
Effects of Long-Term High Carbonate Alkalinity Stress on the Ovarian Development in Exopalaemon carinicauda
by

Xiuhong Zhang
1,2,†,
Jiajia Wang
1,2,†,
Chengwei Wang
1,2,
Wenyang Li
1,2,
Qianqian Ge
3,
Zhen Qin
1,2,
Jian Li
1,2 and
Jitao Li
1,2,* 1
Key Laboratory of Sustainable Development of Marine Fisheries, Ministry of Agriculture and Rural Affairs, Yellow Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Qingdao 266071, China
2
Laboratory for Marine Fisheries Science and Food Production Processes, Pilot National Laboratory for Marine Science and Technology, Qingdao 266071, China
3
Pilot National Laboratory for Marine Science and Technology, Qingdao 266237, China
*
Author to whom correspondence should be addressed.
†
These authors have contributed equally to this work.
Water 2022, 14(22), 3690; https://doi.org/10.3390/w14223690
Submission received: 17 October 2022
/
Revised: 11 November 2022
/
Accepted: 12 November 2022
/
Published: 15 November 2022
(This article belongs to the Special Issue Advances in Aquaculture Ecology Research)
Abstract
:Saline–alkaline water limits the growth and survival of aquatic animals due to its high carbonate alkalinity, high pH, and various ion imbalances. The ridgetail white prawn Exopalaemon carinicauda is strongly adaptable to the saline–alkaline water, making it an excellent candidate species for large-scale aquaculture in saline–alkaline areas. To explore the effect of long-term high carbonate alkalinity stress on ovarian development in E. carinicauda for assisting the development of saline–alkaline aquaculture, we performed ovary histology analysis and RNA–sequencing of the eyestalk and ovary in order to compare the transcriptomic responses of individuals in high carbonate alkalinity (8 mmol/L) with a control group (2 mmol/L) for 60 days. It was found that high carbonate alkalinity stress resulted in a loose arrangement of oogonia and a small number of surrounding follicular cells. A total of 1102 differentially expressed genes (DEGs) in ovary tissue were identified under high carbonate alkalinity stress, and the 18 important DEGs were associated with ovarian development. The majority of the DEGs were enriched in ECM–receptor interaction, Folate biosynthesis, the FoxO signaling pathway, insect hormone biosynthesis, and lysosome, which were involved in the ovarian development of E. carinicauda. A total of 468 DEGs were identified in eyestalk tissue under high carbonate alkalinity stress, and the 13 important DEGs were associated with ovarian development. KEGG enrichment analysis found that ECM–receptor interaction, folate biosynthesis, lysosome, metabolic pathways, and retinol metabolism may be involved in the ovarian development under high carbonate alkalinity stress. Our results provide new insights and reveal the genes and pathways involved in the ovarian development of E. carinicauda under long-term high carbonate alkalinity stress.
1. Introduction
Saline–alkaline water, as the third kind of water, differs from seawater and freshwater and accounts for a large proportion of the world’s water resources [1,2]. There are about 46 million hectares of saline–alkaline water areas, which are widely distributed in the northwest, northeast, and north of China, involving 19 provinces, cities, and autonomous regions [3,4,5]. Saline–alkaline water has a poor buffering capacity, which is characterized by high pH, high carbonate alkalinity, and various types of ion imbalances [1]. Excessive carbonate alkalinity in water substantially affects the development, survival, and reproduction of organisms. Several fishes can adapt to high alkalinity, such as Leuciscus waleckii [6], Gymnocypris przewalskii [7], and tilapia [8], which provide excellent breeding organisms for the development and utilization of carbonate alkalinity waters for aquaculture.
The ridgetail white prawn E. carinicauda is one of the most important commercial shrimps [9], which is widely distributed in the coastal areas of the Yellow Sea and the Bohai Sea in China [10]. Due to its multiple advantages, such as fast growth, high reproductive performance, and good disease resistance [11], the breeding area of E. carinicauda in China has expanded in recent years, which may account for one third of the total output of mixed culture ponds [12,13]. Recently in China, E. carinicauda has been successfully bred and cultured in saline–alkaline ponds at Dongying city, Shandong province (with salinity of 5–8 and carbonate alkalinity of 1.4–8.0 mmol/L) and Cangzhou city, Hebei province (with salinity of 10–20 and carbonate alkalinity of 3.5–13.0 mmol/L), suggesting that E. carinicauda have a high tolerance to saline–alkaline stress [13,14]. E. carinicauda shows strong tolerance to salinity and high carbonate alkalinity, and can carry eggs during culture. Therefore, it is a potential species suitable for large-scale culture in saline–alkaline waters. However, as most studies focused on changes in the physiology and gill transcriptomics in E. carinicauda, the effects of long-term carbonate alkalinity stress on the reproductive mechanisms in E. carinicauda still remains unknown.
Carbonate alkalinity has been considered to be the main stress source affecting the survival, growth, and reproductive traits of aquatic animals in saline–alkaline water [15,16,17]. High carbonate alkalinity can alter normal metabolism, osmotic pressure regulation capacity, and antioxidant capacity [18]. Yao et al. [3] reported that when the carbonate alkalinity exceeded 15.7 mmol/L, the survival rate of medaka (Oryzias latipes) decreased, and morphological abnormalities such as embryo coagulation, embryonic development stagnation, and hatching failure were present. Previous studies have shown that the growth, development, and reproduction indexes in Moina mongolica Daday are optimal when the alkalinity is 2.05–4.58 mmol/L. However, when the alkalinity is 6.43–8.98 mmol/L, all indicators showed a downward trend [19]. Xu et al. [20] found that survival rate was not affected by water with a salinity < 3.2 and alkalinity < 14.32 mmol/L in a 72 h embryo tolerance experiment with Barbus capito. Water with salinity < 5.1 and alkalinity < 14.32 mmol/L did not affect the 96 h survival of larvae. Liu et al. [21] also found the growth and reproduction of E. carinicauda was not affected by low carbonate alkalinity. E. carinicauda is better adapted to environments with high carbonate alkalinity, as it adjusts immune enzyme activities. However, research on the influence of carbonate alkalinity in aquatic organisms mainly focuses on survival and growth. Few reports address carbonate alkalinity influences on the mechanisms involved in gonadal development. Therefore, we urgently need to understand the reproduction of aquatic organisms in saline–alkaline waters to aid aquaculture development, especially breeding in saline–alkaline water.
In recent years, transcriptome sequencing technology has been widely used to study the differential expression and molecular pathways of genes under specific environmental stresses [22,23]. For example, RNA–seq compares the transcriptomic responses of Litopenaeus vannamei under salinity stress [24]. Li et al. [25] used transcriptome sequencing to reveal the genes and pathways related to salt stress in Eriocheir sinensis. However, research on the effect of carbonate alkalinity on aquatic organisms focuses on osmoregulation, with relatively few studies involved in ovarian development. This is the first study involving the transcriptome of the E. carinicauda ovary and eyestalk being sequenced by RNA–seq technology. We analyzed the transcriptome data of the eyestalks and ovaries under high carbonate alkalinity stress in order to identify the genes and pathways involved in ovarian development. This study helps to clarify the ovarian development mechanisms in E. carinicauda when adapting to high carbonate alkalinity.
2. Materials and Methods
2.1. Sample Collection
Adult female shrimps (body length 2.67 ± 0.20 cm, body weight 0.12 ± 0.08 g) were collected from Haichen Aquaculture Co., Ltd., in Rizhao, Shandong province, China. The shrimps were domesticated in the environment (25 °C) for two weeks. The experiment was performed in 200 L PVC tanks. One hundred and eighty shrimps were divided into two groups randomly, including the high carbonate alkalinity group (carbonate alkalinity 8 ± 0.5 mmol/L, salinity 25 ppt, temperature 26 ± 0.5 °C, pH 8.1 ± 0.3, dissolved oxygen 7.5 ± 0.5 mg L−1) and the control group (carbonate alkalinity 2 ± 0.5 mmol/L, salinity 25 ppt, temperature 26 ± 0.5 °C, pH 8.8 ± 0.5, dissolved oxygen 7.5 ± 0.5 mg L−1). The experimental design included three replicates of 30 shrimps in each group. The shrimp were fed 3–5% of their body weight twice daily (8:00 and 18:00) during the experimental period. The water was aerated and 30% of the water was changed daily (with the new sea-water adjusted to maintain the original carbonate alkalinity).
After 60 days, thirty–six female shrimps (6 individuals × 3 three replicates × 2 groups) were randomly sampled for Illumina (San Diego, CA, USA) RNA–seq. The ovary and eyestalk samples were obtained and rapidly frozen in liquid nitrogen, then stored at −80 °C until RNA isolation, respectively. The ovary samples in the high carbonate alkalinity group are labeled HC_O and the eyestalk samples are labeled HC_E. The ovary samples in the control group are labeled CG_O and the eyestalk samples are labeled CG_E. Six female shrimps (2 individuals × 3 three replicates × 2 groups) were sacrificed for the tissue slices.
2.2. Ovary Histology
The ovaries were fixed in 4% paraformaldehyde for 24 h before washing with 1 × PBS and dehydrated using a graded ethanol series (80% ethanol for 1 h, followed by 95% ethanol for 1 h, followed by 100% ethanol for 1 h). Transparency was improved using xylene (pure ethanol: xylene (1:1) for 1 h, then xylene for 1 h). Samples were infiltrated with paraffin (xylene: paraffin (1:1) at 62 °C for 1 h, then paraffin at 62 °C for 2 h) and processed for paraffin embedding. Sections were cut to 6 µm before staining with hematoxylin and eosin. Samples were scanned using a microscope slide scanner (Pannoramic MIDI, 3DHISTECH Ltd., Budapest, Hungary).
