

Influence of Seawater Salinity on the Survival, Growth, Development and Neonate Production of Scapholeberis mucronata (O. F. Müller) (Crustacea: Cladocera)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Incubation and Preparation
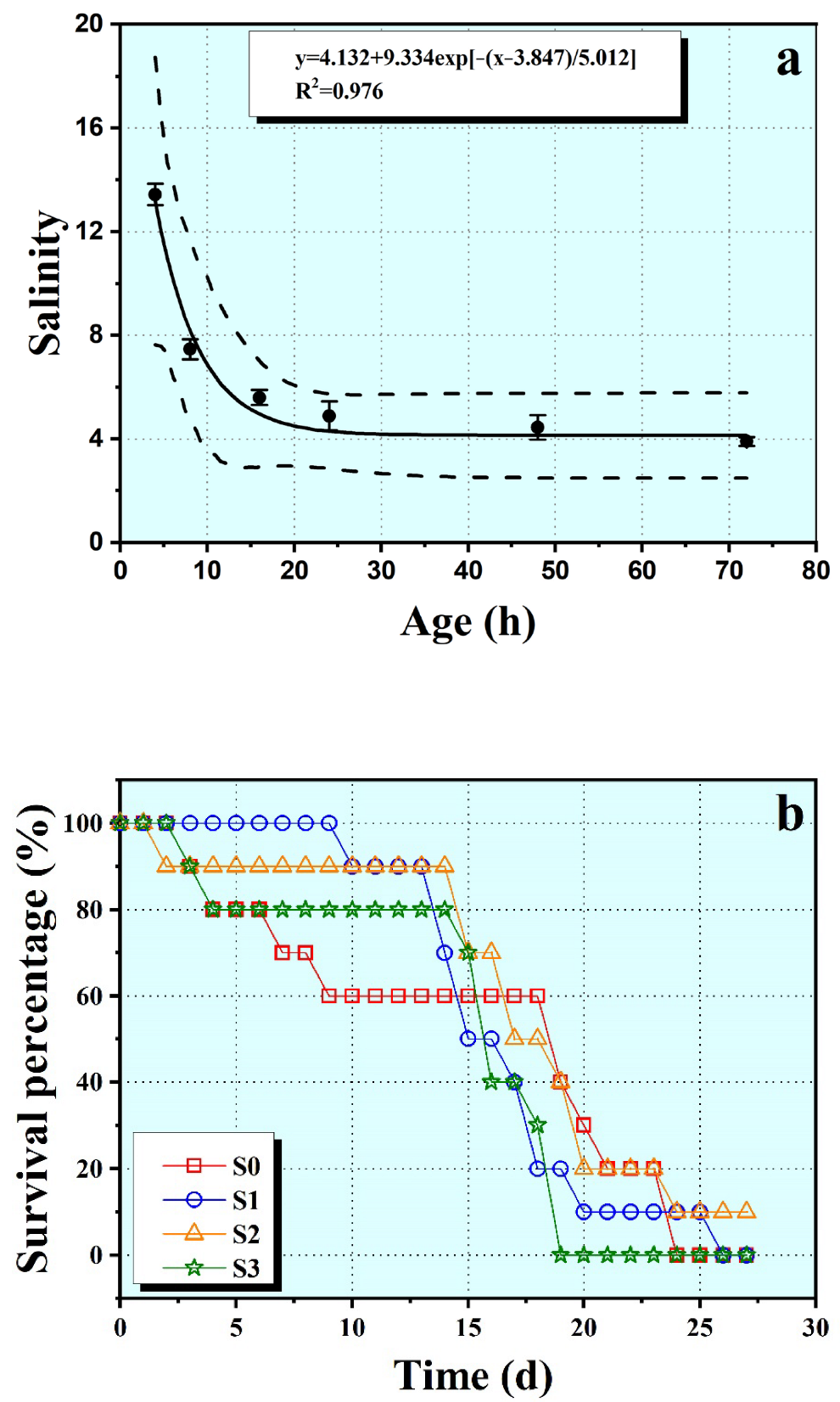
2.2. Median Lethal Concentration (LC50) Experiment
2.3. Life Table Analysis and Reproduction
2.4. Embryonic Development
2.5. Statistical Analysis
3. Results
3.1. Survival
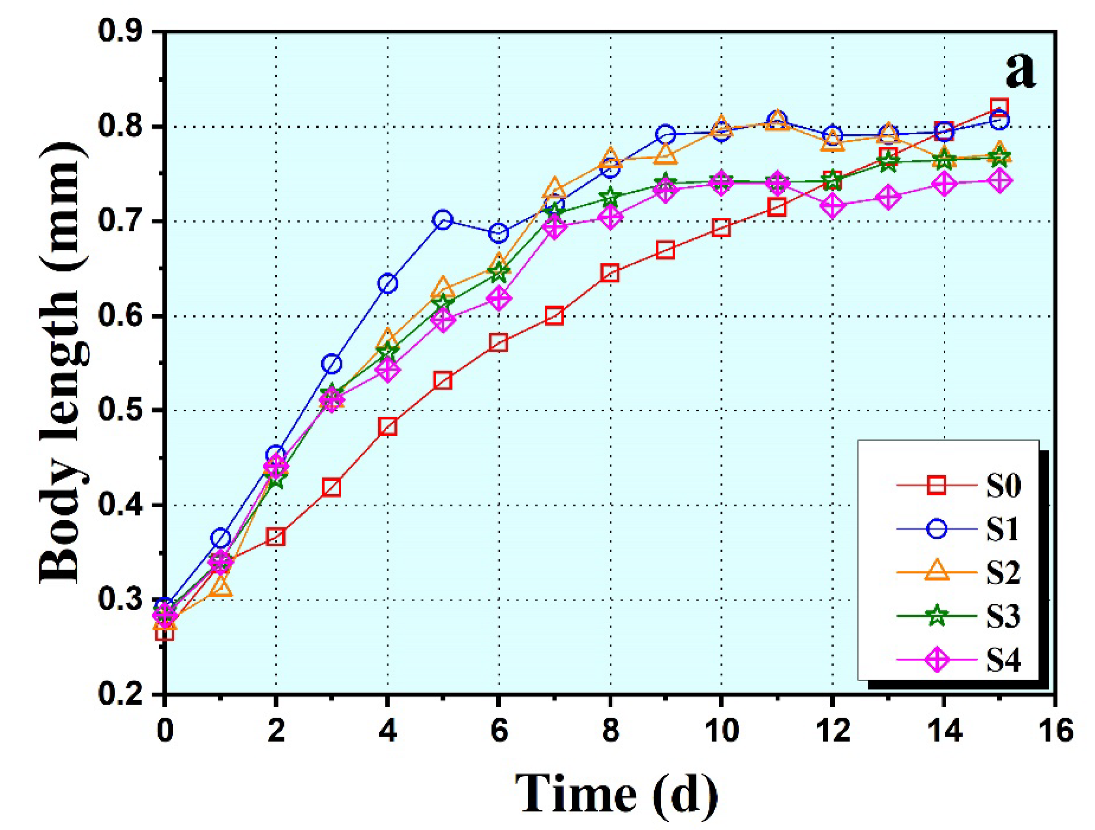
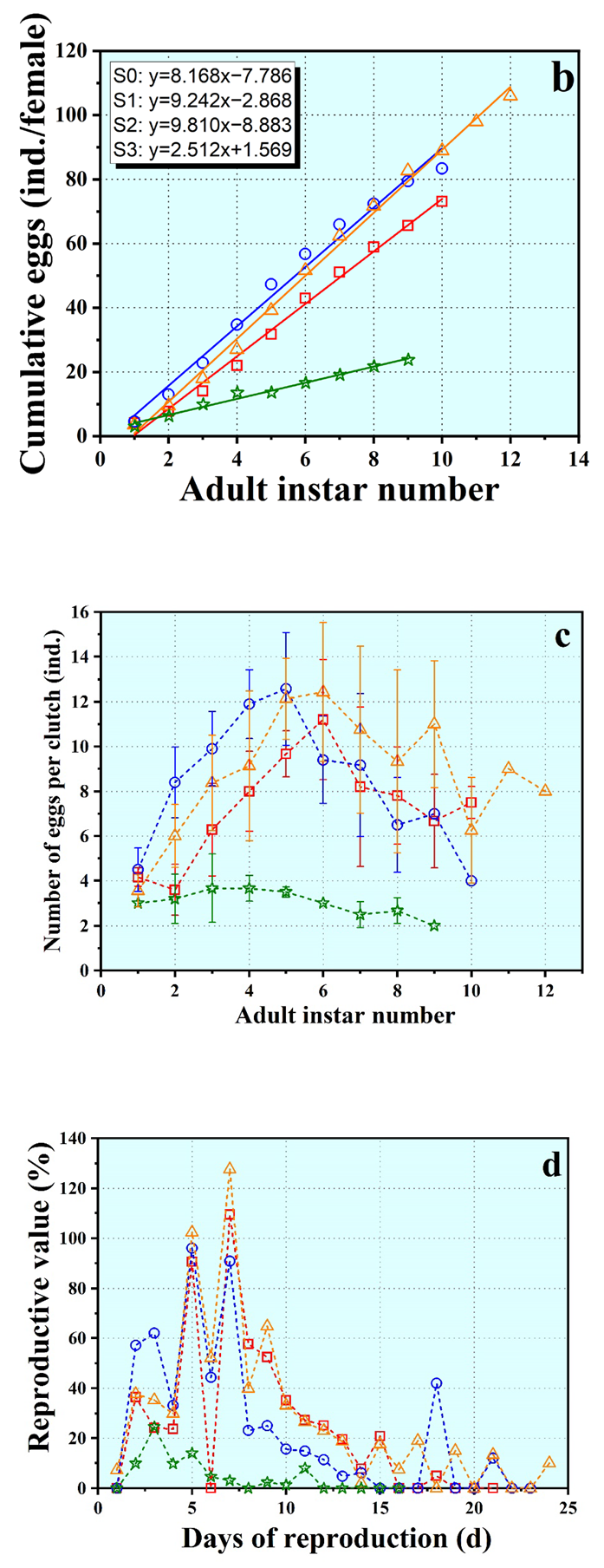
3.2. Growth and Reproduction
3.3. Life Table
3.4. Embryonic Development
3.4.1. Embryonic Development Process of S. mucronata
- Stage 1–2: The egg changed from elongate columniform to oval, and the ovoplasm changed from even to translucent with a transparent peripheral zone. The central region of the egg had fat cells surrounded by cleaved peripheral granulated cells. At this stage, both the outer egg membrane as well as the inner naupliar membrane were seen (Figure 3a–c).
- Stage 3–6: The egg divided from one cell to multiple cells. The vegetal and the animal pole appeared, and cell differentiation began (Figure 3d–g).
- Stage 7–8: The blastula and gastrula formed in order (Figure 3h,i).
- Stage 9–11: The antennae elongated and formed a “T”-type embryo. The head rudiment was formed. Then, distinct head and limb rudiments were formed, but the eyes were not apparent (Figure 3j–l).
- Stage 12–13: The embryo had two very small pink eyes, but these rapidly increased in size and became two large black eyes. Then, a distinct single compound eye was formed, and the embryonic heart began to beat. Juveniles in the brood chamber turned around usually (Figure 3m,n).
