alpine watercourses. Freshw. Biol. 2020, 65, 2096–2107. [Google Scholar] [CrossRef]
- Doretto, A.; Bona, F.; Falasco, E.; Morandini, D.; Piano, E.; Fenoglio, S. Stay with the flow: How macroinvertebrate communities recover during the rewetting phase in Alpine streams affected by an exceptional drought. River Res. Appl. 2020, 36, 91–101. [Google Scholar] [CrossRef] [Green Version]
- Stubbington, R.; Bogan, M.T.; Bonada, N.; Boulton, A.J.; Datry, T.; Leigh, C.; Vander Vorste, R. The biota of intermittent rivers and ephemeral streams: Aquatic invertebrates. In Intermittent Rivers and Ephemeral Streams; Academic Press: Cambridge, MA, USA, 2017; pp. 217–243. [Google Scholar]
- Gruppuso, L.; Doretto, A.; Piano, E.; Falasco, E.; Bruno, M.C.; Bona, F.; Fenoglio, S. Effects of flow intermittence on ecosystem processes in mountain streams: Are artificial and field experiments comparable? Fundam. Appl. Limnol. 2021, 195, 39–59. [Google Scholar] [CrossRef]
- Datry, T.; Larned, S.T.; Fritz, K.M.; Bogan, M.T.; Wood, P.J.; Meyer, E.I.; Santos, A.N. Broad-scale patterns of invertebrate richness and community composition in temporary rivers: Effects of flow intermittence. Ecography 2014, 37, 94–104. [Google Scholar] [CrossRef] [Green Version]
- Herbst, D.B.; Cooper, S.D.; Medhurst, R.B.; Wiseman, S.W.; Hunsaker, C.T. Drought ecohydrology alters the structure and function of benthic invertebrate communities in mountain streams. Freshw. Biol. 2019, 64, 886–902. [Google Scholar] [CrossRef]
- Piano, E.; Doretto, A.; Falasco, E.; Gruppuso, L.; Bona, F.; Fenoglio, S. Flow intermittency negatively affects three phylogenetically related shredder stoneflies by reducing CPOM availability in recently intermittent Alpine streams in SW-Italian Alps. Hydrobiologia 2020, 847, 4049–4061. [Google Scholar] [CrossRef]
- Acuña, V.; Muñoz, I.; Giorgi, A.; Omella, M.; Sabater, F.; Sabater, S. Drought and postdrought recovery cycles in an intermittent Mediterranean stream: Structural and functional aspects. J. N. Am. Benthol. Soc. 2005, 24, 919–933. [Google Scholar] [CrossRef]
- Ylla, I.; Sanpera-Calbet, I.; Vázquez, E.; Romaní, A.M.; Munoz, I.; Butturini, A.; Sabater, S. Organic matter availability during pre-and post-drought periods in a Mediterranean stream. In Global Change and River Ecosystems—Implications for Structure, Function and Ecosystem Services; Springer Nature: Cham, Switzerland, 2010; pp. 217–232. [Google Scholar]
- Vicente-Serrano, S.M.; Lopez-Moreno, J.I.; Beguería, S.; Lorenzo-Lacruz, J.; Sanchez-Lorenzo, A.; García-Ruiz, J.M.; Espejo, F. Evidence of increasing drought severity caused by temperature rise in southern Europe. Environ. Res. Lett. 2014, 9, 044001. [Google Scholar] [CrossRef]
- Bruno, M.C.; Doretto, A.; Boano, F.; Ridolfi, L.; Fenoglio, S. Role of the Hyporheic Zone in Increasing the Resilience of Mountain Streams Facing Intermittency. Water 2020, 12, 2034. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Leaf Type | Variable | Estimate | SE | t | p |
|---|---|---|---|---|---|
| Days | −0.02 | 0.002 | −11.46 | <0.001 | |
| Site2Sanfront | −0.36 | 0.10 | −3.55 | <0.001 | |
| Quercus robur | Site3Revello | −0.61 | 0.10 | −6.02 | <0.001 |
| Days:Site2Sanfront | 0.01 | 0.00 | 9.85 | <0.001 | |
| Days:Site3Revello | 0.02 | 0.00 | 15.33 | <0.001 | |
| Days | −0.04 | 0.00 | −18.40 | <0.001 | |
| Site2Sanfront | −0.34 | 0.19 | −1.74 | 0.08 | |
| Castanea sativa | Site3Revello | −0.81 | 0.19 | −4.09 | <0.001 |
| Days:Site2Sanfront | 0.02 | 0.00 | 9.81 | <0.001 | |
| Days:Site3Revello | 0.04 | 0.00 | 14.54 | <0.001 |
| Metric | Leaf Type | Int | SE | t | Site | F | p |
|---|---|---|---|---|---|---|---|
| Quercus robur | 27.6303 | 0.5674 | 48.7 | Crissolo | 5.933 | 0.003 | |
| Sanfront | 7.804 | 0.007 | |||||
| C:N | Revello | 9.003 | 0.004 | ||||
| Castanea sativa | 28.9510 | 0.6734 | 42.99 | Crissolo | 32.31 | <0.001 | |
| Sanfront | 14.81 | <0.001 | |||||
| Revello | 33.35 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gruppuso, L.; Doretto, A.; Falasco, E.; Fenoglio, S.; Freppaz, M.; Benbow, M.E.; Bona, F. Flow Intermittency Affects Leaf Decomposition and Benthic Consumer Communities of Alpine Streams: A Case Study along the Po River. Water 2022, 14, 258. https://doi.org/10.3390/w14020258
Gruppuso L, Doretto A, Falasco E, Fenoglio S, Freppaz M, Benbow ME, Bona F. Flow Intermittency Affects Leaf Decomposition and Benthic Consumer Communities of Alpine Streams: A Case Study along the Po River. Water. 2022; 14(2):258. https://doi.org/10.3390/w14020258
Chicago/Turabian StyleGruppuso, Laura, Alberto Doretto, Elisa Falasco, Stefano Fenoglio, Michele Freppaz, Mark Eric Benbow, and Francesca Bona. 2022. "Flow Intermittency Affects Leaf Decomposition and Benthic Consumer Communities of Alpine Streams: A Case Study along the Po River" Water 14, no. 2: 258. https://doi.org/10.3390/w14020258