2.3. RNA Isolation, Library Construction and Illumine Sequencing
TRIzol® reagent (Invitrogen, San Diego, CA, USA) extracted the total RNA by following the manufacturer’s instructions. The total RNA was treated with DNase I to implement DNA digestion and obtain pure RNA products. Finally, the RNA purity was examined using NanoPhotometer® spectrophotometer NanoDrop 2000 (IMPLEN, Westlake Village, CA, USA). The RNA integrity and concentration was determined using Agilent 2100/4200 system software (Agilent Technologies, Santa Clara, CA, USA). Next, equal amounts of RNA from the same group of different individuals were pooled in order to construct the library. Next–generation sequencing library preparations were constructed according to the manufacturer’s instructions (NEBNext® Ultra™ RNA Library Prep Kit for Illumina® (NEB, Ipswich, MA, USA)). After the mRNA library successfully passed the quality inspection, PE150 sequencing was performed using the Illumina NovaSeq 6000 platform (Thermo, Waltham, MA, USA).
2.4. Basic Analysis of Sequencing Data
We removed any technical sequences or their fragments (including adapters, polymerase chain reaction (PCR) primers), and any low quality sequences (with a base quality < 20). We achieved this by filtering the data (in the fastq format) using Trimmomatic (v0.30) to provide only high–quality, clean data in our analysis. Firstly, the whole genome sequence of E. carinicauda assembled by our research group was taken as the reference genome (the data have not been published yet). Next, Hisat2 (v2.0.1) was indexed to the reference genome sequence. Finally, we mapped the clean reads to the Silva database to remove the rRNA. All subsequent date analyses were based on clean data without rRNA.
2.5. Differential Expression Genes (DEGs) Analysis and Enrichment Analysis
The DESeq2 and edgeR [26] methods were used to perform the differential expression analysis. The influence of gene length and sequencing volume on the calculated gene expression level was eliminated by the fragments per kilobase per million reads (FPKM) method. The calculated gene expression level can be directly used to compare the expression variations between different genes. We used FPKM to normalize the data. The Benjamini and Hochberg’s approach controlled the false discovery rate. |log2FC| ≥ 1 and p-values < 0.05 were set to detect any significant DEGs. Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genome (KEGG) enrichment analyses of differentially expressed gene sets were implemented by the topG and KOBAS packages 3.0 [27], respectively.
2.6. RNA-Seq Data Validation by Real–Time Quantitative PCR
Ten DEGs were randomly selected for quantitative reverse transcription–PCR (qRT–PCR) analysis to validate the differential expression of mRNAs. The qRT–PCR assay was performed using SYBR Green PCR Master Mix (Life Technologies, MASS, Waltham, MA, USA) in the 7500 fast Real–Time PCR system (Applied Biosystems, Foster, CA, USA) according to the manufacturer’s agreement. The 18S rRNA of E. carinicauda was used as the internal reference [28]. All primer sequences and 18S rRNA sequences are listed in Table S1. The relative expression of target genes was calculated with 2−∆∆CT methods. One–way ANOVA method and Duncan’s test in the statistical software SPSS 22.0 (SPSS, Chicago, IL, USA) were used for statistical analysis. The results are presented as the mean ± standard error, and differences in gene expression were considered statistically significant at p < 0.05.
3. Results
3.1. Histopathology of Ovary
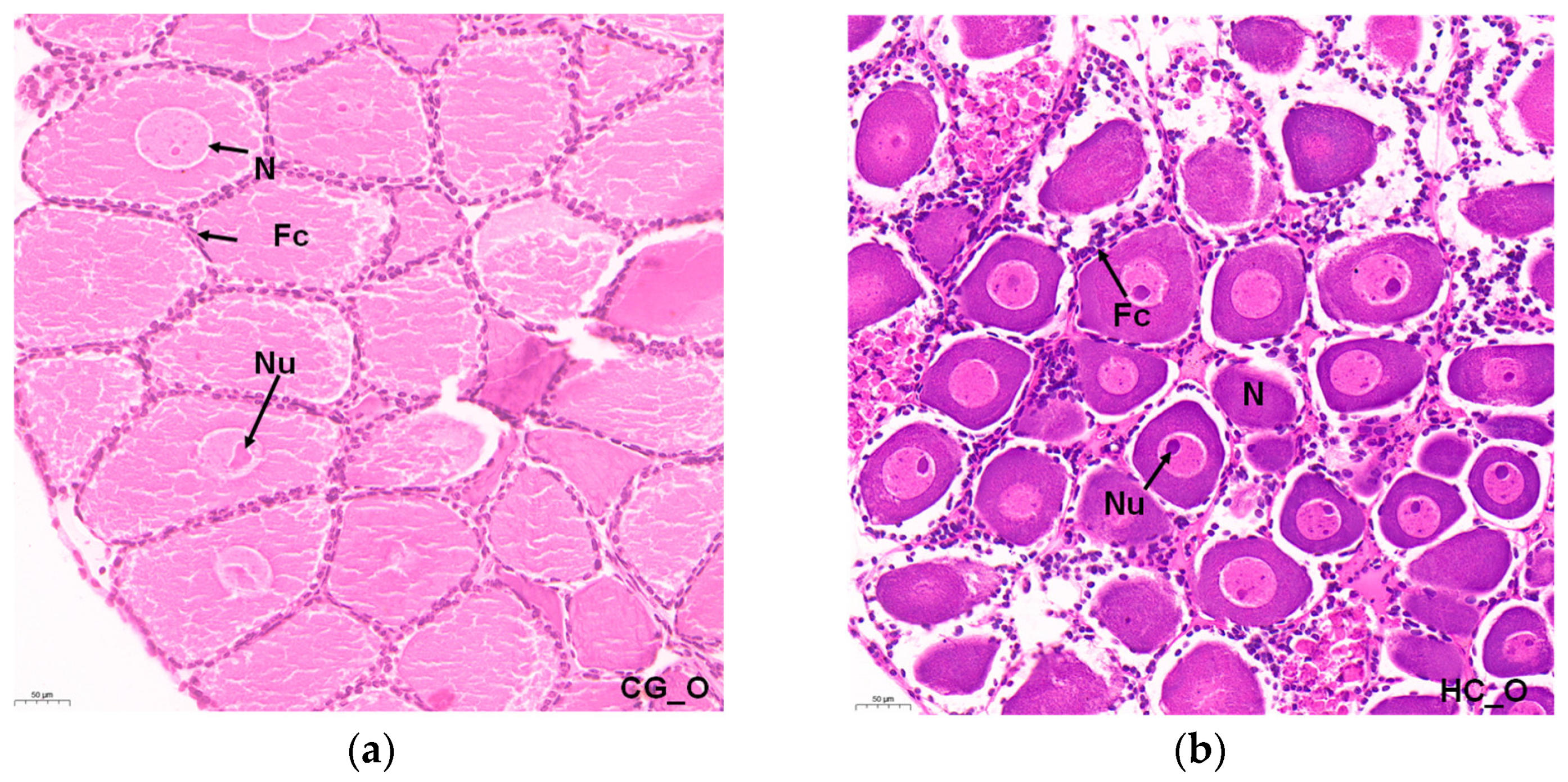
Normal reproduction characteristics were observed in the histological sections of the ovary. The E. carinicauda ovary was in Phase I. The oogonia were oval and proliferative, and the nuclei were round. Most cells had one nucleolus, while a few had two nucleoli. The nucleolus stained the deepest color. The cells of the oogonia were closely arranged. A single layer of follicle cells was closely arranged around the oogonia (Figure 1a). After 60 days of high carbonate alkalinity stress, most of the cells in the ovarian cavity were previtellogenic follicles, and the nuclei were large and original, accounting for about half of the cell volume (Figure 1b).
3.2. Summary of the RNA–Sequencing Data
Six mRNA libraries (n = 6) were constructed from the ovary and eyestalk, respectively, to identify the effects of high carbonate alkalinity on the underlying molecular signaling pathways in E. carinicauda ovarian development. The raw data was submitted to NCBI with accession numbers PRJNA881755 and PRJNA881756. A total of 273,545,646 clean reads were obtained from ovaries, with the Q30 (%) varying from 92.63–94.20%. Of these reads, 70.91–72.74% mapped to the reference genome of E. carinicauda. A total of 264,648,418 clean reads were obtained from eyestalks, with the Q30 (%) varying from 91.72–92.67%. Of these reads, 79.84–83.02% mapped to the reference genome of E. carinicauda (Table S2).