- Stage 14–15: The egg membrane was cast off, and characteristics of adult morphology such as the straight ventral margin of the shell ending posteriorly in a spine, the quadrate shape, the fine short setae on the ventral margin of the shell and the reddish color developed. Then juveniles were released from the brood chamber and moved freely (Figure 3o-3q).
3.4.2. Embryonic Development at Different Salinities
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dodson, S.I.; Frey, D.G. Cladocera and other Branchiopoda. In Ecology and Classification of North American Freshwater Invertebrates; Thorp, J.H., Covich, A.P., Eds.; Academic Press: San Diego, CA, USA, 1991; pp. 723–786. [Google Scholar]
- Lemly, A.D.; Dimmick, J.F. Growth of young-of-the-year and yearling centrarchids in relation to zooplankton in the littoral zone of lakes. Copeia 1982, 2, 305–321. [Google Scholar] [CrossRef]
- Mengestou, S.; Fernando, C.H. Biomass and production of the major dominant crustacean zooplankton in a tropical Rift Valley lake, Awasa, Ethiopia. J. Plankton Res. 1991, 13, 831–851. [Google Scholar] [CrossRef]
- Mavuti, K.M. Durations of development and production estimates by two crustacean zooplankton species Thermocyclops oblongatus Sars (Copepoda) and Diaphanosoma excisum Sars (Cladocera), in Lake Naivasha, Kenya. Hydrobiologia 1994, 272, 185–200. [Google Scholar] [CrossRef]
- Aladin, N.V. Salinity tolerance and morphology of the osmoregulation organs in Cladocera with special reference to Cladocera from the Aral sea. Hydrobiologia 1991, 225, 291–299. [Google Scholar] [CrossRef]
- Hart, B.T.; Bailey, P.; Edwards, R.; Hortle, K.; James, K.; McMahon, A.; Meredith, C.; Swadling, K. A Review of salt sensitivity of the Australian freshwater biota. Hydrobiologia 1991, 210, 105–144. [Google Scholar] [CrossRef]
- Williams, W.D. Salinity as a determinant of the structure of biological communities in salt lakes. Hydrobiologia 1998, 381, 191–201. [Google Scholar] [CrossRef]
- Latta, L.C.; Weider, L.J.; Colbourne, J.K.; Pfrender, M.E. The evolution of salinity tolerance in Daphnia: A functional genomics approach. Ecol. Lett. 2012, 15, 794–802. [Google Scholar] [CrossRef]
- He, Z.; An, S. Effects of salinity of seawater on the Moina rectirostris (Cladocera). J. Zool. 1986, 21, 25–27. (In Chinese) [Google Scholar]
- Shadrin, N.; Yakovenko, V.; Anufriieva, E. Gammarus aequicauda and Moina salina in the Crimean saline waters: New experimental and field data on their trophic relation. Aquac. Res. 2020, 51, 3091–3099. [Google Scholar] [CrossRef]
- Shadrin, N.V.; Yakovenko, V.A.; Anufriieva, E.V. Appearance of a New Species of Cladocera (Anomopoda, Chydoridae, Bosminidae) in the Hypersaline Moynaki Lake, Crimea. Biol. Bull. 2021, 48, 934–937. [Google Scholar] [CrossRef]
- Kikuchi, S. The fine structure of the gill epithelium of a fresh-water flea, Daphnia magna (Crustacea: Phyllopoda) and changes associated with acclimation to various salinities. Cell Tissue Res. 1983, 229, 253–268. [Google Scholar] [CrossRef]