3.3. Differential Gene Expression (DEGs) Analysis
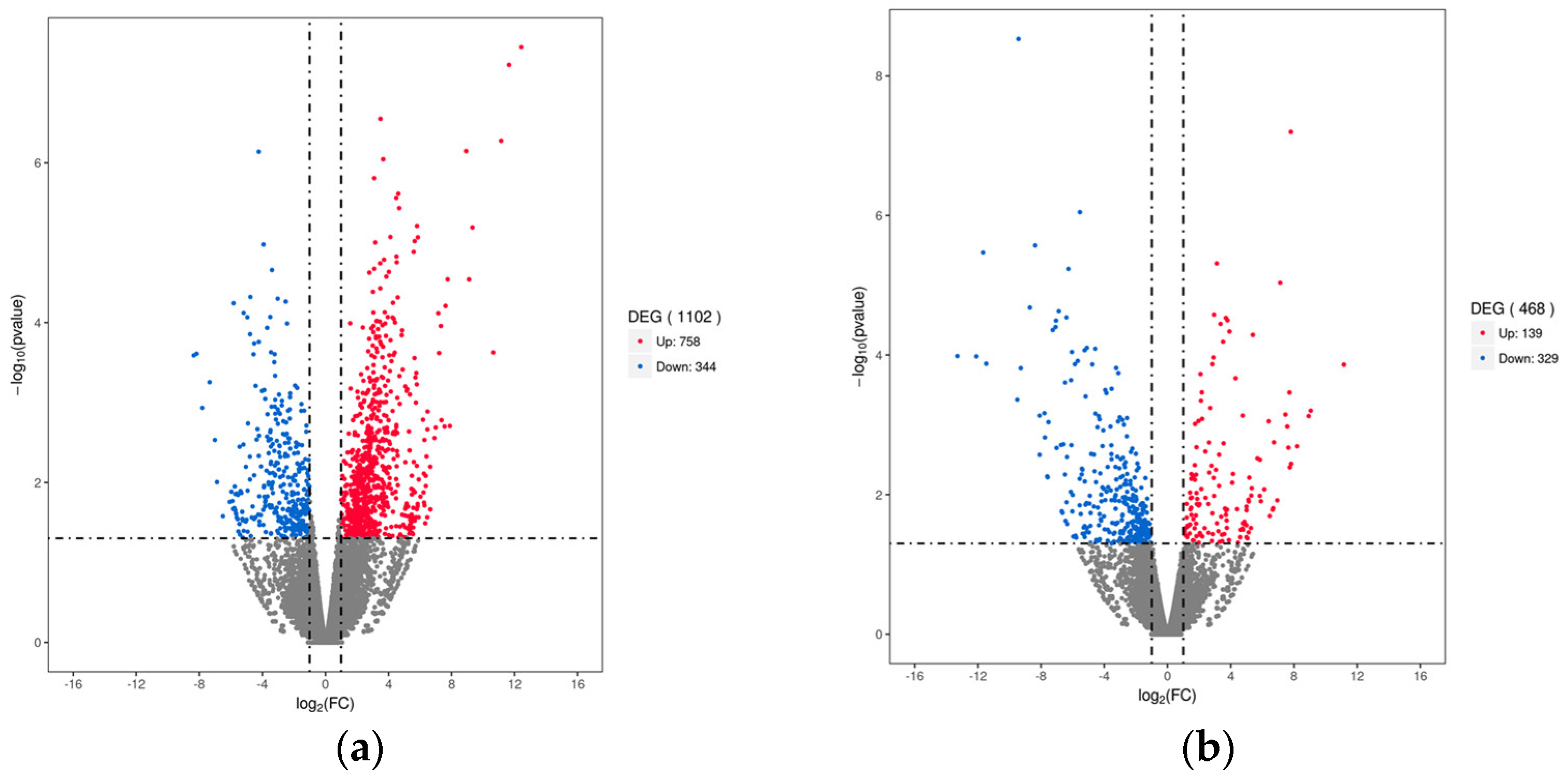
A total of 1102 significant DEGs were identified between the HC_O and CG_O groups. Compared to the control group, 758 up−regulated genes and 344 down−regulated genes were expressed in the ovarian tissue of the HC group (Figure 2a). Compared with the CG_E group, 468 DEG (139 up−regulated and 329 down−regulated) were identified in the HC_E group (Figure 2b). These results suggest that carbonate alkalinity has a significant effect on transcription in the ovary and eyestalk.
3.4. Gene Ontology (GO)Analysis of Significant DEGs
The DEGs of the comparison of HC_O and CG_O were classified into biological process (BP), cellular component (CC), and molecular function (MF). Compared with the CG_O group, up−regulated genes in the HC_O group had enriched steroid metabolic process, sterol homeostasis, molting cycle, follicle cell microvillus organization, steroid biosynthetic process, sterol transport, and estrogen secretion. Conversely, the down−regulated genes in the HC_O group had an enriched positive regulation of ovulation, regulation of ovulation, sterol transport, ovulation, and sterol homeostasis (Table 1).
The DEGs of the comparison of HC_E and CG_E were also classified into biological process (BP), cellular component (CC), and molecular function (MF). Compared with the CG_E group, the up−regulated genes in the HC_E group were enriched in the positive regulation of sterol transport, regulation of sterol transport, embryonic morphogenesis, embryonic organ morphogenesis, embryo development, entry into reproductive diapause, and sterol metabolic process. The down−regulated genes in the HC_E group were enriched in sterol transport, regulation of metaphase/anaphase transition of meiosis I, ecdysteroid metabolic process, sterol homeostasis, maternal placenta development, metaphase/anaphase transition of meiosis I, and the molting cycle (Table 2).
3.5. Kyoto Encyclopedia of Genes and Genomes (KEGG) Analysis
Comparing the DEGs to the KEGG database of pathway enrichment, the potential functions of the significant DEGs were analyzed in order to further understand the ovarian development of E. carinicauda under high carbonate alkalinity stress.
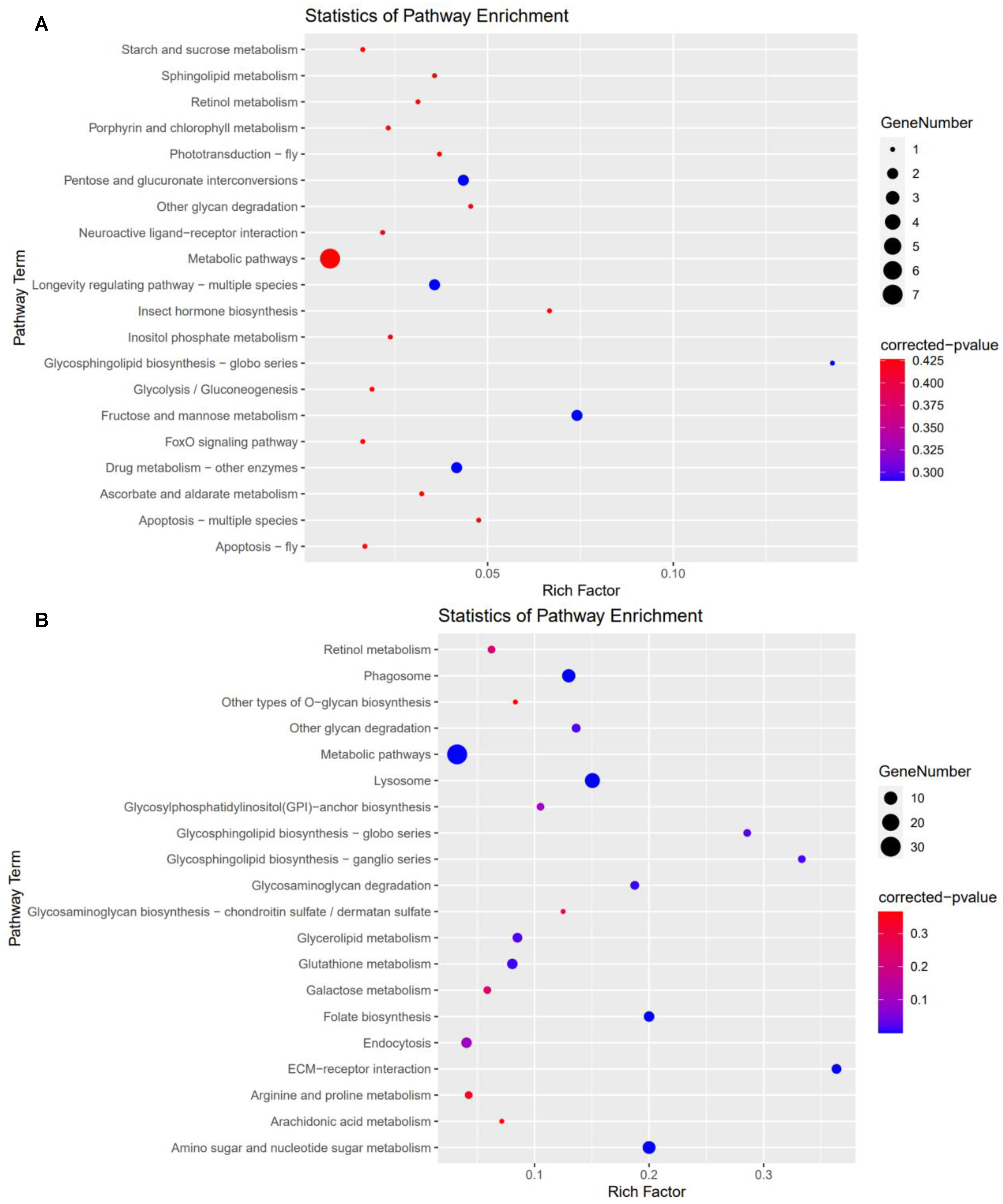
For high carbonate alkalinity stress treatment, 24 KEGG pathways (19 up−regulated and 5 down−regulated pathways) were significantly enriched in the HC_O group (Figure 3). Among these KEGG pathways, some were associated with ovarian development, such as ECM–receptor interaction, folate biosynthesis, FoxO signaling pathway, insect hormone biosynthesis, lysosome, metabolic pathways and neuroactive ligand–receptor interaction.
In the comparison of HC_E and CG_E group, 17 KEGG pathways (5 up−regulated and 12 down−regulated pathways) were significantly enriched (Figure 4). Concurrently, the ECM-receptor interaction, folate biosynthesis, lysosome, metabolic pathways, and retinol metabolism were associated with ovarian development.
3.6. DEGs Involved in Ovarian Development
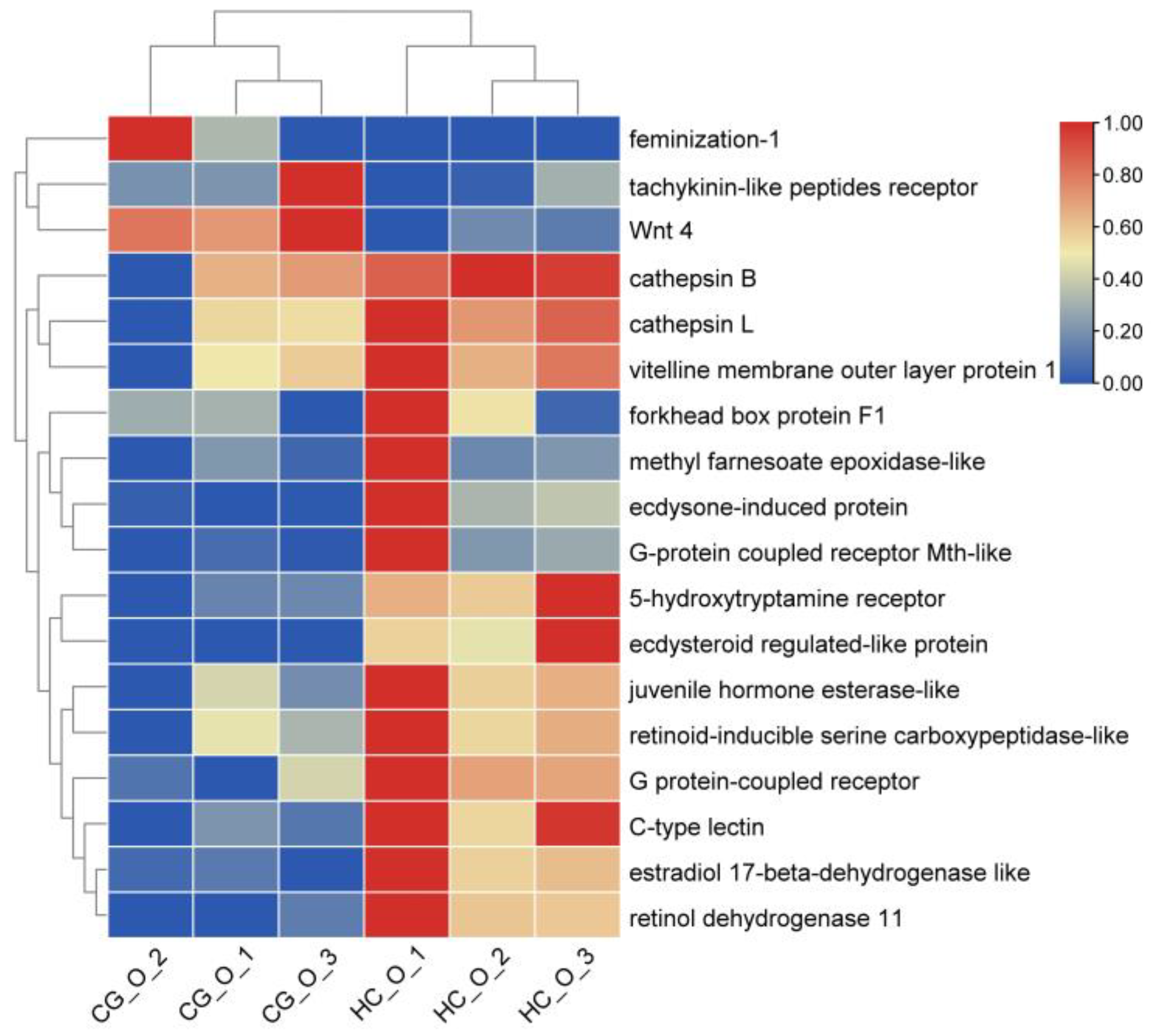
In the ovary, differential genes involved in ovarian development were selected high carbonate alkalinity stress, such as the G protein−coupled receptor, vitelline membrane outer layer protein 1 (VMO−1), retinol dehydrogenase 11, ecdysone−induced protein 78, ecdysteroid regulated−like protein, voltage−dependent calcium channel gamma−7, methyl farnesoate epoxidase, and estradiol 17−beta−dehydrogenase. A clustering heatmap of these results is provided (Figure 5).
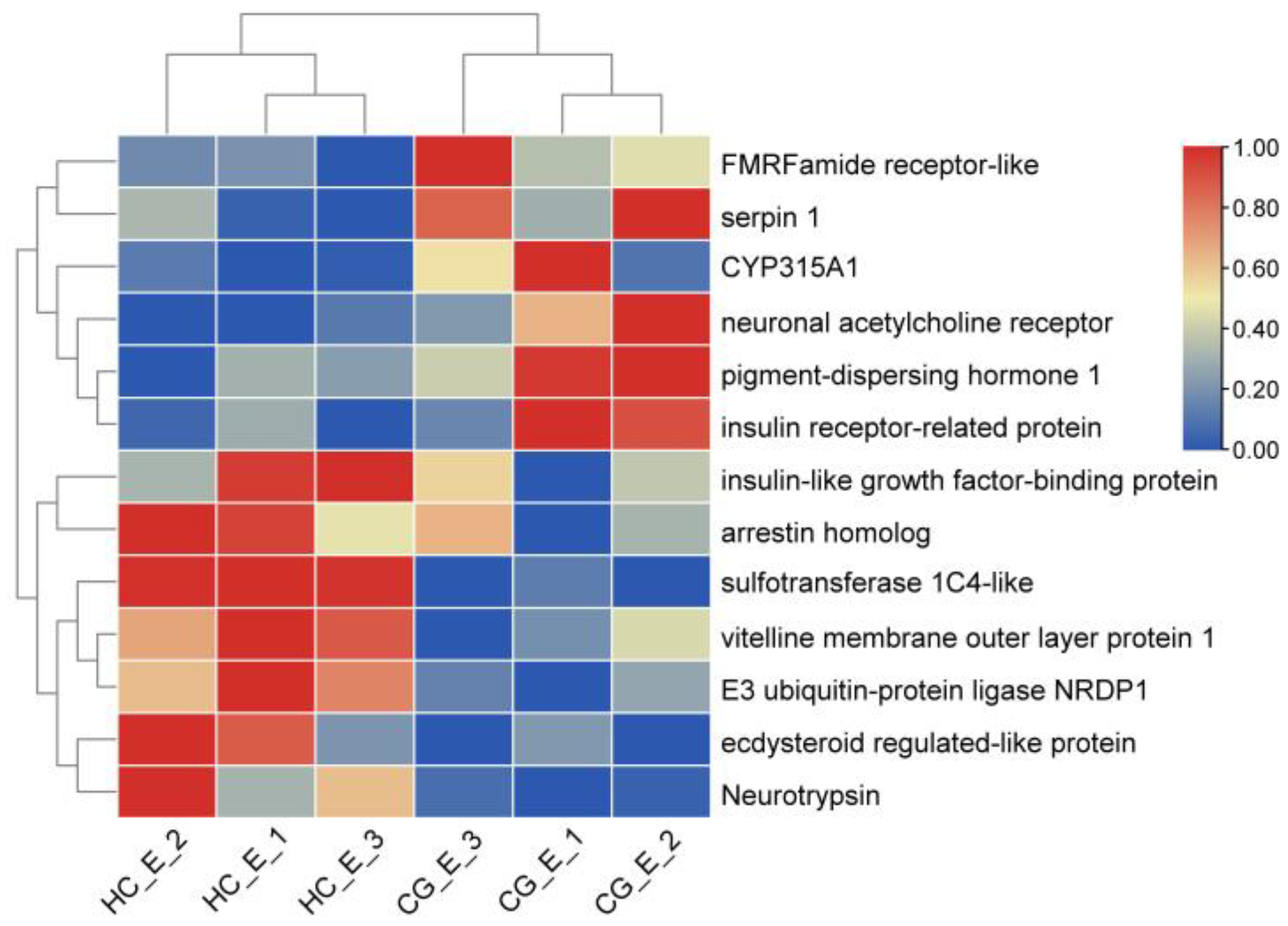
In the eyestalk, the participation of differential genes in ovarian development was observed, and genes were selected which showed high carbonate alkalinity stress. Some DEGs related to ovarian development were screened, such as the vitelline membrane outer layer protein 1, pigment−dispersing hormone 1, insulin−like growth factor binding protein, insulin receptor−related protein, NF−kappa B inhibitor alpha, neuronal acetylcholine receptor, neuroligin−2, and DNA replication complex GINS. A clustering heatmap of these results is provided (Figure 6).
3.7. The Validation of DEGs by qRT−PCR
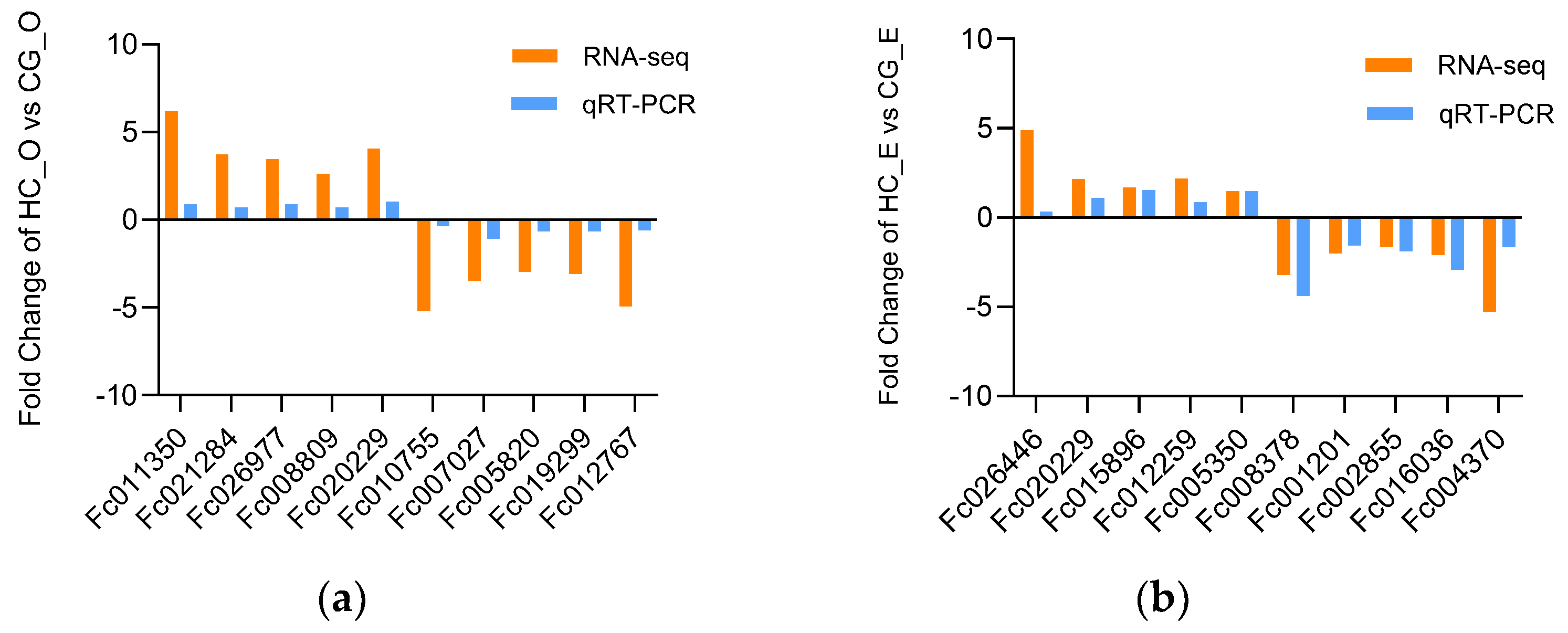
In order to further validate the reliability of the DEGs identified by RNA−Seq, we randomly selected 10 DEGs from two comparisons: HC_O vs. CG_O and HC_E vs. CG_E. The qRT−PCR results were consistent with those of RNA−seq, indicating that the RNA−Seq data was accurate (Figure 7).