- Baillieul, M.; Wachter, B.D.; Blust, R. Effect of Salinity on the Swimming Velocity of the Water Flea Daphnia magna. Physiol. Zool. 1998, 71, 703–707. [Google Scholar] [CrossRef]
- He, Z. The effects of temperature on neonate production and intrinsic increasing rate (rm) of Moina macrocopa Straus. J. Dalian Fish. Univ. 1983, 5, 13–19. (In Chinese) [Google Scholar]
- Achuthankutty, C.T.; Shrivastava, Y.; Mahambre, G.G.; Goswami, S.C.; Madhupratap, M. Parthenogenetic reproduction of Diaphanosoma celebensis (Crustacea: Cladocera): Influence of salinity on feeding, survival, growth and neonate production. Mar. Biol. 2000, 137, 19–22. [Google Scholar] [CrossRef]
- Meester, L.D.; Maas, S.; Dierckens, K.; Dumont, H.J. Habitat selection and patchiness in Scapholeberis: Horizontal distribution and migration of S. mucronata in a small pond. J. Plankton Res. 1993, 15, 1129–1139. [Google Scholar] [CrossRef]
- Lemke, A.M.; Benke, A.C. Growth and reproduction of three cladoceran species from a small wetland in the south-eastern U.S.A. Freshw. Biol. 2003, 48, 589–603. [Google Scholar] [CrossRef]
- Kawabata, K.; Urabe, J. Length–weight relationships of eight freshwater planktonic crustacean species in Japan. Freshw. Biol. 2002, 39, 199–205. [Google Scholar] [CrossRef]
- Nebeker, A.V.; Schuytema, G.S. Chronic Effects of the Herbicide Diuron on Freshwater Cladocerans, Amphipods, Midges, Minnows, Worms, and Snails. Arch. Environ. Contam. Toxicol. 1998, 35, 441–446. [Google Scholar] [CrossRef]
- Gotelli, N.J. A Primer of Ecology; Sinauer Associates, Inc.: Sunderland, MA, USA, 1995. [Google Scholar]
- Murugan, N.; Sivaramakrishnan, K.G. Laboratory studies on the longevity, instar duration, growth, reproduction and embryonic development in Scapholeberis kingi Sars (1903) (Cladocera: Daphnidae). Hydrobiologia 1976, 50, 75–80. [Google Scholar] [CrossRef]
- Krebs, C.J. Ecology: The Experimental Analysis of Distribution and Abundance; Harper Collins: New York, NY, USA, 1994. [Google Scholar]
- Bottrell, H.H. Generation time, length of life, instar duration and frequency of moulting, and their relationship to temperature in eight species of cladocera from the River Thames, reading. Oecologia 1975, 19, 129–140. [Google Scholar] [CrossRef]
- Carmouze, J.P.; Durand, J.R.; Lěvêque, C.L. Lake Chad Ecology and Productivity of a Shallow Tropical Ecosystem; Kluwer Academic Publishers Group: Hague, The Netherlands; Boston, MA, USA; Lancaster, UK, 1983. [Google Scholar]
- He, Z.; Zhang, J.; Jiang, H. Effects of salinity of seawater on the survival and intrinsic rate of increase of two populations of Daphnia Magna. J. Dalian Fish. Univ. 1996, 11, 1–7. (In Chinese) [Google Scholar]
- He, Z.; Qin, J.; Wang, H. Studies on the saline and hypersaline zooplankton from JinNan and YinChuan regions. Acta Hydrobiol. Sin. 1989, 13, 24–38. (In Chinese) [Google Scholar]