4. Discussion
Ovarian maturation is an elaborate and complex molecular process, and a large number of genes need to be regulated to ensure the normal development of the oocytes. In this study, we found that 8 mmol/L carbonate alkalinity stress retarded E. carinicauda ovarian development, even in non−developing females [21]. The ovaries and eyestalks are important organs in crustacean ovarian development. Eyestalk ablation is commonly used to induce ovarian maturation in shrimp farming [29]. Although the mechanism by which eyestalk ablation leads to ovarian maturation remains inconclusive, some genes expressed in the eyestalk regulate ovarian development. The histological study investigates the influence of high carbonate alkalinity on ovary and oocyte development. We used high-throughput sequencing of ovaries and eyestalks for the first time to understand the ovarian development mechanisms in E. carinicauda when in high carbonate alkalinity osmotic stress.
KEGG pathway enrichment can identify major biochemical metabolic and signal transduction pathways involved in genes. In the HC_O group, the insect hormone biosynthesis pathway was significantly enriched, with methyl farnesoate epoxidase, which can catalyze methyl farnesoate into juvenile hormone (JH) III, differentially up−regulated. Methyl farnesoate is a non−epoxidized form of insect juvenile hormone III, which is secreted by the mandibular organs of crustaceans and has a significant stimulating effect on Vg synthesis in various Decapoda species [30,31,32]. Although JH Ⅲ plays a very important role in regulating life processes such as growth, molting, and reproduction, it is considered to be an important endogenous hormone in insects and other arthropods [33,34]. Up-regulated methylfarnesoate epoxidase may lead to a decrease in methylfarnesoate synthesis. However, in Macrobrachium rosenbergii [35] and Sagmariasus verreauxi [36], metabolic enzymes convert methyl farnesoate into JH, which supports the view that JH is also an active hormone in crustaceans. Furthermore, methyl farnesoate epoxidase can directly convert farnesoic acid into JH III acid to form JH III. Therefore, the up−regulation of methyl farnesoate epoxidase promotes the synthesis of JH, thus affecting ovarian development in crustaceans.
Lysosomes were the most significant pathways, with a larger number of DEGs varying between the HC_O and CG_O groups as well as between the HC_E and CG_E groups. Lysosomes are important for intracellular trafficking, metabolic signaling, lipid metabolism, and immune responses [37]. Lysosomes are involved in the preparation of free cholesterol for steroidogenesis and degradation of steroidogenesis regulators, as well as follicle rupture during ovulation in the ovaries of vertebrates [38]. As the central digestive organ of cells, various macromolecules are sent to lysosomes for degradation. Vitellogenin is an important precursor of egg yolk in nearly all oviparous animals [39]. Lysosomes play an important role in the degradation of vitellogenin, which is internalized by endocytosis [40]. Lysosomes are related to the hydrolysis of vitellogenin and energy demand during Macrobrachium nipponense ovarian maturation [41]. Lysosomal enzymes, especially cathepsin B and L, are associated with ovarian development in crustaceans [42,43]. We also found that cathepsin B and L in the ovaries of E. carinicauda were significantly up−regulated in response to carbonate alkalinity stress, which was consistent with the above results. In addition, we found that the lysosomal pathway was significantly up−regulated in the ovaries and significantly down−regulated in the eyestalks. Therefore, we speculate that this change in lysosomes is closely related to the ovarian development of E. carinicauda under carbonate alkalinity stress. However, the specific mechanism is unknown.
In this study, significant differential genes provide evidence of response to high carbonate alkalinity stress in the ovary and eyestalk, which are the target tissues that regulate ovarian development. As the largest membrane receptor family in eukaryotes, G protein coupled receptors are involved in regulating many key physiological and biochemical processes, including sexual maturation and reproduction [44,45]. There are many important factors controlling the development and maturation of shrimp ovaries, such as neurotransmitters, hormones, and their receptors [46,47], which mainly bind and activate G protein coupled receptors on the cell surface [48] and initiate multiple downstream cascades [49]. In this study, G protein−coupled receptor and Mth were significantly up−regulated in the HC_O group, which indicated that the expression of G protein−coupled receptor and Mth may regulate the ovarian development of E. carinicauda under high carbonate alkalinity stress.
As a glycoprotein bound to the cell surface, lectin specifically recognizes carbohydrates [50] and plays an important role in the innate immunity of invertebrates [51,52]. Qin et al. found that pmcl1 plays an important role in the immune response to pathogen infection and ammonia nitrogen stress [53]. Tateno et al. suggests that lectins in fish oocytes may prevent polysperm fertilization, regulate carbohydrate metabolism, participate in the formation of the fertilization shell after binding with glycoproteins, determine the source of disease, and have antibacterial effects [50]. Therefore, we speculate that C−type lectins may perform similar functions in E. carinicauda, but the specific role requires further research. Additionally, lectin and vitellin VMO1/2 are closely bound to ovomucin to form the basic skeleton of the outer membrane of vitellin [54]. The VMO−1 protein is essential for the formation of the outer membrane in chicken eggs [55], and the adhesion between yolk membranes may be related to the VMO−2 protein in quail (Coturnix japonica) eggs [56]. VMO−1 is an important protein in the development of oocytes [57], and its main function is to prevent mixing between yolk and protein in crayfish [58]. In this study, the expression of VMO−1 and C−type lectin was up−regulated in the HC_O group, suggesting that VMO−1 and lectin affects ovarian development in E. carinicauda under osmotic stress.
The crustacean eyestalk is known to regulate reproduction, molting, and energy metabolism [59,60]. Removing the eyestalk can induce ovarian maturation and oviposition in many crustaceans [61,62]. Eyestalk−derived neuropeptides regulate vitellogenesis in crustaceans. Pigment−dispersing hormone (PDH) is involved in the regulation of ovarian maturation in crustaceans [63]. The PDH may participate in vitellogenesis according to their spatiotemporal expression patterns, which maintained a high level from the pre−vitellogenesis stage and decreased significantly in the mature stage in Scylla paramamosain [64,65]. Wei et al. provided evidence of the inductive effect of PDH on oocyte meiotic maturation in E. sinensis [66]. In this study, the PDH was up−regulated in the HC_E group, suggesting that it may participate in the ovarian development of E. carinicauda under high carbonate alkalinity stress.
Retinol and its derivatives play key roles in the meiosis of mammalian fetal ovarian germ cells [67], follicular development [68], ovarian steroidogenesis [69], and oocyte maturation [70]. The retinol dehydrogenase (RDH) is a member of the short−chain dehydrogenase/reductase (SDR) superfamily, which includes three RDHs, RDH11, RDH12, and RDH13 in transcriptome sequences. The expression level of RDH13 in a vitellogenesis ovary of zebrafish was significantly higher than that in a non−vitellogenesis ovary [71]. In addition, knockout of RDH11 resulted in decreased transcription of vitellogenin and vitellogenin receptors in Procambarus clarkii [72], suggesting that RDH11 might play an important role in the synthesis and conveyance of vitellogenin in crustaceans. Our previous study revealed that RDH11 is critical for ovarian development in E. carinicauda [73]. In this study, RDH11, associated with ovarian development in E. carinicauda, was significantly expressed under high carbonate alkalinity stress.
5. Conclusions
In the present study, the transcriptome analysis of the ovary and eyestalk of E. carinicauda under high carbonate alkalinity stress, as well as the effects of high carbonate alkalinity on ovarian development-related genes and signaling pathways, are described for the first time. Eighteen and thirteen DEGs in ovary and eyestalk tissue were identified, respectively. The key genes were identified to be involved in the folate biosynthesis, insect hormone biosynthesis, lysosome, and retinol metabolism, which play an essential role in the response of the ovaries and eyestalks of E. carinicauda to high carbonate alkalinity stress. This study provided new insights into the ovarian development of E. carinicauda under high carbonate alkalinity stress, which could be useful for saline–alkaline water aquaculture and related studies on the reproduction of crustaceans.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/w14223690/s1, Table S1: Primers of RT−PCR designed for validation experiment of DEGs; Table S2: Number of high-throughput clean reads and mapped clean reads generated from E. carinicauda ovary and eyestalk mRNA library.
Author Contributions
X.Z., J.W. and J.L. (Jitao Li) conceived and designed the research; X.Z., J.W., C.W., W.L., Q.G. and Z.Q. performed the experiments and analyzed the data; J.L. (Jian Li) provided research ideas for the experiments. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Key R&D Program of China (2019YFD0900404-03), National Natural Science Foundation of China (32072974), China Agriculture Research System of MOF and MARA (CARS-48) and Central Public-Interest Scientific Institution Basal Research Fund, CAFS (2020TD46).
Institutional Review Board Statement
No human subjects were included in this study. The animal study was reviewed and approved by the Animal Ethics Committee of Yellow Sea Research Institute, CAFS (ID Number: YSFRI–2022033).
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: Center for Biotechnology Information (NCBI) with the accession number PRJNA881755 and PRJNA881756.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sharma, B.R.; Minhas, P.S. Strategies for managing saline/alkali waters for sustainable agricultural production in South Asia. Agric. Water Manag. 2005, 78, 136–151. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, C.; Liu, G.; Jing, J. Development of a cDNA microarray to identify gene expression of Puccinellia tenuiflora under saline-alkali stress. Plant Physiol. Bioch. 2007, 45, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.L.; Lai, Q.F.; Zhou, K.; Rizalita, R.E.; Wang, H. Developmental biology of medaka fish (Oryzias latipes) exposed to alkalinity stress. J. Appl. Ichthyol. 2010, 26, 397–402. [Google Scholar] [CrossRef]
- Fan, Z.; Wu, D.; Zhang, Y.Y.; Li, J.; Xu, Q.Y.; Wang, L.S. Carbonate alkalinity and dietary protein levels affected growth performance, intestinal immune responses and intestinal microflora in Songpu mirror carp (Cyprinus carpio Songpu). Aquaculture 2021, 545, 737135. [Google Scholar] [CrossRef]
- Lin, T.; Lai, Q.; Yao, Z.; Lu, J.; Zhou, K.; Wang, H. Combined effects of carbonate alkalinity and pH on survival, growth and haemocyte parameters of the Venus clam Cyclina sinensis. Fish Shellfish Immun. 2013, 35, 525–531. [Google Scholar] [CrossRef]
- Xu, J.; Ji, P.; Wang, B.; Zhao, L.; Wang, J.; Zhao, Z.; Zhang, Y.; Li, J.; Xu, P.; Sun, X. Transcriptome sequencing and analysis of wild amur ide (Leuciscus waleckii) inhabiting an extreme alkaline–saline lake reveals insights into stress adaptation. PLoS ONE 2013, 8, e59703. [Google Scholar] [CrossRef]
- Li, H.; Lai, Q.; Yao, Z.; Liu, Y.; Sun, Z. Ammonia excretion and blood gas variation in naked carp (Gymnocypris przewalskii) exposed to acute hypoxia and high alkalinity. Fish Physiol. Biochem. 2020, 46, 1981–1990. [Google Scholar] [CrossRef]
- Su, H.; Ma, D.; Zhu, H.; Liu, Z.; Gao, F. Transcriptomic response to three osmotic stresses in gills of hybrid tilapia (Oreochromis mossambicus female × O. urolepis hornorum male). BMC Genom. 2020, 21, 110. [Google Scholar] [CrossRef] [Green Version]
- Li, J.T.; Li, J.; Duan, Y.F.; Chen, P.; Liu, P. The roles of heat shock proteins 70 and 90 in Exopalaemon carinicauda after WSSV and Vibrio anguillarum challenges. J. Ocean Univ. China 2018, 17, 399–406. [Google Scholar] [CrossRef]
- Feng, Y.Y.; Zhai, Q.Q.; Wang, J.J.; Li, J.T.; Li, J. Comparison of florfenicol pharmacokinetics in Exopalaemon carinicauda at different temperatures and administration routes. J. Vet. Pharmacol. Ther. 2019, 42, 230–238. [Google Scholar] [CrossRef]
- Li, J.T.; Li, J.; Chen, P.; Liu, P.; He, Y.Y. Transcriptome analysis of eyestalk and hemocytes in the ridgetail white prawn Exopalaemon carinicauda: Assembly, annotation and marker discovery. Mol. Biol. Rep. 2015, 42, 135–147. [Google Scholar] [CrossRef] [PubMed]
- Li, J.T.; Ma, P.; Ping, L.; Chen, P.; Jian, L. The roles of Na+/K+-ATPase α-subunit gene from the ridgetail white prawn Exopalaemon carinicauda in response to salinity stresses. Fish Shellfish Immun. 2015, 42, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Ge, Q.Q.; Li, J.; Wang, J.J.; Li, Z.D.; Li, J.T. Characterization, functional analysis, and expression levels of three carbonic anhydrases in response to pH and saline–alkaline stresses in the ridgetail white prawn Exopalaemon carinicauda. Cell Stress Chaperon. 2019, 24, 503–515. [Google Scholar] [CrossRef] [PubMed]
- Chang, Z.Q.; Neori, A.; He, Y.Y.; Li, J.T.; Li, J. Development and current state of seawater shrimp farming, with an emphasis on integrated multi–trophic pond aquaculture farms, in China–a review. Rev. Aquacult. 2020, 4, 2544–2558. [Google Scholar] [CrossRef]
- González-Vera, C.; Brown, J.H. Effects of alkalinity and total hardness on growth and survival of postlarvae freshwater prawns, Macrobrachium rosenbergii (De Man 1879). Aquaculture 2017, 473, 521–527. [Google Scholar] [CrossRef]
- Yao, Z.L.; Wang, H.; Chen, L.; Zhou, K.; Ying, C.Q.; Lai, Q.F. Transcriptomic profiles of Japanese medaka (Oryzias latipes) in response to alkalinity stress. Genet. Mol. Res. 2012, 11, 2200–2246. [Google Scholar] [CrossRef]
- Mcfarland, K.; Donaghy, L.; Volety, A.K. Effect of acute salinity changes on hemolymph osmolality and clearance rate of the non-native mussel, Perna viridis, and the native oyster, Crassostrea virginica, in Southwest Florida. Aquat. Invasions 2013, 8, 299–310. [Google Scholar] [CrossRef]
- Sun, Y.C.; Han, S.C.; Yao, M.Z.; Liu, H.B.; Wang, Y.M. Exploring the metabolic biomarkers and pathway changes in crucian under carbonate alkalinity exposure using high–throughput metabolomics analysis based on UPLC–ESI–QTOF–MS. RSC Adv. 2020, 10, 1552–1571. [Google Scholar] [CrossRef]
- Zhao, W.; Wang, C.; Zhang, L.; Wei, J.; Yang, W.; Teng, L. Effects of alkalinity and pH on the survival, growth and neonate production of two strains of Moina mongolica Daday. Acta Ecol. Sin. 2009, 2, 589–598, (In Chinese with English Abstract). [Google Scholar]
- Xu, W.; Geng, L.W.; Li, C.T.; Jin, G.X.; Liu, X.Y. The artificial propagation, embryonic development and saline–alkali tolerant experiment of Barbus capito. J. Fish. China 2011, 2, 255–260, (In Chinese with English Abstract). [Google Scholar]
- Liu, F.; Li, J.; Li, J.T.; Ge, Q.Q.; Ge, H.X.; Shen, M.M. Effects of carbonate alkalinity stress on the survival, growth, reproduction, and immune enzyme activities of Exopalaemon carinicauda. J. Fish. Sci. China 2016, 5, 1137–1147, (In Chinese with English Abstract). [Google Scholar]
- Dugas, D.V. Functional annotation of the transcriptome of Sorghum bicolor in response to osmotic stress and abscisic acid. BMC Genom. 2011, 12, 514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teets, N.M.; Peyton, J.T.; Colinet, H.; Renault, D.; Denlinger, D.L. Gene expression changes governing extreme dehydration tolerance in an Antarctic insect. Proc. Natl. Acad. Sci. USA 2013, 109, 20744–20749. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Li, E.C.; Li, T.Y.; Xu, C.; Wang, X.D.; Lin, H.Z.; Qin, J.G.; Chen, L.Q. Transcriptome and molecular pathway analysis of the hepatopancreas in the pacific white shrimp Litopenaeus vannamei under chronic low–salinity stress. PLoS ONE 2015, 10, e131503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, E.; Wang, S.; Li, C.; Wang, X.; Chen, K.; Chen, L. Transcriptome sequencing revealed the genes and pathways involved in salinity stress of Chinese mitten crab, Eriocheir sinensis. Physiol. Genom. 2014, 46, 177–190. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; Mccarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Biogeosciences 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Bu, D.H.; Luo, H.T.; Huo, P.P.; Wang, Z.H.; Zhang, S.; He, Z.H.; Wu, Y.; Zhao, L.H.; Liu, J.J.; Guo, J.C.; et al. KOBAS–i: Intelligent prioritization and exploratory visualization of biological functions for gene enrichment analysis. Nucleic Acids Res. 2021, 49, 317–325. [Google Scholar] [CrossRef]
- Duan, Y.F.; Liu, P.; Li, J.T.; Li, J.; Gao, B.Q. cDNA cloning, characterization and expression analysis of peroxiredoxin 5 gene in the ridgetail white prawn Exopalaemon carinicauda. Mol. Biol. Rep. 2013, 40, 6569–6577. [Google Scholar] [CrossRef] [Green Version]
- Sittikankaew, K.; Pootakham, W.; Sonthirod, C.; Sangsrakru, D.; Yoocha, T.; Khudet, J.; Nookaew, I.; Uawisetwathana, U.; Rungrassamee, W.; Karoonuthaisiri, N. Transcriptome analyses reveal the synergistic effects of feeding and eyestalk ablation on ovarian maturation in black tiger shrimp. Sci. Rep. 2020, 10, 3229. [Google Scholar]
- Mak, A.S.C.; Choi, C.L.; Tiu, S.H.K.; Hui, J.H.L.; He, J.G.; Tobe, S.S.; Chan, S.M. Vitellogenesis in the red crab Charybdis feriatus: Hepatopancreas–specific expression and farnesoic acid stimulation of vitellogenin gene expression. Mol. Reprod. Dev. 2005, 70, 288–300. [Google Scholar] [CrossRef]
- Yang, Y.; Ye, H.H.; Huang, H.Y.; Jin, Z.X.; Li, S.J. Cloning, expression and functional analysis of farnesoic acid O-methyltransferase (FAMeT) in the mud crab, Scylla paramamosain. Mar. Freshw. Behav. Phy. 2012, 45, 209–222. [Google Scholar] [CrossRef]
- Miyakawa, H.; Toyota, K.; Sumiya, E.; Iguchi, T. Comparison of JH signaling in insects and crustaceans. Curr. Opin. Insect Sci. 2014, 1, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Veeran, S.; Wang, J.; Li, S.; Liu, S.N. Dual roles of juvenile hormone signaling during early oogenesis in Drosophila. Insect Sci. 2019, 27, 665–674. [Google Scholar] [CrossRef]
- Suang, S.; Hiruma, K.; Yu, K.; Manaboon, M. Diapause hormone directly stimulates the prothoracic glands of diapause larvae under juvenile hormone regulation in the bamboo borer, Omphisa fuscidentalis Hampson. Arch. Insect Biochem. 2019, 102, e21603. [Google Scholar] [CrossRef] [PubMed]
- Tomer, V.; Rivka, M.; Aflalo, E.D.; Vered, C.C.; Simy, W.; Omri, S.; Amir, S.; Yu, J. Post–embryonic transcriptomes of the prawn Macrobrachium rosenbergii: Multigenic succession through metamorphosis. PLoS ONE 2013, 8, e55322. [Google Scholar]