- Alonso, M. Anostraca, Cladocera and Copepoda of Spanish saline lakes. Hydrobiologia 1990, 197, 221–231. [Google Scholar] [CrossRef]
- Williams, W.D. Life in Inland Waters; Blackwell Scientific Publications: Melbourne, Australia, 1983. [Google Scholar]
- Pennak, R.W. Freshwater Invertebrates of the United States, 3rd ed.; John Wiley & Sons: New York, NY, USA, 1989. [Google Scholar]
- Vernberg, W.B.; Vernberg, F.J. Environmental Physiological of Marine Animals; Springer: Berlin, Germany, 1972. [Google Scholar]
- Cole, L.C. The population consequences of life-history phenomena. Q. Rev. Biol. 1954, 29, 103–137. [Google Scholar] [CrossRef] [PubMed]
- Meats, A. The relative importance to population increase of fluctuations in mortality, fecundity and the time variables of the reproductive schedule. Oecologia 1971, 6, 223–237. [Google Scholar] [CrossRef]
- Snell, T.W. Fecundity, developmental time, and population growth rate. Oecologia 1978, 32, 119–125. [Google Scholar] [CrossRef]
- Lynch, M. The evolution of cladoceran life histories. Q. Rev. Biol. 1980, 55, 23–42. [Google Scholar] [CrossRef]
- Huang, X. Effect of temperature on the development, growth and egg production in Moina affinis (Cladocera, Moinidae). Acta Hydrobiol. Sin. 1983, 8, 105–112. (In Chinese) [Google Scholar]
- Yang, H.Y. The effect of salinity of salinity on assimilation, metabolism, growth and carbon budget of Daphnia magna. J. Fish. Sci. China 1997, 4, 33–38. (In Chinese) [Google Scholar]
- Qiu, Y.D. Effect of salinity on the energy budget of carp. J. Fish. China 1995, 19, 35–42. (In Chinese) [Google Scholar]
- Masclaux, H.; Bec, A.; Kainz, M.J.; Desvilettes, C.; Jouve, L.; Bourdier, G. Combined effects of food quality and temperature on somatic growth and reproduction of two freshwater cladocerans. Limnol. Oceanogr. 2009, 54, 1323–1332. [Google Scholar] [CrossRef]
- Yuslan, A.; Najuwa, S.; Hagiwara, A.; Ghaffar, M.A.; Suhaimi, H.; Rasdi, N.W. Production Performance of Moina macrocopa (Straus 1820) (Crustacea, Cladocera) Cultured in Different Salinities: The Effect on Growth, Survival, Reproduction, and Fatty Acid Composition of the Neonates. Diversity 2021, 13, 105. [Google Scholar] [CrossRef]
- Madhupratap, M.; Nehring, S.; Lenz, J. Resting eggs of zooplankton (Copepoda and Cladocera) from the Kiel Bay and adjacent waters (southwestern Baltic). Mar. Biol. 1996, 125, 77–87. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Salinity | AR | No. of Clutches | No. of Neonates | No. of Neonates | Mean Lifespan | Mean Interval | a Value |
|---|---|---|---|---|---|---|---|
| Treatments | (Days) | Per Female | Per Female | Per Clutch | (Days) | Between Clutches (Days) | |
| control | 4.28 a (0.76) | 7.14 a (2.16) | 52.00 a (22.46) | 8.37 a (1.77) | 14.30 a (8.19) | 1.75 a (0.18) | 1.3238 |
| 1 | 4.00 a (0.00) | 6.20 b (1.69) | 54.90 a (16.23) | 8.84 a (0.85) | 16.05 a (4.31) | 1.66 a (0.37) | 1.2809 |
| 2 | 4.00 a (0.00) | 7.67 a (1.94) | 67.33 a (17.78) | 8.79 a (0.90) | 17.30 a (6.72) | 1.76 a (0.35) | 1.3902 |
| 3 | 5.25 b (1.04) | 2.75 c (2.75) | 9.38 b (4.17) | 3.38 b (0.83) | 14.20 a (6.01) | 4.48 b (2.23) | 0.9209 |
| 4 | - | - | - | - | 14.55 a (5.31) | - | - |
| Salinity Treatments | rm | G (d) | R0 | λ |
|---|---|---|---|---|
| control | 0.777 | 4.56 | 34.60 | 2.175 |
| 1 | 1.021 | 3.94 | 55.80 | 2.776 |
| 2 | 0.903 | 4.55 | 60.60 | 2.467 |
| 3 | 0.481 | 3.95 | 6.70 | 1.617 |
| 4 | - | - | - | - |
| Development Phase | Duration of Each Phase | Figures | |||
|---|---|---|---|---|---|
| Control | 1 | 2 | 3 | ||
| One-cell stage | 2~4 min | 2~4 min | 2~4 min | 2~4 min | 1 |
| 0.5~1 min | 0.5~1 min | 0.5~1 min | 0.5~1 min | 2 | |
| Membrane lift | 2.1 ± 0.4 h | 2.1 ± 0.6 h | 1.9 ± 0.7 h | 2.4 ± 0.8 h | 3 |
| Two-cell stage | 1.8 ± 0.4 h | 1.9 ± 0.4 h | 1.6 ± 0.5 h | 1.9 ± 0.4 h | 4 |
| Four-cell stage | 2.4 ± 0.5 h | 2.3 ± 0.5 h | 2.3 ± 0.8 h | 2.8 ± 0.7 h | 5 |
| Eight-cell stage | 1.6 ± 0.8 h | 1.5 ± 0.6 h | 1.5 ± 0.5 h | 1.8 ± 0.6 h | 6 |
| Many-cell stage | 2.5 ± 1.0 h | 2.4 ± 0.9 h | 2.2 ± 1.0 h | 2.8 ± 1.0 h | 7 |
| Blastula stage | 2.0 ± 0.6 h | 1.9 ± 0.5 h | 1.8 ± 0.8 h | 2.2 ± 0.3 h | 8 |
| Gastrula stage | 1.7 ± 0.5 h | 1.6 ± 0.3 h | 1.5 ± 0.4 h | 1.6 ± 0.4 h | 9 |
| Formation of “T”-type embryo | 1.8 ± 1.1 h | 1.7 ± 1.0 h | 1.5 ± 0.9 h | 2.0 ± 0.8 h | 10 |
| Formation of antenna rudiment | 2.0 ± 0.8 h | 2.0 ± 0.8 h | 2.0 ± 0.6 h | 2.5 ± 0.8 h | 11 |
| Formation of pereiopod rudiment | 1.7 ± 0.3 h | 1.8 ± 0.4 h | 1.6 ± 0.5 h | 1.8 ± 0.5 h | 12 |
| Formation of 2 compound eyes | 3.6 ± 1.1 h | 3.4 ± 1.3 h | 3.4 ± 1.2 h | 4.0 ± 0.5 h | 13 |
| Single compound eye stage | 1.4 ± 0.3 h | 1.4 ± 0.4 h | 1.3 ± 0.4 h | 1.3 ± 0.5 h | 14 |
| Membrane rive | 0.8 ± 0.4 h | 0.8 ± 0.3 h | 0.6 ± 0.5 h | 0.9 ± 0.8 h | 15~16 |
| Expulsion from matrix | 2~4 min | 2~4 min | 2~4 min | 2~4 min | 16 |
| Cumulative duration of each phase | 25.4 h a | 24.8 h a | 23.2 h b | 28.0 h c | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Zhao, W.; Huo, Y.; Yin, X.; Wei, J.; Wang, S.; Wang, Y. Influence of Seawater Salinity on the Survival, Growth, Development and Neonate Production of Scapholeberis mucronata (O. F. Müller) (Crustacea: Cladocera). Water 2022, 14, 3706. https://doi.org/10.3390/w14223706
Wang L, Zhao W, Huo Y, Yin X, Wei J, Wang S, Wang Y. Influence of Seawater Salinity on the Survival, Growth, Development and Neonate Production of Scapholeberis mucronata (O. F. Müller) (Crustacea: Cladocera). Water. 2022; 14(22):3706. https://doi.org/10.3390/w14223706
Chicago/Turabian StyleWang, Lei, Wen Zhao, Yuanzi Huo, Xuwang Yin, Jie Wei, Shan Wang, and Yujie Wang. 2022. "Influence of Seawater Salinity on the Survival, Growth, Development and Neonate Production of Scapholeberis mucronata (O. F. Müller) (Crustacea: Cladocera)" Water 14, no. 22: 3706. https://doi.org/10.3390/w14223706