- Ventura, T.; Fitzgibbon, Q.P.; Battaglene, S.C.; Elizur, A. Redefining metamorphosis in spiny lobsters: Molecular analysis of the phyllosoma to puerulus transition in Sagmariasus verreauxi. Sci. Rep. 2015, 5, 13537. [Google Scholar] [CrossRef] [Green Version]
- Lamming, D.W.; Bar-Peled, L. Lysosome: The metabolic signaling hub. Traffic 2019, 20, 27–38. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Wang, Z.; Andersen, C.L.; Ye, X. Functions of lysosomes in mammalian female reproductive system. Reprodu. Dev. Med. 2020, 4, 109–122. [Google Scholar] [CrossRef]
- Harwood, G.; Amdam, G. Vitellogenin in the honey bee midgut. Apidologie 2021, 52, 837–847. [Google Scholar] [CrossRef]
- Carnevali, O.; Cionna, C.; Tosti, L.; Lubzens, E.; Maradonna, F. Role of cathepsins in ovarian follicle growth and maturation. Gen. Comp. Endocr. 2006, 146, 195–203. [Google Scholar] [CrossRef]
- Zhang, Y.N.; Jiang, S.F.; Qiao, H.; Xiong, Y.W.; Fu, H.T.; Zhang, W.Y.; Gong, Y.S.; Jin, S.B.; Wu, Y. Transcriptome analysis of five ovarian stages reveals gonad maturation in female updates Macrobrachium nipponense. BMC Genom. 2021, 22, 510. [Google Scholar] [CrossRef] [PubMed]
- Luo, B.Y.; Qian, H.L.; Jiang, H.C.; Xiong, X.Y.; Ye, B.Q.; Liu, X.; Guo, Z.Q.; Ma, K.Y. Transcriptional changes revealed water acidification leads to the immune response and ovary maturation delay in the Chinese mitten crab Eriocheir sinensis. Comp. Biochem. Phys. D 2021, 39, 100868. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Qi, D.; Chen, L.Q.; Zhang, H.; Zhang, X.W.; Qin, J.G.; Hu, S.N. Gene discovery from an ovary cDNA library of oriental river prawn Macrobrachium nipponense by ESTs annotation. Comp. Biochem. Phys. D 2009, 4, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.V.; Rotllant, G.E.; Cummins, S.F.; Elizur, A.; Ventura, T. Insights into sexual maturation and reproduction in the norway lobster (Nephrops norvegicus) via in silico prediction and characterization of neuropeptides and G protein–coupled receptors. Front. Endocrinol. 2018, 9, 430. [Google Scholar] [CrossRef] [Green Version]
- Flaherty, P.; Radhakrishnan, M.L.; Dinh, T.; Rebres, R.A.; Roach, T.I.; Jordan, M.I.; Arkin, A.P. A dual receptor crosstalk model of G protein coupled signal transduction. PLoS Comput. Biol. 2008, 4, e1000185. [Google Scholar] [CrossRef]
- Subramoniam, T. Mechanisms and control of vitellogenesis in crustaceans. Fish. Sci. 2011, 77, 1–21. [Google Scholar] [CrossRef]
- Crown, A.; Clifton, D.K.; Steiner, R.A. Neuropeptide signaling in the integration of metabolism and reproduction. Neuroendocrinology 2016, 86, 175–182. [Google Scholar] [CrossRef]
- Sharabi, O.; Manor, R.; Weil, S.; Aflalo, E.D.; Lezer, Y.; Levy, T.; Aizen, J.; Ventura, T.; Mather, P.B.; Khalaila, I. Identification and characterization of an insulin–like receptor involved in crustacean reproduction. Endocrinology 2016, 157, 928–941. [Google Scholar] [CrossRef] [Green Version]
- Christie, A.E.; Stemmler, E.A.; Dickinson, P.S. Crustacean neuropeptides. Cell. Mol. Life Sci. 2010, 67, 4135–4169. [Google Scholar] [CrossRef]
- Tateno, H.; Saneyoshi, A.; Ogawa, T.; Muramoto, K.; Kamiya, H.; Saneyoshi, M. Isolation and characterization of rhamnose-binding lectins from eggs of steelhead trout (Oncorhynchus mykiss) homologous to low density lipoprotein receptor superfamily. J. Biol. Chem. 1998, 273, 19190–19197. [Google Scholar] [CrossRef] [Green Version]
- Dodd, R.B.; Kurt, D. Lectin-like proteins in model organisms: Implications for evolution of carbohydrate–binding activity. Glycobiology 2001, 11, 71–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Z.; Zhang, J.Q.; Li, F.H.; Liu, C.Z.; Xiang, J.H. Cloning of a novel C–type lectin LvLec2 from the shrimp Litopenaeus vannamei and its immune response to different challenges. Mar. Sci. 2010, 34, 103–110. [Google Scholar]
- Qin, Y.; Jiang, S.; Huang, J.; Zhou, F.; Yang, Q.; Jiang, S.; Yang, L. C–type lectin response to bacterial infection and ammonia nitrogen stress in tiger shrimp (Penaeus monodon). Fish Shellfish Immun. 2019, 90, 188–198. [Google Scholar] [CrossRef]
- Kido, S.; Morimoto, A.; Kim, F.; Doi, Y. Isolation of a novel protein from the outer layer of the vitelline membrane. Biochem. J. 1992, 286, 17–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Back, J.F.; Bain, J.M.; Vadehra, D.V.; Burley, R.W. Proteins of the outer layer of the vitelline membrane of hen’s eggs. Biochim. Biophys. Acta. 1982, 705, 12–19. [Google Scholar] [CrossRef]
- Rahman, M.A.; Akihiko, M.; Atsushi, I.; Norio, Y. VMO–II mediates the binding of the chalaziferous layer with the vitelline membrane in quail eggs. J. Poult. Sci. 2009, 46, 240–248. [Google Scholar] [CrossRef]
- Gismondi, E.; Thome, J.P.; Urien, N.; Uher, E.; Baiwir, D.; Mazzucchelli, G.; De Pauw, E.; Fechner, L.C.; Lebrun, J.D. Ecotoxicoproteomic assessment of the functional alterations caused by chronic metallic exposures in gammarids. Environ. Pollut. 2017, 225, 428–438. [Google Scholar] [CrossRef]
- Sricharoen, S.; Kim, J.J.; Tunkijjanukij, S.; Soderhall, I. Exocytosis and proteomic analysis of the vesicle content of granular hemocytes from a crayfish. Dev. Comp. Immunol. 2005, 29, 1017–1031. [Google Scholar] [CrossRef]
- Philip, B.; Abigail, E.; Richard, W.; Cummins, S.F.; Wayne, K. Gene expression profiling of the cephalothorax and eyestalk in penaeus monodon during ovarian maturation. Int. J. Biol. Sci. 2012, 8, 328–343. [Google Scholar]
- Jung, H.T.; Lyons, R.E.; Hurwood, D.A.; Mather, P. Genes and growth performance in crustacean species: A review of relevant genomic studies in crustaceans and other taxa. Rev. Aquacult. 2013, 5, 77–110. [Google Scholar] [CrossRef]
- Koshio, S.; Teshima, S.I.; Kanazawa, A. Effects of unilateral eyestalk ablation and feeding frequencies on growth, survival, and body compositions of juvenile freshwater prawn Macrobrachium rosenbergii. Nippon Suisan Gakk. 1992, 58, 1419–1425. [Google Scholar] [CrossRef]
- Qiao, H.; Xiong, Y.; Zhang, W.; Fu, H.; Jiang, S.; Sun, S.; Bai, H.; Jin, S.; Gong, Y. Characterization, expression, and function analysis of gonad–inhibiting hormone in Oriental River prawn, Macrobrachium nipponense and its induced expression by temperature. Comp. Biochem. Phys. A 2015, 185, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Rotllant, G.; Nguyen, T.V.; Aizen, J.; Suwansa-ard, S.; Ventura, T. Toward the identification of female gonad–stimulating factors in crustaceans. Hydrobiologia 2018, 825, 91–119. [Google Scholar] [CrossRef]
- Huang, X.; Ye, H.; Huang, H.; Yu, K.; Huang, Y. Two beta-pigment–dispersing hormone (β–PDH) isoforms in the mud crab, Scylla paramamosain: Implication for regulation of ovarian maturation and a photoperiod-related daily rhythmicity. Anim. Reprod. Sci. 2014, 150, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Bao, C.; Yang, Y.; Huang, H.; Ye, H. Neuropeptides in the cerebral ganglia of the mud crab, Scylla paramamosain: Transcriptomic analysis and expression profiles during vitellogenesis. Sci. Rep. 2015, 5, 17055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, L.L.; Chen, T.T.; Luo, B.Y.; Qiu, G.F. Evidences for Red Pigment Concentrating Hormone (RPCH) and Beta-Pigment Dispersing Hormone (beta–PDH) Inducing Oocyte Meiotic Maturation in the Chinese Mitten Crab, Eriocheir sinensis. Front. Endocrinol. 2021, 12, 802768. [Google Scholar] [CrossRef]
- Jiang, Y.W.; Li, C.J.; Chen, L.; Wang, F.G.; Zhou, X. Potential role of retinoids in ovarian physiology and pathogenesis of polycystic ovary syndrome. Clin. Chim. Acta 2017, 469, 87–93. [Google Scholar] [CrossRef]
- Kawai, T.; Yanaka, N.; Richards, J.S.; Shimada, M. De novo–synthesized retinoic acid in ovarian antral follicles enhances FSH–mediated ovarian follicular cell differentiation and female fertility. Endocrinology 2016, 157, 2160–2172. [Google Scholar] [CrossRef] [Green Version]
- Damdimopoulou, P.; Chiang, C.; Flaws, J.A. Retinoic acid signaling in ovarian folliculogenesis and steroidogenesis. Reprod. Toxicol. 2019, 87, 32–41. [Google Scholar] [CrossRef]
- Tahaei, L.S.; Eimani, H.; Yazdi, P.E.; Ebrahimi, B.; Fathi, R. Effects of retinoic acid on maturation of immature mouse oocytes in the presence and absence of a granulosa cell co–culture system. J. Assist. Reprod. Gen. 2011, 28, 553–558. [Google Scholar] [CrossRef] [Green Version]
- Levi, L.; Ziv, T.; Admon, A.; Levavi-Sivan, B.; Lubzens, E. Insight into molecular pathways of retinal metabolism, associated with vitellogenesis in zebrafish. Am. J. Physiol.-Endoc. Metab. 2012, 302, 626–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, P.F.; Mao, B.; Fan, C.; Wang, Y.F. Transcriptomic information from the ovaries of red swamp crayfish (Procambarus clarkii) provides new insights into development of ovaries and embryos. Aquaculture 2019, 505, 333–343. [Google Scholar] [CrossRef]
- Wang, J.J.; Li, J.T.; Ge, Q.Q.; Li, W.Y.; Li, J. Full–Length transcriptome sequencing and comparative transcriptomic analysis provide insights into the ovarian maturation of Exopalaemon carinicauda. Front. Mar. Sci. 2022, 9, 906730. [Google Scholar] [CrossRef]
Figure 1.
Histological sections of the ovarian status of E. carinicauda: (a) CG_O group; (b) HC_O group. Bar: 50 µm. N: nucleus; Nu: nucleolus; Fc: follicular cell.
Figure 1.
Histological sections of the ovarian status of E. carinicauda: (a) CG_O group; (b) HC_O group. Bar: 50 µm. N: nucleus; Nu: nucleolus; Fc: follicular cell.

Figure 2.
The number of differentially expressed genes between different groups. (a) HC_O vs. CG_O group; (b) HC_E vs. CG_E group. Significantly up−regulated and down−regulated genes are indicated in red and blue, respectively, and those not significantly different are in gray.
Figure 2.
The number of differentially expressed genes between different groups. (a) HC_O vs. CG_O group; (b) HC_E vs. CG_E group. Significantly up−regulated and down−regulated genes are indicated in red and blue, respectively, and those not significantly different are in gray.

Figure 3.
The KEGG significant enrichment pathway in HC_O vs. CG_O group. (A) Up−regulation pathways in HC_O vs. CG_O group; (B) down−regulation pathways in HC_O vs. CG_O group.
Figure 3.
The KEGG significant enrichment pathway in HC_O vs. CG_O group. (A) Up−regulation pathways in HC_O vs. CG_O group; (B) down−regulation pathways in HC_O vs. CG_O group.

Figure 4.
The KEGG significant enrichment pathway in HC_E vs. CG_E group. (A) up−regulation pathways in HC_E vs. CG_E group; (B) down−regulation pathways in HC_E vs. CG_E group.
Figure 4.
The KEGG significant enrichment pathway in HC_E vs. CG_E group. (A) up−regulation pathways in HC_E vs. CG_E group; (B) down−regulation pathways in HC_E vs. CG_E group.

Figure 5.
The heatmap of DEGs related to ovarian development in the HC_O vs. CG_O group. The columns and rows indicate individuals and genes, respectively. The color scale represents FPKM after standard normalization, the same as below.
Figure 5.
The heatmap of DEGs related to ovarian development in the HC_O vs. CG_O group. The columns and rows indicate individuals and genes, respectively. The color scale represents FPKM after standard normalization, the same as below.

Figure 6.
The heatmap of DEGs related to ovarian development in the HC_E vs. CG_E group.

Figure 7.
The results were verified by qRT−PCR. (a) Relative fold change of DEGs between qRT−PCR and RNA−seq results in HC_O vs. CG_O group. (b) Relative fold change of DEGs between qRT-PCR and RNA−seq results in HC_E vs. CG_E group. Relative expression levels from the RNA−seq results were calculated as log2FC values. Fc011350 ecdysteroid regulated; Fc021284 legumain; Fc026977 lysosome−associated membrane glycoprotein 1; Fc008809 baculoviral IAP repeat-containing; Fc020229 N−acetylated−alpha−linked acidic dipeptidase; Fc010755 high−affinity choline transporter 1; Fc007027 Wnt 4; Fc019299 tachykinin−like peptides receptor; Fc005820 reelin 3; Fc012767 facilitated trehalose transporter; Fc026446 neuroligin 2; Fc015896 insulin receptor−related; Fc012259 serpin 1; Fc005350 UDP−glucosyltransferase 2; Fc008378 crustacyanin−C1; Fc001201 legumain; Fc002855 cathepsin L; Fc016036 multidrug resistance−associated protein; Fc004370 macrophage mannose receptor 1.
Figure 7.
The results were verified by qRT−PCR. (a) Relative fold change of DEGs between qRT−PCR and RNA−seq results in HC_O vs. CG_O group. (b) Relative fold change of DEGs between qRT-PCR and RNA−seq results in HC_E vs. CG_E group. Relative expression levels from the RNA−seq results were calculated as log2FC values. Fc011350 ecdysteroid regulated; Fc021284 legumain; Fc026977 lysosome−associated membrane glycoprotein 1; Fc008809 baculoviral IAP repeat-containing; Fc020229 N−acetylated−alpha−linked acidic dipeptidase; Fc010755 high−affinity choline transporter 1; Fc007027 Wnt 4; Fc019299 tachykinin−like peptides receptor; Fc005820 reelin 3; Fc012767 facilitated trehalose transporter; Fc026446 neuroligin 2; Fc015896 insulin receptor−related; Fc012259 serpin 1; Fc005350 UDP−glucosyltransferase 2; Fc008378 crustacyanin−C1; Fc001201 legumain; Fc002855 cathepsin L; Fc016036 multidrug resistance−associated protein; Fc004370 macrophage mannose receptor 1.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The GO term of related to ovarian development in the HC_O vs. CG_O group.
| Term | Significant | Up/Down | ID | p Value |
|---|---|---|---|---|
| Steroid metabolic process | 31 | up | 0008202 | 5.70 × 10−7 |
| Sterol homeostasis | 11 | up | 0055092 | 9.50 × 10−5 |
| Molting cycle | 22 | up | 0042303 | 0.00016 |
| Follicle cell microvillus organization | 3 | up | 0032529 | 0.00031 |
| Retinoid metabolic process | 12 | up | 0001523 | 0.00052 |
| Steroid biosynthetic process | 16 | up | 0006694 | 0.00076 |
| Sterol transport | 11 | up | 0015918 | 0.00486 |
| Response to estrogen | 9 | up | 0043627 | 0.00875 |
| Estrogen secretion | 2 | up | 0035937 | 0.01058 |
| Positive regulation of ovulation | 3 | down | 0060279 | 0.00099 |
| Regulation of ovulation | 3 | down | 0060278 | 0.00176 |
| Sterol transport | 6 | down | 0015918 | 0.02138 |
| Ovulation | 3 | down | 0030728 | 0.03153 |
| Sterol homeostasis | 4 | down | 0055092 | 0.04769 |
Table 2.
The GO term of related to ovarian development in the HC_E vs. CG_E group.
| Term | Significant | Up/Down | ID | p Value |
|---|---|---|---|---|
| Positive regulation of sterol transport | 2 | up | 0032373 | 0.00482 |
| Regulation of sterol transport | 2 | up | 0032371 | 0.01030 |
| Embryonic morphogenesis | 10 | up | 0048598 | 0.02561 |
| Embryonic organ morphogenesis | 5 | up | 0048562 | 0.02572 |
| Embryo development | 16 | up | 0009790 | 0.02894 |
| Entry into reproductive diapause | 1 | up | 0055116 | 0.04250 |
| Sterol metabolic process | 5 | up | 0008202 | 0.02501 |
| Sterol transport | 7 | down | 0015918 | 0.00226 |
| Regulation of metaphase/anaphase transition of meiosis I | 1 | down | 1905186 | 0.01765 |
| Ecdysteroid metabolic process | 3 | down | 0045455 | 0.02708 |
| Sterol homeostasis | 4 | down | 0055092 | 0.02759 |
| Maternal placenta development | 2 | down | 0001893 | 0.02798 |
| Metaphase/anaphase transition of meiosis I | 1 | down | 1905186 | 0.01765 |
| Molting cycle | 8 | down | 0042303 | 0.03756 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhang, X.; Wang, J.; Wang, C.; Li, W.; Ge, Q.; Qin, Z.; Li, J.; Li, J. Effects of Long-Term High Carbonate Alkalinity Stress on the Ovarian Development in Exopalaemon carinicauda. Water 2022, 14, 3690. https://doi.org/10.3390/w14223690
AMA Style
Zhang X, Wang J, Wang C, Li W, Ge Q, Qin Z, Li J, Li J. Effects of Long-Term High Carbonate Alkalinity Stress on the Ovarian Development in Exopalaemon carinicauda. Water. 2022; 14(22):3690. https://doi.org/10.3390/w14223690
Chicago/Turabian StyleZhang, Xiuhong, Jiajia Wang, Chengwei Wang, Wenyang Li, Qianqian Ge, Zhen Qin, Jian Li, and Jitao Li. 2022. "Effects of Long-Term High Carbonate Alkalinity Stress on the Ovarian Development in Exopalaemon carinicauda" Water 14, no. 22: 3690. https://doi.org/10.3390/w14223690
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.